
Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans

National Institute of Ecology, Seo-Cheon Gun 33657, Republic of Korea
*
Author to whom correspondence should be addressed.
Water 2022, 14(22), 3781; https://doi.org/10.3390/w14223781
Submission received: 27 September 2022
/
Revised: 15 November 2022
/
Accepted: 17 November 2022
/
Published: 21 November 2022
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
Abstract
:Fish predation is restricted in areas with high aquatic macrophyte coverage; however, bluegill sunfish (Lepomis macrochirus), an exotic fish species, can consume prey even in such spaces. Here, we hypothesized that the cladoceran community structures in three wetlands (Upo, Jangcheok, and Beongae) with abundant macrophytes within the Nakdong River basin, South Korea, were influenced by high abundance and foraging activities of L. macrochirus. The study areas were dominated by L. macrochirus with body sizes < 10 cm, and their cladoceran consumption was higher than that of larger size classes. Most of the cladoceran species that were highly consumed by L. macrochirus were pelagic (i.e., Daphnia obtusa and Daphnia pulex); epiphytic species were consumed less (i.e., Chydorus shpaericus, Alona retangula, and Pleroxus laevis). We hypothesize that epiphytic cladoceran species are not easily explored by L. macrochirus. Selective consumption of pelagic species by L. macrochirus was reflected in the seasonal cladoceran distribution. Epiphytic species were the most abundant, whereas pelagic species abundance was relatively low. The low density of pelagic cladoceran species further increased the epiphytic species abundance. We conclude that the dominance of L. macrochirus introduced into Korean wetlands has been sufficient to change the cladoceran community structure; therefore, the abundance of epiphytes cladocerans in other countries or regions needs to be provided priority to the dominance and consumption pattern of L. macrochirus.
1. Introduction
In freshwater ecosystems, biological interactions such as competition and predation are important factors for understanding the community structure and species diversity [1,2]. Compared with terrestrial and marine ecosystems, freshwater ecosystems not only have a very limited range but also low continuity; hence, contact between individuals or communities is frequent, and relatively diverse organisms coexist in one space [3]. Therefore, biological interactions are closely related to the decrease or increase in species diversity or abundance within a specific region [4]. Predation often has a stronger effect than competition on the structure and species diversity of biological communities [5,6]. Although primary producer groups such as phytoplankton and aquatic macrophytes are mainly affected by physical and chemical disturbances (e.g., environmental change, pollution, and nutrients) rather than predation [7,8], primary consumer groups are largely affected by predators. Empirical studies have suggested that the foraging activity of fish, which play apex predator roles within freshwater food webs, has a strong influence on the temporal and spatial distributions of invertebrates such as zooplankton, Diptera, Ephemeroptera, and Odonata [9,10,11]. Although some adult fish consume other fish (i.e., juvenile fish) as food sources, including piscivorous fishes, small invertebrates are their main food source [12]. Therefore, fish predation must be considered a major factor in understanding the community structure or distribution of primary consumers such as invertebrates.
In freshwater ecosystems, cladoceran communities with small body sizes and relatively slow movements are frequently used as food sources for various fish species [13]. Moreover, they have relatively rapid population growth and are often present at a high density [14,15]; therefore, they are more strongly affected by fish predation (especially young fish) than other invertebrates. This food consumption pattern often has a negative effect on the cladoceran species diversity or density, and the strong foraging of fish in some areas sometimes leads to the extinction of the cladoceran community [16,17]. This prey–predator relationship has long been maintained and is sufficient to lead to various defensive responses in prey groups (i.e., cladocerans). Previous studies have suggested that increasing head and spine length, which is frequently seen in some cladoceran species (e.g., Daphnia spp.), are primary morphological changes to prevent fish predation [18,19]; however, their effectiveness in avoiding fish predation is relatively low. In contrast, the behavioral change (i.e., diel vertical migration) of cladoceran species that migrate between the upper and bottom layers in deep lakes during the day and night is effective in sustaining their survival and density as individuals inhabit the bottom layer during the daytime when fish predation is high [20]. However, the bottom layers of wetlands or reservoirs with relatively low water levels are not avoided by fish, making it difficult for cladocerans to use these areas as a refuge. Therefore, cladocerans inhabiting wetlands or shallow reservoirs require different defense strategies to avoid fish predation compared with those in deep lakes.
Previous studies have suggested that aquatic macrophytes are frequently used as refuges for cladocerans to avoid fish predation [21,22,23]. Wetlands or shallow reservoirs with shallow water levels and abundant nutrients are suitable for aquatic macrophyte growth, which strongly mediate biological interactions such as predation and competition. Choi et al. [22] found that spaces covered by diverse aquatic macrophyte species with different forms serve as more suitable habitats for cladocerans than those covered by one or two aquatic macrophyte species. The aquatic habitat structure is determined by the arrangement and shape of aquatic macrophyte leaves and stems [24], and the dominance of aquatic macrophytes in complex forms creates more complex structures in specific areas. In particular, areas that were highly covered by submerged macrophytes had a more complex habitat structure than those covered by other macrophyte forms (emergent, free-floating, and floating-leaved). Space that is highly covered by aquatic macrophytes is disturbed by the visual food exploration of fish; therefore, such areas can be supported by a high cladoceran abundance [25]. Although fish usually prefer areas covered by aquatic macrophytes or open waters, they remain on the edge of the vegetation bed to acquire a stable food source [26,27]. Aquatic macrophytes are used as substrate surfaces for some cladoceran species. For example, cladoceran species belonging to the family Chydoridae are closely related to aquatic macrophytes because they utilize aquatic macrophyte leaf and stem surfaces as attachment substrates and for predator avoidance (i.e., Chydorus shpaericus and Alona affinis; [28,29]). Although these species are frequently observed in wetlands and shallow reservoirs where aquatic macrophytes are abundant, they are extremely rare in deep lakes where aquatic plants are absent or have low coverage [30]. This suggests that wetlands or shallow reservoirs where aquatic macrophytes are abundant may be inhabited by more diverse cladoceran species than other freshwater ecosystem types.
In this study, we focused on the cladoceran communities in wetlands and shallow reservoirs in South Korea, which are mainly inhabited by cladoceran species belonging to the family Chydoridae. In particular, the Nakdong River basin in South Korea is scattered with wetlands of various sizes and is supported by high relative abundance of cladoceran species such as Alona guttata, C. sphaericus, and Pleurous aduncus. Empirical studies have suggested that it is difficult for Chydoridae to gain regional dominance due to their unique environmental preferences (i.e., preference for aquatic macrophytes and high water temperature [31,32]) and relatively low population growth rates [33]. From this perspective, the cladoceran community structure dominated by species belonging to the Chydoridae family in wetlands or shallow reservoirs in South Korea is interpreted as a result of being influenced by specific environmental variables, and the type and foraging activities of predators distributed in each region were hypothesized to be strong influencing factors. Introduced into Korea in the 1970s, the bluegill sunfish (Lepomis macrochirus) is frequently found throughout South Korea, and due to its strong disruption effect of indiscriminate consumption of invertebrates such as zooplankton and Odonata larvae, South Korea manages them as an “ecosystem disturbances species” [27,34]. Lepomis macrochirus is mainly distributed in the littoral vegetated area to avoid competition and predation by bass (Micropterus salmoides), which is also an ecosystem disturbance species, [35] and has a strong preference for consuming abundant pelagic cladocerans (Daphnia obtusa, Ceriodaphnia dubia, and Simochephalus vetulus) in this region [27]. Furthermore, some studies in Lake Biwa in Japan have revealed that L. macrochirus exhibit clear spatial changes in growth, maturation, and reproduction [36,37]. This is because the topographical characteristics of this area cause a wide range of water temperatures. In areas with low water temperatures during the pre-spawning season, ovary maturation in L. macrochirus is delayed, while high temperatures help them mature quickly [37].
We predicted that the selective consumption of pelagic cladocerans by L. macrochirus would provide an opportunity for other cladoceran species to thrive. The selective consumption of L. macrochirus serves as an advantage for epiphytic cladoceran species (i.e., A. guttata, A. retangula, C. sphaericus, and P. aduncus), which actively utilize areas with high aquatic macrophyte coverage in wetlands or shallow reservoirs as their habitat. This study aimed to elucidate: (1) the response of cladoceran community distributions to environmental variations, (2) the effect of changes in the cladoceran distribution on different microhabitat characteristics in relation to aquatic macrophytes, and (3) the influence of the cladoceran community structure on fish predation. To test our hypothesis, we investigated the effects of the L. macrochirus distribution and food consumption patterns on the relative abundance and seasonality of cladocerans in three wetlands in South Korea with a high abundance of L. macrochirus, and the effect of the introduction of exotic species on regional biodiversity was examined.
2. Materials and Methods
2.1. Study Site
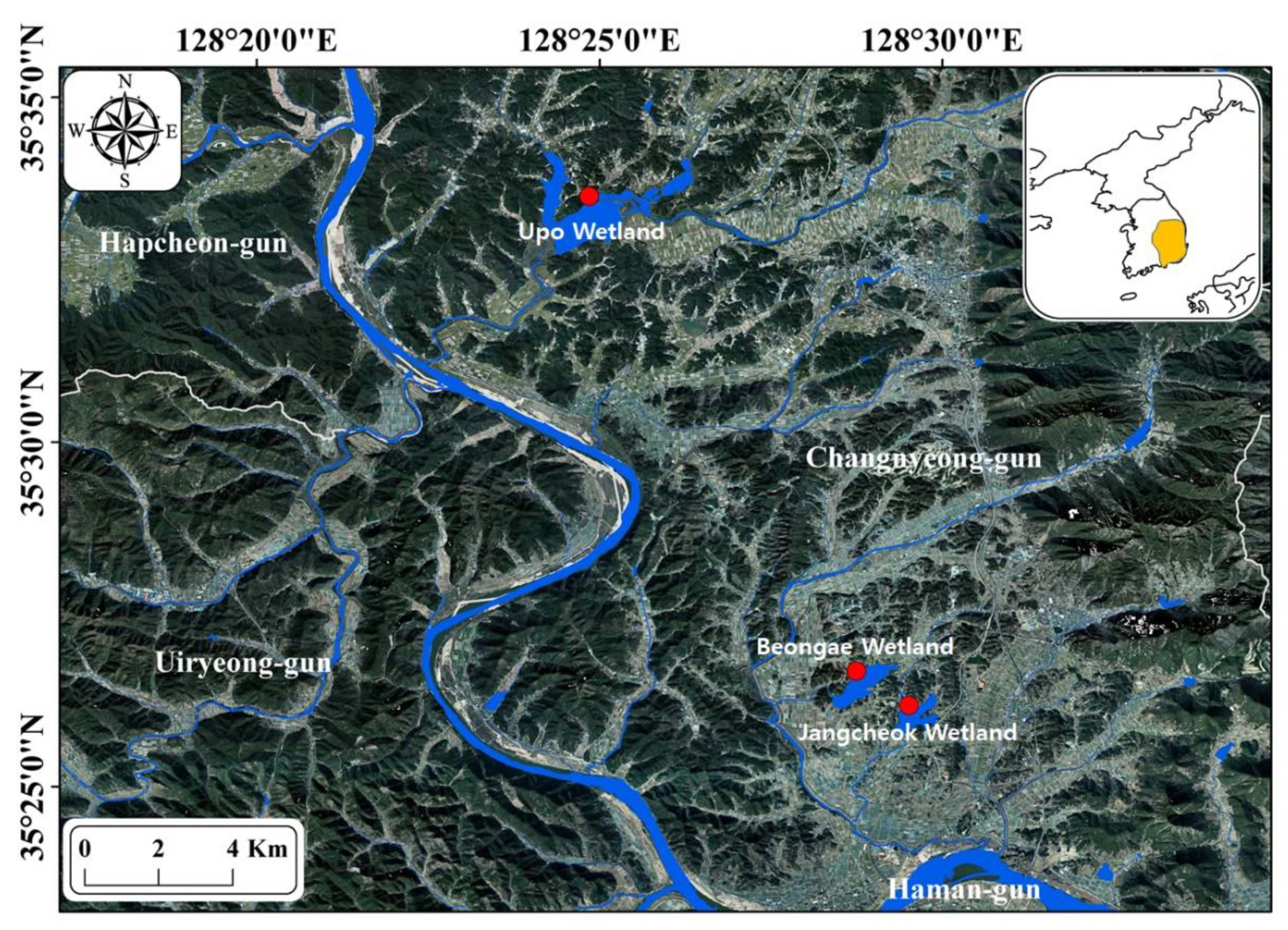
The freshwater network in South Korea occupies most of the area of the country and is divided into four large rivers (Nakdong, Geum, Han, and Yeongsan rivers). The Nakdong River, located in the southeastern part of South Korea, is the longest among the four large rivers (506.17 km) and has the largest catchment area (23,384.21 km2). The Nakdong River basin is frequently flooded by concentrated summer rainfall, and the surrounding area is composed of soil with low drainage capacity, which is advantageous for frequent flooding and wetland or reservoir formation. In particular, numerous wetlands are scattered in the confluence section of the Nakdong and Nam rivers, including international wetlands such as Upo Wetland and Junam Reservoir (approximately 200 places [22]). Nutritional salts such as phosphorus and nitrogen flow into these wetlands from the surrounding agricultural land and residential areas, which provides favorable environmental conditions for the growth of various aquatic macrophytes.
South Korea has a temperate climate zone and four distinct seasons; however, due to recent climate change, the southern part of South Korea has subtropical climate characteristics. Invasion and settlement of various subtropical organisms have been confirmed, and there have been simultaneous decreases in native species [38]. Rainfall concentrated in summer is a typical characteristic of the Nakdong River basin and acts as a major disturbance factor to reduce the abundance of biological communities grown in spring [39,40].
To understand the influence of aquatic macrophytes on L. macrochirus foraging activity, we selected three wetlands (Upo, Jangcheok, and Beongae wetlands) covered by similar plant species and with shallow water depths based on previous studies (Figure 1; [22]). The sizes of these wetlands are 1.84, 1.23, and 1.41 ha, respectively, and they are used as an agricultural water supply. The littoral zones are shallow (0.5–0.9 m depth) and the central areas are deeper (1.4–2.7 m depth). The study sites were almost completely covered with macrophytes from spring (May) to autumn (October). In this study, we identified seven aquatic macrophyte species: Phragmites australis, Paspalum distichum, Zizania latifolia, Salvinia natans, Trapa japonica, Ceratophyllum demersum, and Hydrilla verticillate.
2.2. Monitoring Strategy
We conducted biweekly monitoring at the study sites over a 3-year period from 2015 to 2017. Prior to monitoring, we searched for candidate sampling locations within the wetlands that were characterized by similar macrophyte species compositions and depths (average depths 0.6–0.7 m). At the study location, three 1 × 1 m quadrats were used to monitor environmental variables, macrophyte biomass, and cladoceran communities.
Environmental variables (water temperature, percent saturation of dissolved oxygen (DO), conductivity, pH, turbidity, and chlorophyll a (Chl a)) were measured, and cladocerans were enumerated using water samples collected from the quadrats. A DO meter (YSI Model 58; Yellow Springs, OH, USA) was used to measure the water temperature and DO, and conductivity and pH were measured using a conductivity meter (YSI model 152; Yellow Springs, OH, USA) and pH meter (Orion Model 250 A; Orion Research, Beverly, MA, USA), respectively. Water samples were transported within 4 h to the laboratory to measure turbidity and Chl a. A turbidity meter (Model 100 B; HF Scientific Inc., Ft. Myers, FL, USA) was used to measure turbidity. To determine the Chl a concentration, water samples were filtered through 0.45 µm mixed cellulose ester membrane filters (A045A047A; Advantech Co. Ltd., Taipei, Taiwan). The filtered membranes were placed in cold 90% acetone in darkness at 20 °C for 4 h. To improve extraction, the cells were disintegrated for 2 min in an ultrasonic bath. To remove cell debris and filter particles, the pigment extract was centrifuged at 5000 rpm for 5–10 min. The extinction coefficient was estimated at 600 and 750 nm using a spectrophotometer (Japan Fantec Research Institute, Shizuoka, Japan), with the sample placed in a 1 cm glass cuvette [41]. The Chl a concentration was estimated using the following equation:
where Va is the extract volume (mL) and Vb is the sample volume (mL).
Chl a = 11,403 × (A600 − A750) × Va × Vb−1,
For cladoceran enumeration, we collected 10 L water samples from each quadrat using a 10 L column water sampler (length: 20 cm; width: 30 cm; height: 70 cm). The sampler was placed vertically in the water to collect rotifers from the entire water column of the quadrat. The sampled water was filtered through a plankton net (32 mm mesh) and the filtrate was preserved in sugar formalin (final concentration: 4% formaldehyde [42]). Cladoceran enumeration and identification at the species level were performed using a microscope (ZEISS, Model Axioskop 40; ×200 magnification), with identification based on the classification key published by Mizuno and Takahachi [43].
After cladoceran collection, we investigated the aquatic macrophyte biomass in each quadrat. We removed emergent organs above the water surface (i.e., stalks and flowers) of all aquatic macrophytes because cladocerans inhabit underwater environments. In emergent and floating-leaved macrophytes, only a small part of the plant body is submerged; whereas, in free-floating and submerged macrophytes, complex parts of the plant body are submerged. The collected macrophytes were dried in the laboratory at 60 °C for 2 days, and the dry plant weight was estimated for each quadrat.
To understand the effect of fish predation on cladoceran community structures, we investigated the seasonal distribution of fish communities using cast nets (7 × 7 mm) and scoop nets (5 × 5 mm) along 300 m transects in each wetland in 2017. The cast and scoop nets were deployed for 30 min and 15 min, respectively. Each of the collected fish was identified to the species level according to Kim and Park [44] and the classification system of Nelson [45]. The fish assemblage collected from the study site was dominated by L. macrochirus (approximately > 70%), with low density and frequency of other fish species (Micropterus salmoides, Pseudorasbora parva, Carassius Carassius, Misgurnus anguillicaudatus, and Odontobutis platycephala). Thus, we only used information on L. macrochirus for the analysis in this study. To understand the seasonal distribution and food preferences of L. macrochirus collected from each wetland, we conducted seasonal body size measurements and gut content analyses. The L. macrochirus body size was measured from head to tail using a 50 cm tape measure. Based on the body size, L. macrochirus was divided into three size classes (>5 cm, 6–10 cm, and 11–15 cm). The guts of each of the 20 individuals in each of the three size classes were immediately fixed in a methanol-formaldehyde solution. We identified and counted all the cladoceran species in the L. macrochirus gut contents. Prey item abundance was calculated based on the number of each item in the gut relative to the total number of items.
Furthermore, we investigated cladoceran and fish distributions in five different microhabitat types based on the heterogeneity of the macrophytic composition in each wetland: (1) open water area (without macrophytes) (Ow), (2) area covered by P. distichum (Pd), (3) area covered by T. japonica (Tj), (4) area covered by floating-leaved macrophytes (Hydrocharis dubia and S. natans) (floating), and (5) area covered by floating-leaved and submerged macrophytes (H. dubia, S. natans, C. demersum, H. verticillata, and Najas graminea) (mixed). Fifty randomly selected sampling points were surveyed at each site from September–October 2017. Quadrats (1 × 1 m) were established at each sampling point for monitoring. Ten sampling points were assigned for each microhabitat type.
2.3. Data Analysis
We used non-metric multidimensional scaling (NMDS) to examine the seasonal distribution patterns of cladocerans according to environmental variation in the three wetlands. NMDS ordination plots were generated based on the Euclidean distance and the goodness-of-fit was assessed in terms of the loss of stress. Each variation was log-transformed after being assessed for normality using the Shapiro–Wilk test. The stress value for the two-dimensional solution was 0.162, which is lower than the generally accepted maximum stress value of <0.2 [46]. The significance of the fitted vectors was assessed using 3000 permutations, with p < 0.05 considered significant. The NMDS ordination was conducted using the R package “vegan” (version 2.5-3 [47]).
Furthermore, a one-way analysis of variance (ANOVA) was used to examine the effects of the microhabitat type on the mean densities of L. macrochirus and cladocerans. Tukey’s test was used for additional post-hoc comparison analysis to determine statistically significant differences. Statistical analyses, including ANOVA, were conducted using SPSS ver. 20 (released 2011; IBM SPSS Statistics for Windows, version 20.0. Armonk, NY, USA: IBM Corp.). Differences and relationships were considered statistically significant at p < 0.05.
3. Results
3.1. Environmental Variables and Cladoceran Distributions
Seven environmental variables measured in Upo, Jangcheok, and Beongae wetlands reflected the typical seasonal characteristics of temperate climates (Table 1). The dry weight of macrophytes and water temperature were low in spring and winter and high in summer and autumn; whereas DO, conductivity, pH, turbidity, and Chl a showed contrasting seasonal patterns (i.e., high in spring and winter and low in summer and autumn). The seasonality of these environmental variables was supported by a similar pattern over the 3 years of the study, and there was minimal difference between the three wetlands.
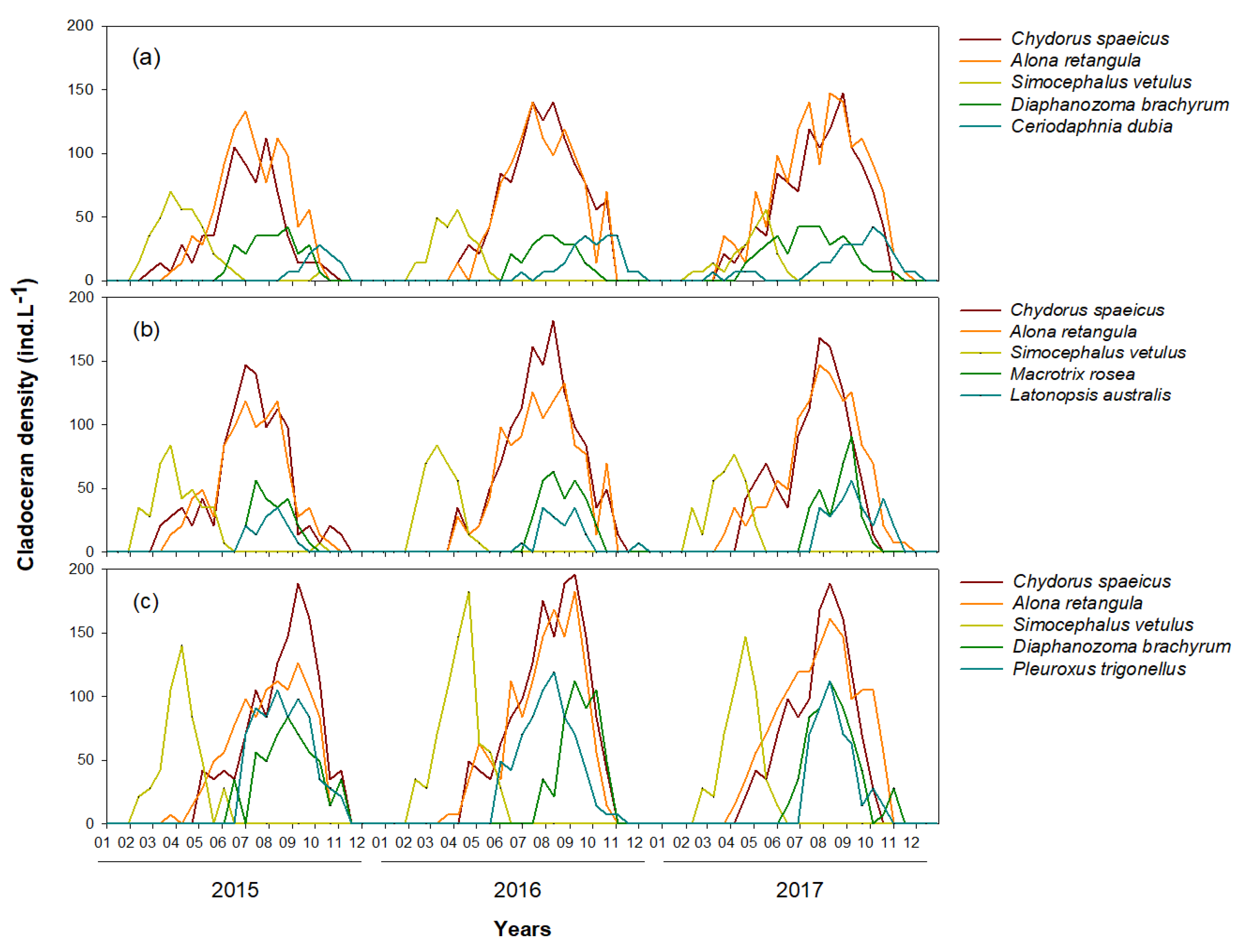
The number of cladoceran species was similar among the three wetlands (8–10 species); whereas, the dominant species differed between the wetlands (Figure 2). Three cladoceran species (i.e., C. shpaericus, Alona retangula, and S. vetulus) were abundant in the three wetlands; however, the abundance of the other cladoceran species varied depending on the wetland. Diaphanozoma brachyrum was the fourth most common cladoceran species in Upo and Beongae wetlands; whereas, its density was relatively low in Jangcheok Wetland. The fifth most dominant cladoceran species was clearly different among the three wetlands (Upo, C. dubia; Jangcheok, Latonopsis australis; Beongae, Pleuroxus trignellus). The five dominant cladoceran species in each wetland accounted for >90% of the total cladoceran abundance. The dominant cladoceran species exhibited distinct seasonality. Chydorus shpaericus and A. retangula were abundant in summer and autumn (August–September); whereas, S. vetulus was the most abundant only in spring (March–April). D. brachyrum was observed for a relatively long period from early summer to late autumn, and C. dubia was abundant in autumn. Macrotrix rosea, L. australis, and P. trignellus were also observed mainly in summer and autumn.
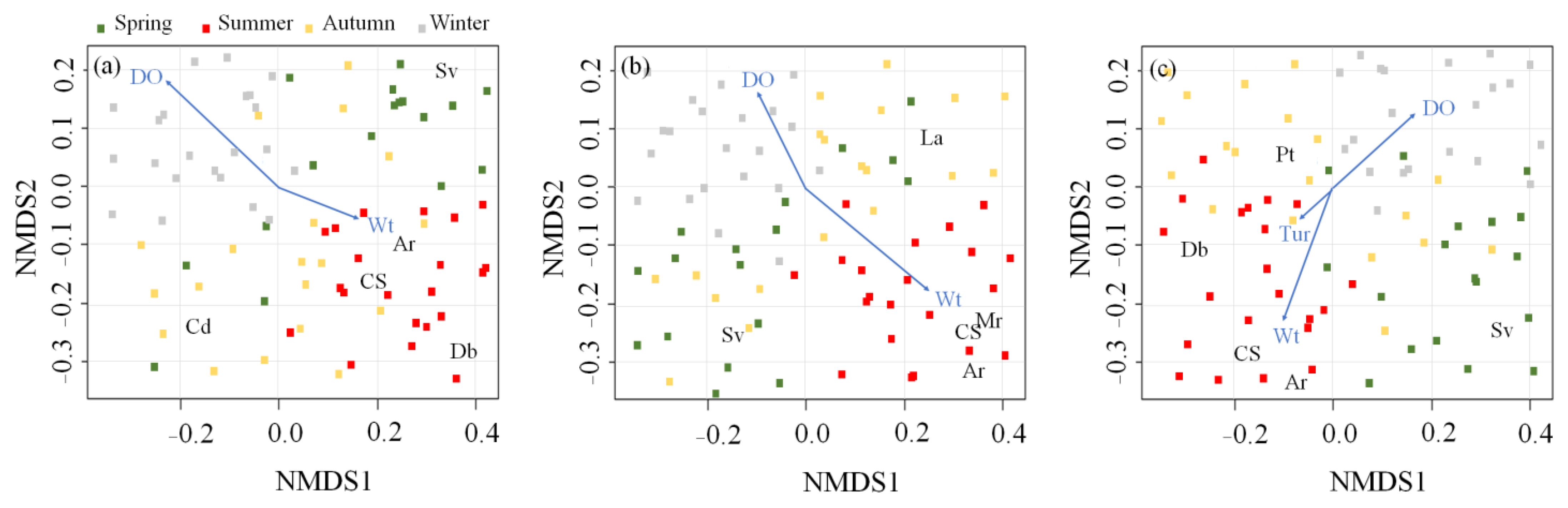
We fitted the dominant cladoceran species that appeared in each wetland to the NMDS ordination axes and selected two or three environmental variables that were significantly correlated with those axes (p < 0.05; Figure 3). Although the cladoceran species differed in each wetland, the relationship between cladocerans and environmental variables was similar. Chydorus shpaericus (Cs), A. retangula (Ar), D. brachyrum (Db), and M. rosea (Mr) were associated with higher water temperatures and lower DO. Whereas distributions of S. vetulus (Sv), C. dubia (Cd), L. australis (La), and Pleuroxus trigonellus (Pt) were relatively irregular.
3.2. Effect of Microhabitat Types on Cladoceran Distribution
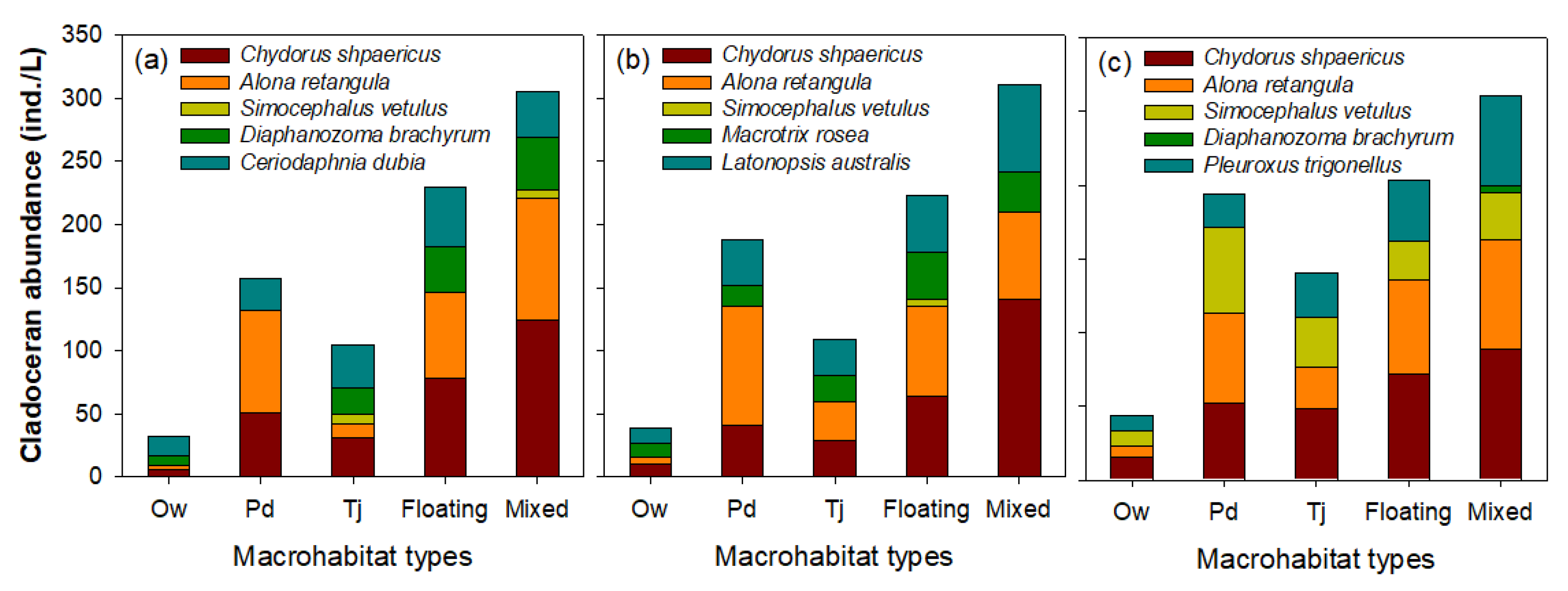
The abundance of the five dominant cladoceran species in each wetland differed according to the microhabitat type based on the spatial coverage rate of aquatic macrophytes (Figure 4). The highest cladoceran abundance was found in the ‘mixed’ type, followed by the ‘floating’ and ‘Pd’ types. In contrast, the ‘Ow’ type had the lowest abundance of cladocerans. This cladoceran distribution pattern was similar in all wetlands. Three microhabitat types (Pd, floating, and mixed) where high cladoceran abundance was observed were predominantly dominated by high abundance of C. sphaericus and A. retangula. S. vetulus was abundant in all microhabitat types related to macrophytes in the Beongae Wetland; whereas its abundance was low in the Upo and Jangcheok Wetlands. The other dominant cladoceran species showed irregular distribution characteristics depending on the wetland or habitat type.
3.3. Effect of Microhabitat Characteristics on L. macrochirus Distribution
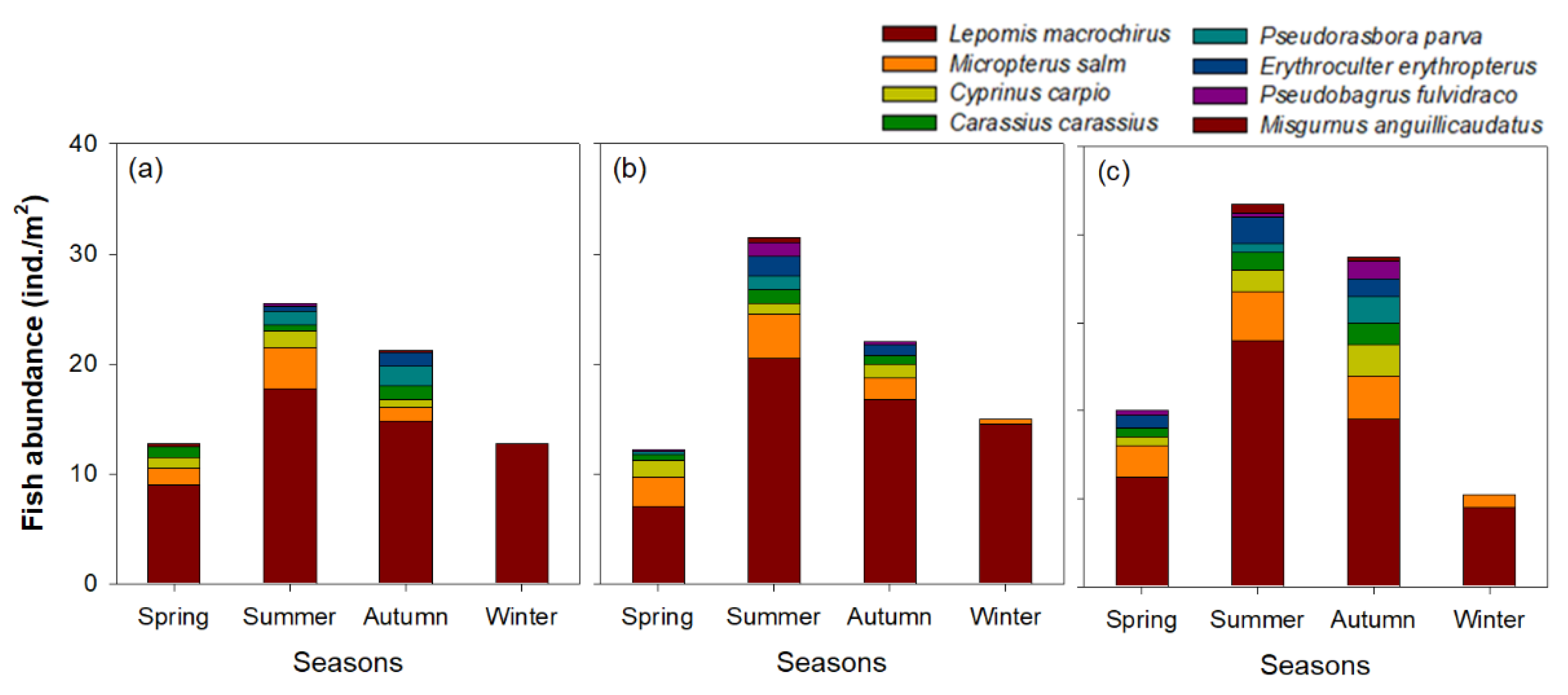
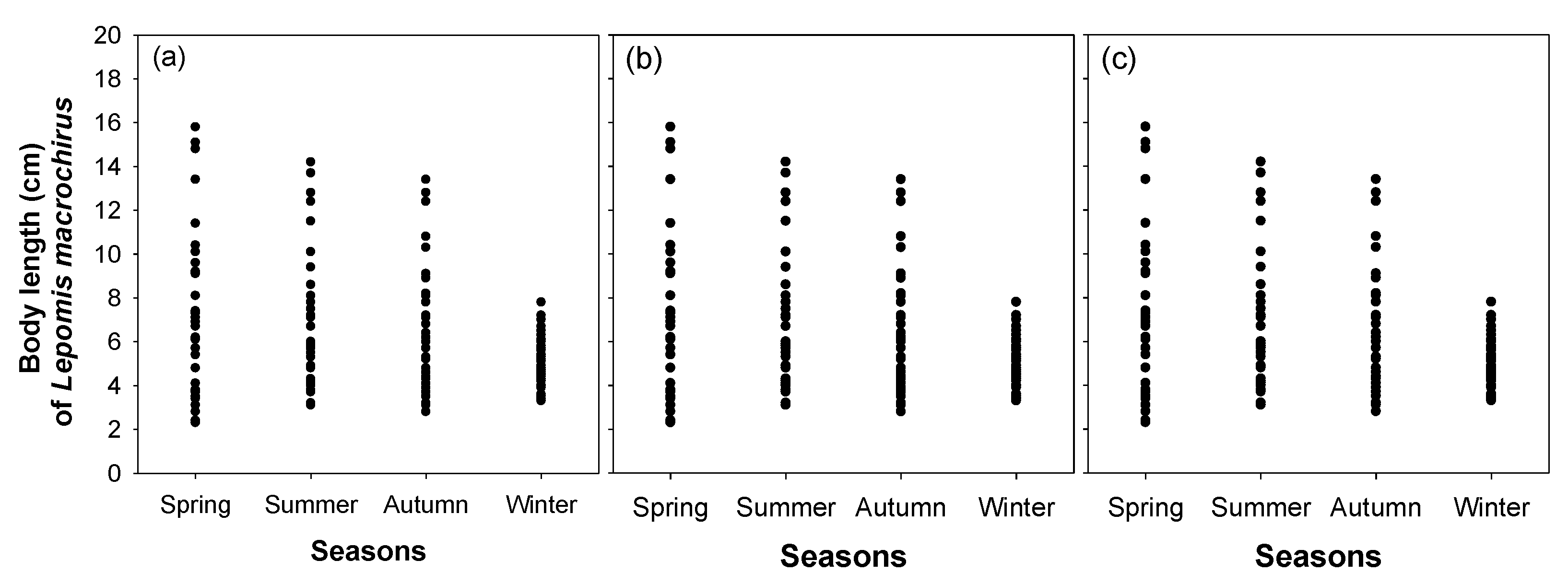
The seasonal distribution patterns of the fish communities were similar among the three wetlands (Figure 5). A total of eight fish species were commonly observed in all wetlands, of which the relative abundance of L. macrochirus was >70% (approximately 61.4~75.1%). The species number and abundance of fish communities were highest in summer and relatively low in spring and winter. The relative abundance of L. macrochirus was highest in winter and relatively low from spring–autumn due to the increase in the abundance of other fish species. In particular, in summer, L. macrochirus abundance increased as other fish species (including the sub-dominant species M. salmoides) became abundant; whereas, the relative abundance of M. salmoides decreased. L. macrochirus dominating the three wetlands mostly had body sizes < 10 cm (Figure 6). L. macrochirus individuals > 10 cm were observed from spring–autumn but were less abundant than individuals < 10 cm. The body size range of L. macrochirus was the widest in spring (2.3–16.2 cm) and gradually decreased from spring–winter. In winter, only L. macrochirus with body sizes < 10 cm were abundant.
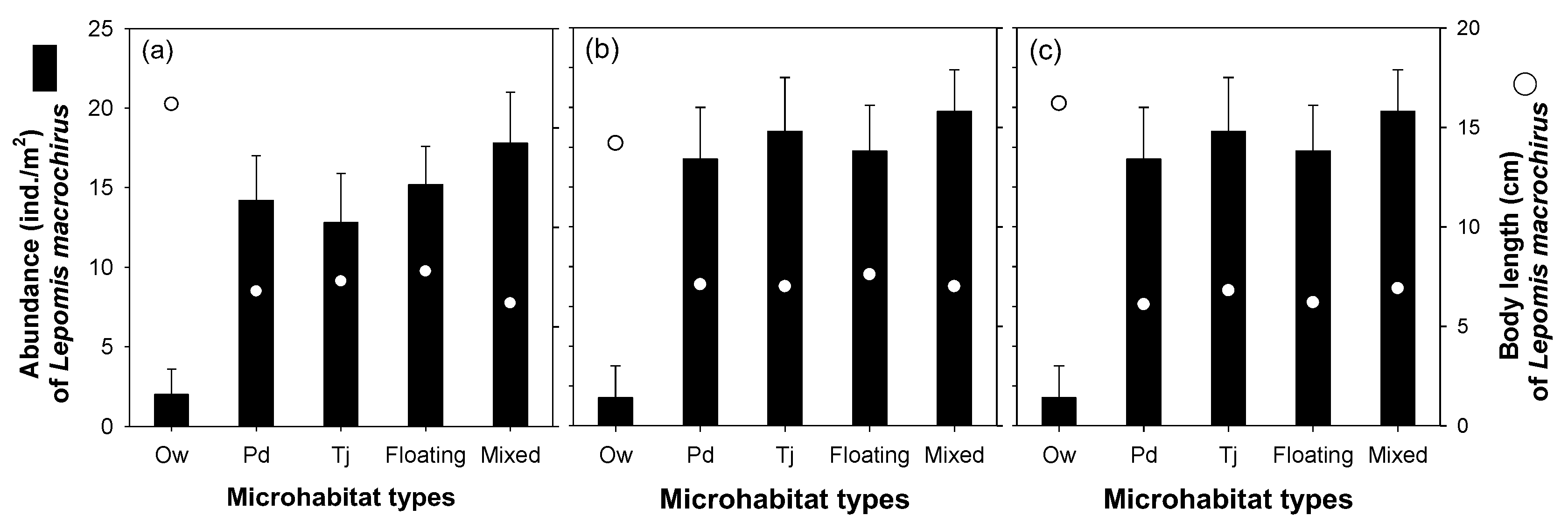
L. macrochirus abundance had minimal influence on the five microhabitats, based on the aquatic macrophyte spatial coverage rate (Figure 7). Although the L. macrochirus abundance was extremely low in the ‘Ow’ type, there were few statistical differences between the four microhabitat types (Pd, Tj, floating, and mixed) other than the ‘Ow’ type (one-way ANOVA, p > 0.05). In addition, there was no significant difference in the L. macrochirus body sizes between the habitat types (one-way ANOVA, p > 0.05). The body sizes and spatial distribution patterns of L. macrochirus in the ‘Ow’ microhabitat type were common among all three wetlands.
3.4. Utilization of Cladocerans as Food Source for L. macrochirus
The consumption patterns of cladoceran species collected from each wetland differed according to the season (Figure 8).
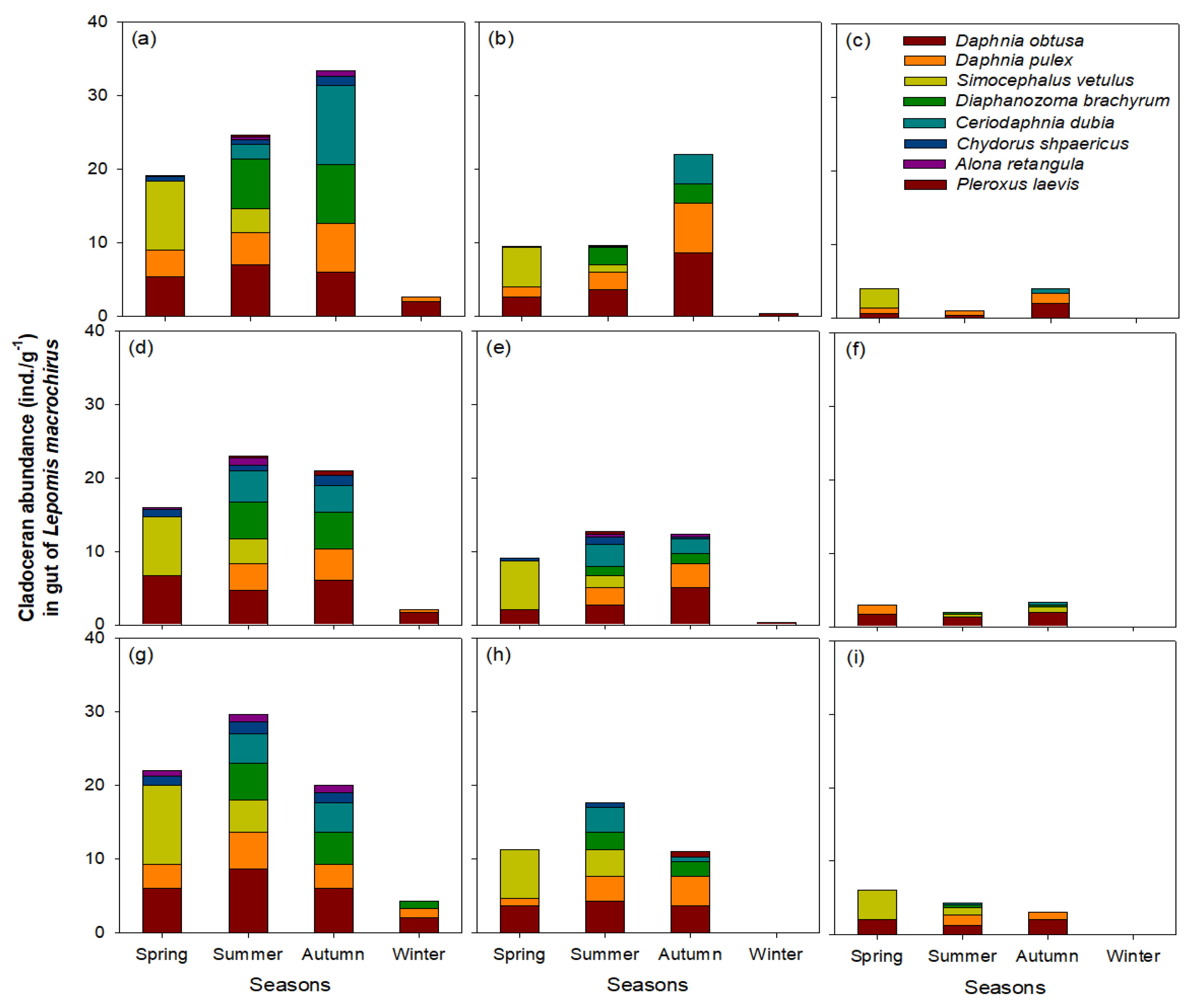
D. obtusa and D. pulex were commonly used as the main food sources for L. macrochirus in most seasons, while the consumption of the other cladoceran species by L. macrochirus depended on the season. S. vetulus was consumed largely by L. macrochirus in spring; whereas, D. brachyrum and C. dubia were frequent food sources for L. macrochirus in summer and autumn. However, the consumption of C. sphaericus, A. retangula, and P. laevis by L. macrochirus was low in all three wetlands. The seasonal consumption patterns of these cladoceran species by L. macrochirus were similar in accordance with all factors such as wetlands, seasons, and L. macrochirus body sizes.
The consumption of cladoceran species by L. macrochirus showed a clear quantitative difference according to the L. macrochirus body size (one-way ANOVA, p < 0.01). The highest cladoceran consumption by L. macrochirus was found for L. macrochirus with a body size < 5 cm, followed by individuals with a body size of 6–10 cm. Cladoceran consumption by L. macrochirus with body sizes of 11–15 cm was relatively low.
4. Discussion
4.1. Distribution Patterns of Cladocerans and Fish Communities
In this study, high densities of cladoceran communities were observed mainly in summer and autumn. Summer and autumn not only have the highest water temperatures among the four seasons but are also abundant in food sources such as phytoplankton and bacteria, making these seasons suitable for the growth of various aquatic fauna, including cladocerans. However, cladoceran communities, which play a role as primary consumers in freshwater food webs, can often be supported by low densities because high density of their predator (e.g., invertebrates and fish) as well as cladocerans occur during summer and autumn [48,49,50]. Cladocerans are preferred food sources for predators such as invertebrates and fish because of their low swimming speed and rapid population growth rate [51]. These biological factors, as well as environmental factors, such as concentrated summer rainfall, are major factors that decrease the density of cladoceran communities in summer and autumn [40,52]. The turbulence that occurs due to heavy rainfall and rapid water flow can reduce the abundance of cladoceran species that do not have excellent swimming capabilities. An empirical study suggested that summer rainfall causes a major disruption to the spring growth of zooplankton communities in rivers and streams and induces new succession in autumn [53]. Nevertheless, frequent observations of high cladoceran densities in summer and autumn in various wetlands and shallow reservoirs, including in this study, are closely related to aquatic macrophytes, which excessively cover surfaces during these seasons. The space covered by macrophytes has complex microhabitat structures and heterogeneity, with biological and non-biological effects [54,55,56]. Fish foraging activity has been proven to be significantly reduced in spaces with excessive aquatic macrophyte coverage [57], allowing for consistent population growth of prey groups such as cladocerans. In addition, surface coverage caused by macrophytes mediates physical disturbances (such as summer rainfall) and prevents small aquatic organisms (such as zooplankton) from being scattered or damaged [40,53]. Choi et al. [58] found that strong summer rainfall consecutively increased epiphytic rotifer abundance on aquatic macrophyte leaves and stems in Upo Wetland in South Korea. This is interpreted as a strategy to prevent physical disturbances caused by heavy rainfall. In this study, unlike pelagic species such as D. obtusa and D. pulex, the epiphytic species C. sphaericus, A. retangula, and P. laevis, which predominated in this study area, have characteristics that enable them to crawl on the leaves and stem surfaces of aquatic plants [59,60,61]; hence, they are easily distributed in areas with highly aquatic macrophyte coverage. In particular, because submerged macrophytes have higher heterogeneity of leaves and stems than other plant types (e.g., emergent, free-floating, and floating-leaved macrophytes), their high coverage can greatly contribute to the suitability of microhabitats for epiphytic species (i.e., predation avoidance and suitability as attachment substrates) [62,63]. We also found high cladoceran density in the “mixed” habitat which was more highly covered by submerged macrophytes than the other microhabitat types. The “mixed” habitat type has a physically complex microhabitat structure and a strong effect of restricting the foraging activities of fish that visually search for food.
Although previous studies have suggested that fish foraging activity is strongly restricted by aquatic macrophytes [64,65], the dominant fish species in this study, L. macrochirus, was abundant in spaces with high aquatic macrophyte coverage. The spatial distribution of L. macrochirus is considered a behavioral strategy adapted from interactions between fish communities, such as competition and predation. L. macrochirus and M. salmoides have similar food preferences; however, M. salmoides has a higher piscivorous tendency than L. macrochirus [66]. Young L. macrochirus and M. salmoides utilize similar food sources (i.e., zooplankton or invertebrates); however, these two species have different consumption patterns during the adult life stage. L. macrochirus adults continue to rely on zooplankton or invertebrates; whereas, M. salmoides adults usually consume young fish, such as Pseudorasbora parva and L. macrochirus [67]. M. salmoides has a preference for consuming native fish over L. macrochirus; however, the current continued decline in native fish population in Korean wetlands has accelerated L. macrochirus consumption. L. macrochirus, which is relatively vulnerable to predation by M. salmoides, is thought to be mainly distributed in areas that are covered by aquatic macrophytes to facilitate avoidance of M. salmoides. The majority of L. macrochirus inhabiting littoral vegetation in this study were < 10 cm in body size, which is similar to the findings of other studies. M. salmoides are mainly distributed in areas with minimal aquatic macrophytes coverage or in open waters due to the difficulty in searching for food in areas that are covered by aquatic macrophytes [27].
However, the spatial distribution of L. macrochirus was not affected by the five microhabitat types based on aquatic macrophytes. L. macrochirus was distributed at a similar abundance in the “mixed” habitat type, which has the most complex structure, as in the other microhabitat types. However, from these spatial distributions of L. macrochirus, it is difficult to confirm that L. macrochirus foraging activities were not affected by the microhabitat type. Because each microhabitat type was connected, rather than being an independent space, the possibility that L. macrochirus collected from one microhabitat type performed foraging activities in another cannot be ruled out. Moreover, pelagic cladoceran species, D. obtusa and D. pulex, which are mainly consumed by L. macrochirus, were distributed at already low abundance in all microhabitat types, making it difficult to elucidate the impact of foraging activity of L. macrochirus on pelagic cladoceran species. Because the density of M. salmoides, a predator of L. macrochirus, was low in all microhabitats that were covered by aquatic macrophytes, L. macrochirus individuals do not have to select a refuge space based on aquatic macrophytes. If L. macrochirus have sufficient feeding activity in other microhabitat types and are restricted from foraging activity in “mixed” habitats, they are only able to reside in the “mixed” habitat for a short time. Since the proportion of the area covered by ‘mixed’ habitat within the entire area covered by aquatic macrophytes was relatively low, this was not sufficient to lead to different spatial distributions of L. macrochirus. This can be interpreted as a result of the larger space covered by aquatic macrophytes; regardless of the physical complexity of the habitat, L. macrochirus population growth can be promoted.
4.2. Influence of L. macrochirus on Cladoceran Community Structure
In this study, we identified the food preferences and consumption patterns of L. macrochirus in wetlands that were abundant in aquatic macrophytes through gut content analyses. High consumption of cladocerans by L. macrochirus was found mainly for L. macrochirus individuals < 5 cm in body size, and cladoceran consumption by L. macrochirus with body sizes > 5 cm was relatively low. In general, the size of food items consumed by fish also increases as their body size increases, which ensures the cost-effectiveness of foraging activities [68]. L. macrochirus individuals < 5 cm consume cladocerans and copepods; however, as they grow they develop a tendency to consume large food sources, such as invertebrates and young fish [69]. However, when relatively large food items (such as invertebrates) are scarce, L. macrochirus consume small food items such as zooplankton [70]. The dietary shift of fish depends on the plasticity of available resources in the habitat [71]. Considering that cladoceran species are relatively small among the available food resources for fish in freshwater ecosystems, if L. macrochirus with a body size > 5 cm consume cladocerans it means that large food items (e.g., invertebrates) are lacking or that there is a high preference for cladocerans. However, the latter (high preference for cladocerans) is more likely than the former, given the suggestion that the study sites have high invertebrate species diversity and abundance [72]. From this finding, the continuous consumption of pelagic cladocerans by L. macrochirus has greatly contributed to the cladoceran species composition and community structure in the wetlands in the present study.
L. macrochirus prefers pelagic cladocerans as a food source over epiphytic cladocerans. We frequently observed pelagic cladoceran species (e.g., D. obtusa, D. pulex, and S. vetulus) in the gut contents of L. macrochirus; whereas, few epiphytic cladoceran species (i.e., C. sphaericus, A. retangula, and P. laevis) were observed. These findings corresponded to all investigated wetlands. The selective consumption of pelagic cladoceran species by L. macrochirus is interpreted as a result of the efficiency of locating prey in areas with high aquatic macrophyte coverage. The frequent movement of pelagic species to consume food items, such as phytoplankton and bacteria, contributed positively to the search for food by L. macrochirus [72], which led to the high consumption of pelagic cladoceran species. In contrast, epiphytic cladoceran species are difficult for L. macrochirus to search because they have relatively little movement [73]. Although the cladoceran community had relatively rapid population growth, the abundance of pelagic cladocerans was low, and epiphytic cladoceran species were abundant due to the selective consumption of pelagic cladoceran species by L. macrochirus. Epiphytic cladocerans are mainly distributed by attachment to the leaves and stem surfaces of aquatic macrophytes. Therefore, in summer and winter, when aquatic macrophytes are abundant, they not only provide suitable habitats for epiphytic cladocerans but also provide conditions for population growth at a high density by increasing the refuges that are available to cladocerans to avoid predators.
4.3. Effect of Exotic Fish Introduction on Cladoceran Diversity
Based on this study, we surmised that there were three major requirements for the introduction and stable settlement of L. macrochirus (i.e., an exotic fish species) in wetlands or shallow reservoirs located in the Nakdong River basin, including the study sites. First, the space covered by aquatic macrophytes was large. Agricultural lands widely distributed around wetlands promote the inflow of nutrients such as nitrogen and phosphorus, which provide favorable conditions for aquatic macrophyte growth. Second, the diversity and abundance of food sources were high. The area covered by aquatic macrophytes was used as a microhabitat and refuge for cladocerans, which would have contributed to the high cladoceran species diversity and abundance. Prior to the introduction of L. macrochirus, aquatic macrophytes were an effective refuge for pelagic and epiphytic cladoceran species. Therefore, it is speculated that areas covered by aquatic macrophytes have high cladoceran species diversity and abundance. Third, predators of L. macrochirus were less abundant in the Korean freshwater ecosystem. Although Channa argus is the only predator of L. macrochirus, it is frequently captured by humans in the Nakdong River basin where the study sites are located; therefore, C. argus has minimal impact on L. macrochirus populations [27]. Therefore, L. macrochirus settled stably in wetlands and shallow reservoirs where aquatic macrophytes were abundant and likely had a negative effect on various native fish and invertebrates such as cladocerans.
Prey–predator interactions, such as relationships between fish and cladocerans in freshwater ecosystems, are formed by coexistence for a considerably long-term period and by maintaining appropriate relationships based on the efficacy of various defense strategies for avoiding predators based on evolutionary arms races [74]. In this regard, epiphytic species such as C. sphaericus and A. retangula have highly efficient morphological defense strategies to avoid fish predation, which are advantageous for adaptation in areas where aquatic macrophytes are abundant [75]. Vertical and horizontal migration [76,77,78] to avoid predators incurs high costs across all generations; however, morphological changes incur relatively low costs [79]. For example, pelagic cladocerans, to avoid fish predators, move quickly or migrate to a space (e.g., deep water and/or areas covered by aquatic macrophytes) that fish do not prefer; whereas, epiphytic cladocerans have minimal movement and are not easily located by fish foraging activities as they attach to the leaves and stems of aquatic macrophytes. Thus, the distribution of epiphytic cladocerans is more dependent on changes in the microhabitat structure associated with the seasonal growth of aquatic macrophytes than the effects of fish predation [80]. Hence, epiphytic cladoceran species can dominate over pelagic cladoceran species in areas where aquatic macrophytes are abundant.
Nevertheless, C. sphaericus and A. retangula, which belong to the family Chydoridae, were rarely dominant in the study areas. These species prefer summer and autumn, when the water temperature is high, and are closely related to the aquatic macrophyte abundance; therefore, it is difficult for these species to exist at a high density unless these two conditions are met [81,82]. In addition, even if they are less competitive in terms of prey consumption, they are likely to lag in food competition when coexisting with pelagic cladoceran species such as Daphnia spp. Filtration consumption of Daphnia on phytoplankton is powerful enough to cause a ‘Clear-water Phase’ [83]. Epiphytic cladocerans consume algae that are attached to aquatic macrophyte leaves and stems; however, food competition is inevitable when coexisting with pelagic cladoceran species distributed in similar spaces because they primarily feed on phytoplankton or bacteria attached to the leaves and stems of aquatic macrophytes [61]. In addition, pelagic cladocerans can easily dominate because they have higher fecundity and ability to occupy than epiphytic cladocerans; therefore, the presence of pelagic cladocerans can lead to a decrease in epiphytic cladocerans. Therefore, even if the environmental conditions of high water temperature and high plant abundance are satisfied, the condition of a low density of pelagic cladocerans is required to allow epiphytic cladocerans to dominate.
Although the areas with high aquatic macrophyte coverage in South Korea facilitated stable settlement of L. macrochirus and the resulting change in the cladoceran community structure, it can be said that they positively contributed to cladoceran species diversity. The use of the cladoceran community as a food source also occurs in other fish species (i.e., P. parva; [84]), and fish predation had an extremely negative effect on cladoceran survival and population growth in freshwater ecosystems where aquatic macrophytes are minimal or absent. Most freshwater ecosystems in which water sources are permanently maintained are supported by a moderate abundance of fish, which is a major cause of low cladoceran species diversity. In this regard, higher density and species diversity of epiphytic cladocerans can be found in wetlands where macrophytes are abundant, including the study sites, than in other freshwater ecosystems (e.g., streams or reservoirs). Our study shows that the foraging activity of L. macrochirus in areas with high aquatic macrophyte coverage can also contribute to high cladoceran species diversity. In the absence of biological disturbances such as fish predation, cladocerans can accelerate interactions such as intra-competition which can lead to the dominance of one or two species.
In areas where aquatic macrophytes are abundant, food consumption by L. macrochirus is likely to expand to predation on epiphytic cladoceran species when the density of pelagic cladoceran species is lower in the future. Frequent consumption of pelagic cladocerans by L. macrochirus in such areas can lead to a decrease or extinction of pelagic cladocerans; consequently, a decrease in pelagic cladocerans can encourage consumption of an alternative food source by L. macrochirus. Such a result may require different defense strategies in another direction for epiphytic cladocerans to avoid predators. This evolutionary direction can also lead to other prey–predator interactions. Consequently, this change is likely to lead to a trophic cascade effect throughout freshwater food webs in the study area.
5. Conclusions
The results of this study suggest that selective consumption of pelagic cladoceran species (D. obtusa, D. pulex, and S. vetulus) by L. macrochirus can change the cladoceran community structure. L. macrochirus was able to efficiently prey on pelagic cladocerans, even in areas with excessive aquatic macrophyte coverage, which led to the re-examination of the role of aquatic macrophytes as a refuge for cladocerans to avoid fish predation, as suggested in previous studies. Visual foraging activities of L. macrochirus facilitated the search for pelagic cladocerans with frequent movements (due to preying on phytoplankton or bacteria), which led to higher consumption compared with that of epiphytic cladocerans with relatively little movement. Based on the consumption tendency and characteristics of L. macrochirus, there is a high possibility of this species dispersing and settling around wetlands or shallow reservoirs with high aquatic macrophyte coverage, thereby leading to a trophic cascade change in freshwater food webs.
Author Contributions
Conceptualization, J.-Y.C.; methodology, S.-K.K.; validation, J.-Y.C. and S.-K.K.; formal analysis, J.-Y.C.; investigation, J.-Y.C. and S.-K.K.; resources, J.-Y.C.; data curation, J.-Y.C.; writing—original draft preparation, J.-Y.C. and S.-K.K.; writing—review and editing, J.-Y.C.; visualization, J.-Y.C.; supervision, J.-Y.C.; project administration, J.-Y.C.; funding acquisition, S.-K.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by a grant from the National Institute of Ecology (NIE), funded by the Ministry of Environment (MOE) of South Korea (NIE-A-2022-01).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available upon request from the corresponding author. The data are not publicly available because of restrictions on the right to privacy.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Caswell, H. Community structure: A neutral model analysis. Ecol. Monogr. 1976, 46, 327–354. [Google Scholar] [CrossRef]
- Devries, P.J.; Walla, T.R. Species diversity and community structure in neotropical fruit-feeding butterflies. Biol. J. Linn. Soc. Lond. 2001, 74, 1–15. [Google Scholar] [CrossRef]
- Heino, J. A macroecological perspective of diversity patterns in the freshwater realm. Freshw. Biol. 2011, 56, 1703–1722. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Violle, C.; Spasojevic, M.J.; McGill, B.; Damschen, E.; Harrison, S.; Enquist, B.J. Intra-specific and inter-specific variation in specific leaf area reveal the importance of abiotic and biotic drivers of species diversity across elevation and latitude. J. Veg. Sci. 2013, 24, 921–931. [Google Scholar] [CrossRef]
- Menge, B.A. Organization of the New England rocky intertidal community: Role of predation, competition, and environmental heterogeneity. Ecol. Monogr. 1976, 46, 355–393. [Google Scholar] [CrossRef]
- Chesson, P.; Kuang, J.J. The interaction between predation and competition. Nature 2008, 456, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Hambright, K.D.; Zohary, T. Phytoplankton species diversity control through competitive exclusion and physical disturbances. Limnol. Oceanogr. 2000, 45, 110–122. [Google Scholar] [CrossRef]
- Del Pozo, R.; Fernández-Aláez, C.; Fernández-Aláez, M. The relative importance of natural and anthropogenic effects on community composition of aquatic macrophytes in Mediterranean ponds. Mar. Freshw. Res. 2011, 62, 101–109. [Google Scholar] [CrossRef]
- Ball, S.L.; Baker, R.L. The non-lethal effects of predators and the influence of food availability on life history of adult Chironomus tentans (Diptera: Chironomidae). Freshw. Biol. 1995, 34, 1–12. [Google Scholar] [CrossRef]
- Casatti, L.; Mendes, H.F.; Ferreira, K.M. Aquatic macrophytes as feeding site for small fishes in the Rosana Reservoir, Paranapanema River, Southeastern Brazil. Bra. J. Biol. 2003, 63, 213–222. [Google Scholar] [CrossRef]
- Nunn, A.D.; Tewson, L.H.; Cowx, I.G. The foraging ecology of larval and juvenile fishes. Rev. Fish Biol. Fish. 2012, 22, 377–408. [Google Scholar] [CrossRef]
- Wipfli, M.S. Terrestrial invertebrates as salmonid prey and nitrogen sources in streams: Contrasting old-growth and young-growth riparian forests in southeastern Alaska, USA. Can. J. Fish Aquat. Sci. 1997, 54, 1259–1269. [Google Scholar] [CrossRef]
- Dodson, S.L.; Cáceres, C.E.; Rogers, D.C. Cladocera and other Branchiopoda. In Ecology and Classification of North American Freshwater Invertebrates; Academic Press: Cambridge, MA, USA, 2010; pp. 773–827. [Google Scholar]
- Goulden, C.E.; Henry, L.L.; Tessier, A.J. Body size, energy reserves, and competitive ability in three species of Cladocera. Ecology 1982, 63, 1780–1789. [Google Scholar] [CrossRef]
- Ramírez-García, P. Life table demography and population growth of Daphnia laevis (Cladocera, Anomopoda) under different densities of Chlorella vulgaris and Microcystis aeruginosa. Crustaceana 2000, 73, 1273–1286. [Google Scholar] [CrossRef]
- Kirk, K.L.; Gilbert, J.J. Suspended clay and the population dynamics of planktonic rotifers and cladocerans. Ecology 1990, 71, 1741–1755. [Google Scholar] [CrossRef]
- Gayosso-Morales, M.A.; Nandini, S.; Martínez-Jeronimo, F.F.; Sarma, S.S.S. Fish-mediated zooplankton community structure in shallow turbid waters: A mesocosm study. Wetl. Ecol. Manag. 2019, 27, 651–661. [Google Scholar] [CrossRef]
- Tollrian, R.; Dodson, S.I. Inducible defenses in Cladocera: Constraints, costs, and multipredator environments. In The Ecology and Evolution of Inducible Defenses; Princeton University Press: Princeton, NJ, USA, 1999; pp. 177–202. [Google Scholar]
- Dzialowski, A.R.; Lennon, J.T.; O’Brien, W.J.; Smith, V.H. Predator-induced phenotypic plasticity in the exotic cladoceran Daphnia lumholtzi. Freshw. Biol. 2003, 48, 1593–1602. [Google Scholar] [CrossRef] [Green Version]
- Von Elert, E.; Pohnert, G. Predator specificity of kairomones in diel vertical migration of Daphnia: A chemical approach. Oikos 2000, 88, 119–128. [Google Scholar] [CrossRef]
- Jeppesen, E.; Lauridsen, T.L.; Kairesalo, T.; Perrow, M.R. Impact of submerged macrophytes on fish-zooplankton interactions in lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 91–114. [Google Scholar]
- Choi, J.Y.; Jeong, K.S.; Kim, S.K.; La, G.H.; Chang, K.H.; Joo, G.J. Role of macrophytes as microhabitats for zooplankton community in lentic freshwater ecosystems of South Korea. Ecol. Inform. 2014, 24, 177–185. [Google Scholar] [CrossRef]
- Zeng, L.; He, F.; Dai, Z.; Xu, D.; Liu, B.; Zhou, Q.; Wu, Z. Effect of submerged macrophyte restoration on improving aquatic ecosystem in a subtropical, shallow lake. Ecol. Eng. 2017, 106, 578–587. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Thomaz, S.M.; Warfe, D.M. Habitat complexity: Approaches and future directions. Hydrobiologia 2012, 685, 1–17. [Google Scholar] [CrossRef]
- Rossa, D.C.; Lansac-Tôha, F.A.; Bonecker, C.C.; Velho, L.F.M. Abundance of cladocerans in the littoral regions of two environments of the upper Paraná river floodplain, Mato Grosso do Sul, Brazil. Braz. J. Biol. 2001, 61, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Dibble, E.D.; Pelicice, F.M. Influence of aquatic plant-specific habitat on an assemblage of small neotropical floodplain fishes. Ecol. Freshw. Fish 2010, 19, 381–389. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. Effects of aquatic macrophytes on spatial distribution and feeding habits of exotic fish species Lepomis macrochirus and Micropterus salmoides in shallow reservoirs in South Korea. Sustainability 2020, 12, 1447. [Google Scholar] [CrossRef] [Green Version]
- Sakuma, M.; Hanazato, T.; Saji, A.; Nakazato, R. Migration from plant to plant: An important factor controlling densities of the epiphytic cladoceran Alona (Chydoridae, Anomopoda) on lake vegetation. Limnology 2004, 5, 17–23. [Google Scholar]
- Basińska, A.M.; Antczak, M.; Świdnicki, K.; Jassey, V.E.; Kuczyńska-Kippen, N. Habitat type as strongest predictor of the body size distribution of Chydorus sphaericus (OF Müller) in small water bodies. Int. Rev. Hydrobiol. 2014, 99, 382–392. [Google Scholar] [CrossRef]
- Belyaeva, M.A. Littoral Cladocera (Crustacea: Branchiopoda) from the Altai mountain lakes, with remarks on the taxonomy of Chydorus sphaericus (OF Müller, 1776). Arthropoda Sel. 2003, 12, 171–182. [Google Scholar]
- Adamczuk, M. Niche separation by littoral-benthic Chydoridae (Cladocera, Crustacea) in a deep lake-potential drivers of their distribution and role in littoral-pelagic coupling. J. Limnol. 2014, 73, 490–501. [Google Scholar] [CrossRef] [Green Version]
- De Eyto, E.; Irvine, K. The response of three chydorid species to temperature, pH and food. Hydrobiologia 2001, 459, 165–172. [Google Scholar] [CrossRef]
- Muro-Cruz, G.; Nandini, S.; Sarma, S.S.S. Comparative life table demography and population growth of Alona rectangula and Macrothrix triserialis (Cladocera: Crustacea) in relation to algal (Chlorella vulgaris) food density. J. Freshw. Ecol. 2002, 17, 1–11. [Google Scholar] [CrossRef]
- Jang, M.H.; Kim, J.G.; Park, S.B.; Jeong, K.S.; Cho, G.I.; Joo, G.J. The current status of the distribution of introduced fish in large river systems of South Korea. Int. Rev. Hydrobiol. 2002, 87, 319–328. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. Effect of the human utilization of northern snakehead (Channa argus Cantor, 1842) on the settlement of exotic fish and cladoceran community structure. Sustainability 2021, 13, 2486. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Tsukada, H.; Shiah, F.K. Latitudinal gradient in the ovary weight of bluegill Lepomis macrochirus in Lake Biwa, Japan. Ethol. Ecol. Evol. 2011, 23, 291–299. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Shiah, F.K. Spatial variation in reproductive measures of female bluegill Lepomis macrochirus Rafinesque in a small lake with diverse thermal conditions. Zool. Stud. 2013, 52, 19. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K.; Kim, J.C.; Kwon, S.J. Habitat preferences and trophic position of Brachydiplax chalybea flavovittata Ris, 1911 (Insecta: Odonata) larvae in Youngsan River wetlands of South Korea. Insects 2020, 11, 273. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.Y.; Jeong, K.S.; Kim, H.W.; Chang, K.H.; Joo, G.J. Inter-annual variability of a zooplankton community: The importance of summer concentrated rainfall in a regulated river ecosystem. J. Ecol. Environ. 2011, 34, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K. Responses of rotifer community to microhabitat changes caused by summer-concentrated rainfall in a shallow reservoir, South Korea. Diversity 2020, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, R.G.; Likens, G.E. Limnological Analyses; Springer: New York, NY, USA, 2000. [Google Scholar]
- Haney, J.F.; Hall, D.J. Sugar-coated Daphnia: A preservation technique for Cladocera. Limnol. Oceanogr. 1973, 18, 331–333. [Google Scholar] [CrossRef]
- Mizuno, T.; Takahashi, E. An Illustrated Guide to Freshwater Zooplankton in Japan; Tokai University Press: Tokyo, Japan, 1999; p. 532. [Google Scholar]
- Kim, I.S.; Park, J.Y. Freshwater Fish of Korea; Kyo Hak Publishing: Seoul, Korea, 2002. [Google Scholar]
- Nelson, J.S.; Grande, T.C.; Wilson, M.V. Fishes of the World; John Wiley & Sons: Hoboken, NJ, USA, 2016. [Google Scholar]
- Clarke, K.R. Non-parametric multivariate analysis of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Package ‘Vegan’. Community Ecology Package, Version 2. 2013. Available online: http://cran.r-project.org/web/packages/vegan/index.html (accessed on 23 March 2022).
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Fenger-Grøn, M.; Bramm, M.E.; Sandby, K.; Møller, P.H.; Rasmussen, H.U. Impact of fish predation on cladoceran body weight distribution and zooplankton grazing in lakes during winter. Freshw. Biol. 2004, 49, 432–447. [Google Scholar] [CrossRef]
- Ginter, K.; Blank, K.; Haberman, J.; Kangur, A.; Kangur, K. Fish predation pressure on zooplankton in a large northern temperate lake: Impact of adult predators versus juvenile predators. Proc. Est. Acad. Sci. 2018, 65, 356–367. [Google Scholar] [CrossRef]
- Choi, J.Y.; Kim, S.K. A study of the distribution of Daphnia obtusa and Simocephalus vetulus in response to varying environmental conditions using field and microcosm approaches. Water 2020, 12, 815. [Google Scholar] [CrossRef]
- Graeb, B.D.; Dettmers, J.M.; Wahl, D.H.; Cáceres, C.E. Fish size and prey availability affect growth, survival, prey selection, and foraging behavior of larval yellow perch. Trans. Am. Fish. Soc. 2004, 133, 504–514. [Google Scholar] [CrossRef]
- Kim, S.K.; Joo, G.J.; Choi, J.Y. Effect of gated weir opening on the topography and zooplankton community of Geum River, South Korea. Land 2022, 11, 529. [Google Scholar] [CrossRef]
- Kim, H.W.; Choi, J.Y.; La, G.H.; Jeong, K.S.; Jo, G.J. Relationship between rainfall and zooplankton community dynamics in a riverine wetland ecosystem (Upo). Korean J. Ecol. Environ. 2010, 43, 129–135. [Google Scholar]
- Thomaz, S.M.; Cunha, E.R.D. The role of macrophytes in habitat structuring in aquatic ecosystems: Methods of measurement, causes and consequences on animal assemblages’ composition and biodiversity. Acta Limnol. Bras. 2010, 22, 218–236. [Google Scholar] [CrossRef]
- Kaufmann, P.R.; Hughes, R.M.; Van Sickle, J.; Whittier, T.R.; Seeliger, C.W.; Paulsen, S.G. Lakeshore and littoral physical habitat structure: A field survey method and its precision. Lake Reserv. Manag. 2014, 30, 157–176. [Google Scholar] [CrossRef]
- Son, S.H.; Kwon, S.J.; Im, J.H.; Kim, S.K.; Kong, D.; Choi, J.Y. Aquatic macrophytes determine the spatial distribution of invertebrates in a shallow reservoir. Water 2021, 13, 1455. [Google Scholar] [CrossRef]
- Rantala, M.J.; Ilmonen, J.; Koskimäki, J.; Suhonen, J.; Tynkkynen, K. The macrophyte, Stratiotes aloides, protects larvae of dragonfly Aeshna viridis against fish predation. Aquat. Ecol. 2004, 38, 77–82. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; Joo, G.J. Rainfall as dominant driver of rotifer dynamics in shallow wetlands: Evidence from a long-term data record (Upo wetlands, South Korea). Int. Rev. Hydrobiol. 2015, 100, 21–33. [Google Scholar] [CrossRef]
- Cheruvelil, K.S.; Soranno, P.A.; Madsen, J.D.; Roberson, M.J. Plant architecture and epiphytic macroinvertebrate communities: The role of an exotic dissected macrophyte. J. N. Am. Benthol. Soc. 2002, 21, 261–277. [Google Scholar] [CrossRef]
- Choi, J.Y.; Jeong, K.S.; La, G.H.; Kim, S.K.; Joo, G.J. Sustainment of epiphytic microinvertebrate assemblage in relation with different aquatic plant microhabitats in freshwater wetlands (South Korea). J. Limnol. 2014, 73, 197–202. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.Y.; Kim, S.K.; Jeong, K.S.; Joo, G.J. Distribution pattern of epiphytic microcrustaceans in relation to different macrophyte microhabitats in a shallow wetland (Upo wetlands, South Korea). Oceanol. Hydrobiol. Stud. 2015, 44, 151–163. [Google Scholar] [CrossRef]
- Kemp, W.M.; Batleson, R.; Bergstrom, P.; Carter, V.; Gallegos, C.L.; Hunley, W.; Karrh, L.; Koch, E.W.; Landwehr, J.M.; Moore, K.A.; et al. Habitat requirements for submerged aquatic vegetation in Chesapeake Bay: Water quality, light regime, and physical-chemical factors. Estuaries 2004, 27, 363–377. [Google Scholar] [CrossRef]
- Lu, J.; Wang, Z.; Xing, W.; Liu, G. Effects of substrate and shading on the growth of two submerged macrophytes. Hydrobiologia 2013, 700, 157–167. [Google Scholar] [CrossRef]
- Manatunge, J.; Asaeda, T.; Priyadarshana, T. The influence of structural complexity on fish–zooplankton interactions: A study using artificial submerged macrophytes. Environ. Biol. Fishes 2000, 58, 425–438. [Google Scholar] [CrossRef]
- Deosti, S.; de Fátima Bomfim, F.; Lansac-Tôha, F.M.; Quirino, B.A.; Bonecker, C.C.; Lansac-Tôha, F.A. Zooplankton taxonomic and functional structure is determined by macrophytes and fish predation in a Neotropical river. Hydrobiologia 2021, 848, 1475–1490. [Google Scholar] [CrossRef]
- Post, D.M. Individual variation in the timing of ontogenetic niche shifts in largemouth bass. Ecology 2003, 84, 1298–1310. [Google Scholar] [CrossRef]
- Jang, M.H.; Joo, G.J.; Lucas, M.C. Diet of introduced largemouth bass in Korean rivers and potential interactions with native fishes. Ecol. Freshw. Fish 2006, 15, 315–320. [Google Scholar] [CrossRef]
- Szedlmayer, S.T.; Lee, J.D. Diet shifts of juvenile red snapper (Lutjanus campechanus) with changes in habitat and fish size. Fish. Bull. 2004, 102, 366–375. [Google Scholar]
- Huss, M.; Byström, P.; Persson, L. Resource heterogeneity, diet shifts and intra-cohort competition: Effects on size divergence in YOY fish. Oecologia 2008, 158, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Holmes, T.H.; McCormick, M.I. Size-selectivity of predatory reef fish on juvenile prey. Mar. Ecol. Prog. Ser. 2010, 399, 273–283. [Google Scholar] [CrossRef]
- Dabrowski, K.; Portella, M.C. Feeding plasticity and nutritional physiology in tropical fishes. Fish. Physiol. 2005, 21, 155–224. [Google Scholar]
- Yamada, S.; Urabe, J. Role of sediment in determining the vulnerability of three littoral cladoceran species to odonate larvae predation. Inland Waters 2021, 11, 154–161. [Google Scholar] [CrossRef]
- Theel, H.J.; Dibble, E.D. An experimental simulation of an exotic aquatic macrophyte invasion and its influence on foraging behavior of bluegill. J. Freshw. Ecol. 2008, 23, 79–89. [Google Scholar] [CrossRef]
- Nolfi, S.; Floreano, D. Coevolving predator and prey robots: Do “arms races” arise in artificial evolution? Artif. life 1998, 4, 311–335. [Google Scholar] [CrossRef] [Green Version]
- Boeing, W.J.; Wissel, B.; Ramcharan, C.W. Can costs and benefits of Daphnia defense against fish be measured in a field experiment? Fundam. Appl. Limnol. 2010, 176, 1. [Google Scholar] [CrossRef]
- Berge, J.; Cottier, F.; Varpe, Ø.; Renaud, P.E.; Falk-Petersen, S.; Kwasniewski, S.; Griffiths, C.; Søreide, J.E.; Johnsen, G.; Aubert, A.; et al. Arctic complexity: A case study on diel vertical migration of zooplankton. J. Plankton Res. 2014, 36, 1279–1297. [Google Scholar] [CrossRef]
- Kim, S.K.; Choi, J.Y. Differences in the vertical distribution of two cladoceran species in the Nakdong River estuary, South Korea. Water 2020, 12, 2154. [Google Scholar] [CrossRef]
- Iglesias, C.; Goyenola, G.; Mazzeo, N.; Meerhoff, M.; Rodo, E.; Jeppesen, E. Horizontal dynamics of zooplankton in subtropical Lake Bianca (Uruguay) hosting multiple zooplankton predators and aquatic plant refuges. In Shallow Lakes in a Changing World; Springer: Dordrecht, The Netherlands, 2007; pp. 179–189. [Google Scholar]
- Diel, P.; Kiene, M.; Martin-Creuzburg, D.; Laforsch, C. Knowing the enemy: Inducible defences in freshwater zooplankton. Diversity 2020, 12, 147. [Google Scholar] [CrossRef] [Green Version]
- Kuczyńska-Kippen, N.M.; Nagengast, B. The influence of the spatial structure of hydromacrophytes and differentiating habitat on the structure of rotifer and cladoceran communities. Hydrobiologia 2006, 559, 203–212. [Google Scholar] [CrossRef]
- Ali, M.M.; Mageed, A.A.; Heikal, M. Importance of aquatic macrophyte for invertebrate diversity in large subtropical reservoir. Limnologica 2007, 37, 155–169. [Google Scholar] [CrossRef] [Green Version]
- Bucak, T.; SaraoĞLu, E.C.E.; Levi, E.E.; Nihan Tavşanoğlu, Ü.; Çakiroğlu, A.I.; Jeppesen, E.; Beklioğlu, M. The influence of water level on macrophyte growth and trophic interactions in eutrophic Mediterranean shallow lakes: A mesocosm experiment with and without fish. Freshw. Biol. 2012, 57, 1631–1642. [Google Scholar] [CrossRef]
- Van Onsem, S.; De Backer, S.; Triest, L. Microhabitat–zooplankton relationship in extensive macrophyte vegetations of eutrophic clear-water ponds. Hydrobiologia 2010, 656, 67–81. [Google Scholar] [CrossRef]
- Musil, M.; Novotná, K.; Potužák, J.; Hůda, J.; Pechar, L. Impact of topmouth gudgeon (Pseudorasbora parva) on production of common carp (Cyprinus carpio)-question of natural food structure. Biologia 2014, 69, 1757–1769. [Google Scholar] [CrossRef]
Figure 1.
Study site locations. The study sites (Upo, Jangcheok, and Beongae wetlands) located in southeast South Korea are indicated by solid circles (●). The small map in the upper right-hand corner shows the Korean Peninsula, and the Nakdong River is indicated in yellow.
Figure 1.
Study site locations. The study sites (Upo, Jangcheok, and Beongae wetlands) located in southeast South Korea are indicated by solid circles (●). The small map in the upper right-hand corner shows the Korean Peninsula, and the Nakdong River is indicated in yellow.

Figure 2.
Time-series fluctuations in dominant cladoceran species in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland from 2015 to 2017.
Figure 2.
Time-series fluctuations in dominant cladoceran species in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland from 2015 to 2017.

Figure 3.
Non-metric multidimensional scaling (NMDS) of five dominant rotifer species (letters) and 79 sampling times (dots). Blue arrows represent the associations with environmental variables. Sampling times are divided into four seasons (spring, summer, autumn, and winter). Wt: water temperature; DO: dissolved oxygen; Tur: turbidity; Cs: Chydorus shpaericus; Ar: Alona retangula; Sv: Simocephalus vetulus; Db: Diaphanozoma brachyrum; Mr: Macrotrix rosea; Cd: Ceriodaphnia dubia; La: Latonopsis australis; Pt: Pleuroxus trigonellus. (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.
Figure 3.
Non-metric multidimensional scaling (NMDS) of five dominant rotifer species (letters) and 79 sampling times (dots). Blue arrows represent the associations with environmental variables. Sampling times are divided into four seasons (spring, summer, autumn, and winter). Wt: water temperature; DO: dissolved oxygen; Tur: turbidity; Cs: Chydorus shpaericus; Ar: Alona retangula; Sv: Simocephalus vetulus; Db: Diaphanozoma brachyrum; Mr: Macrotrix rosea; Cd: Ceriodaphnia dubia; La: Latonopsis australis; Pt: Pleuroxus trigonellus. (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.

Figure 4.
Abundance of dominant cladoceran species according to five microhabitat types. Ow: open water; Pd: the area covered by Paspalum distichum; Tj: the area covered by Trapa japonica; Floating: the area covered by floating-leaved macrophytes (Hydrocharis dubia and Salvinia natans); Mixed: the area covered by floating-leaved and submerged macrophytes (Hydrocharis dubia, Salvinia natans, Ceratophyllum demersum, Hydrilla verticillata, and Najas graminea). Chydorus shpaericus and Alona retangula are epiphytic cladocerans species, and the remaining three species (Simocephalus vetulus, Diaphanozoma brachyrum, and Latonopsis australis) are pelagic. (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.
Figure 4.
Abundance of dominant cladoceran species according to five microhabitat types. Ow: open water; Pd: the area covered by Paspalum distichum; Tj: the area covered by Trapa japonica; Floating: the area covered by floating-leaved macrophytes (Hydrocharis dubia and Salvinia natans); Mixed: the area covered by floating-leaved and submerged macrophytes (Hydrocharis dubia, Salvinia natans, Ceratophyllum demersum, Hydrilla verticillata, and Najas graminea). Chydorus shpaericus and Alona retangula are epiphytic cladocerans species, and the remaining three species (Simocephalus vetulus, Diaphanozoma brachyrum, and Latonopsis australis) are pelagic. (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.

Figure 5.
Seasonal distributions of fish communities in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.
Figure 5.
Seasonal distributions of fish communities in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.

Figure 6.
Seasonal body lengths of Lepomis macrochirus in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.
Figure 6.
Seasonal body lengths of Lepomis macrochirus in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.

Figure 7.
Lepomis macrochirus abundance according to five microhabitat types. Ow: open water; Pd: area covered by Paspalum distichum; Tj: area covered by Trapa japonica; Floating: area covered by floating-leaved macrophytes (Hydrocharis dubia and Salvinia natans); Mixed: area covered by floating-leaved and submerged macrophytes (Hydrocharis dubia, Salvinia natans, Ceratophyllum demersum, Hydrilla verticillata, and Najas graminea). in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.
Figure 7.
Lepomis macrochirus abundance according to five microhabitat types. Ow: open water; Pd: area covered by Paspalum distichum; Tj: area covered by Trapa japonica; Floating: area covered by floating-leaved macrophytes (Hydrocharis dubia and Salvinia natans); Mixed: area covered by floating-leaved and submerged macrophytes (Hydrocharis dubia, Salvinia natans, Ceratophyllum demersum, Hydrilla verticillata, and Najas graminea). in (a) Upo Wetland, (b) Jangcheok Wetland, and (c) Beongae Wetland.

Figure 8.
Seasonal food utilization of cladoceran groups by Lepomis macrochirus in three L. macrochirus size ranges (i.e., >5 cm, 6–10 cm, and 11–15 cm) based on L. macrochirus body length in three wetlands: (a) <5 cm body length in Upo Wetland, (b) 6–10 cm body length in Upo Wetland, (c) 11–15 cm body length in Upo Wetland, (d) <5 cm body length in Jangcheok Wetland, (e) 6–10 cm body length in Jangcheok Wetland, (f) 11–15 cm body length in Jangcheok Wetland, (g) <5 cm body length in Beongae Wetland, (h) 6–10 cm body length in Beongae Wetland, and (i) 11–15 cm body length in Beongae Wetland. Chydorus shpaericus, Alona retangula, and Pleroxus laevis are epiphytic cladocerans species, and the remaining species are pelagic.
Figure 8.
Seasonal food utilization of cladoceran groups by Lepomis macrochirus in three L. macrochirus size ranges (i.e., >5 cm, 6–10 cm, and 11–15 cm) based on L. macrochirus body length in three wetlands: (a) <5 cm body length in Upo Wetland, (b) 6–10 cm body length in Upo Wetland, (c) 11–15 cm body length in Upo Wetland, (d) <5 cm body length in Jangcheok Wetland, (e) 6–10 cm body length in Jangcheok Wetland, (f) 11–15 cm body length in Jangcheok Wetland, (g) <5 cm body length in Beongae Wetland, (h) 6–10 cm body length in Beongae Wetland, and (i) 11–15 cm body length in Beongae Wetland. Chydorus shpaericus, Alona retangula, and Pleroxus laevis are epiphytic cladocerans species, and the remaining species are pelagic.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Seasonal variations in environmental variables measured at three wetlands (Upo, Jangcheok, and Beongae wetlands). Dwm, dry weight (g) of macrophyte; WT, water temperature (°C); DO, % saturation of dissolved oxygen; Cond., conductivity (µS cm−1); Tur., Turbidity (NTU); Chl a, chlorophyll a (µg L−1). Each season consisted of 3 months: Spring, March–May; Summer, June–August; Autumn, September–November; Winter, December–February.
Table 1.
Seasonal variations in environmental variables measured at three wetlands (Upo, Jangcheok, and Beongae wetlands). Dwm, dry weight (g) of macrophyte; WT, water temperature (°C); DO, % saturation of dissolved oxygen; Cond., conductivity (µS cm−1); Tur., Turbidity (NTU); Chl a, chlorophyll a (µg L−1). Each season consisted of 3 months: Spring, March–May; Summer, June–August; Autumn, September–November; Winter, December–February.
| Sites | Variable | Spring (n = 18) | Summer (n = 21) | Autumn (n = 19) | Winter (n = 21) |
|---|---|---|---|---|---|
| Upo | Dwm | 16.2 ± 4.8 | 31.5 ± 10.4 | 48.2 ± 11.6 | 5.1 ± 1.8 |
| WT | 16.3 ± 5.1 | 25.6 ± 2.4 | 16.1 ± 5.8 | 4.9 ± 1.7 | |
| DO | 101 ± 38 | 38 ± 27.8 | 34 ± 26.2 | 84 ± 31.8 | |
| Cond. | 408 ± 134 | 420 ± 152 | 361 ± 81 | 483 ± 131 | |
| pH | 7.8 ± 0.4 | 7.1 ± 0.5 | 7.2 ± 0.5 | 7.7 ± 0.3 | |
| Tur. | 36 ± 14.6 | 27 ± 14.8 | 56 ± 36.8 | 39 ± 25.6 | |
| Chl a | 13.8 ± 4.6 | 18 ± 5.4 | 18 ± 3.2 | 32 ± 2.6 | |
| Jangcheok | Dwm | 12.4 ± 3.7 | 34.8 ± 11.9 | 43.7 ± 14.2 | 8.2 ± 4.7 |
| WT | 16.7 ± 5.7 | 25.9 ± 1.6 | 16.9 ± 5.8 | 3.4 ± 2.6 | |
| DO | 96 ± 40.3 | 26 ± 11.6 | 35 ± 17.1 | 73 ± 37.4 | |
| Cond. | 341 ± 66.7 | 320 ± 243 | 275 ± 156 | 384 ± 246 | |
| pH | 7.6 ± 0.4 | 6.8 ± 0.4 | 6.9 ± 0.6 | 7.8 ± 0.4 | |
| Tur. | 26 ± 14.6 | 28 ± 20.6 | 31 ± 21.2 | 28 ± 13.4 | |
| Chl a | 25 ± 2.3 | 36 ± 4.1 | 31 ± 2.7 | 43 ± 4.1 | |
| Beongae | Dwm | 15.1 ± 7.3 | 37.4 ± 11.4 | 41.8 ± 13.7 | 8.0 ± 3.7 |
| WT | 17.3 ± 5.6 | 26.8 ± 1.8 | 18.2 ± 5.9 | 4.7 ± 2.7 | |
| DO | 97 ± 62.3 | 20 ± 31.2 | 23 ± 14.3 | 90 ± 43.2 | |
| Cond. | 289 ± 117 | 238 ± 118 | 269 ± 184 | 348 ± 216 | |
| pH | 7.4 ± 0.6 | 6.9 ± 0.4 | 7.3 ± 0.3 | 7.7 ± 0.4 | |
| Tur. | 29 ± 17.1 | 27 ± 14.8 | 40 ± 14.7 | 31 ± 24.7 | |
| Chl a | 14 ± 3.4 | 32 ± 3.5 | 27 ± 4.8 | 33 ± 3.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kim, S.-K.; Choi, J.-Y. Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans. Water 2022, 14, 3781. https://doi.org/10.3390/w14223781
AMA Style
Kim S-K, Choi J-Y. Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans. Water. 2022; 14(22):3781. https://doi.org/10.3390/w14223781
Chicago/Turabian StyleKim, Seong-Ki, and Jong-Yun Choi. 2022. "Selective Consumption of Pelagic Cladocerans by Bluegill Sunfish (Lepomis macrochirus Rafinesque) Contributes to Dominance of Epiphytic Cladocerans" Water 14, no. 22: 3781. https://doi.org/10.3390/w14223781
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.