
Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio)
by
Rong Shen
1,*,
Yichang Yu
2,
Qinfang Liu
3,
Ruili Wang
1,
Rong Lan
1,
Hui Feng
1,
Ran Yu
1,
Lihua Jin
1 and
Ye Li
1 1
College of Bioengineering, Beijing Polytechnic, Beijing 100176, China
2
Center of Water Ecological Environment, Chongqing Academy of Ecological and Environmental Science, Southwest Branch of Chinese Academy of Environmental Sciences, Chongqing 401120, China
3
Anhui Provincial Institute for Food and Drug Control, Hefei 230051, China
*
Author to whom correspondence should be addressed.
Water 2022, 14(23), 3857; https://doi.org/10.3390/w14233857
Submission received: 29 September 2022
/
Revised: 21 November 2022
/
Accepted: 24 November 2022
/
Published: 27 November 2022
(This article belongs to the Section Biodiversity and Functionality of Aquatic Ecosystems)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Pefloxacin (PFLX) is a type of fluoroquinolone (FQ) that has been widely used in aquaculture and animal husbandry. It is also a new kind of pseudo-persistent pollutant, which has some toxic effects. In this study, zebrafish (Danio rerio) embryos were used as exposure models to analyze the toxicity of PFLX. The results showed that PFLX was absorbed on the outer membrane surface at the beginning, and then a few PFLX slowly entered the cytoplasm. The adsorption complied with the Langmuir isotherm adsorption model. The surface morphology of the embryonic membrane was changed. PFLX exposure had no effect on fetal autonomous movement, but it significantly inhibited the embryo hatching rate. After PFLX exposure, the embryo and larval survival rates showed a clear dose–effect relationship. Though PFLX exposure did not cause teratogenic toxicity, it still affected cardiovascular function. The potential toxicity mechanism and developmental toxicity of PFLX to zebrafish were revealed. These results could be important for studies concerning FQ toxicology.
1. Introduction
Fluoroquinolone (FQ) is a kind of general medicine for humans and animals [1]. Because of its broad antibacterial spectrum, strong antibacterial activity and no cross-resistance with other antibacterial drugs, it is widely used in many fields [2,3]. It has been detected in urban sewage effluent, surface water, groundwater, and even drinking water [4,5,6] in concentrations ranging from ng/L to mg/L [7,8,9]. Due to the lack of biodegradation pathways and high adsorption affinity, it has become a pseudo-persistent organic pollutant in the environment with a half-life of 10.6 days in surface water and up to 580 days in soil [10]. Some studies have shown that FQs in the environment may be toxic to organisms [11,12,13], such as the inhibition of growth and parthenogenetic reproduction for Daphnia magna [14], adverse effects on the growth of cabbage, tomato, wheat and other crops [15], and even hepatic and renal toxicity for human beings [16]. Zhang et al. [17] reported that the EC50 value of the mixed FQs was 481.3 mg/L, and the mixed FQs showed adverse effects on zebrafish hatching, mortality and malformation. Gatifloxacin and ciprofloxacin have cardiovascular toxicity for zebrafish when the concentrations are higher than 1238 and 1407 mg/L, respectively [18]. Enrofloxacin is ecotoxic for the development, growth and the detoxifying and oxidative stress enzymes of anuran amphibian larvae when the concentration is greater than or equal to 10 μg/L [19].
Pefloxacin (PFLX) is one of the most common FQ pollutants [20] and is harmful to the ecological environment when it enters into the soil and water environment [21,22]. In addition, when PFLX is used in livestock production, it may remain in animal foodstuffs [23], thereby harming human health through the food chain [24]. Based on the risk assessment results, the Ministry of Agriculture and Rural Affairs of China has stopped using PFLX antibacterial drugs for food animals since December 31, 2016. Due to the low metabolic rate of PFLX in humans and animals, most PFLXs enter the water environment through medical waste water, domestic sewage and excreta, causing pollution to the water environment [25]. Moreover, PFLX has toxic effects on animals. It adversely affects the biomechanics and histopathology of the achilles tendon in wistar rats, which undergoes hyaluronic degeneration, fiber dislocation and mucus degeneration [26].
Zebrafish (Danio rerio) is a tropical freshwater fish with the characteristics of convenient breeding, a short breeding cycle, a large egg production, in vitro fertilization of embryos, in vitro development, a transparent embryo body, etc. These characteristics make them an important model species for researchers [27,28]. Furthermore, the zebrafish is highly similar to mammals in biological structure and physiological functions, and has up to 87% homology to human genes, and its signal transduction pathways are basically similar. It has become a model organism for assessing chemical environments and human health risks [29,30,31]. It is also a standard fish for experimental use in the instruction manual of Organisation for Economic Co-operation and Development (OECD), an experimental fish species recommended by International Standardization Organization (ISO), and a new biological substitute animal recommended by the European Centre for the Validation of Alternative Methods (ECVAM) [32]. As a small vertebrate model organism, the zebrafish is a good complement to the biological gap between traditional in vitro cytology and other biological detection methods and mammalian experimental detection, which helps to comprehensively analyze the results of in vitro and in vivo experiments to improve the accuracy and efficiency of drug toxicity analysis.
As far as we know, there is little research on the toxicity and toxicity mechanism of PFLX. However, some researchers have studied how PFLX reacts with embryonic membrane. If PFLX causes developmental toxicity to zebrafish embryos, there must be some reaction between PFLX and the embryonic membrane. In order to understand the toxic mechanism of PFLX, it is necessary to comprehensively analyze the correlation between PFLX and zebrafish toxicity performance. In this study, the interaction between PFLX and the zebrafish embryonic membrane were investigated. The effects of PFLX on embryonic autonomic movement and hatchability, embryo/larval survival rate, heart rate and teratogenic toxicity were also studied. The aim of this work was to elucidate the potential toxicity mechanism and developmental toxicity of PFLX in zebrafish.
2. Experimental
2.1. Apparatus and Materials
High-performance liquid chromatography (HPLC) (Model LC-10A, Shimadzu, Japan), a fluorescence detector (RF-20A, Shimadzu, Japan), a ZORBAX Eclipse Plus C18 reversed-phase column (Eclipse Plus 5 µm, 4.6 × 250 mm, Agilent, Santa Clara, CA, USA), a freezer dryer (K750X, Nanjing kangshi health care products Co., Ltd, Nanjing, China), a scanning electron microscope (SEM) (S-4800, Hitachi, Tokyo, Japan), an anatomical stereoscopic microscope (SZX7, OLYMPUS, Japan), an incubator (THZ-300C, Yiheng scientific instrument Co., Ltd, Shanghai, China), a precision electronic balance (CP214, OHAUS, Parsippany, NJ, USA) and an ultrapure water system (CASCADAIXMK2, PALL, Port Washington, NY, USA) were used for this project.
Analytical standards pefloxacin (PFLX), pefloxacin mesylate dihydrate (≥99%), tetrabutylammonium bromide (ion-pair chromatography, ≥99%), phosphoric acid (chromatography grade, 85–90%) and dimethyl sulfoxide were bought from Aladdin holdings (group) Co., Ltd. (Shanghai, China). Acetonitrile and methanol (HPLC) were bought from Fisher (Boston, USA). Hydrochloric acid, NaHCO3, KCl, CaCl2⋅2H2O and MgSO4⋅7H2O were bought from Sinopharm Chemical Reagent (Shanghai, China). All of them were used without further treatment.
PFLX stock solution was prepared in ultra-pure water by using pefloxacin mesylate dihydrate as solute at a concentration of 0.1 mol/L. It was stored in a refrigerator at 4 °C and diluted with ultrapure water as needed before use.
Reconstituted buffer (RB): 0.294 g CaCl2⋅2H2O, 0.123 g MgSO4⋅7H2O, 0.065 g NaHCO3 and 0.006 g KCl were dissolved in 1000 mL ultrapure water (ISO 6341-1982). It was used for the culture of zebrafish embryos after being fully aerated.
2.2. Zebrafish Husbandry and Embryo Exposure
Mature zebrafish were raised in a 25.0 L glass tank and were obtained from an aquatic products market in Beijing. The water quality condition was as follows: hardness was 250 mg/L measured in CaCO3, pH for 7–8, dissolved oxygen was (10.5 ± 0.5) mg/L. The cultivation conditions met the U.S. Environmental Protection Agency (EPA) standards. Zebrafish were fed 2 times/day with frozen red blood worms, and 1/3 of the total amount of water was changed every day. The light cycle of zebrafish spawning was 14 h of light and 10 h of darkness, and the temperature was between 25 and 27 °C. Male and female zebrafish (1:2) were put into the spawning box the night before the experiment. The zebrafish spawned within 30 min after turning on the light in next morning. Then, the embryos were collected and further treated for the subsequent.
To ensure that the experiments produced valid results, normal embryos were screened with an inverted microscope and the fertilized eggs were obtained only from spawns with a fertilization rate higher than 95%. Six embryos were incubated in a 1.0 mL solution, a control group were incubated in RB, and the experimental exposure group were incubated in different concentrations of PFLX solution. The control group and experimental exposure group both set 5 replicates, with 30 embryos exposed per group. The culture period of the exposed embryos was 12 h/12 h (light/dark). The temperature of the light incubator was set to 26 ± 0.5 °C. The death of the embryo was judged by observing heartbeat, body movements and reaction after stimulation. Dead embryos were immediately removed from the exposed solution. After exposure for a certain period of time, an embryo was randomly selected for observation from each well with a microscope. The embryo mortality, hatching rate, heart rate, exercise status and deformity characteristics were calculated and recorded.
Embryos were not fed for 9 days after fertilization and can survive on nutrients provided by their own yolk sac. After the experiment, zebrafish at various developmental stages were overexposed with tricaine mesylate (0.3 mg/mL) to be killed by anesthesia. The procedure was in accordance with the requirements of the American Veterinary Medical Association (AVMA) for animal anesthesia.
2.3. PFLX Determination
The concentration of PFLX was measured by HPLC. Chromatographic condition was as follows: the ratio of tetrabutylammonium bromide solution (pH 3.1) to acetonitrile was 11:89 (v/v), which was used as mobile phase. Regarding automatic injection, the injection volume was 10 μL, the flow rate was 1.5 mL/min, and the column temperature was 40 °C. A fluorescence detector was used, the excitation wavelength was 280 nm, and the emission wavelength was 450 nm.
2.4. The Interaction between PFLX and Embryonic Membrane
Kinetic experiment: Five embryos were put into a 1 mL 1.00 mM PFLX solution. After a certain period of time, the concentration of the remaining PFLX (cL) was analyzed by HPLC.
Temperature experiment: Five embryos were put into 1 mL 1.00 mM PFLX solution at different temperatures. After 8 h, the concentration of the remaining PFLX (cL) was analyzed by HPLC.
Adsorption experiment: Five embryos were put into a 1 mL PFLX solution. The concentration of PFLX (c0) was set as 0.20, 0.40, 0.60, 0.80, 1.00, 1.21, 1.43, 1.65, 1.87, 2.09, 2.20, 3.00 mM. After 8 h, the concentration of the remaining PFLX (cL) were analyzed by HPLC.
All of the above experiments were set up in three groups in parallel.
SEM morphology of the embryonic membrane surface: After 8 h of culture and exposure, the embryos of the blank control group and the PFLX exposure group (c0 = 1.00 mM) were taken out of the solution. Each group had five embryos. The moisture on the surface of embryo was absorbed and then precooled in the refrigerator at −4 °C for 1 h. The precooled embryos were transferred to a freeze-drying cabinet and freeze-dried at −55 °C for 24 h. After the fixation, the freeze-dried embryos were recorded by SEM [33].
2.5. Embryo Behavior and Toxicity Experiment
The embryos were exposed to PFLX solution in semi-static conditions and 1/3 of the exposure solution was renewed every day. The concentration of PFLX before and after exposure were analyzed to make sure that the difference between these two is controlled within 10% of the newly replaced solution.
Autonomous movement: The embryos (2 h post fertilization, 2 hpf) were randomly selected and added to the PFLX solution, whose concentration were 2.13, 4.26, 6.39, 10.65 and 15.97 mM. The blank control group was also set. After 24 h exposure, the time interal of autonomous movement of embryos within 60 s was recorded.
Hatching, survival and heart rate: The embryos (24 hpf), whose concentrations were 2.13, 4.26, 6.39, 10.65 and 15.97 mM, were randomly selected and added to the PFLX solution. The blank control group was also set. After 24 h exposure, the hatching rate of embryos was recorded. After a certain period of treatment, the survival and heart rate of embryos was recorded.
Embryo teratogenic experiment: The embryos (2 hpf) were randomly selected and added to the PFLX solution. The concentrations were 4.26 and 10.65 mM. The blank control group was also set. After 1 and 2 d exposure, the teratogenicity of embryos was recorded.
2.6. Determination of Exposure Concentration
In the pre-experiment, when the exposure concentration was lower than 2.13 mM, the survival rates of the embryos and larvae were both higher than 90% after 1 d exposure. When the exposure concentration was higher than 15.97 mM, all embryos and larvae were dead after 6 d exposure. These results were used to determine the exposure concentration range (2.13–15.97 mM). The other concentrations were obtained in decreasing proportion.
2.7. Larva Behavior and Toxicity Experiment
Survival and heart rate: The larvae (2 h post hatch, 2 hph), whose concentrations were 2.13, 4.26, 6.39, 10.65 and 15.97 mM were randomly selected and added to the PFLX solution. The blank control group was also set. After 24 and 48 h of treatment, the survival and heart rates of the embryos were recorded.
Larva teratogenic experiment: The larvae (2 hph), whose concentrations were 4.26 and 10.65 mM, were randomly selected and added to the PFLX solution. The blank group and vehicle group were set. After 1, 2 and 3 d of treatment, the teratogenicity of larva was recorded.
2.8. Data Analysis
All data were analyzed using the GraphPad Prism 5.00 (GraphPad Software, San Diego, CA, USA) software. Values were expressed as mean ± standard deviation (SD) and each datum was repeated 3 times, p < 0.05 indicated a significant difference.
3. Results and Discussion
3.1. The Interaction between PFLX and Embryonic Membrane
The binding ratio (γ) between the PFLX and the embryo can be expressed by the following Equation (1):
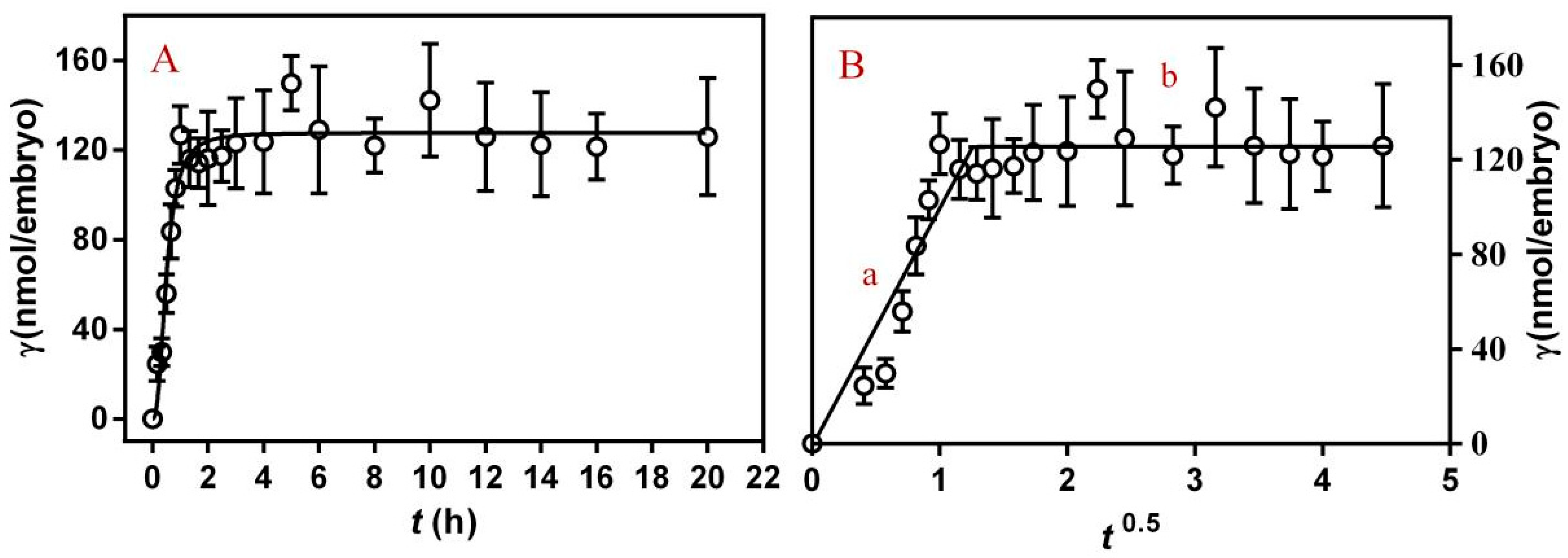
c0 represents the inital concentration of PFLX, cL represents the remaining concentration of PFLX, V represents the volume of PFLX solution and n represents the number of embryos. Figure 1A shows the change of γ with time when PFLX interacts with the embryos. In the first 0.5 h, PFLX was rapidly absorbed on the surface of the embryonic membrane, and the value of γ reached 45% of equilibrium. After 2 h exposure, the value of γ reached the equilibrium and had no obvious change with the increase of exposure time. This is because PFLX has hydrophilicity, which can bind to hydrophilic groups on the surface of embryonic membranes. The equilibrium time between the embryo and PFLX is 2 h. The first 6 h of embryo development is the rapid division of cells. The dividing cells become smaller and smaller, and the cells have similar morphology and structure without being differentiated. Then, the embryos began to differentiate and the adsorption of PFLX had essentially reached equilibrium. Therefore, in the subsequent experiments, 8 h was selected as the equilibrium time, which can ensure all embryonic cells complete division and enter a relatively stable stage of differentiation.
The adsorption process can be analyzed by a diffusion-controlled kinetic model, and its equation is as follows:
γt is the average amount of PFLX adsorbed on each embryo after t h exposure, D is the diffusion coefficient, and S is the surface area of the embryo membrane. The curve is shown in Figure 1B. The whole process is divided into stage a and stage b. Stage a represents a rapid adsorption process. After 2 h of rapid adsorption, the binding of PFLX on the surface of the embryonic membrane is nearly saturated, and then it enters a relatively slow transport process (stage b). In stage b, γt slightly increases with the increase of exposure time. In this stage, part of PFLX molecule adsorbed on the surface of embryo membrane can be carried into the membrane through transport of membrane proteins.
3.2. Effects of PFLX Exposure on Embryonic Membranes
After exposure to different concentration of PFLX for 8 h, the value of γ was calculated according to Equation (1). As shown in Figure 2A, the value of γ shows a nearly linear increase trend with the increasing initial concentration of PFLX. When c0 exceeds 1.21 mM, the value of γ increases but the rate of increase decreases. The adsorption of PFLX on the embryo does not reach saturation during this process. This is because the concentration range of PFLX used in this experiment is narrow and the saturation concentration of PFLX adsorption by the embryo has not been reached. In addition, at a relatively low concentration range, PFLX adsorbed to the membrane is continuously metabolized and transported to different parts of the embryo during embryonic development.
To better illustrate the interaction between PFLX and embryonic membrane, the classical Langmuir isothermal adsorption equation [34] is used to simulate the interaction process. The equation is as follows:
K is the binding constant when PFLX interacts with embryonic membrane, and N is the saturation binding number. The fitting curve was shown in Figure 2B. The embryonic adsorption of PFLX agrees with the Langmuir isothermal adsorption model, and the fitting results indicate that the value of N is 3.79 μmol/embryo, which means one embryo can bind up to 3.79 μmol of PFLX molecule, K is 4.00 × 102 L/μmol.
The large amount of PFLX adsorption on the embryonic membrane, indicating that most of PFLX is trapped outside the membrane and fails to cross the membrane into the cytoplasm. It is difficult to directly cause target “poisoning” and biological “three causes” effects, but it may cause the blockage of the embryonic membrane channels, embryo expansion or even asphyxiation death. PFLX on the surface of the embryonic membrane through the attraction between its positively charged piperazine group (N2C4H9−) with a negatively charged >PO4− group of the embryonic membrane surface [35,36,37]. The negatively charged groups, such as glutamic acid, phosphoethanolamine and phosphoserine, distributing on the surface of the embryonic membrane, could also be attracted to the positively changed piperazine group in the PFLX molecule. While the negatively charged group (–COO−) in the PFLX molecule could be attracted to the positively changed –NR4+ in membrane bilayer. In addition, a variety of hydrophilic groups such as –COOH, –OH and –NH2 are also distributed on the surface of the embryonic membrane. These groups could be combined with the carboxyl group and amino group in the PFLX molecule through hydrogen bonding and van der Waals force.
3.3. Effect of Temperature on the Action of PFLX and Embryo
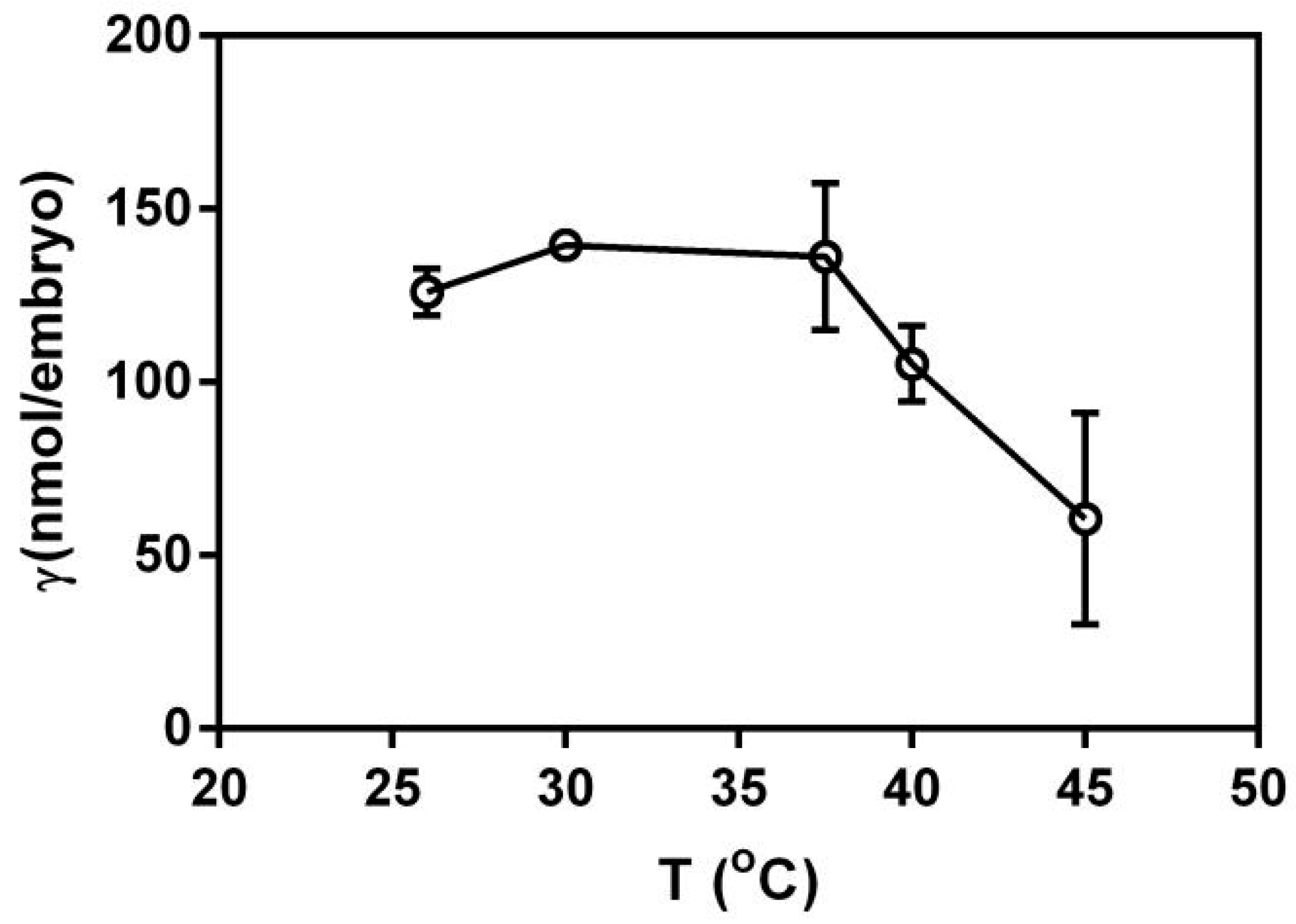
As shown in Figure 3, the value of γ (the binding ratio) shows an increasing trend with the increase of temperature. These results indicate that a higher temperature was conducive to the adsorption of embryos to PFLX. This might be due to the fact that the higher temperature could accelerate the transfer of embryonic membrane proteins and the rotation of membrane phospholipids, making more PFLX transport into the intramembrane matrix. But when the temperature is higher than 37.5 °C, the value of γ begins to decrease. This is because the normal physiological activity of the embryo is affected at high temperatures, which is not conducive to its adsorption of PFLX. The biological activity of the embryonic membrane decreases rapidly when the temperature is over 35 °C and the membrane protein transport function is seriously affected.
3.4. SEM Analysis
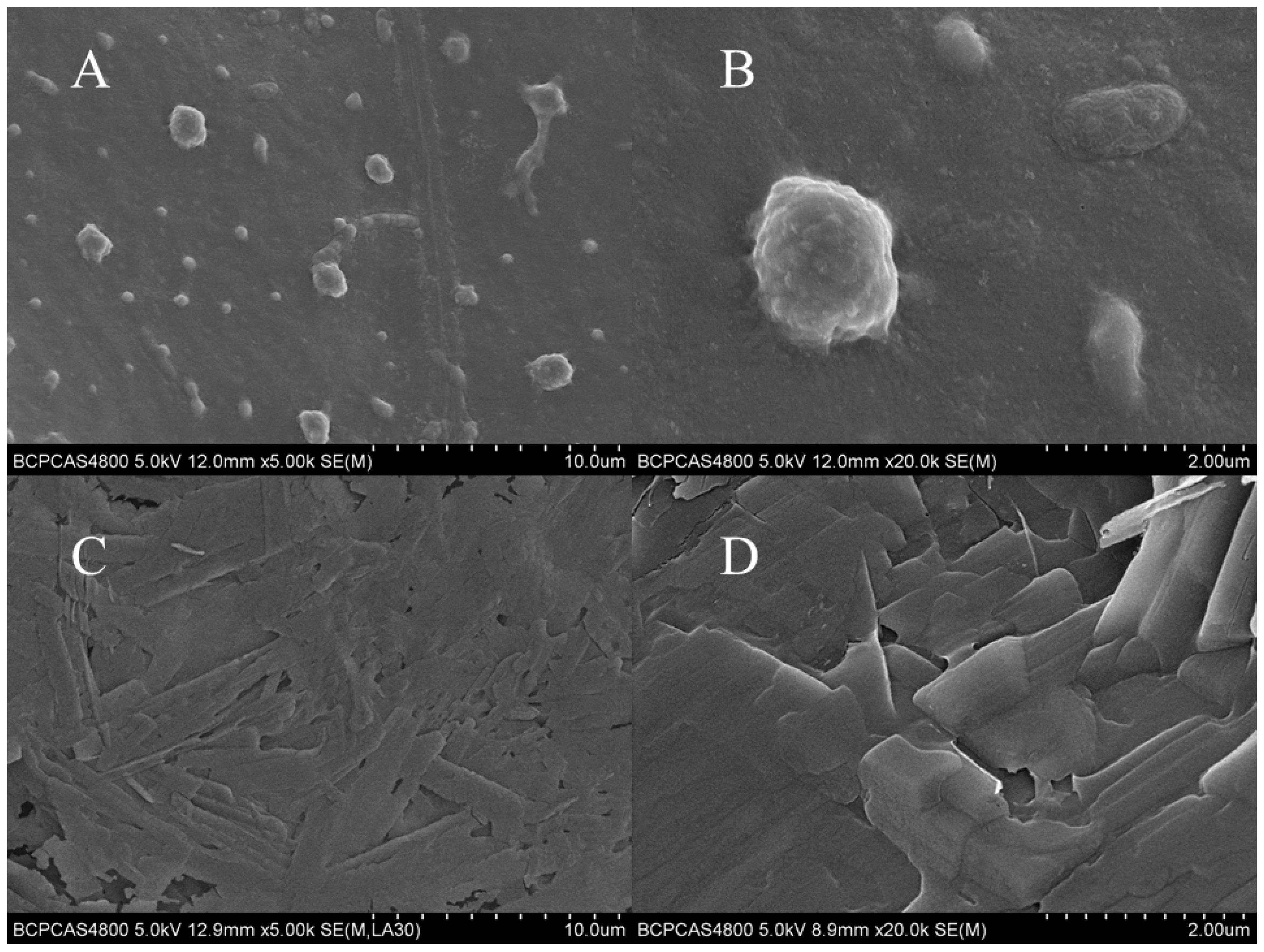
Figure 4 shows the SEM images of the embryonic membranes of the embryos exposed to blank control group and PFLX solution for 8 h. The surface characteristics of the embryonic membrane of the blank control group is shown in Figure 4A,B. After PFLX exposure, it caused obvious morphological changes on the embryonic membrane surface (Figure 4C,D). The structure of the embryonic membrane became thinner and the particles on the surface of the embryonic membrane became flakes. This is because PFLX enriched on the surface of the embryonic membrane affects the fluidity of membrane phospholipids, the transport function of membrane proteins, the absorption of nutrients and the discharge of metabolites, which further changes the surface structure of embryo membrane.
3.5. Effects of PFLX Exposure on Embryonic Autonomic Movement and Hatchability
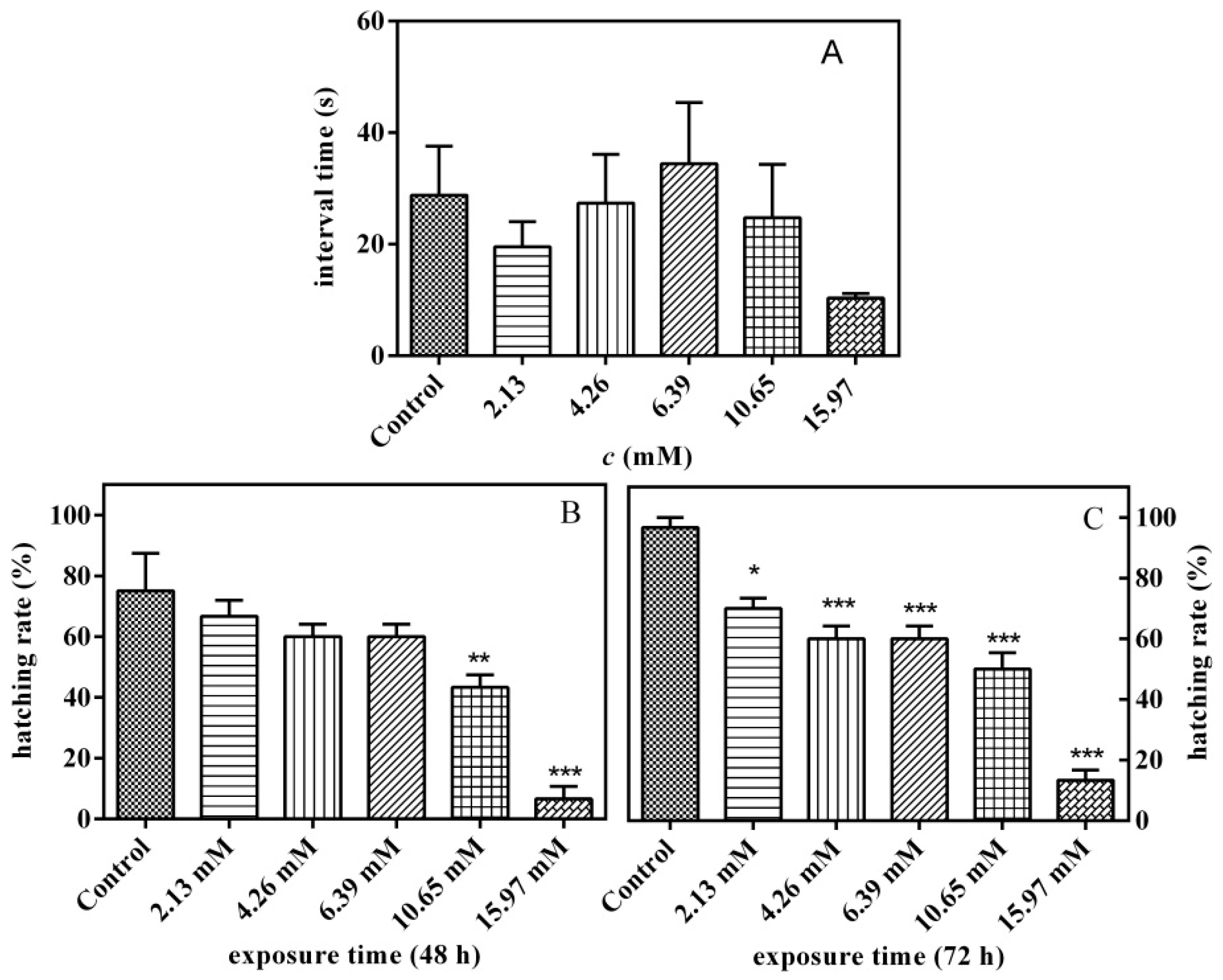
Figure 5A shows the effect of PFLX exposure for 24 h on the time interval between two consecutive voluntary movements of zebrafish embryos. The spontaneous movement of embryos in the chorion was promoted under low concentration (<4.26 mM) and was inhibited under medium concentration (6.39 mM), and when the concentration was further increased, it turned into a promoting effect. However, whether the PFXC exposure process has a promotion effect or an inhibition effect, the impact is small, and there is no significant difference compared with the control group (p > 0.05).
The effect of PFLX exposure on embryo hatchability was investigated. After 48 h of exposure, embryos in the blank control group and all concentration groups began to hatch (Figure 5B). The PFXC exposure groups all have inhibitory effects on hatching and show a typical dose–effect relationship. When the PFXC concentration increases to 10.65 mM, the embryo hatching rate is 43.2%, which is significantly lower than that of the blank control group (75.0%) (p < 0.01). After 72 h of exposure, most of the embryos had hatched out of the membrane, and all exposed groups of PFXC had a significant inhibitory effect on embryo hatchability (p < 0.05) (Figure 5C). The hatching rate was 60% under 4.26 mM PFLX exposure, which is much lower than that of the blank control group (96.7%) (p < 0.001). Therefore, the hatching rate decreases with the increase of PFLX concentration. When the concentration of PFLX increased to 15.97 mM, the hatching rate was only 13.36%.
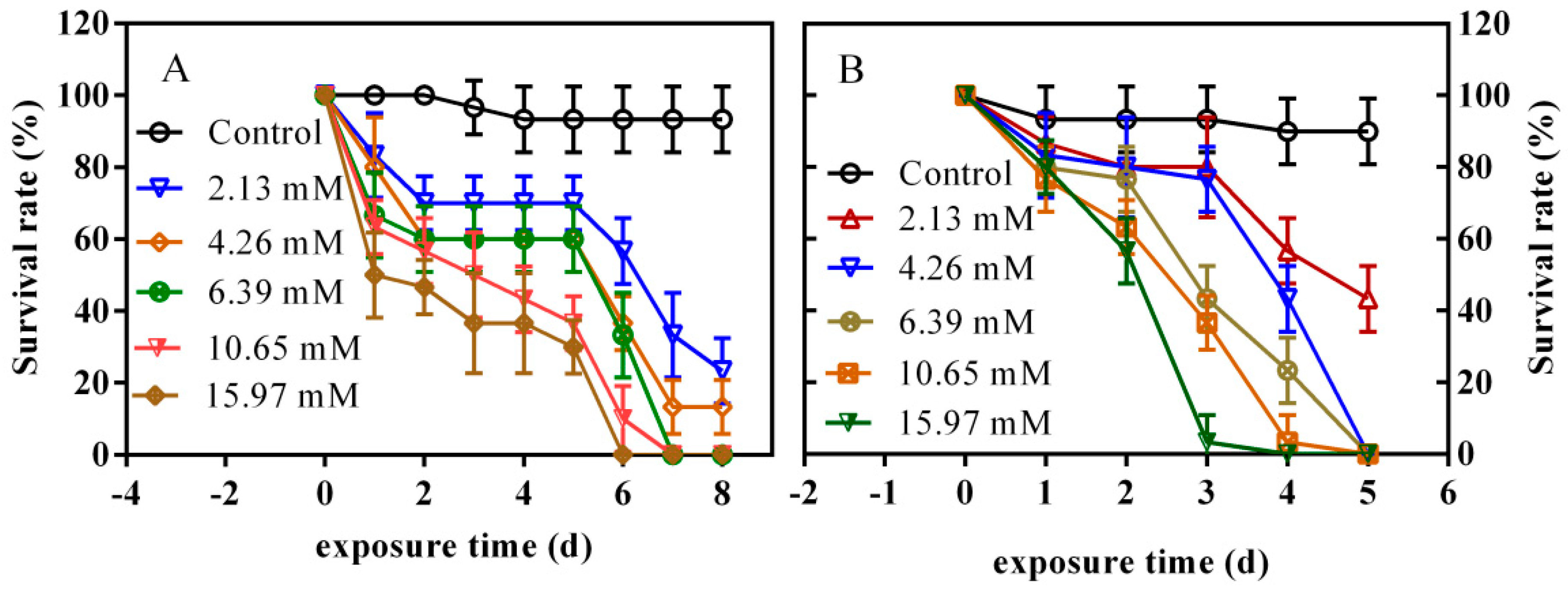
3.6. Effects of PFLX Exposure on the Survival Rate of Zebrafish Embryos and Larvae
Figure 6A shows the effect of PFLX exposure on the survival rate of zebrafish. After 1 day PFLX exposure, the survival rate of zebrafish decreased with the increase of PFLX concentration. The survival rate of the lowest concentration group (2.13 mM) was 83.3%, which is 50% for the highest concentration group (15.97 mM). The survival rate of 4.26 mM PFLX exposure group already shows a significant difference (p < 0.05) compared with the blank control group. PFLX exhibits obvious acute lethal toxicity for zebrafish embryos. After 2 days PFLX exposure, the survival rate of all exposure group further decreased, and all surviving embryos basically hatched into larvae. After 2–5 days of PFLX exposure, the survival rate of two high-concentration (10.65 mM and 15.97 mM) exposure groups continued to decline. The lower concentration exposure groups showed no obvious changes in survival rate. A possible reason for this phenomenon is that the zebrafish larvae gradually adapt to the environment of the lower concentration exposure group. However, when the exposure time reaches 6 days, the survival rate of zebrafish larvae decreased rapidly, manifesting as a sudden outbreak of death. The survival rate of the lowest concentration group (2.13 mM) declined to 56.7%, and all zebrafish larvae of the highest concentration group (15.97 mM) were dead. After 7 days exposure, no zebrafish larvae of 6.39 mM and 10.65 mM concentration groups were alive. After 8 days exposure, the survival rate of blank control group was 93.3%, while the 2.13 mM and 4.26 mM concentration groups were 23.3% and 13.4%, respectively. There is a clear dose–effect relationship between survival rate and exposure concentration.
After studying the effect of PFLX exposure on embryo survival, the effect of PFLX exposure on larvae was further studied, and the results are shown in Figure 6B. After 1 day exposure, the survival rate of all the concentration groups showed no significant difference with the blank control group, which only showed a slight decline. When the exposure time increased to 2 days, the survival rate of the highest concentration group (15.97 mM) declined to 56.7%, which shows a significant difference (p < 0.001) with that of the blank control group (93.3%). With the increase in the exposure time, the survival rate of each concentration group got lower and lower. After 4 days exposure, the survival rate of all concentration group showed a significant difference (p < 0.001) with that of the blank control group. When the exposure time reached 5 days, only the lowest concentration group (2.13 mM) had 43.3% of larvae alive, and all of the other concentration groups have no alive larvae. The effect of PFLX exposure on zebrafish larvae is similar to that of the zebrafish embryo, which shows acute lethal at high concentration and sublethal at low concentration.
Lethal Concentration 50 (LC50) is the concentration of the agent that killed 50% of the exposed individuals. It is an important parameter to measure the toxicity of pollutants present in water to aquatic animals, which is closely related to the exposure time. The LC50 and 95% confidence limit of the zebrafish embryos and larvae after 4 days PFLX exposure were calculated. The LC50 of embryo and larvae were 8.45 and 5.73 mM, respectively, and the 95% confidence limit were 6.13–11.64 and 4.90–6.69 mM, respectively. The lower LC50 of larvae means larvae are more sensitive to PFLX exposure than embryos. This is because embryo is protected by chorion, while larvae are more likely to be attacked by PFLX without this protection and show a higher lethal rate.
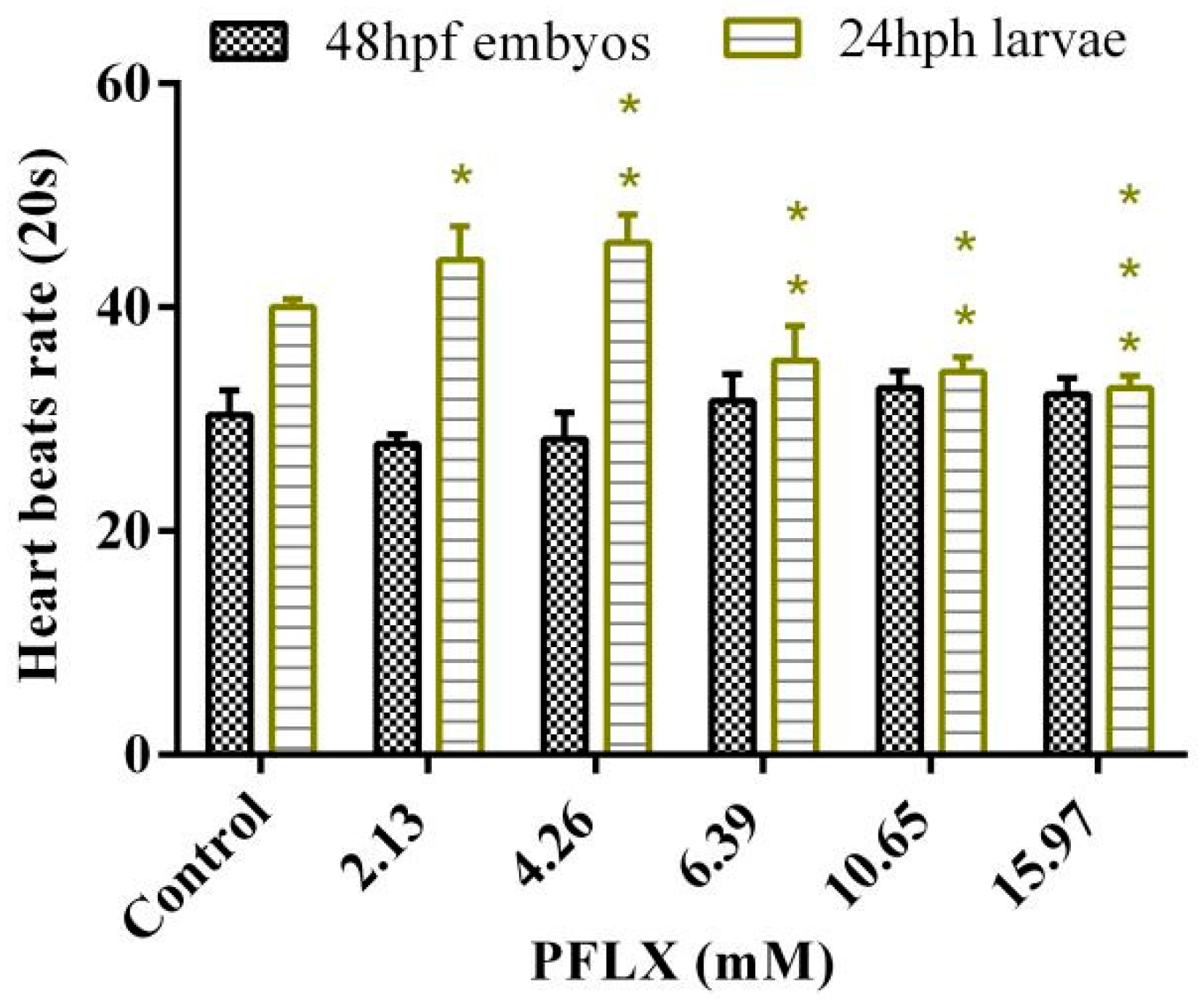
3.7. Effects of PFLX Exposure on the Heart Rate of Zebrafish Embryo and Larva
As shown in Figure 7, the effect of PFLX exposure on the heart rate of the 48 hpf embryo shows an inhibitory effect at lower concentrations (2.13 mM and 4.66 mM), and the inhibitory effect gradually declines with the increase of exposure concentration. When the exposure concentration increases to 6.39 mM, it turns out to have a promoting effect for the heart rate, which slightly increases with the increase of exposure concentration. However, no matter whether it is an inhibitory effect or a promoting effect on embryo heart rate, the effect is small and shows no significant difference (p > 0.05) compared with the blank control group.
The same as the effect of PFLX exposure on the zebrafish survival rate, the heart rates of larvae are more sensitive to PFLX exposure (Figure 7). However, unlike the effect of PFLX exposure on the heart rate of embryos, PFLX exposure shows low concentration promotion and high concentration inhibition on larval heart rate, which both increase with the increase of PFLX concentration. Even the heart rate of the lowest concentration group (2.13 mM) is significantly higher (p < 0.05) than that of the blank control group. When the exposure concentration increases to 4.26 mM, the promotion effect also enhances, and the difference from the blank control group is even more significant (p < 0.01). When larvae are exposed to PFLX, the heart’s ability to transport nutrients and oxygen is significantly reduced. In order to maintain their energy demand, it is necessary to increase the number of heartbeats to make up for the lack of nutrients and oxygen delivered by each heartbeat. When PFLX exposure concentration reaches 6.39 mM, it shows an inhibitory effect for the heart rate, which increases with the increase of PFLX exposure concentration.
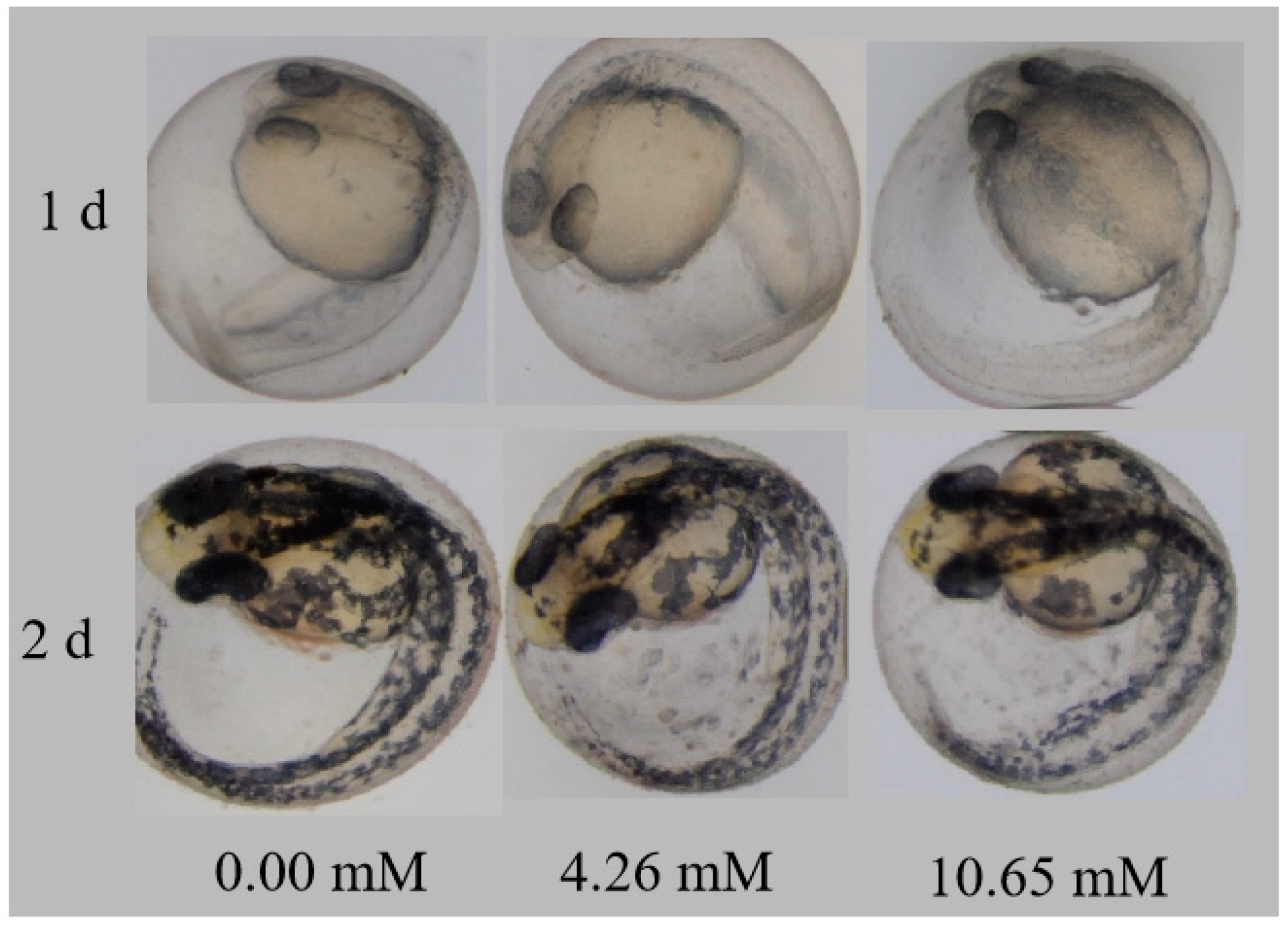
3.8. Teratogenic Effect of PFLX on Zebrafish Embryos and Larvae
The teratogenic effects of PFLX exposure on an embryo are shown in Figure 8. Compared with the blank control group, the embryos in each concentration group are basically normal, with no obvious abnormalities. This is because PFLX is adsorbed on the surface of the embryonic membrane and fails to enter the embryo. The presence of the embryonic membrane protects the early development of the embryo. As indicated in the previous section, PFLX exposure affects the embryo’s heart rate, but it shows no influence of abnormality. This indicates that impaired cardiovascular function caused by PFLX exposure may not have changes in cardiovascular morphology for the zebrafish embryo.
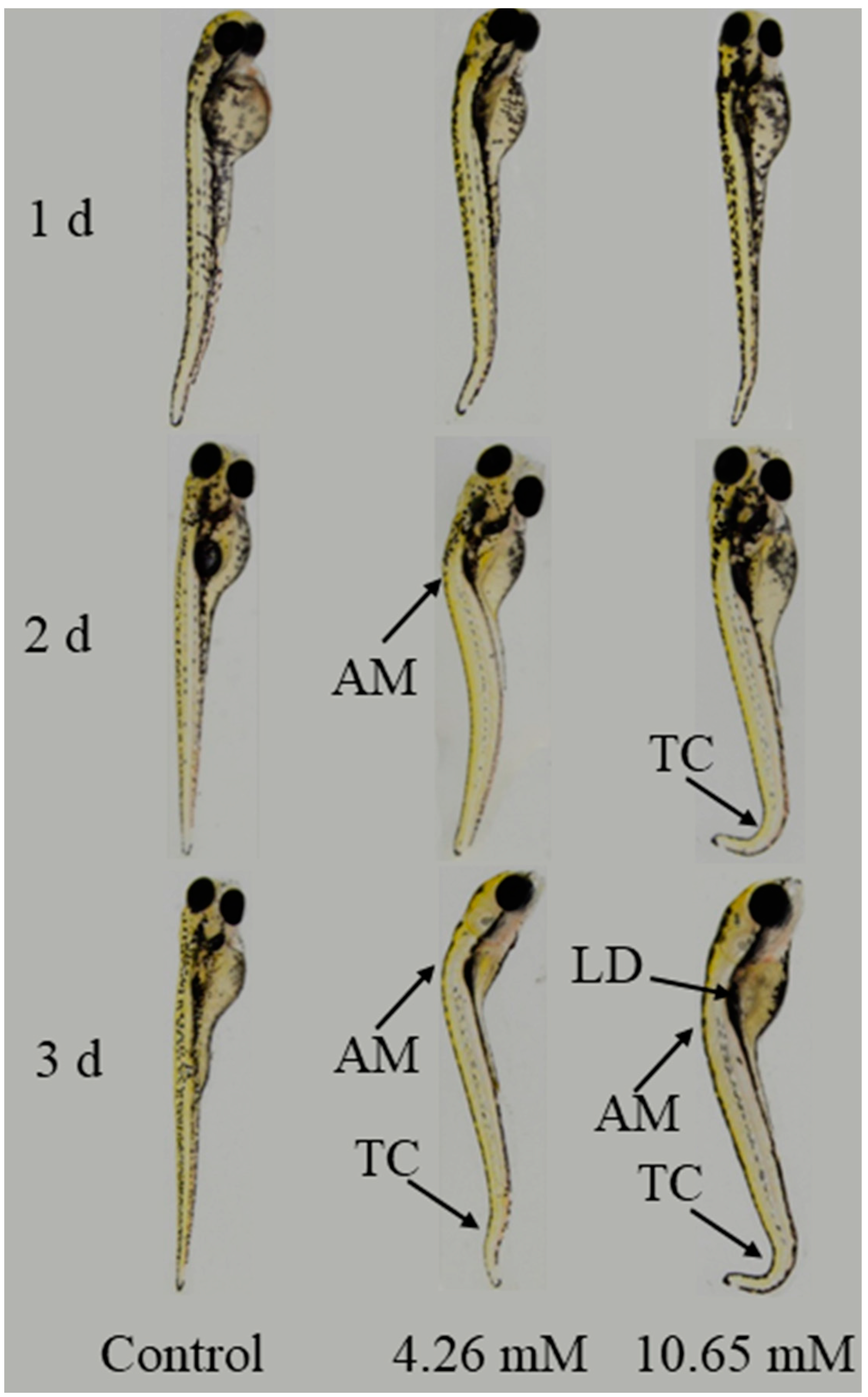
As shown in Figure 9, the teratogenic effect of PFLX on zebrafish larvae was studied. The abnormality characteristics become more and more obvious with the increase in exposure time. The development of larvae exposed to 4.26 mM and 10.65 mM PFLX are basically normal for 1 day exposure. After 2 days exposure, the larvae show axial malformation (AM) and tail curving (TC). When the exposure time reaches 3 days, the symptoms of AM and TC continue and they show a new symptom as liver degeneration (LD). PFLX exposure has a time–effect relationship with the teratogenic toxicity of larvae.
3.9. Correlation Analysis of Membrane Effect and Toxic Effect of PFLX
During PFLX exposure, PFLX is mainly adsorbed on the embryo membrane, which is in accordance with the Langmuir isotherm adsorption model. The value of N is 3.7 μmol/embryo, while that of K is 4.00 × 102 L/μmol. PFLX exposure makes the embryonic membrane structure thin and the membrane surface is in the shape of strip. The adsorbed PFLX affects the rotation of embryonic membrane phospholipids and the transport of membrane proteins, which affects the absorption of nutrients and the excretion of metabolic waste during the early development of the embryo, but it does not cause teratogenic toxicity. While the larvae without the protection of an embryonic membrane are directly exposed to the PFLX solution and show abnormal symptoms such as AM, TC and LD. As the exposure time increases, the above abnormal symptoms become more and more serious.
4. Conclusions
The transmembrane process of PFLX is divided into two stages: rapid diffusion and slow transport. The adsorption between PFLX and the embryonic membrane conforms to the Langmuir isotherm adsorption model. High temperature will reduce the biological activity of the embryonic membrane, thereby reducing the adsorption of PFLX. The concentration of PFLX has basically no effect on the autonomous movement of the embryo, but it will significantly affect the embryo hatching rate. In addition, PFLX exposure shows a clear dose–effect relationship on the survival rate of embryos and larvae. By comparing the LC50 values, it was found that larvae is more sensitive to PFLX exposure than embryos, which can be attributed to the effect of embryo chorionic. PFLX exposure has no teratogenic toxicity to embryos because the amount of PFLX entering the embryonic membrane is too small to induce morphological abnormalities. While PFLX exposure has no significant effect on embryo heart rate (p > 0.05), it does have a certain impact on the heart rate of larval. However, PFLX exposure does not induce any symptoms of cardiovascular malformations for larvae, which indicates that the function of the cardiovascular system may be damaged even without any cardiovascular malformations. This is useful for the toxicity study of PFLX exposure to the zebrafish cardiovascular system.
Author Contributions
Conceptualization and methodology, R.S.; data analysis, Y.Y.; formal analysis and validation, Q.L.; writing—original draft preparation, Y.Y. and R.W.; writing—review, editing and revision, R.L., H.F. and R.Y.; funding acquisition, L.J., R.S. and Y.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Key Science and Technology Programs of Beijing Polytechnic (2020Z004-KXZ and 2022X002-KXZ), Special Research Fund of Beijing Polytechnic (SG030801) and Beijing Outstanding Talents Training Fund (Top Natural Science) (2020Z002-002-KWT).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the support from Beijing Polytechnic.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, H.; Liu, F.-F.; Wang, S.-C.; Huang, T.-Y.; Li, M.-R.; Zhu, Z.-L.; Liu, G.-Z. Sorption of fluoroquinolones to nanoplastics as affected by surface functionalization and solution chemistry. Environ. Pollut. 2020, 262, 114347. [Google Scholar] [CrossRef] [PubMed]
- Von Ah, S.; Stephan, R.; Zurfluh, K.; Sidler, X.; Kümmerlen, D. Occurrence of quinolone-resistant Escherichia coli in environmental samples from a sow pool system in Switzerland. Schweiz. Arch. Tierh. 2019, 161, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Kindle, P.; Zurfluh, K.; Nüesch-Inderbinen, M.; Von Ah, S.; Sidler, X.; Stephan, R.; Kümmerlen, D. Phenotypic and genotypic characteristics of Escherichia coli with non-susceptibility to quinolones isolated from environmental samples on pig farms. Porc. Health Manag. 2019, 5, 9. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Song, W.; Lin, H.; Wang, W.; Du, L.; Xing, W. Antibiotics and antibiotic resistance genes in global lakes: A review and meta-analysis. Environ. Int. 2018, 116, 60–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Shi, W.; Liu, W.; Li, H.; Zhang, W.; Hu, J.; Ke, Y.; Sun, W.; Ni, J. A duodecennial national synthesis of antibiotics in China’s major rivers and seas (2005–2016). Sci. Total Environ. 2018, 615, 906–917. [Google Scholar] [CrossRef]
- Du, J.; Zhao, H.; Wang, Y.; Xie, H.; Zhu, M.; Chen, J. Presence and environmental risk assessment of selected antibiotics in coastal water adjacent to mariculture areas in the Bohai Sea. Ecotoxicol. Environ. Saf. 2019, 177, 117–123. [Google Scholar] [CrossRef]
- Dorival-García, N.; Zafra-Gómez, A.; Cantarero, S.; Navalón, A.; Vílchez, J. Simultaneous determination of 13 quinolone antibiotic derivatives in wastewater samples using solid-phase extraction and ultra performance liquid chromatography–tandem mass spectrometry. Microchem. J. 2013, 106, 323–333. [Google Scholar] [CrossRef]
- Teglia, C.M.; Perez, F.A.; Michlig, N.; Repetti, M.R.; Goicoechea, H.C.; Culzoni, M.J. Occurrence, Distribution, and Ecological Risk of Fluoroquinolones in Rivers and Wastewaters. Environ. Toxicol. Chem. 2019, 38, 2305–2313. [Google Scholar] [CrossRef]
- Wang, Z.; Du, Y.; Yang, C.; Liu, X.; Zhang, J.; Li, E.; Zhang, Q.; Wang, X. Occurrence and ecological hazard assessment of selected antibiotics in the surface waters in and around Lake Honghu, China. Sci. Total Environ. 2017, 609, 1423–1432. [Google Scholar] [CrossRef]
- Rosendahl, I.; Siemens, J.; Kindler, R.; Groeneweg, J.; Zimmermann, J.; Czerwinski, S.; Lamshöft, M.; Laabs, V.; Wilke, B.-M.; Vereecken, H.; et al. Persistence of the Fluoroquinolone Antibiotic Difloxacin in Soil and Lacking Effects on Nitrogen Turnover. J. Environ. Qual. 2012, 41, 1275–1283. [Google Scholar] [CrossRef]
- Gatti, M.; Bianchin, M.; Raschi, E.; De Ponti, F. Assessing the association between fluoroquinolones and emerging adverse drug reactions raised by regulatory agencies: An umbrella review. Eur. J. Intern. Med. 2020, 75, 60–70. [Google Scholar] [CrossRef]
- Wang, D.; Ning, Q.; Dong, J.; Brooks, B.W.; You, J. Predicting mixture toxicity and antibiotic resistance of fluoroquinolones and their photodegradation products in Escherichia coli. Environ. Pollut. 2020, 262, 114275. [Google Scholar] [CrossRef] [PubMed]
- Thompson, A.M. Ocular toxicity of fluoroquinolones. Clin. Exp. Ophthalmol. 2007, 35, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Luo, T.; Chen, J.; Li, X.; Zhang, S.; Yao, H.; Peijnenburg, W.J. Effects of lomefloxacin on survival, growth and reproduction of Daphnia magna under simulated sunlight radiation. Ecotoxicol. Environ. Saf. 2018, 166, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Luo, Y.; Wu, L.; Huang, Y.; Christie, P. Residues and potential ecological risks of veterinary antibiotics in manures and composts associated with protected vegetable farming. Environ. Sci. Pollut. Res. 2015, 22, 5908–5918. [Google Scholar] [CrossRef]
- Lu, T.; Zhu, Y.; Ke, M.; Peijnenburg, W.; Zhang, M.; Wang, T.; Chen, J.; Qian, H. Evaluation of the taxonomic and functional variation of freshwater plankton communities induced by trace amounts of the antibiotic ciprofloxacin. Environ. Int. 2019, 126, 268–278. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, X.; Yin, X.; Shi, M.; Dahlgren, R.A.; Wang, H. Toxicity Assessment of Combined Fluoroquinolone and Tetracycline Exposure in Zebrafish (Danio rerio). Environ. Toxicol. 2016, 31, 736–750. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Yu, Y.; Lan, R.; Yu, R.; Yuan, Z.; Xia, Z. The cardiovascular toxicity induced by high doses of gatifloxacin and ciprofloxacin in zebrafish. Environ. Pollut. 2019, 254, 112861. [Google Scholar] [CrossRef]
- Peltzer, P.M.; Lajmanovich, R.C.; Attademo, A.M.; Junges, C.M.; Teglia, C.M.; Martinuzzi, C.; Curi, L.; Culzoni, M.J.; Goicoechea, H.C. Ecotoxicity of veterinary enrofloxacin and ciprofloxacin antibiotics on anuran amphibian larvae. Environ. Toxicol. Pharmacol. 2017, 51, 114–123. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, X.; He, Q.; Song, Q.; Wu, F.; Zhang, C. Solvothermal synthesis of InNbO4 cubes for efficient degradation of pefloxacin. Spectrochim. Acta A 2020, 234, 118247. [Google Scholar] [CrossRef]
- Lee, H.-J.; Kim, K.Y.; Hamm, S.-Y.; Kim, M.; Kim, H.K.; Oh, J.-E. Occurrence and distribution of pharmaceutical and personal care products, artificial sweeteners, and pesticides in groundwater from an agricultural area in Korea. Sci. Total Environ. 2019, 659, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, L.; Liu, X.; Ding, J. Ranking and prioritizing pharmaceuticals in the aquatic environment of China. Sci. Total Environ. 2019, 658, 333–342. [Google Scholar] [CrossRef] [PubMed]
- Sha, L.; Tang, X.; Liu, D.; Xu, Y.; Ding, Y.; Ding, F. Detection and Quantitation of Lomefloxacin and Pefloxacin Residues in the Organ Tissues and Eggs of Laying Hens. J. Food Prot. 2018, 81, 810–814. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; He, K.; Zhang, D.; Huang, Z. Production and characterization of a monoclonal antibody for Pefloxacin and mechanism study of antibody recognition. Biosci. Biotechnol. Biochem. 2019, 83, 633–640. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, F.; Minhaz, A.; Shah, M.R.; Ain, N.U.; Khan, A.A.; Shah, K.; Ullah, S.; Ishaq, M. Highly selective and sensitive chemosensor for detection of Pefloxacin in tap water based on click generated triazole. Microchem. J. 2019, 146, 332–338. [Google Scholar] [CrossRef]
- Qu, S.; Wei, J.; Wang, Q.; Li, Y.; Jin, X.; Chaib, L. Oral toxicity of pefloxacin, norfloxacin, ofloxacin and ciprofloxacin: Comparison of biomechanical and histopathological effects on Achilles tendon in rats. J. Toxicol. Sci. 2011, 36, 339–345. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; van Mil, H.G.; Richardson, M.K. Large-scale assessment of the zebrafish embryo as a possible predictive model in toxicity testing. PLoS ONE 2011, 6, e21076. [Google Scholar] [CrossRef] [Green Version]
- Di Paolo, C.; Seiler, T.-B.; Keiter, S.; Hu, M.; Muz, M.; Brack, W.; Hollert, H. The value of zebrafish as an integrative model in effect-directed analysis—A review. Environ. Sci. Eur. 2015, 27, 8. [Google Scholar] [CrossRef] [Green Version]
- Marques, I.J.; Lupi, E.; Mercader, N. Model systems for regeneration: Zebrafish. Development 2019, 146, dev167692. [Google Scholar] [CrossRef] [Green Version]
- Shao, Y.; Xiao, H.; Di Paolo, C.; Deutschmann, B.; Brack, W.; Hollert, H.; Seiler, T.B. Integrated zebrafish-based tests as an investigation strategy for water quality assessment. Water Res. 2019, 150, 252–260. [Google Scholar] [CrossRef]
- Babić, S.; Barišić, J.; Stipaničev, D.; Repec, S.; Lovrić, M.; Malev, O.; Martinovic-Weigelt, D.; Čož-Rakovac, R.; Klobučar, G. Assessment of river sediment toxicity: Combining empirical zebrafish embryotoxicity testing with in silico toxicity characterization. Sci. Total Environ. 2018, 643, 435–450. [Google Scholar] [CrossRef] [PubMed]
- Stainier, D.Y. Zebrafish genetics and vertebrate heart formation. Nat. Rev. Genet. 2001, 2, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Song, C.; Gao, N.Y.; Gao, H.W. Transmembrane distribution of kanamycin and chloramphenicol: Insights into the cytotoxicity of antibacterial drugs. Mol. Biosyst. 2010, 6, 1901–1910. [Google Scholar] [CrossRef]
- Yu, Y.; Hu, Z.; Wang, Y.; Gao, H. Magnetic SN-functionalized diatomite for effective removals of phenols. Int. J. Miner. Process. 2017, 162, 1–5. [Google Scholar] [CrossRef]
- Xiao, C.; Han, Y.; Liu, Y.; Zhang, J.; Hu, C.-Q. Relationship Between Fluoroquinolone Structure and Neurotoxicity Revealed by Zebrafish Neurobehavior. Chem. Res. Toxicol. 2018, 31, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Asadipour, A.; Moshafi, M.H.; Khosravani, L.; Moghimi, S.; Amou, E.; Firoozpour, L.; Ilbeigi, G.; Beiki, K.; Soleimani, E.; Foroumadi, A. N-substituted piperazinyl sarafloxacin derivatives: Synthesis and in vitro antibacterial evaluation. DARU 2018, 26, 199–207. [Google Scholar] [CrossRef]
- Zhang, Z.; Xie, X.; Yu, Z.; Cheng, H. Influence of chemical speciation on photochemical transformation of three fluoroquinolones (FQs) in water: Kinetics, mechanism, and toxicity of photolysis products. Water Res. 2019, 148, 19–29. [Google Scholar] [CrossRef]
Figure 1.
Adsorption kinetic (A) and diffusion control kinetic model (B) of PFLX (1.00 mM) on embryos (n = 5) after incubation at different times at 25 °C, a: diffusion stage; b: transfer stage.
Figure 1.
Adsorption kinetic (A) and diffusion control kinetic model (B) of PFLX (1.00 mM) on embryos (n = 5) after incubation at different times at 25 °C, a: diffusion stage; b: transfer stage.

Figure 2.
(A): The change of γ binding on zebrafish embryos (n = 5) after the embryos had been incubated with PFLX for 8 h; (B): Plot of 1/γ vs. 1/cL.
Figure 2.
(A): The change of γ binding on zebrafish embryos (n = 5) after the embryos had been incubated with PFLX for 8 h; (B): Plot of 1/γ vs. 1/cL.

Figure 3.
Effects of temperature on g of PFLX (1.00 mM) on embryos (n = 5).

Figure 4.
SEM images of the outer membrane surface of the zebrafish embryo (n = 5) after incubation for 8 h. (A,B): Blank control in the reconstituted medium. (C,D): 1.0 mM PFLX.
Figure 4.
SEM images of the outer membrane surface of the zebrafish embryo (n = 5) after incubation for 8 h. (A,B): Blank control in the reconstituted medium. (C,D): 1.0 mM PFLX.

Figure 5.
Effects of PFLX on the interval time (s) between two connected spontaneous movements of the embryos in the chorion for 24 h exposure. (A) Effects of PFLX exposure on hatching of embryos, (B): 48 h, (C): 72 h. The asterisk symbols (*) denote no significantly different results compared to control group, * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 5.
Effects of PFLX on the interval time (s) between two connected spontaneous movements of the embryos in the chorion for 24 h exposure. (A) Effects of PFLX exposure on hatching of embryos, (B): 48 h, (C): 72 h. The asterisk symbols (*) denote no significantly different results compared to control group, * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 6.
Effect of PFLX exposure on the survival rate of the zebrafish embryo (A) and larvae (B).

Figure 7.
Effects of PFLX on the heart beats rate of embryos and larvae: The asterisk symbols (*) denote significantly different results compared to control at * p < 0.05, ** p < 0.01, *** p < 0.001.
Figure 7.
Effects of PFLX on the heart beats rate of embryos and larvae: The asterisk symbols (*) denote significantly different results compared to control at * p < 0.05, ** p < 0.01, *** p < 0.001.

Figure 8.
Teratogenic effect of PFLX on a zebrafish embryo.

Figure 9.
Teratogenic effect of PFLX on a zebrafish larva.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shen, R.; Yu, Y.; Liu, Q.; Wang, R.; Lan, R.; Feng, H.; Yu, R.; Jin, L.; Li, Y. Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio). Water 2022, 14, 3857. https://doi.org/10.3390/w14233857
AMA Style
Shen R, Yu Y, Liu Q, Wang R, Lan R, Feng H, Yu R, Jin L, Li Y. Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio). Water. 2022; 14(23):3857. https://doi.org/10.3390/w14233857
Chicago/Turabian StyleShen, Rong, Yichang Yu, Qinfang Liu, Ruili Wang, Rong Lan, Hui Feng, Ran Yu, Lihua Jin, and Ye Li. 2022. "Toxic Effects and Mechanism of Pefloxacin on the Early Life Stage of Zebrafish (Danio rerio)" Water 14, no. 23: 3857. https://doi.org/10.3390/w14233857
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.