

Effect of Using Trichoderma spp. on Turfgrass Quality under Different Levels of Salinity

1
Department of Horticulture and Crop Sciences, School of Agriculture, University of Jordan, Amman 11942, Jordan
2
Department of Plant Pathology, School of Agriculture, University of Jordan, Amman 11942, Jordan
*
Author to whom correspondence should be addressed.
Water 2022, 14(23), 3943; https://doi.org/10.3390/w14233943
Submission received: 11 November 2022
/
Revised: 27 November 2022
/
Accepted: 28 November 2022
/
Published: 4 December 2022
(This article belongs to the Special Issue Desalination Treatment of Irrigation Water)
Abstract
:Lawns achieve environmental, functional, and aesthetical roles in urban environments. The objectives of this research were to assess the effect of different salinity levels on Trichoderma isolates and to study the effect of Trichoderma spp. on perennial ryegrass under different levels of salinity. T. harzianum (ThLem2017-01) and T. atroviride (TaDP2019-01) isolates had a higher mycelium growth rate than T. atroviride (TaDP2019-02) when salinity levels were low. In contrast, the mycelium growth rate of T. atroviride (TaDP2019-02) isolate at high salinity levels had superior results. Turfgrass seeds that were inoculated with (TaDP2019-02) isolate maintained high radicle length, coleoptile length, and leaf length under high salinity levels. Increasing salinity level decreased clippings’ fresh weight (FW), dry weight (DW), and shoot and root dry weight of perennial ryegrass. Interestingly, perennial ryegrass pots that were treated with (TaDP2019-02) isolate had increased FW and DW by 16 to 114% and 24 to 76%, respectively. Soils that were inoculated with Trichoderma (TaDP2019-02) had higher CO2 respiration (75%) than the control. Therefore, using T. atroviride (TaDP2019-02) isolate revealed promising results in increasing plant biomass and as an environmentally friendly alternative factor to overcome salinity stress.
1. Introduction
Turfgrasses are one of the main components of the landscape [1]. Turfgrasses play an essential role in urban environments by reducing soil erosion, filtering air and water and consequently reducing pollution [2,3]. In addition, lawns add an aesthetic value to the landscape and increase a property’s value [4].
Turfgrasses are used to enhance the space of beauty [5], recreation, and sport [1]. It is used as a playground because it gives a cushioning influence that decreases pains to players when compared to ordinary soils or artificial grass, particularly in contact sports such as rugby and football [3]. Turfgrass is considered as billion-dollar industry [6,7]. Perennial ryegrass (Lolium perenne L.) is one of the most common cool-season turfgrasses. It is native to temperate Asia, Europe, and North Africa [8]. It is used as a forage species with high yield and quality, and is appropriate for parks, home courtyards, cemeteries, golf courses, and roadsides [9]. Perennial ryegrass is often used for quickly re-establishing devastated lawns because it is distinguished by rapid seed germination, growth, and establishment [3]. Interestingly, perennial ryegrass has been used widely for clean-up of heavy metal-polluted soils (remediation) [10].
Turfgrass is affected by numerous abiotic and biotic stresses such as salinity, intensive fertilization schemes, heat, cold, drought, soil compaction, diseases, pests, and pesticide and herbicide application [11,12]. Soil salinity is consistently considered as one of the major problems around the globe. High salt levels in soil ECe values of ˂1.0–24.0 dS m−1 significantly decrease plant performance and quality [13]. However, seed germination and the seedling stage are the most sensitive phases to saline water [14]. In fact, at early growth stages (germination and seedling) salt stress can induce critical osmotic potential that inhibits the water uptake and mineral nutrients of the germinated seeds or/and increase assimilation of toxic ions (especially Na) that possibly alter hormonal or enzymatic activities of the plants [15,16].
Jordan is among the poorest countries in terms of water resources worldwide, facing serious problems related to water shortages [17]. High demand for irrigation water coupled with a significant rise in population recently due to refugees fleeing from the Syrian civil war increase the over-abstraction of groundwater (the main source of fresh water in Jordan) and consequently reduce water quality [18]. In fact, the continuous over-abstraction of water reduced the levels of groundwater and increased the salinity readings [19]. To overcome these problems, it is imperative to modulate some horticultural practices through decreasing the application of chemical fertilizers, proper leaching and drainage, and deep plowing [12,20].
Perennial ryegrass is classified as a cool-season turfgrass, with moderate irrigation requirements, good traffic tolerance, fine texture leaf type, and moderate tolerance to salinity [3,21]. The salinity thresholds of perennial ryegrass ranged from 4 to 8 dS m−1 [22]. This tolerance in perennial ryegrass is partially attributed to antioxidant genes expression efficiency as well as antioxidant enzymes activation [9]. Plant growth (shoot and root biomass, density) and aesthetic visual component (leaf greenness or color) are critical variables to assess turf grass responses to salt stress [23,24]. Growing perennial ryegrass in a saline environment (6 dS m−1) for six weeks significantly reduced shoot and root dry weight and visual quality [24].
Trichoderma spp. Fungi inoculation has been recommended as an environmentally friendly approach to overcome salinity problems [25]. These fungi are used to enhance nutrient uptake and increase plant growth, induce systemic resistance and as a biological control for many plant’s fungal pathogens and pests [26,27,28,29]. Trichoderma symbioses with wheat plants, which ameliorates the negative impact of salt stress by inducing the shoots to accumulate more protective amino acids [30]. In addition, Trichoderma spp. decreases the need for chemical fertilizers and pesticides. In this context, the positive interaction between fungi and plants can achieve the global trend toward sustainable agriculture, by promoting salinity tolerance and lowering agrichemical inputs [18,31]. Therefore, T. harzianum fungi inoculation can be a promising practice for lawn management in saline soils. However, the effect of salt stress on T. harzianum is not well investigated. In addition, no study we are aware of has discussed the influence of T. atroviride inoculation on perennial ryegrass growth and physiology under saline conditions. The objectives of this study were (1) to evaluate the effect of different salt stress levels on Trichoderma isolates growth and (2) to assess the influence Trichoderma spp. inoculation on plant growth and physiology of perennial ryegrass under saline conditions.
2. Materials and Methods
2.1. Experiments Layout
In this research, laboratory and greenhouse experiments were conducted to evaluate the effect of Trichoderma sp. on perennial ryegrass (Lolium perenne L.) under salinity stress. Two experiments were conducted in the laboratory while the third one was in the greenhouse. In the first experiment, the mycelium growth rate of three Trichoderma isolates (Table 1) under different sodium chloride (NaCl) concentrations was assessed. In the second experiment, the performance of perennial ryegrass seeds inoculated with Trichoderma spp. isolates was assessed under saline conditions. In the third experiment, the best Trichoderma isolate from experiment two was used to inoculate perennial ryegrass and evaluated in the greenhouse under a saline environment.
2.2. Trichoderma spp. Isolates
Three Trichoderma isolates belonging to two species, isolated from two plants from the Jordan Valley, Jordan, were used in this study. Isolates TaDP2019-01 and TaDP2019-02 were isolated in 2019 from date palm fronds and identified as T. atroviride; isolate ThLem2017-01 was isolated in 2017 from lemon leaves and identified as T. harzianum. The isolates were molecularly identified by sequencing the internal transcribed spacer (ITS1/ITS4) [32]. Deoxyribonucleic acid (DNA) extraction, polymerase chain reaction (PCR) amplification, and sequencing were conducted at Macrogen Inc, Seoul, South Korea. Sequences were received as FASTA files (stands for “FAST-All”), edited via MEGA 7 software (Institute for Genomics and Evolutionary Medicine, Temple University, Philadelphia, PA, USA) [33] and BLASTn program (National Center for Biotechnology Information, Bethesda, MA, USA). Reference accessions with high similarity were used for phylogenetic analysis. Final sequences with their identity were submitted to the Genbank and accession numbers were received for ITS1/4 (MT626715, MT626716, and MT626718).
2.3. Experiment 1: Mycelial Growth Rate of the Three Trichoderma spp. Isolates under Different Salt Concentrations
Trichoderma spp. isolates (TaDP2019-01, TaDP2019-02, ThLem2017-01) (Table 1) were sub-cultured on potato dextrose agar (PDA) (TCHNO PHARMCHEM, Delhi, India) medium (39.3 g L−1 water (sterile-distilled water) amended with different NaCl concentrations (50, 100, 150, 200, 300, 400, 500, 750, and 1000 mM) (they are equivalent to 4, 8, 12, 18, 28, 38, 44, 64, and 84 dS m−1) by using sodium chloride extra pure (Scharlau), Sentmenat, Spain. Sterile-distilled water was used as a control. Media were sterilized by autoclaving at 121 °C and 15 psi then poured into 9 cm diameter Petri dishes under aseptic conditions [34].
One disc (2 mm in diameter) from 10-day-old cultures of each Trichoderma spp. was placed in the middle of the PDA Petri dishes. Plates were incubated in the growth chamber at 25 ± 5 °C and at 70% ± 5% relative humidity. Mycelium growth rate (mm day−1) readings were recorded after 3, 6, and 9 days. An average of two perpendicular measurements was recorded [35]. A completely randomized design (CRD) with three replicates per NaCl concentration was used.
2.4. Experiment 2: In Vitro Evaluation of Trichoderma spp. on Turfgrass Seeds under Different Levels of Salinity
Turfgrass seeds were cultured on instant agar agar medium (MERK KGaA, Darmstadt, Germany) (20 g L−1). The electrical conductivity (EC) of the medium was adjusted to three NaCl concentrations (50, 100, and 150 mM) and control (sterile-distilled water) by using sodium chloride extra pure (Scharlau), Sentmenat, Spain. Media was sterilized by autoclaving at 121 °C at 15 psi and poured in a 9 cm diameter Petri dishes under aseptic conditions.
Seeds were soaked in sterile-distilled water for 15 min, sterilized in 2.5% of commercial sodium hypochlorite solution (6%) for five minutes then rinsed three times in sterile-distilled water under aseptic conditions [36].
The spore suspension was prepared by mixing two plugs (5 mm) of 10-day-old isolates culture for each Trichoderma spp. (ThLem2017-01, TaDP2019-01, and TaDP2019-02) in 5 mL distilled water for 30 min. Spore count was adjusted to 8 × 106 spore mL−1. Seeds of turfgrass were soaked for 15 min in the spore suspension, dried in aseptic conditions [35], and plated on agar agar-medium amended with different NaCl concentrations. Ten seeds were immersed gently in each Petri dish and were incubated in the growth chamber at 25 ± 1 °C with 70% ± 5 relative humidity for 20 days. Four replicates were used for each NaCl concentration. Measurements including radicle length and coleoptile length, and the number of leaves was recorded. The Trichoderma isolate that showed best results in this assay was selected for the following experiment. Completely randomized design (CRD) with four replicates per NaCl concentration was considered. Therefore, there were 64 experimental units (10 seeds of turfgrass per each experimental unit). The readings were taken for all the seeds in each Petri dish so that 10 readings from each measurement were taken per experiment. The 10 values from each replicate were averaged to one value before statistical analysis.
2.5. Experiment 3: In-Vivo Effect of Trichoderma Isolate on Perennial Ryegrass under Different Levels of Salinity
Greenhouse experiments were conducted at the greenhouse facility, School of Agriculture, University of Jordan, Amman, Jordan (32°00′40.4″ N 35°52′20.7″ E from 27 October 2020 to 3 May 2021. The minimum and maximum average temperatures in the greenhouse were 10 °C and 25 °C, respectively.
Inoculum of Trichoderma isolate that had been selected from the second experiment was prepared on wheat as a carrier seed. Wheat seeds were rinsed and soaked in tap water for 12–24 h, loaded into sterilized mason jars and sterilized by autoclaving at 121 °C and 15 Psi. Two plugs (5 mm) of 10-day-old cultures of the selected Trichoderma isolate were added to the jars in aseptic conditions and incubated in the dark for 21 days. Jars were shaken at least once every day [37].
2.5.1. Perennial Ryegrass Cultivation and Treatments
Perennial ryegrass seeds were sown on 27 October 2020 by sowing the entire surface of the pot with approximately 100 seeds. Sterilized pots (1.4 L) were filled with sterilized, washed silica sand and were covered with 0.5 cm of wet peat moss. The growing pots are made of plastic material and each pot has five drainage holes. The soil surface area for each pot is 136 cm2 and the base area is 96 cm2. The top diameter of the pot is 13.17 cm and the base diameter is 11.06 cm. The height of each pot is 12.19 cm. A geotextile (white fabric) layer was placed in the bottom of each pot to prevent sand from passing through the pot’s holes. Pots were irrigated once every two days by pouring 500 mL of water using a beaker (500 mL) for each pot. After two weeks, pots were fertigated once a week for one month using water-soluble fertilizer 20N-20P2O5-20K2O plus trace elements. The final composition of the fertigation solution was 28.6 mmol L−1 nitrogen, 5.6 mmol L−1 phosphorus, 8.5 mmol L−1 potassium, 0.6 mmol L−1 magnesium, 0.05 mmol L−1 iron, 0.02 mmol L−1 boron, 0.04 mmol L−1 manganese, 0.03 mmol L−1 zinc, and 0.01 mmol L−1 copper pouring 500 mL of fertigated water using a beaker (500 mL) per pot. After 20 days, seven seeds of the selected Trichoderma sp. inoculum were added to each pot to inoculate the well-established perennial ryegrass plants.
After 20 days of treating pots with Trichoderma (TaDP2019-02) isolate (17 November 2020), four salinity treatments (levels) were considered (4 dS m−1 (2.9 g L−1), 8 dS m−1 (5.8 g L−1), 12 dS m−1 (8.8 g L−1), and tap water (0.30–0.90 dS m−1)) by pouring 500 mL of saline water using a beaker (500 mL) into each pot every two days.. In this experiment, pots were arranged in a completely randomized design (four levels of NaCl concentrations and two levels of Trichoderma) with five replicates.
2.5.2. Measurements
Leaves of turfgrass were mowed to a length of 7.0 cm. Clipped leaves from each pot were collected and weighed to derive the fresh weight (FW) then were oven-dried at 80 °C for 24 hr to record dry weight (DW). Dry weight was performed on a monthly basis to estimate crop production. Leaf color co-ordinates (L*, a*, and b*) were measured every two weeks by Chroma Meter measuring head (CHROMA METER CR-400, KONICA MINOLTA, Osaka, Japan). Electrical conductivity and pH of the leachate water were measured to estimate the water quality and how salt builds in soil over time. EC was measured using a pH/Conductivity Meter (Extech EC500 Waterproof EcStik П, Extech, Boston, MA, USA) and pH was measured using a pH meter (pp-203 pH meter, GOnDO Electronic Co, Taipei, Taiwan). EC and pH were recorded five times during the experiment. The pH meter was calibrated frequently between measurements by inserting the electrode in the standardizing solution buffers (first with pH 7, then with pH 4, and finally with pH 10) according to the manual protocol.
Soil respiration was determined at the end of experiment using the Solvita® kit (Solvita, CO2-Burst, Mount Vernon, ME, USA). Soil respiration is considered as an indicator of microbial activity. This test was made on two levels of salinity treatments (12 dS m−1 and control) with three replicates.
The presence or absence of the selected isolate was confirmed after six months of seeding by plating perennial ryegrass leaves and roots on PDA media. From each treatment of perennial ryegrass, five segments (5 mm) from roots and leaves were surface-sterilized in 1% hypochlorite for 3 min then rinsed five times by distilled water and plated on PDA supplemented with ampicillin. The plates were incubated at 28 °C for one week and the presence or absence of Trichoderma was determined microscopically and based on culture morphology [38].
2.6. Data Analysis
Data analyses were conducted using SAS software (Version 9.4 for Windows; SAS Institute, Cary, NC, USA). Two-way analysis of variance using general linear model PROC GLM were used for all experiments. The least significant difference test (LSD) (p ≤ 0.05) was used to identify differences between treatments [39].
3. Results
3.1. Mycelial Growth Rate of the Three Trichoderma spp. Isolates under Different Salt Concentrations
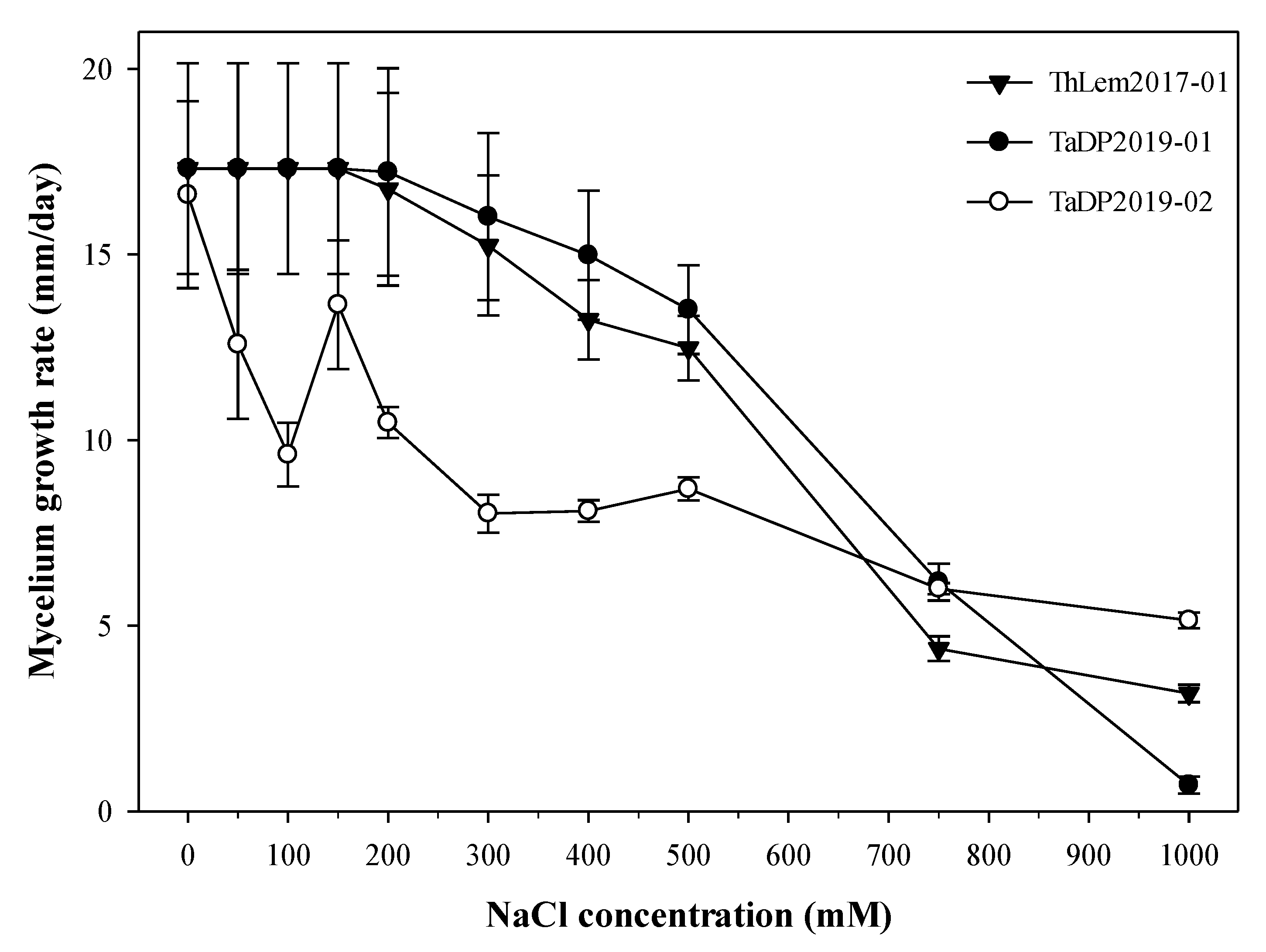
Salt stress significantly reduced mycelium growth rate of Trichoderma spp. (Figure 1). Salinity stress reduced mycelium growth rate of Trichoderma spp. by 13.8% at 200 mM, 32.4% at 500 mM, 67.7% at 750 mM and 82.4% at 1000 mM. ThLem2017-01 and TaDP2019-01 had a higher mycelium growth rate than TaDP2019-02 when salinity levels were 200, 300, 400, and 500 mM. Conversely, TaDP2019-02 had a higher mycelium growth rate at 1000 mM. Although TaDP2019-02 isolate had a lower mycelium growth rate at low levels of salinity, it remained stable at high levels of salinity comparing to ThLem2017-01 and TaDP2019-01.
3.2. In Vitro Evaluation of Trichoderma spp. on Turfgrass Seeds under Different Levels of Salinity
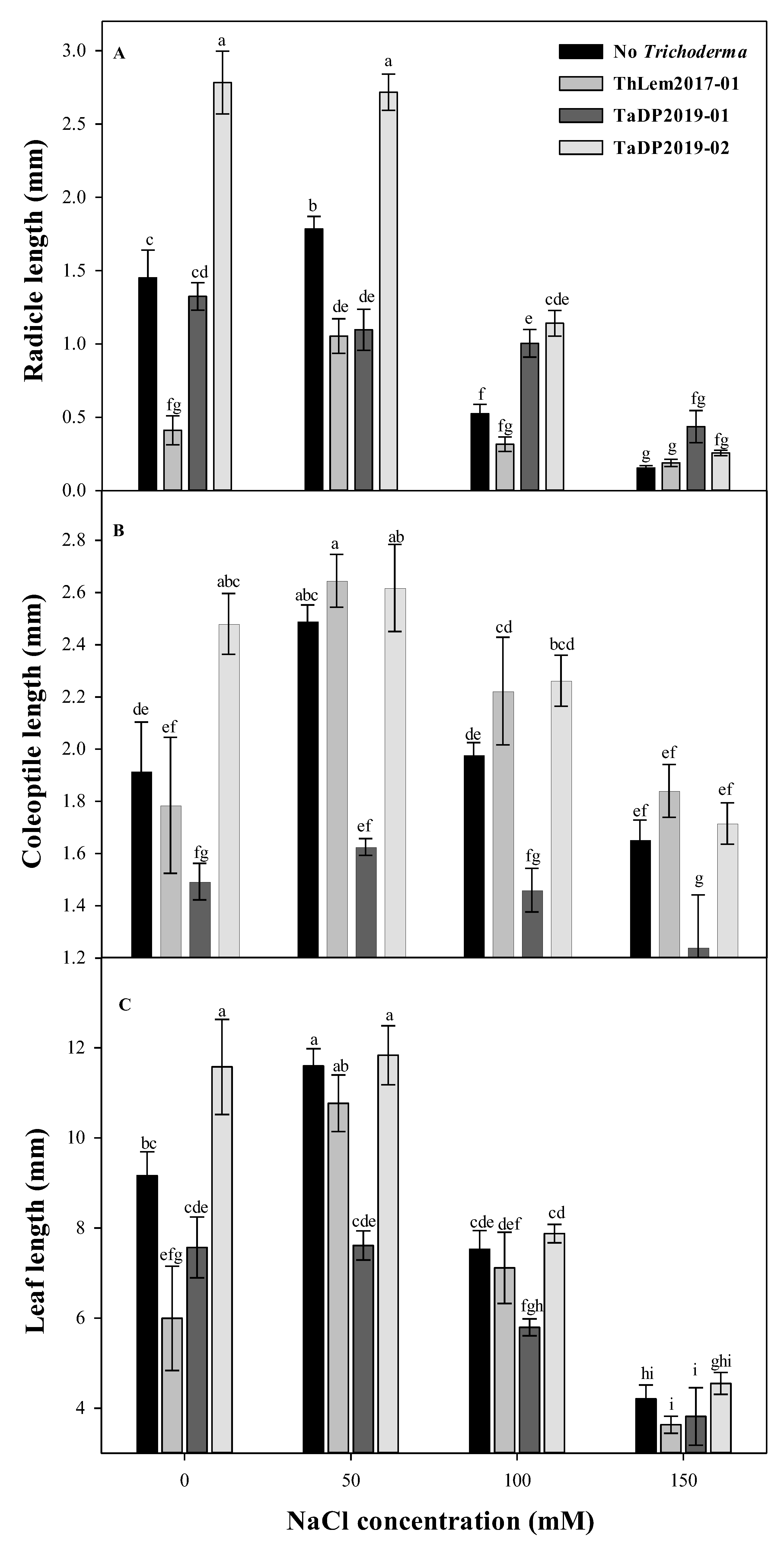
The growth of turfgrass seeds was significantly affected by salt levels and Trichoderma isolates. Increasing salinity levels led to a reduction of radicle length, coleoptile length, and leaf length (Figure 2). The radicle length of plants that inoculated with TaDP2019-02 isolate has increased by 91 to 575% at control compared to other isolates at the same salinity level. While at 4 dS m−1 the radicle length increased by 52 to 158% compared to the other isolates. Interestingly, TaDP2019-02 maintained high radicle length, coleoptile length, and leaf length for each level of salinity. In contrast, the ThLem2017-01 isolate had the lowest radicle length at 0, 50, and 100 mM. In addition, the TaDP2019-01 isolate had the lowest coleoptile length among all isolates and the lowest leaf length at 0, 50, 100, and 150 mM. As a result, the TaDP2019-02 isolate maintained the best morphological performance at all levels of salinity. Therefore, TaDP2019-02 isolate was selected to be used for the greenhouse experiment.
3.3. In-Vivo Effect of Trichoderma Isolate on Perennial Ryegrass under Different Levels of Salinity
Clipping FW and DW of perennial ryegrass were significantly affected by salt levels and Trichoderma treatment (Table 1). The interaction between the salinity and T. atroviride (TaDP2019-02) treatments was not significantly different. The increase in salinity levels (4, 8, and 12 dS m−1) decreased clipping FW and DW. Plants inoculated with Trichoderma had increased FW and DW of leaf clippings. FW and DW of perennial ryegrass that was inoculated with Trichoderma significantly increased by 16 to 114% and 24 to 76% than uninoculated ones, respectively (Table 1). The root mass of perennial ryegrass was more branched and longer when treated with T. atroviride (TaDP2019-02) at all levels of salinity compared to non-treated plants (Figure 3). Roots and shoots dry weight of perennial ryegrass were significantly affected by T. atroviride (TaDP2019-02) and salinity levels. When the salinity level increases the root and shoot dry weight decreases. Trichoderma atroviride (TaDP2019-02) isolate had enhanced root dry weight by 22% and shoot dry weight by 14.3% (Table 1).
The Solvita soil respiration test showed higher soil respiration (Soil-CO2-C, mg kg−1) levels in soil samples that had been inoculated with TaDP2019-02 isolate (Table 2). Soils that were inoculated with T. atroviride (TaDP2019-02) increased soil respiration by 75% compared to non-inoculated plants. There was no effect of salinity levels on CO2 concentration.
Increasing salinity levels decreased the leaf color co-ordinate ‘a*’, a greenness indicator (Table 3). However, Trichoderma (TaDP2019-02) isolate did not affect the color co-ordinate ‘a*’ of perennial ryegrass leaves. The salinity level at 12 dS m−1 increased the color ‘a*’ values by 9 to 16% than the control treatment (tap water). The interaction between the salinity and T. atroviride (TaDP2019-02) treatments was not statistically significant.
4. Discussion
The mycelium growth rate of all Trichoderma spp. decreased by increasing NaCl concentrations. The maximum colony growth was observed at 0 mM NaCl. Current results agreed with Poosapati et al. [35] who found that 50% of the isolates lost their viability when the salinity concentration reached 0.75 M NaCl. In addition, germination rates of most isolates were decreased to 60–70% at 1 M NaCl concentration. Mohamed and Haggag [40] proved that salinity stress had significant negative effects on colony growth and sporulation of wild type of T. harzianum. The response of ThLem2017-01, TaDP2019-01, and TaDP2019-02 isolates to an increase in salinity level were significantly varied among isolates. The response of mycelium growth rate to increase in salinity levels of TaDP2019-02 seems more constant than of ThLem2017-01 and TaDP2019-01 isolates. Rawat et al. [41] had tested 45 T. harzianum isolates, only five isolates tolerated salinity and grew at 240 mM NaCl. De Donno et al. [42] discovered that Trichoderma has in fact better colonized Lake Alimini Grande in Italy. These are saprophytic fungi that belong to ubiquitous species and are capable of colonising aquatic environments and brackish water. In particular, the tested Trichoderma isolates in this current experiment maintain high mycelium growth rates up to 200 mM (18 dS m−1). As a result, these Trichoderma isolates should be effective when irrigated with brackish water. Sedat et al. [43] revealed that several T. harzianum isolates were able to grow their mycelium and sporulate in the growth medium (PDA) at a concentration up to 350 mM salt. In another in vitro study, high levels of salinity (250 mM) caused morphological and physiological changes for the Trichoderma isolates [44]. According to their screening, T. harzianum (T103) was classified as the most salt-tolerant isolate [44]. In case of water reduction in the growing medium, a buildup of specific intracellular substances caused by an increase in synthetic metabolic pathways is one of the physiological changes in this kind of micro-organism [45]. The accumulated substances are neutral osmolites that can play a role as osmoprotective agents [46].
In the in vitro evaluation of the effect of Trichoderma spp. on turfgrass seeds under different levels of salinity, radicle, coleoptile, and leaf length were reduced when salinity levels increased. Salinity is an important abiotic stress that leads to negative effects on many biochemical and physiological mechanisms in seed germination, plant growth and development. In addition, salinity stress causes toxicity (ion accumulation), cell dehydration, and limit water absorption [47]. Shiade and Boelt [48] conducted an experiment on eight varieties of tall fescue at 6 to 15 dS m−1 EC. They stated that seed growth parameters (roots length, seedling, and shoots vigor index) showed a gradual decrease with increasing NaCl concentration. When salinity levels increased over 8 dS m−1, root and shoot growth of perennial ryegrass suffered seriously compared to lower salinity levels (0, 2, and 4 dS m−1) [49].
Although the mycelium growth rate of Trichoderma TaDP2019-02 isolate did not give the superior results all the time under different levels of salinity in the first laboratory experiment, plants that were inoculated with T. atroviride (TaDP2019-02) isolate maintained high radicle length, coleoptile length, and leaf length for each level of salinity. In the second experiment. T. atroviride TaDP2019-02 isolate increased seedling radicle and coleoptile length compared to a seedling that was not treated with Trichoderma sp. Trichoderma atroviride increases the percentage of germination, growth, phenolic content, and the essential oil content of salinity-stressed seedlings of fennel [50]. Lo and Lin [51] reported that cucumber plants that were treated separately with T. harzianum and T. virens strains showed a significant increase in plant height, leaf area, and root dry weight compared to non-treated seedlings. Moreover, Hoyos-Carvajal et al. [52] found that early stages of growth in bean plants were stimulated by Trichoderma strains. Trichoderma spp. promotes plant growth by enhancing nutrient uptake through increased root growth and increasing the availability of essential nutrients around the root zone [53].
In the current research, FW and DW of perennial ryegrass biomass that was inoculated with Trichoderma increased by 16 to 114% and 24 to 76% compared to uninoculated ones, respectively (Table 1). However, exposure to high levels of salinity reduced plant growth in both plants that were inoculated and non-inoculated with Trichoderma compared to plants that had non-saline treatments. The current results are consistent with previous studies which have proven that perennial ryegrass reduced growth rate and turf quality under salt stress. Decreasing in leaf growth in perennial ryegrass is considered as the earliest response to salinity stress [9,54].
The mutual symbiosis of plants with endophytic fungi can improve salt tolerance and reduce crop yield losses caused by saline conditions [55,56]. In Indian mustard, high levels of NaCl (200 mM) reduced mustard plant height by 33.7%, and plant dry weight by 34.5% [57]. However, under the same salinity conditions, shoot and plant DW for plants that were inoculated with T. harzianum increased by 13.8 and 16.7%, respectively [57]. Contreras-Cornejo et al. [58] suggested that T. virens and T. atroviride produce higher levels of indole-3-acetic acid (IAA) once it is grown in a medium supplied with 100 mM NaCl to enhance seed germination and plant growth under salt stress. Sekmen Cetinel et al. [59] found that Trichoderma citrinoviride reduced H2O2 content, which might be responsible for the protection of membrane lipids from peroxidation.
Soil respiration has been used as an indicator for soil health by measuring the CO2 emission that resulted from the respiration of soil micro-organisms [60]. Respiration gives a reliable and scientific basis on which to evaluate microbial activity [61]. In this study, the Solvita soil respiration-CO2 test proved that soils that were inoculated with Trichoderma (TaDP2019-02) had higher respiration in perennial ryegrass rhizospheres. This result is consistent with Wiedow et al. [62] who observed a significant increase in soil respiration in the treatments that were inoculated with T. saturnisporum. In addition, Yadav et al. [63] found that T. viride inoculation increased soil basal respiration.
Leaf color is one of the most crucial parameters to determine the color quality of tissue surfaces. Trichoderma (TaDP2019-02) isolate did not affect the greenness lightness ‘a’ that indicates chromaticity co-ordinates from green (−) to red (+) of perennial ryegrass leaves. High salinity levels have been reported to affect perennial ryegrass color and visual quality [64]. In addition, salt stress reduces the photosynthetic pigments due to a reduction in the synthesis of many enzymes, such as photochlorophyllide reductase and δ-aminolevulinic acid dehydratase [65]. These enzymes have a role in the impairment of ion supply (Mg2+, Fe+, and Mn2+) that are essential for the synthesis of chlorophyll [66]. In contrast, some other researchers found that high levels of salinity increased chloroplast concentration per unit in leaf area. According to their explanation, leaves became thicker and smaller [67]. Hu et al. [9] stated that chlorophyll content in perennial ryegrass had higher values with abiotic stress under greenhouse conditions. Heidari et al. [68] reported that salinity decreased chlorophyll content only in the long-term while chlorophyll content increased by increasing leaf thickness in the short-term of salinity. Inoculation of Trichoderma by seed biopriming improved wheat plants’ net photosynthesis, chlorophyll fluorescence, greenness, and stomatal conductance [69]. Khomari et al. [70] suggested that pretreatment of soybean seedlings with salt-tolerant Trichoderma repressed the decrease in leaf greenness and chlorophyll fluorescence.
This research focused on using Trichoderma isolates on turfgrass (perennial ryegrass) which is commonly used in public and residential landscapes and in sports fields. However, the robust performance of Trichoderma isolate could be applied to other related grass crops and some cereal crops and forage crops in areas that suffer from salinity stress. This should increase the forage production for the purpose of grazing, silage, and hay.
5. Conclusions
Trichoderma atroviride (TaDP2019-02) inoculation holds promise as a beneficial micro-organism and an environmentally friendly alternative to mitigate salinity. The results showed that increasing salinity levels decreased the mycelium growth rate of all Trichoderma isolates. ThLem2017-01 and TaDP2019-01 had a higher mycelium growth rate than TaDP2019-02 when salinity levels were 200, 300, 400, 500 mM while TaDP2019-02 had a higher mycelium growth rate at 1000 mM. The response of mycelium growth rate to an increase in salinity levels of TaDP2019-02 was more constant than the response of ThLem2017-01 and TaDP2019-01. In the second experiment, the results revealed that Trichoderma inoculated-seedlings had higher radicle length, coleoptile length, and leaf length than uninoculated plants. Mainly, TaDP2019-02 isolate maintained the best morphological performance at all levels of salinity. Therefore, TaDP2019-02 isolate was selected to be used for further assessment in the greenhouse experiment. In this experiment, perennial ryegrass clippings FW and DW that were inoculated with TaDP2019-02 isolate were 16 to 114% and 24 to 76% higher than uninoculated ones, respectively. In addition, soils that were inoculated with Trichoderma (TaDP2019-02) had higher CO2 respiration (75%) than the control. T. atroviride (TaDP2019-02) inoculation holds promise as a beneficial micro-organism and an environmentally friendly alternative to mitigate salinity. Therefore, applying Trichoderma isolate to turfgrass and other related grasses has the potential to enhance turfgrass quality and even increase forage production in regions with saline stress.
Author Contributions
Conceptualization, M.G.A.-A. and K.M.A.; methodology, N.S.A.-S.; validation, M.G.A.-A., K.M.A. and Y.A.O.; formal analysis, M.G.A.-A.; investigation, N.S.A.-S.; resources, N.S.A.-S. and M.G.A.-A.; data curation, N.S.A.-S.; writing—original draft preparation, N.S.A.-S.; writing—review and editing M.G.A.-A., K.M.A. and Y.A.O.; supervision, M.G.A.-A. and K.M.A.; project administration, M.G.A.-A.; funding acquisition, M.G.A.-A. All authors have read and agreed to the published version of the manuscript.
Funding
This research was fully funded by the Deanship of Scientific Research at the University of Jordan, Funding number 19/2020/961.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
The authors would like to thank the Deanship of Scientific Research at the University of Jordan, Jordan, for the full funding of this project.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Villwock, M.R.; Meyer, E.G.; Powell, J.W.; Fouty, A.J.; Haut, R.C. Football playing surface and shoe design affect rotational traction. Am. J. Sports Med. 2009, 37, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Roberts, E.C.; Beverly, C.R. Lawn and Sports Turf Benefits; Lawn Institute: East Dundee, IL, USA, 1989. [Google Scholar]
- Christians, N.E.; Aaron, J.P.; Quincy, D. Law. In Fundamentals of Turfgrass Management; John Wiley & Sons: New York, NY, USA, 2016. [Google Scholar]
- Turgeon, A.J. Turfgrass Management, 3rd ed.; Prentice-Hall Inc.: Hoboken, NJ, USA, 1991. [Google Scholar]
- Geren, H.; Riza, A.; Melis, C. Performances of some warm-season turfgrasses under Mediterranean conditions. Afr. J. Biotechnol. 2009, 8, 4469–4474. [Google Scholar]
- Sakr, W.R.A. Response of paspalum turfgrass grown in sandy soil to trinexapac-ethyl and irrigation water salinity. J. Hortic. Sci. Ornam. Plants 2009, 1, 15–26. [Google Scholar]
- Chawla, S.L.; Roshni, A.; Patel, M.; Patil, S.; Shah, H.P. Turfgrass: A billion dollar industry. In Proceedings of the 2018 National Conference on Floriculture for Rural and Urban Prosperity in the Scenerio of Climate Change, Ranipool, India, 16–18 February 2018. [Google Scholar]
- Jensen, K.B.; Asay, K.H.; Waldron, B.L. Dry matter production of orchardgrass and perennial ryegrass at five irrigation levels. Crop Sci. 2001, 41, 479–487. [Google Scholar] [CrossRef] [Green Version]
- Hu, L.; Li, H.; Pang, H.; Fu, J. Responses of antioxidant gene, protein and enzymes to salinity stress in two genotypes of perennial ryegrass (Lolium perenne) differing in salt tolerance. J. Plant Physiol. 2012, 169, 146–156. [Google Scholar] [CrossRef]
- Wang, Q.; Liang, X.; Dong, Y.; Xu, L.; Zhang, X.; Hou, J.; Fan, Z. Effects of exogenous nitric oxide on cadmium toxicity, element contents and antioxidative system in perennial ryegrass. Plant Growth Regul. 2013, 69, 11–20. [Google Scholar] [CrossRef]
- Fan, J.; Zhang, W.; Amombo, E.; Hu, L.; Kjorven, J.O.; Chen, L. Mechanisms of environmental stress tolerance in turfgrass. Agronomy 2020, 10, 522. [Google Scholar] [CrossRef] [Green Version]
- Gross, C.M.; Angle, J.S.; Welterlen, M.S. Nutrient and sediment losses from turfgrass. J. Environ. Qual. 1990, 19, 663–668. [Google Scholar] [CrossRef]
- Pitman, M.G.; Läuchli, A. Global impact of salinity and agricultural ecosystems. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 3–20. [Google Scholar]
- Shalhevet, J. Using water of marginal quality for crop production: Major issues. Agric. Water Manag. 1994, 25, 233–269. [Google Scholar] [CrossRef]
- Atak, M.; Kaya, M.D.; Kaya, G.; Çikili, Y.; Çiftçi, C.Y. Effects of NaCl on the germination, seedling growth and water uptake of triticale. Turk. J. Agric. For. 2006, 30, 39–47. [Google Scholar]
- Othman, Y.; Al-Karaki, G.; Al-Tawaha, A.R.; Al-Horani, A. Variation in germination and ion uptake in barley genotypes under salinity conditions. World J. Agric. Sci 2006, 2, 11–15. [Google Scholar]
- Hussein, H. Lifting the veil: Unpacking the discourse of water scarcity in Jordan. Environ. Sci. Policy 2018, 89, 385–392. [Google Scholar] [CrossRef]
- Othman, Y.A.; Tahat, M.; Alananbeh, K.M.; Al-Ajlouni, M. Arbuscular Mycorrhizal Fungi Inoculation Improves Flower Yield and Postharvest Quality Component of Gerbera Grown under Different Salinity Levels. Agriculture 2022, 12, 978. [Google Scholar] [CrossRef]
- Odeh, T.; Mohammad, A.H.; Hussein, H.; Ismail, M.; Almomani, T. Over-pumping of groundwater in Irbid governorate, northern Jordan: A conceptual model to analyze the effects of urbanization and agricultural activities on groundwater levels and salinity. Environ. Earth Sci. 2019, 78, 40. [Google Scholar] [CrossRef]
- Zhang, Q.; Wang, S.; Rue, K. Salinity tolerance of 12 turfgrasses in three germination media. HortScience 2011, 46, 651–654. [Google Scholar] [CrossRef]
- Tang, J.; Yu, X.; Luo, N.A.; Xiao, F.; Camberato, J.J.; Jiang, Y. Natural variation of salinity response, population structure and candidate genes associated with salinity tolerance in perennial ryegrass accessions. Plant Cell Environ. 2013, 36, 2021–2033. [Google Scholar] [CrossRef] [PubMed]
- Ali Harivandi, M.; Butler, J.D.; Wu, L. Salinity and turfgrass culture. Turfgrass 1992, 32, 207–229. [Google Scholar]
- Alshammary, S.F.; Qian, Y.L.; Wallner, S.J. Growth response of four turfgrass species to salinity. Agric. Water Manag. 2004, 66, 97–111. [Google Scholar] [CrossRef]
- Marcum, K.B.; Pessarakli, M. Salinity tolerance of ryegrass turf cultivars. HortScience 2010, 45, 1882–1884. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, G.; Bigirimana, J.; Elad, Y.; Höfte, M. Induced systemic resistance in Trichoderma harzianum T39 biocontrol of Botrytis cinerea. Eur. J. Plant Pathol. 1998, 104, 279–286. [Google Scholar] [CrossRef]
- Alfano, G.; Ivey, M.L.L.; Cakir, C.; Bos, J.I.B.; Miller, S.A.; Madden, L.V.; Kamoun, S.; Hoitink, H.A.J. Systemic modulation of gene expression in tomato by Trichoderma hamatum 382. Phytopathology 2007, 97, 429–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermosa, R.; Viterbo, A.; Chet, I.; Monte, E. Plant-beneficial effects of Trichoderma and of its genes. Microbiology 2012, 158, 17–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mastouri, F.; Björkman, T.; Harman, G.E. Seed treatment with Trichoderma harzianum alleviates biotic, abiotic, and physiological stresses in germinating seeds and seedlings. Phytopathology 2010, 100, 1213–1221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naeimi, S.; Okhovvat, S.M.; Javan-Nikkhah, M.; Vágvölgyi, C.; Khosravi, V.; Kredics, L. Biological control of Rhizoctonia solani AG1-1A, the causal agent of rice sheath blight with Trichoderma strains. Phytopathol. Mediterr. 2010, 49, 287–300. [Google Scholar]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Alleviation of the adverse effects of salinity stress in wheat (Triticum aestivum L.) by seed biopriming with salinity tolerant isolates of Trichoderma harzianum. Plant Soil 2011, 347, 387–400. [Google Scholar] [CrossRef]
- Papavizas, G.C. Trichoderma and Gliocladium: Biology, ecology, and potential for biocontrol. Annu. Rev. Phytopathol. 1985, 23, 23–54. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 18, pp. 315–322. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Keswani, C.; Ray, S.; Upadhyay, R.S.; Singh, D.P.; Prabha, R.; Sarma, B.K.; Singh, H.B. Isolation and screening of high salinity tolerant Trichoderma spp. with plant growth property and antagonistic activity against various soilborne phytopathogens. Arch. Phytopathol. Plant Prot. 2019, 52, 667–680. [Google Scholar] [CrossRef]
- Poosapati, S.; Ravulapalli, P.D.; Tippirishetty, N.; Vishwanathaswamy, D.K.; Chunduri, S. Selection of high temperature and salinity tolerant Trichoderma isolates with antagonistic activity against Sclerotium rolfsii. SpringerPlus 2014, 3, 641. [Google Scholar] [CrossRef] [Green Version]
- Mukhtar, I. Influence of Trichoderma species on seed germination in okra. Mycopath 2008, 6, 47–50. [Google Scholar]
- Sharma, V.P.; Kumar, S. Spawn Production Technology; Mushrooms Cultivation, Marketing and Consumption, Directorate of Mushroom Research (ICAR): Chambaghat, India, 2011; pp. 35–42. [Google Scholar]
- Dawar, S.; Khaliq, S.; Tariq, M. Comparative effect of plant extract of Datura alba Nees and Cynodon dactylon (L.) Pers., alone or in combination with microbial antagonists for the control of root rot disease of cowpea and okra. Pak. J. Bot 2010, 42, 1273–1279. [Google Scholar]
- Kuehl, R.O.O. Designs of Experiments: Statistical Principles of Research Design and Analysis; Duxbury Press: Duxbury, MA, USA, 2000. [Google Scholar]
- Mohamed, H.A.-L.A.; Haggag, W.M. Biocontrol potential of salinity tolerant mutants of Trichoderma harzianum against Fusarium oxysporum. Braz. J. Microbiol. 2006, 37, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Rawat, L.; Singh, Y.; Shukla, N.; Kumar, J. Salinity tolerant Trichoderma harzianum reinforces NaCl tolerance and reduces population dynamics of Fusarium oxysporum f. sp. ciceri in chickpea (Cicer arietinum L.) under salt stress conditions. Arch. Phytopathol. Plant Prot. 2013, 46, 1442–1467. [Google Scholar] [CrossRef]
- De Donno, A.; Bagordo, F.; Lugoli, F.; Leopizzi, M.I.; Russo, A.; Napoli, C.; Montagna, M.T. Spatial distribution of fungal microflora in the sediment of a brackish lake (Lake Alimini Grande, Italy) used for fish production and bathing. J. Prev. Med. Hyg. 2008, 49, 148–151. [Google Scholar] [PubMed]
- Sedat, Ç.A.M.; Çiğdem, K.Ü.Ç.Ü.K. The Effect of Salinity on Growth, Antagonistic Potential, Protease Activity, and Proline Content of Trichoderma harzianum. Commagene J. Biol. 2020, 4, 62–66. [Google Scholar]
- Mishra, N.; Khan, S.S.; Sundari, S.K. Native isolate of Trichoderma: A biocontrol agent with unique stress tolerance properties. World J. Microbiol. Biotechnol. 2016, 32, 130. [Google Scholar] [CrossRef]
- Soliman, H.G.; El-Sheikh, H.H.; Lashine, I.F. Influence of salt stress on certain metabolic activities of Aspergillus terreus and A. tamarii isolated from the Mediterranean Sea water. Al-Azhar J. Microbiol 1994, 24, 46–57. [Google Scholar]
- Yeo, A. Molecular biology of salt tolerance in the context of whole-plant physiology. J. Exp. Bot. 1998, 49, 915–929. [Google Scholar] [CrossRef]
- Munns, R. Salinity, growth and phytohormones. In Salinity: Environment-Plants-Molecules; Springer: Dordrecht, The Netherlands, 2002; pp. 271–290. [Google Scholar]
- Shiade, S.R.G.; Boelt, B. Seed germination and seedling growth parameters in nine tall fescue varieties under salinity stress. Acta Agric. Scand. Sect. B-Soil Plant Sci. 2020, 70, 485–494. [Google Scholar] [CrossRef]
- Nizam, I. Effects of salinity stress on water uptake, germination and early seedling growth of perennial ryegrass. Afr. J. Biotechnol. 2011, 10, 10418–10424. [Google Scholar]
- Kalleli, F.; Aissa, E.; M’Hamdi, M. Seed biopriming with endophytic fungi enhances germination, growth, yield and fruit quality of fennel under salinity stress. J. Res. Environ. Earth Sci. 2022, 8, 1–10. [Google Scholar]
- Lo, C.-T.; Lin, C.-Y. Screening strains of Trichoderma spp. for plant growth enhancement in Taiwan. J. Phytopathol. 2002, 11, 215–220. [Google Scholar]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Growth stimulation in bean (Phaseolus vulgaris L.) by Trichoderma. Biol. Control 2009, 51, 409–416. [Google Scholar] [CrossRef]
- Kleifeld, O.; Chet, I. Trichoderma harzianum—interaction with plants and effect on growth response. Plant Soil 1992, 144, 267–272. [Google Scholar] [CrossRef]
- Hu, T.; Li, H.; Zhang, X.; Luo, H.; Fu, J. Toxic effect of NaCl on ion metabolism, antioxidative enzymes and gene expression of perennial ryegrass. Ecotoxicol. Environ. Saf. 2011, 74, 2050–2056. [Google Scholar] [CrossRef]
- Waller, F.; Achatz, B.; Baltruschat, H.; Fodor, J.; Becker, K.; Fischer, M.; Heier, T.; Hückelhoven, R.; Neumann, C.; von Wettstein, D.; et al. The endophytic fungus Piriformospora indica reprograms barley to salt-stress tolerance, disease resistance, and higher yield. Proc. Natl. Acad. Sci. USA 2005, 102, 13386–13391. [Google Scholar] [CrossRef] [Green Version]
- Baltruschat, H.; Fodor, J.; Harrach, B.D.; Niemczyk, E.; Barna, B.; Gullner, G.; Janeczko, A.; Kogel, K.; Schäfer, P.; Schwarczinger, I.; et al. Salt tolerance of barley induced by the root endophyte Piriformospora indica is associated with a strong increase in antioxidants. New Phytol. 2008, 180, 501–510. [Google Scholar] [CrossRef]
- Ahmad, P.; Hashem, A.; Abd-Allah, E.F.; Alqarawi, A.A.; John, R.; Egamberdieva, D.; Guce, S. Role of Trichoderma harzianum in mitigating NaCl stress in Indian mustard (Brassica juncea L.) through antioxidative defense system. Front. Plant Sci. 2015, 6, 868. [Google Scholar] [CrossRef] [Green Version]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Del-Val, E.K.; Larsen, J. Ecological functions of Trichoderma spp. and their secondary metabolites in the rhizosphere: Interactions with plants. FEMS Microbiol. Ecol. 2016, 92, fiw036. [Google Scholar] [CrossRef] [Green Version]
- Cetinel, S.H.A.; Gokce, A.; Erdik, E.; Cetinel, B.; Cetinkaya, N. The Effect of Trichoderma citrinoviride Treatment under Salinity Combined to Rhizoctonia solani Infection in Strawberry (Fragaria × ananassa Duch.). Agronomy 2021, 11, 1589. [Google Scholar] [CrossRef]
- Othman, Y.A.; Leskovar, D. Organic soil amendments influence soil health, yield, and phytochemicals of globe artichoke heads. Biol. Agric. Hortic. 2018, 34, 258–267. [Google Scholar] [CrossRef]
- Qin, K.; Dong, X.; Jifon, J.; Leskovar, D.I. Rhizosphere microbial biomass is affected by soil type, organic and water inputs in a bell pepper system. Appl. Soil Ecol. 2019, 138, 80–87. [Google Scholar] [CrossRef]
- Wiedow, D.; Baum, C.; Leinweber, P. Inoculation with Trichoderma saturnisporum accelerates wheat straw decomposition on soil: (Inokulation mit Trichoderma saturnisporum beschleunigt den Abbau von Weizenstroh auf dem Boden). Arch. Agron. Soil Sci. 2007, 53, 1–12. [Google Scholar] [CrossRef]
- Yadav, R.L.; Shukla, S.K.; Suman, A.; Singh, P.N. Trichoderma inoculation and trash management effects on soil microbial biomass, soil respiration, nutrient uptake and yield of ratoon sugarcane under subtropical conditions. Biol. Fertil. Soils 2009, 45, 461–468. [Google Scholar] [CrossRef]
- Tabatabaei, S.J.; Fakhrzad, F. Foliar and soil application of potassium nitrate affects the tolerance of salinity and canopy growth of perennial ryegrass (Lolium perenne var Boulevard). Am. J. Agric. Biol. Sci. 2008, 3, 544–550. [Google Scholar] [CrossRef] [Green Version]
- Behzadi Rad, P.; Roozban, M.R.; Karimi, S.; Ghahremani, R.; Vahdati, K. Osmolyte accumulation and sodium compartmentation has a key role in salinity tolerance of pistachios rootstocks. Agriculture 2021, 11, 708. [Google Scholar] [CrossRef]
- Padmaja, K.; Prasad, D.D.K.; Prasad, A.R.K. Inhibition of chlorophyll synthesis in Phaseolus vulgaris L. seedlings by cadmium acetate. Photosynthetica 1990, 24, 399–405. [Google Scholar]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651. [Google Scholar] [CrossRef] [Green Version]
- Heidari, A.; Bandehagh, A.; Toorchi, M. Effects of NaCl stress on chlorophyll content and chlorophyll fluorescence in sunflower (Helianthus annuus L.) lines. Yuzuncu Yıl Univ. J. Agric. Sci. 2014, 24, 111–120. [Google Scholar] [CrossRef] [Green Version]
- Rawat, L.; Bisht, T.S.; Kukreti, A.; Prasad, M. Bioprospecting drought tolerant Trichoderma harzianum isolates promote growth and delay the onset of drought responses in wheat (Triticum aestivum L.). Mol. Soil Biol. 2016, 7, 1–15. [Google Scholar] [CrossRef]
- Khomari, S.; Davari, M. Trichoderma-induced enhancement of soybean seedling performance in response to salt stress. J. Plant Physiol. Breed. 2017, 7, 27–39. [Google Scholar]
Figure 1.
The mycelium growth rate of the three isolates of Trichoderma spp. under different NaCl concentrations (50, 100, 150, 200, 300, 400, 500, 750, and 1000 mM) (they are equivalent to 4, 8, 12, 18, 28, 38, 44, 64, and 84 dS m−1). Sterile-distilled water was used as a control. Note that 1000 mM = 58.44 g of NaCl per one litre. Measurements were taken after three, six and nine days of subculturing on PDA media. Closed and open symbols represent the mean ± SE for different Trichoderma isolates (n = 3). Error bars represent standard error of the mean.
Figure 1.
The mycelium growth rate of the three isolates of Trichoderma spp. under different NaCl concentrations (50, 100, 150, 200, 300, 400, 500, 750, and 1000 mM) (they are equivalent to 4, 8, 12, 18, 28, 38, 44, 64, and 84 dS m−1). Sterile-distilled water was used as a control. Note that 1000 mM = 58.44 g of NaCl per one litre. Measurements were taken after three, six and nine days of subculturing on PDA media. Closed and open symbols represent the mean ± SE for different Trichoderma isolates (n = 3). Error bars represent standard error of the mean.

Figure 2.
Radicle length (A), coleoptile length (B), and leaf length (C) readings after 20 days under different NaCl concentrations (50, 100, and 150 mM) (they are equivalent to 4, 8, and 12, dS m−1). Sterile-distilled water was used as a control. Note that 1000 mM = 58.44 g of NaCl per one litre. Lowercase letters that share different letters are significantly different according to the least significant difference (LSD) test (p ≤ 0.05). Closed and open symbols represent the mean ± SE for different Trichoderma spp. isolates (n = 4). Error bars represent standard error of the mean.
Figure 2.
Radicle length (A), coleoptile length (B), and leaf length (C) readings after 20 days under different NaCl concentrations (50, 100, and 150 mM) (they are equivalent to 4, 8, and 12, dS m−1). Sterile-distilled water was used as a control. Note that 1000 mM = 58.44 g of NaCl per one litre. Lowercase letters that share different letters are significantly different according to the least significant difference (LSD) test (p ≤ 0.05). Closed and open symbols represent the mean ± SE for different Trichoderma spp. isolates (n = 4). Error bars represent standard error of the mean.

Figure 3.
The root mass of perennial ryegrass with and without T. atroviride (TaDP2019-02) under different salinity levels (0, 4, 8, and 12 dS m−1). The white areas between roots are a white fabric filter (geotextile) to prevent sand from filtering out of the pots.
Figure 3.
The root mass of perennial ryegrass with and without T. atroviride (TaDP2019-02) under different salinity levels (0, 4, 8, and 12 dS m−1). The white areas between roots are a white fabric filter (geotextile) to prevent sand from filtering out of the pots.


{kind=link}
{kind=link}
{kind=link}
Table 1.
ANOVA of clipping fresh and dry weight and shoot and root dry weight for perennial ryegrass with and without (control) T. atroviride under different levels of salinity (0, 4, 8, and 12 dS m−1).
Table 1.
ANOVA of clipping fresh and dry weight and shoot and root dry weight for perennial ryegrass with and without (control) T. atroviride under different levels of salinity (0, 4, 8, and 12 dS m−1).
| Main Effect | Clipping Dry Weight (g) | Shoot Dry Weight (g) | Root Dry Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Day | 88 | 119 | 185 | 88 | 119 | 185 | 190 | 190 | |
| Trichoderma (T) | |||||||||
| With | 1.51 a 1 | 2.21 a | 12.10 a | 0.35 a | 0.46 a | 3.37 a | 19.64 a | 8.22 a | |
| Without 2 | 1.12 a | 1.35 b | 10.31 b | 0.26 a | 0.26 b | 2.71 b | 17.20 b | 6.77 b | |
| LSD | 0.4263 | 0.3629 | 1.5432 | 0.0978 | 0.0882 | 0.4092 | 2.0274 | 1.0278 | |
| Salinity (S) | |||||||||
| 0 | 2.34 a | 2.81 a | 17.84 a | 0.52 a | 0.57 a | 4.64 a | 22.56 a | 13.29 a | |
| 4 | 1.34 b | 1.89 b | 11.62 b | 0.34 b | 0.40 b | 3.16 b | 18.06 b | 8.60 b | |
| 8 | 0.84 bc | 1.33 c | 10.05 b | 0.20 bc | 0.27 bc | 2.79 b | 17.79 b | 4.76 c | |
| 12 | 0.72 c | 1.07 c | 5.30 c | 0.17 c | 0.21 c | 1.57 c | 15.99 b | 3.79 c | |
| LSD | 0.6029 | 0.5132 | 2.1824 | 0.1383 | 0.1248 | 0.5787 | 2.8378 | 1.4477 | |
| ANOVA | |||||||||
| T | 0.0716 | <0.0001 | 0.0246 | 0.0701 | <0.0001 | 0.0026 | 0.0205 | 0.0055 | |
| S | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0009 | <0.0001 | |
| T × S | 0.7829 | 0.3643 | 0.7724 | 0.6976 | 0.3913 | 0.2526 | 0.8192 | 0.7262 | |
Note(s): 1 Numbers followed by different letters within a column are significantly different according to the least significant difference (LSD) test (p ≤ 0.05). 2 Treatment that did not receive Trichoderma inoculation (without) and had a zero salinity level was considered as a control treatment.
Table 2.
ANOVA of CO2 levels for perennial ryegrass soil samples with and without T. atroviride (TaDP2019-02) under two levels of salinity (0 and 12 dS m−1).
Table 2.
ANOVA of CO2 levels for perennial ryegrass soil samples with and without T. atroviride (TaDP2019-02) under two levels of salinity (0 and 12 dS m−1).
| Main Effect | CO2% |
|---|---|
| Trichoderma (T) | |
| With | 2.67 a 1 |
| Without 2 | 1.50 b |
| LSD | 0.3842 |
| Salinity (S) | |
| 0 | 2.25 a |
| 12 | 2.92 a |
| LSD | 0.3843 |
| ANOVA | |
| T | 0.0066 |
| S | 0.2598 |
| T × S | 0.4876 |
Note(s): 1 Numbers followed by different letters within a column are significantly different according to least significant difference (LSD) test (p ≤ 0.05). 2 Treatment that did not receive Trichoderma inoculation (without) and had a zero salinity level was considered as a control treatment.
Table 3.
ANOVA of color parameters for perennial ryegrass with and without T. atroviride (TaDP2019-02) under different levels of salinity (0, 4, 8, and 12 dS m−1).
Table 3.
ANOVA of color parameters for perennial ryegrass with and without T. atroviride (TaDP2019-02) under different levels of salinity (0, 4, 8, and 12 dS m−1).
| Main Effect | L* 1 | a* 2 | b* 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Day | 28 | 43 | 58 | 28 | 43 | 58 | 28 | 43 | 58 | |
| Trichoderma (T) | ||||||||||
| With | 34.72 a 4 | 34.39 a | 34.69 a | −15.65 a | −14.27 a | −13.68 a | 23.42 a | 22.40 a | 22.19 a | |
| Without 5 | 36.13 a | 34.88 a | 35.65 a | −15.07 a | −13.90 a | −13.00 a | 23.23 a | 21.54 a | 21.67 a | |
| LSD | 1.6001 | 1.3312 | 1.1335 | 0.6914 | 0.8886 | 0.7946 | 1.1955 | 0.8731 | 0.7747 | |
| Salinity (S) | ||||||||||
| 0 | 37.69 a | 36.23 a | 36.15 a | −16.50 b | −15.07 b | −14.66 c | 24.26 a | 23.20 a | 22.81 a | |
| 4 | 36.38 ab | 35.15 a | 35.52 ab | −15.36 a | −14.22 ab | −13.26 b | 23.61 ab | 21.79 bc | 22.39 a | |
| 8 | 34.58 bc | 34.73 a | 34.79 ab | −14.89 a | −13.76 a | −13.59 bc | 22.44 ab | 22.16 ab | 21.98 a | |
| 12 | 33.04 c | 32.43 b | 34.23 b | −14.70 a | −13.39 a | −11.86 a | 22.99 b | 20.75 c | 20.52 b | |
| LSD | 2.2629 | 1.8826 | 1.6031 | 0.9778 | 1.2567 | 1.1238 | 1.6908 | 1.2348 | 1.0956 | |
| ANOVA | ||||||||||
| T | 0.0811 | 0.4552 | 0.0932 | 0.0966 | 0.3995 | 0.0914 | 0.7521 | 0.0527 | 0.1823 | |
| S | 0.0012 | 0.0024 | 0.0999 | 0.0032 | 0.0446 | 0.0002 | 0.1699 | 0.0033 | 0.0011 | |
| T × S | 0.3245 | 0.0333 | 0.2516 | 0.8423 | 0.9124 | 0.0756 | 0.6426 | 0.0103 | 0.6053 | |
Note(s): 1 ‘L*’ indicates lightness. 2 ‘a*’ indicates chromaticity co-ordinates from green (−) to red (+). 3 ‘b*’ indicates chromaticity co-ordinates from blue (−) to yellow (+). 4 Numbers followed by different letters within a column are significantly different according to the least significant difference (LSD) test (p ≤ 0.05). 5 Treatment that did not receive Trichoderma inoculation (without) and had a zero salinity level was considered as a control treatment.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Abu-Shanab, N.S.; Alananbeh, K.M.; Othman, Y.A.; Al-Ajlouni, M.G. Effect of Using Trichoderma spp. on Turfgrass Quality under Different Levels of Salinity. Water 2022, 14, 3943. https://doi.org/10.3390/w14233943
AMA Style
Abu-Shanab NS, Alananbeh KM, Othman YA, Al-Ajlouni MG. Effect of Using Trichoderma spp. on Turfgrass Quality under Different Levels of Salinity. Water. 2022; 14(23):3943. https://doi.org/10.3390/w14233943
Chicago/Turabian StyleAbu-Shanab, Nour S., Kholoud M. Alananbeh, Yahia A. Othman, and Malik G. Al-Ajlouni. 2022. "Effect of Using Trichoderma spp. on Turfgrass Quality under Different Levels of Salinity" Water 14, no. 23: 3943. https://doi.org/10.3390/w14233943
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.