
Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna

1
Papanin Institute for Biology of Inland Waters, Russian Academy of Sciences, 152742 Borok, Russia
2
Scientific and Technological Center of Unique Instrumentation, Russian Academy of Sciences, 117342 Moscow, Russia
*
Author to whom correspondence should be addressed.
Water 2022, 14(23), 3955; https://doi.org/10.3390/w14233955
Submission received: 21 November 2022
/
Revised: 29 November 2022
/
Accepted: 2 December 2022
/
Published: 5 December 2022
(This article belongs to the Section Water and Climate Change)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:The intensity of climatic changes and human activities is increasing every year. The general consequence of these processes for freshwater ecosystems can be a dissolved oxygen decrease. There is also a possibility of a reduction in geomagnetic field intensity due to a reversal of the Earth’s magnetic poles. It is assumed that the magnetic poles’ reversal may proceed relatively quickly and coincide with global climatic changes. To evaluate the influence of these processes on aquatic organisms, we studied the effects of different dissolved oxygen levels (2 mg/L, 5 mg/L, and 8 mg/L) under the geomagnetic field (51.7 ± 0.2 μT) and hypomagnetic field (0 ± 0.2 μT) on the model freshwater crustacean Daphnia magna Straus. It was found that reduced oxygen levels and the hypomagnetic field led to a decrease in the sizes of parental females, a reduction in the number of produced offspring, and an increase in the period between broods. The newborns from the first brood in the hypomagnetic field were larger than that from the geomagnetic field. The dissolved oxygen level and magnetic environment affected the age of the first brood release and caudal spine length. The results imply that the probable coincidence of the geomagnetic pole reversal and the decrease in the dissolved oxygen level due to global climatic and geophysical processes will have a more negative impact on freshwater crustaceans than the occurrence of these processes at different times.
1. Introduction
Dissolved oxygen levels have been declining in water bodies during the last several decades [1]. It is due to two reasons. The first is human activities leading to nutrient overloading and expanding eutrophication in coastal and inland waters [2]. It causes seasonal phytoplankton blooms that accumulate enormous biomass. Dissolved oxygen is consumed to decompose this biomass [3,4]. Another reason for hypoxia is global climate change [5]. Oxygen solubility in water directly depends on temperature: increased temperature causes lower levels of dissolved oxygen [6]. In addition, the metabolism of ectotherms needs more oxygen at higher temperatures [7]. Forecasting models predict rising water temperatures due to global warming [5,8].
There is a possibility that global climate changes would coincide with the reversal of the Earth’s magnetic poles. Paleomagnetic data reveal that these events occurred stochastically at different time intervals. The last reversal occurred, presumably, 780 thousand years ago [9,10]. A decrease in the intensity of the geomagnetic field (GMF) up to zero values has accompanied reversals of the Earth’s magnetic poles [11]. Paleomagnetic data show that the GMF reversal may proceed relatively quickly [12,13]. The chaotic nature of the geodynamo makes it impossible to predict the next magnetic field reversal occurrence [14]. The GMF intensity has decreased by more than 10% during the past century. Current observations have also displayed an acceleration of the magnetic north pole movement velocity. Together these processes suggest that a reversal could happen in the near future [15,16].
We could not find papers on the combined effects of the zero magnetic field (co-called hypomagnetic field (HMF)) and dissolved oxygen decrease on aquatic organisms. The probable coincidence of global climate changes with the reversal of geomagnetic poles makes this issue relevant. We used the crustacean Daphnia magna Straus as a model freshwater organism. This species is an essential link in the trophic chain of freshwater ecosystems. Daphnids are easy to cultivate and reproduce rapidly in the laboratory [17]. This work aimed to study the separate and combined impact of HMF and different dissolved oxygen levels on the morphometric and life-history parameters in D. magna. We did not evaluate the effects of temperature due to the limited volume within Helmholtz coils used for the generation of HMF.
2. Materials and Methods
2.1. Daphnia magna
D. magna obtained from Lomonosov Moscow State University were maintained at the Papanin Institute for Biology of Inland Waters of the Russian Academy of Sciences for three years. The stock culture of the crustaceans was held in a hard water medium [18] at 22 ± 0.5 °C. The dissolved oxygen level was ~7.5 mg/L, conductivity ~460 mS/cm, and the pH was ~7.97. Daphnids were maintained under a 16 h/8 h light–dark cycle with about 1000 lux light intensity. The newborn offspring were withdrawn daily. The medium was changed twice a week. Crustaceans were fed daily with a cell suspension of algae Chlorella vulgaris Beijerinck (3 × 107 cells per 100 cm3 of the medium). Algae were cultivated in Bold’s basal medium.
2.2. Oxygen
We used a hard water medium containing 2 mg/L, 5 mg/L, and 8 mg/L of dissolved oxygen. The last value corresponds to the oxygen level in water bodies of the Yaroslavl oblast [19]. The dissolved oxygen levels of 2 mg/L and 5 mg/L simulate a probable decrease in this parameter due to climate changes [20]. The oxygen content of 8 mg/L was obtained by the aeration of a hard water medium (50 L of the medium for 48 h). Reducing the oxygen level to 2 mg/L and 5 mg/L was reached by barbotage of the water medium with nitrogen. Close to zero oxygen content was obtained after 24-h barbotage of the water medium with nitrogen. The water medium containing 2 mg/L and 5 mg/L of oxygen was prepared by dilution of the water medium with oxygenless water. The dissolved oxygen level was checked twice a day with a portable oximeter HI 9146 N (Hanna instruments, Nusfalau, Romania).
Before the experiments, we observed the dynamics of oxygen levels after the preparation of the medium containing 2 mg/L and 5 mg/L of dissolved oxygen. It was found that oxygen level recovers in 24 h to 4.86 mg/L and 6.22 mg/L, correspondingly, due to gas exchange with air. Based on this, we prepared a new water medium with the studied oxygen levels and changed the medium in all experimental groups daily.
2.3. Magnetic Conditions
The GMF of 51.7 ± 0.2 µT and HMF of 0 ± 0.2 µT were utilized for the experiments. HMF simulates possible changes due to the reversal of Earth’s magnetic poles. Generation of HMF was carried out in a system of three pairs of mutually orthogonal Helmholtz coils (made by the Schmidt Institute of Physics of the Earth). Each coil was 0.5 m in diameter and contained 700 turns of 0.2 mm copper wire. The Helmholtz coils were supplied with a direct current from three DC sources (AKIP-1103, Manson Engineering Ltd., Hong Kong, China). Measurements of the vertical and horizontal GMF components were taken before the experiments using a three-component fluxgate magnetometer NV0302A (ENT, Saint Petersburg, Russia). Afterward, the system of Helmholtz coils was positioned so that its axes would coincide with the measured GMF components. The current from DC sources was supplied to each Helmholtz coil winding so that the induced magnetic field was equivalent in intensity while opposite in direction to the measured GMF components. The magnetic fields generated with Helmholtz coils compensated for the GMF in the central area of the system due to the superposition, and HMF was obtained. We checked the parameters of HMF and GMF with an NV0599C magnetometer (ENT, Saint Petersburg, Russia) twice a day.
2.4. Experimental Procedure and Evaluated Parameters
The experiments were performed in polypropylene vials that contained 50 mL of medium with different oxygen levels. A total of 132 neonates, not older than 24-h, (F2 of one parental specimen) were sampled randomly from the stock culture of D. magna and moved individually into vials with a plastic paster pipette. The crustaceans were split into six groups of 22 individuals. Three groups of daphnids in the vials containing medium with dissolved oxygen levels of 2 mg/L, 5 mg/L, and 8 mg/L were placed in the center of the Helmholtz coils (HMF). Three other groups of D. magna in the vials with the same oxygen levels remained under the GMF. Therefore, the following experimental groups of 22 daphnids were studied:
- GMF, dissolved oxygen 8 mg/L;
- GMF, dissolved oxygen 5 mg/L;
- GMF, dissolved oxygen 2 mg/L;
- HMF, dissolved oxygen 8 mg/L;
- HMF, dissolved oxygen 5 mg/L;
- HMF, dissolved oxygen 2 mg/L.
The exposure of the daphnids to the above treatments lasted for 30 days; temperature, photoperiod, etc., corresponded to the cultivation conditions. The experimental animals were fed daily.
The following characteristics were registered daily: the time of the first brood’s release, the number of offspring for each female in the first five broods, and the mortality of the parental specimens. This procedure took about 20 min for each group. Experimental daphnids were not exposed to HMF for this period. Age at first reproduction and mean brood size were determined from observed life history characteristics.
The body length of newborns from the first brood (from the head to the base of the caudal spine) was measured after the brood’s release prior to their first molt. The body length, the carapace height, and the caudal spine length were also measured in the parental daphnids on the last day of experiments. Crustaceans were measured with an MC-2 microscope (Micromed, Saint-Petersburg, Russia) equipped with a DCM 500 Camera (Hangzhou Huaxin IC Technology, Hangzhou, China), using Image-Pro 3.0 software (Media Cybernetics, Bethesda, MD, USA). The raw data could be found in Supplementary File.
The data were tested for normality (Kolmogorov–Smirnov test) and homoscedasticity (Levene’s test). A two-way ANOVA was performed to test for the influence of magnetic conditions and dissolved oxygen content on morphometric and life-history traits of daphnids. Tukey’s post hoc multiple comparisons tests were performed to determine the significance of differences between studied groups. When data were not normally distributed (age at the first reproduction and the period between broods), a Kruskal–Wallis test and the post hoc Dunn’s multiple comparisons test were performed.
3. Results
The mortality of parental daphnids in all treatments by the end of the experiments was less than 8%. This matches the accidental mortality rate in D. magna during standard testing for chronic toxicity [21].
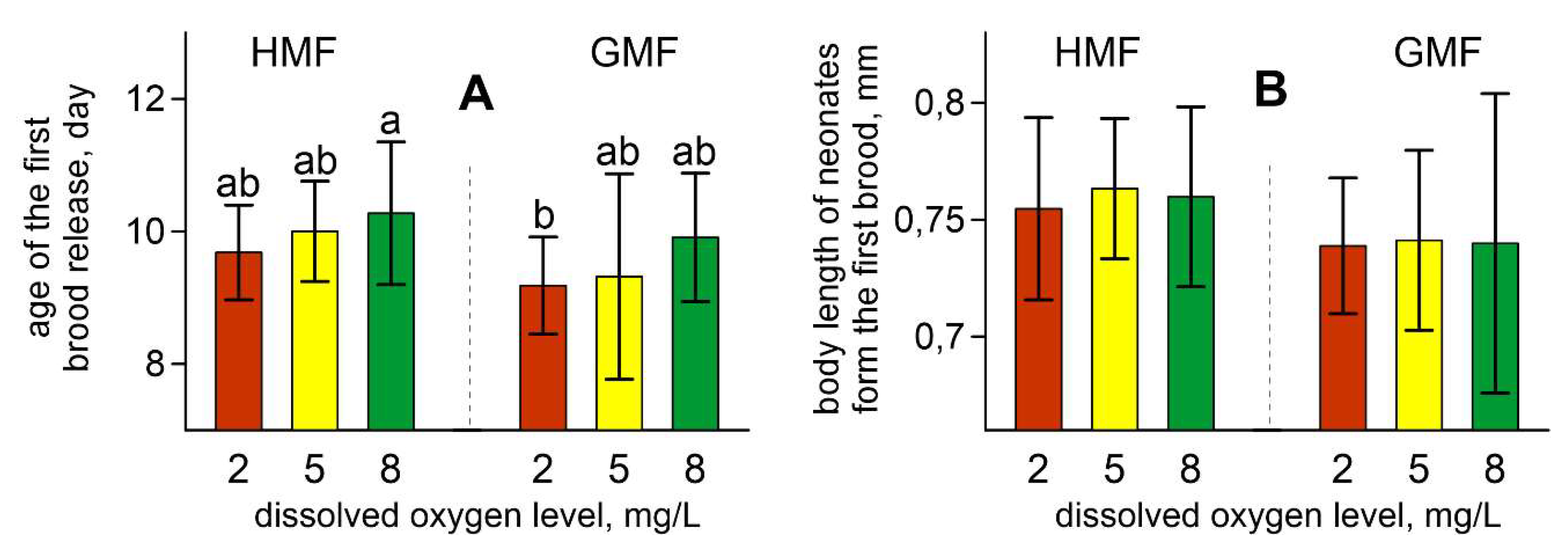
Kruskal–Wallis ANOVA by ranks revealed a significant influence of magnetic conditions on the age of the first brood release (H [1, 132] = 9.92; p < 0.01). The first offspring in HMF appeared later than in GMF (Figure 1A). The dissolved oxygen level also influenced this parameter (H [2, 132] = 9.52; p < 0.01). The age of the first brood release slightly decreased with a reduction of the oxygen concentration. The multiple pairwise comparisons revealed significant differences between daphnids exposed to GMF with an oxygen level of 2 mg/L and exposed to HMF with 8 mg/L of oxygen (Figure 1A).
Magnetic conditions and dissolved oxygen content did not affect the number of offspring in the first brood. However, despite the absence of differences between groups, ANOVA revealed the influence of magnetic conditions on the body length of newborns in the first brood (F [1, 126] = 6.98; p ˂ 0.01; ŋ2 = 0.053). This effect was caused by an increase of this characteristic in the daphnids maintained in HMF compared with those exposed to the GMF (Figure 2). The oxygen level did not affect the body length of newborns in the first brood.
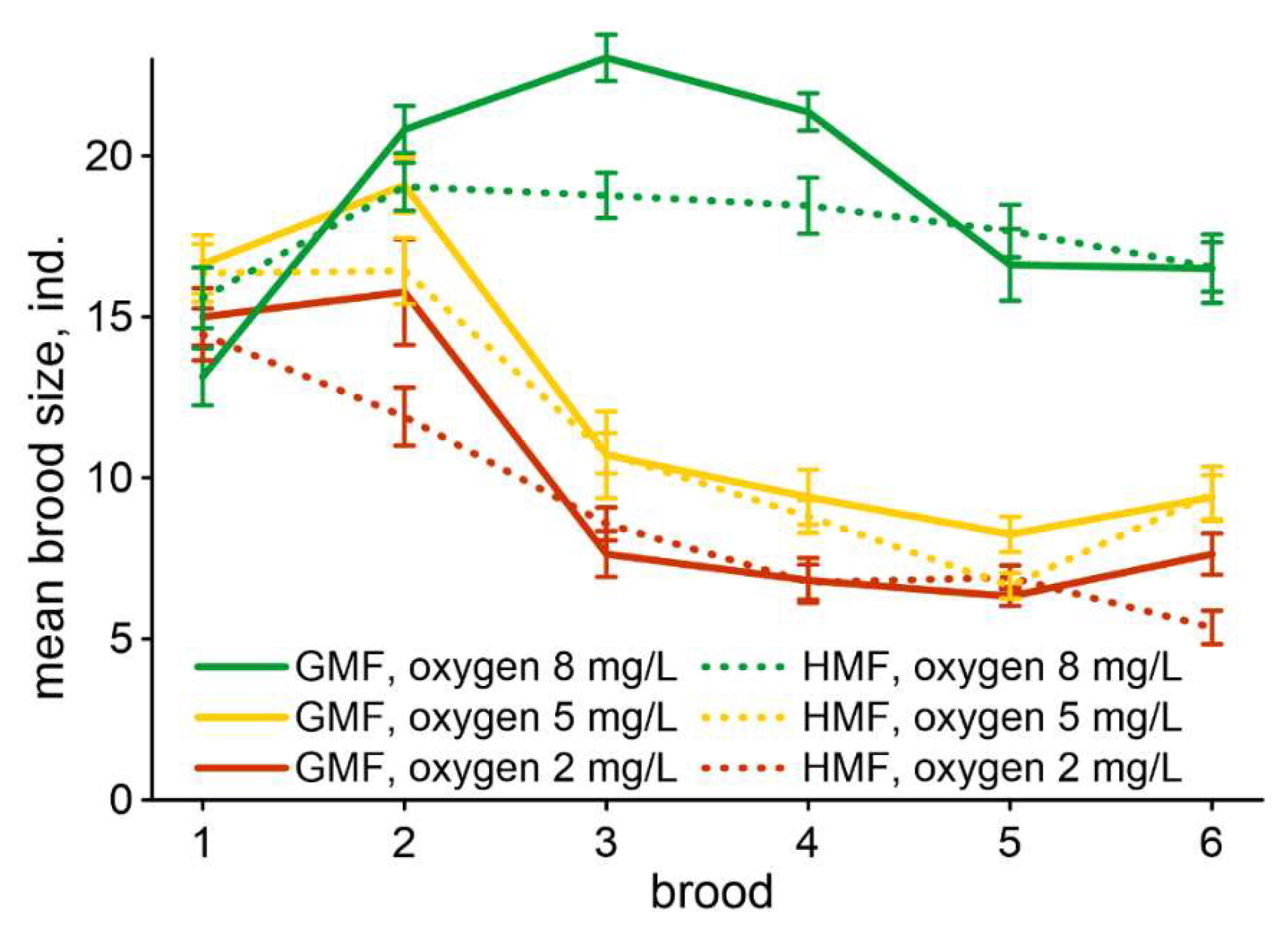
The changes in the number of offspring from the first to the fifth broods are shown in Figure 2. The brood sizes in groups maintained under the oxygen level of 8 mg/L were significantly increased compared to daphnids exposed to lower oxygen levels regardless of magnetic conditions starting from the second brood. Differences between groups in this characteristic are noticeable in the third and fourth broods. The lower the oxygen content, the lower the number of produced offspring. The effect of magnetic background on the brood size is also noticeable in the third and fourth broods in groups exposed to the oxygen level of 8 mg/L: this characteristic in HMF is significantly less than in GMF.
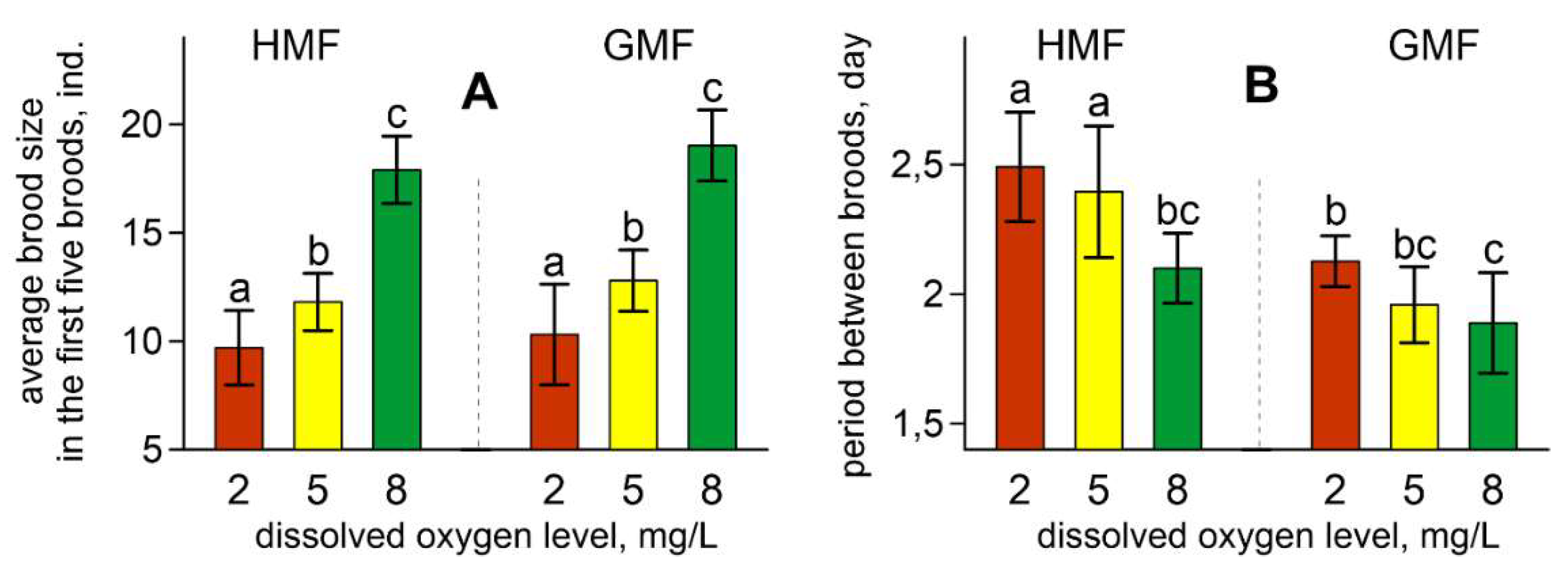
The same pattern can be found when comparing the average brood sizes (Figure 3A). This parameter increases with an increase in the dissolved oxygen level regardless of magnetic conditions. ANOVA revealed the influence of oxygen content on the average brood sizes (F [2, 121] = 286.71; p < 0.001; ŋ2 = 0.826). Magnetic conditions had a weaker effect on this parameter (F [1, 121] = 8.93; p < 0.01; ŋ2 = 0.069) which was caused by a decrease of this parameter under HMF (Figure 3A).
The period between broods was influenced by both dissolved oxygen content (H [2, 130] = 31.27; p < 0.001) and magnetic background (H [1, 130] = 52.78; p < 0.001). The period between broods was longer in HMF compared with GMF. This characteristic increased with a decrease in the oxygen content in the water (Figure 3B).
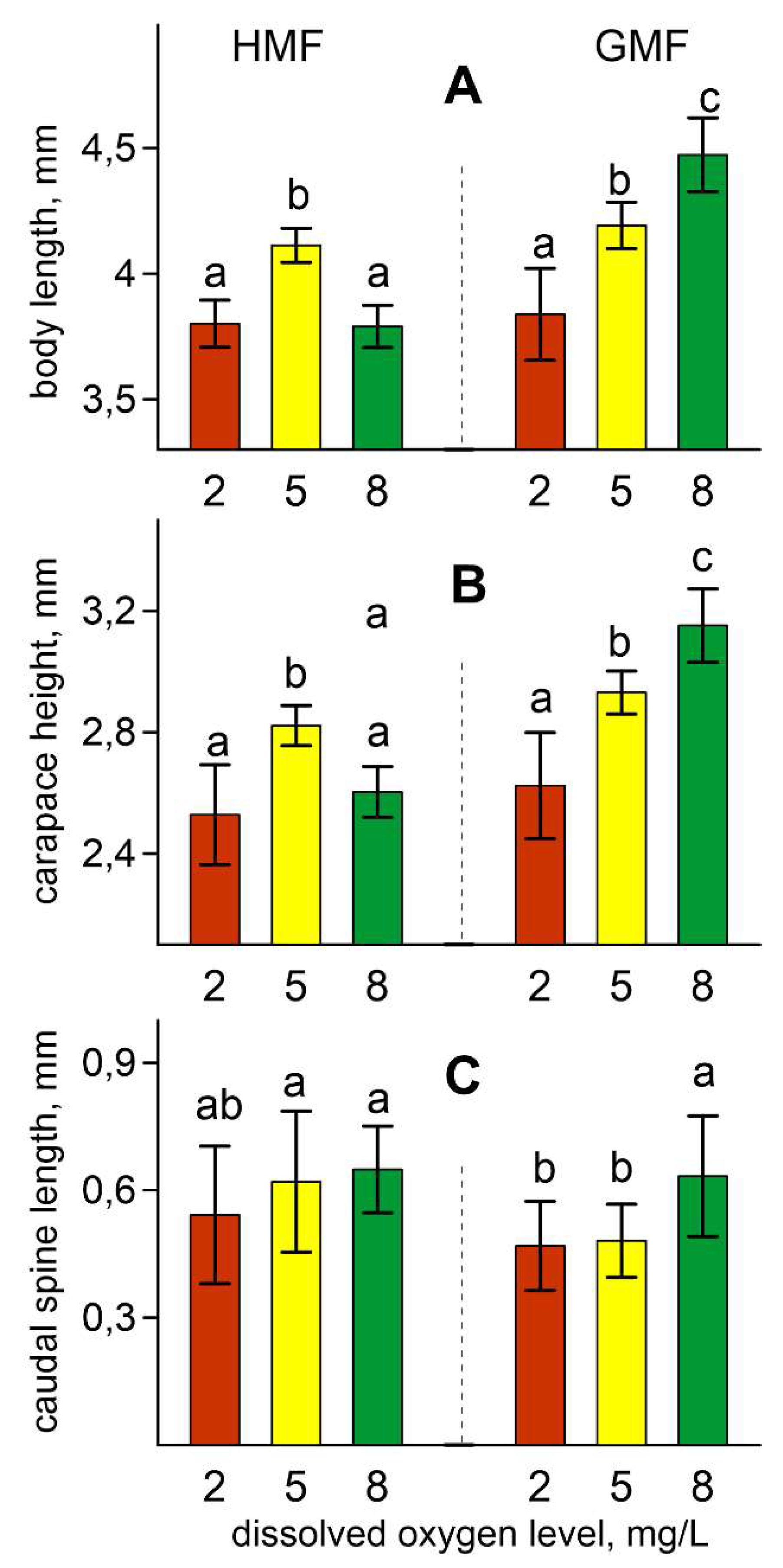
An ANOVA revealed a significant influence of magnetic conditions and dissolved oxygen content on the body length of the parental females on the last day of the experiment (F [1, 119] = 154.91; p < 0.001; ŋ2 = 0.566 and F [2, 119] = 102.60; p < 0.001; ŋ2 = 0.633, correspondingly). These effects arose due to the reduction of this characteristic in daphnids exposed to HMF and the lowest oxygen level of 2 mg/L (Figure 4A). In addition, females’ sizes were influenced by the interaction of magnetic conditions and oxygen level (F [2, 119] = 95.49; p < 0.001; ŋ2 = 0.616). The average body length and carapace height in daphnids exposed to GMF and the maximal dissolved oxygen level of 8 mg/L were significantly increased compared to females exposed to HMF at the same oxygen level (Figure 4A,B).
The oxygen level (F [2, 106] = 11.02; p < 0.001; ŋ2 = 0.172) and magnetic conditions (F [1, 106] = 9.69; p < 0.01; ŋ2 = 0.084) affected the caudal spine length. The first effect is due to the trend towards an increase in this parameter with growth in dissolved oxygen level. The second is associated with an increase in caudal spine length under HMF (Figure 4C).
4. Discussion
D. magna is tolerant to hypoxia species within the Daphnia genus with quite high basal hemoglobin levels [22]. These crustaceans respond to hypoxia by adjustments in hemoglobin concentration and oxygen affinity through changes in gene expression, a decline in filtering and respiration rates, and an increase in heartbeat rate [23,24,25,26]. Daphnids’ reproduction indicators decrease with long-term maintenance in a hypoxic environment since a part of the resources and energy is spent on adaptations to a low level of dissolved oxygen. Thus, the moving of crustaceans to deep layers of the hypolimnion with low oxygen levels in natural habitats led to the depression of reproduction in terms of a high percentage of males in the population and the low egg ratio [27]. A decrease in the reproductive potential of daphnids and an increase in the period between broods with a reduction of dissolved oxygen concentration in experiments were registered earlier [28,29]. Another well-described effect of a decrease in dissolved oxygen level is body size reduction [30]. A smaller body has a greater surface-to-body volume ratio, i.e., the respiratory surface area for oxygen uptake per unit of the body’s volume is lower in a bigger body [31]. Moreover, oxygen is transported over shorter distances within a smaller body [29,32]. Thus, the reduction in the reproductive potential and body size of D. magna in response to a decline in the level of dissolved oxygen revealed in this study is consistent with the previously published data.
The effects of HMF on the reproduction and sizes of daphnids (a reduction in the number of produced offspring, an increase in the period between broods, and a decrease in the size of females) were similar to the effects of hypoxia. The same effects in response to exposure to HMF were reported in D. magna earlier. Thus, in a previous study, we found a reduction of offspring produced in the 2nd, 4th, and 5th broods and a decrease in the size of parental individuals due to the long-term maintenance of daphnids in HMF [33]. A decrease in the reproductive potential and the sizes of parental specimens with a weakening of the geomagnetic field induction was shown in other invertebrate species: small brown planthopper (Laodelphax striatellus) and brown planthopper (Nilaparvata lugens) [34]. A mechanism of HMF influence on organisms is most likely related to the Zeeman effect. This effect consists of the spectral lines splitting in the particles with a magnetic moment in biomolecules into several components under the static magnetic field [35].
Exposure to HMF also increased the size of the produced offspring in the first brood. This effect is due to the general reproductive strategy described for daphnia under adverse conditions [36,37], including magnetic fields [38]. Under unfavorable conditions, crustaceans produce larger and not numerous offspring, which can have a competitive advantage in comparison with numerous broods that consist of small newborns.
Additive adverse effects are noticeable as a result of the combined action of HMF and low oxygen levels on daphnids. The smallest sizes of parental individuals, the lowest number of offspring produced, and the maximum period between broods were recorded in groups exposed to HMF with an oxygen level of 2 mg/L. Thereby, the obtained results imply that the reduction of GMF intensity can be a significant additional factor that aggravates the effects of a decreased dissolved oxygen level. It allows us to assume that the probable coincidence of geomagnetic pole reversal and the decrease of dissolved oxygen level in the water due to global climatic and geophysical processes can have a more adverse impact on freshwater crustaceans than the occurrence of these processes at different times.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w14233955/s1, Data spreadsheet.
Author Contributions
Conceptualization, V.V.K. and A.A.S.; methodology, V.V.K.; formal analysis, A.A.S., D.A.S. and V.V.K.; investigation, A.A.S. and D.A.S.; resources, V.V.K.; data curation, A.A.S., D.A.S. and V.V.K.; writing, V.V.K.; visualization, V.V.K.; project administration, V.V.K. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Russian Science Foundation, project #22-24-20053 (https://rscf.ru/en/project/22-24-20053/ accessed on 5 December 2022).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included as Supplementary Files.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Breitburg, D.; Levin, L.A.; Oschlies, A.; Grégoire, M.; Chavez, F.P.; Conley, D.J.; Garçon, V.; Gilbert, D.; Gutiérrez, D.; Isensee, K.; et al. Declining oxygen in the global ocean and coastal waters. Science 2018, 359, eaam7240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz, R.J.; Rosenberg, R. Introduction to environmental and economic consequences of hypoxia. Int. J. Water Resour. Dev. 2011, 27, 71–82. [Google Scholar] [CrossRef]
- Nixon, S.W. Coastal marine eutrophication: A definition, social causes, and future concerns. Ophelia 1995, 41, 199–219. [Google Scholar] [CrossRef]
- Zhou, F.; Chai, F.; Huang, D.; Wells, M.; Ma, X.; Meng, Q.; Xue, H.; Xuan, J.; Wang, P.; Ni, X.; et al. Coupling and decoupling of high biomass phytoplankton production and hypoxia in a highly dynamic coastal system: The Changjiang (Yangtze River) estuary. Front. Mar. Sci. 2020, 7, 259. [Google Scholar] [CrossRef]
- Whitney, M.M. Observed and projected global warming pressure on coastal hypoxia. Biogeosciences 2022, 19, 4479–4497. [Google Scholar] [CrossRef]
- Roman, M.R.; Brandt, S.B.; Houde, E.D.; Pierson, J.J. Interactive effects of hypoxia and temperature on coastal pelagic zooplankton and fish. Front. Mar. Sci. 2019, 6, 139. [Google Scholar] [CrossRef] [Green Version]
- Vaquer-Sunyer, R.; Duarte, C.M. Temperature effects on oxygen thresholds for hypoxia in marine benthic organisms. Glob. Chang. Biol. 2011, 17, 1788–1797. [Google Scholar] [CrossRef]
- Altieri, A.H.; Gedan, K.B. Climate change and dead zones. Glob. Chang. Biol. 2015, 21, 1395–1406. [Google Scholar] [CrossRef]
- Lutz, T. The magnetic reversal record is not periodic. Nature 1985, 317, 404–407. [Google Scholar] [CrossRef]
- Cande, S.C.; Kent, D.V. Revised calibration of the geomagnetic polarity timescale for the Late Cretaceous and Cenozoic. J. Geophys. Res. 1995, 100, 6093–6095. [Google Scholar] [CrossRef]
- Valet, J.-P.; Fournier, A. Deciphering records of geomagnetic reversals. Rev. Geophys. 2016, 54, 410–446. [Google Scholar] [CrossRef] [Green Version]
- Bogue, S.W.; Glen, J.M.G. Very rapid geomagnetic field change recorded by the partial remagnetization of a lava flow. Geophys. Res. Lett. 2010, 37, L21308. [Google Scholar] [CrossRef] [Green Version]
- Coe, R.S.; Prevot, M.; Camps, P. New evidence for extraordinarily rapid change of the geomagnetic field during a reversal. Nature 1995, 374, 687–692. [Google Scholar] [CrossRef]
- Hulot, G.; Lhuillier, F.; Aubert, J. Earth’s dynamo limit of predictability. Geophys. Res. Lett. 2010, 37, L06305. [Google Scholar] [CrossRef]
- Liu, Q. Will the Earth’s magnetic reverse its polarity in the near future? Solid Earth Sci. 2020, 5, 247–248. [Google Scholar] [CrossRef]
- Cooper, A.; Turney, C.S.M.; Palmer, J.; Hogg, A.; McGlone, M.; Wilmshurst, J.; Lorrey, A.M.; Heaton, T.J.; Russell, J.M.; McCracken, K.; et al. A global environmental crisis 42000 years ago. Science 2021, 371, 811–818. [Google Scholar] [CrossRef]
- Peters, R.H. Daphnia culture. Mem. Ist. Ital. Idrobiol. 1987, 45, 483–495. [Google Scholar]
- ASTM. Standard Practice for Conducting Acute Toxicity Tests with Fishes, Macroinvertebrates and Amphibians; American Standards for Testing and Materials: Philadelphia, PA, USA, 1980. [Google Scholar]
- Bikbulatov, E.S.; Bikbulatova, E.M.; Litvinov, A.S.; Poddubnyi, S.A. Gidrologiya i Gidrokhimiya Ozera Nero (Hydrology and Hydrochemistry of Lake Nero); Rybinskii Dom Pechati: Rybinsk, Russia, 2003; pp. 20–153. [Google Scholar]
- Kay, J.E.; Deser, C.; Phillips, A.; Mai, A.; Hannay, C.; Strand, G.; Arblaster, J.; Bates, S.; Danabasoglu, G.; Edwards, J.; et al. The community Earth system model (CESM) large ensemble project: A community resource for studying climate change in the presence of internal climate variability. Bull. Am. Meteorol. Soc. 2015, 96, 1333–1349. [Google Scholar] [CrossRef] [Green Version]
- OECD. OECD Guideline for Testing of Chemicals, 211: Daphnia magna Reproduction Test; Organization for Economic Cooperation and Development: Paris, France, 2012. [Google Scholar]
- Wilczynski, W.; Babkiewicz, E.; Pukos, S.; Wawrzeńczak, J.; Zebrowski, M.L.; Banasiak, L.; Kudriashov, M.; Maszczyk, P. The effects of hypoxia on threshold food concentrations in different Daphnia Species. Water 2022, 14, 3213. [Google Scholar] [CrossRef]
- Heisey, D.; Porter, K.G. The effect of ambient oxygen concentration on filtering and respiration rates of Daphnia galeata mendotae and Daphnia magna. Limnol. Oceanogr. 1977, 22, 839–845. [Google Scholar] [CrossRef] [Green Version]
- Paul, R.; Colmorgen, M.; Hüller, S.; Tyroller, F.; Zinkler, D. Circulation and respiratory control in millimetre-sized animals (Daphnia magna, Folsomia candida) studied by optical methods. J. Comp. Physiol. B 1997, 167, 399–408. [Google Scholar] [CrossRef]
- Paul, R.J.; Zeis, B.; Lamkemeyer, T.; Seidl, M.; Pirow, R. Control of oxygen transport in the microcrustacean Daphnia: Regulation of haemoglobin expression as central mechanism of adaptation to different oxygen and temperature conditions. Acta Physiol. Scand. 2004, 182, 259–275. [Google Scholar] [CrossRef] [PubMed]
- Zeis, B.; Becher, B.; Goldmann, T.; Clark, R.; Vollmer, E.; Bölke, B.; Bredebusch, I.; Lamkemeyer, T.; Pinkhaus, O.; Pirow, R.; et al. Differential haemoglobin gene expression in the crustacean Daphnia magna exposed to different oxygen partial pressures. Biol. Chem. 2003, 384, 1133–1145. [Google Scholar] [CrossRef]
- Hanazato, T.; Yasuno, M.; Hosomi, M. Significance of a low oxygen layer for a Daphnia population in lake Yunoko, Japan. Hydrobiologia 1989, 185, 19–27. [Google Scholar] [CrossRef]
- Nebeker, A.V.; Dominguez, S.E.; Chapman, G.A.; Onjukka, S.; Stevens, D.G. Effects of low dissolved oxygen on survival, growth and reproduction of Daphnia, Hyalella and Gammarus. Environ. Toxicol. Chem. 1992, 11, 373–379. [Google Scholar] [CrossRef]
- Lyu, K.; Wang, Q.; Yang, Z.; Montagnes, D.J.S. Cumulative effects matter: Multi-brood responses of Daphnia to hypoxia. Freshw. Biol. 2016, 61, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Seidl, M.D.; Paul, R.J.; Pirow, R. Effects of hypoxia acclimation on morpho-physiological traits over three generations of Daphnia magna. J. Exp. Biol. 2005, 208, 2165–2175. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, D.; Morley, S.A.; Hughes, R.N. From cells to colonies: At what levels of body organization does the ‘temperature-size rule’ apply? Evol. Dev. 2006, 8, 202–214. [Google Scholar] [CrossRef]
- Czarnoleski, M.; Ejsmont-Karabin, J.; Angilletta, M.J.; Kozlowski, J. Colder rotifers grow larger but only in oxygenated waters. Ecosphere 2015, 6, 164. [Google Scholar] [CrossRef]
- Krylov, V.V.; Bolotovskaya, I.V.; Osipova, E.A. The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagn. Biol. Med. 2013, 32, 30–39. [Google Scholar] [CrossRef]
- Wan, G.J.; Jiang, S.L.; Zhao, Z.C.; Xu, J.J.; Tao, X.R.; Sword, G.A.; Gao, Y.B.; Pan, W.D.; Chen, F.J. Bio-effects of near-zero magnetic fields on the growth, development and reproduction of small brown planthopper, Laodelphax striatellus and brown planthopper, Nilaparvata lugens. J. Insect Physiol. 2014, 68, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Binhi, V.N.; Prato, F.S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 2017, 12, e0179340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enserink, L.; de la Haye, M.; Maas, H. Reproductive strategy of Daphnia magna: Implications for chronic toxicity tests. Aquat. Toxicol. 1993, 25, 111–123. [Google Scholar] [CrossRef]
- Burns, C.W. Effects of crowding and different food levels on growth and reproductive investment of Daphnia. Oecologia 1995, 101, 234–244. [Google Scholar] [CrossRef]
- Krylov, V.V.; Osipova, E.A. The response of Daphnia magna Straus to the long-term action of low-frequency magnetic fields. Ecotoxicol. Environ. Saf. 2013, 96, 213–219. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Age of the first brood release (A) and newborns’ body length (B) in studied groups of daphnids. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) after Kruskal–Wallis test and Dunn’s post-hoc test. For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.
Figure 1.
Age of the first brood release (A) and newborns’ body length (B) in studied groups of daphnids. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) after Kruskal–Wallis test and Dunn’s post-hoc test. For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.

Figure 2.
Dynamics of changing mean brood size in studied groups of daphnids. Bars denote standard error.
Figure 2.
Dynamics of changing mean brood size in studied groups of daphnids. Bars denote standard error.

Figure 3.
Average brood size in the first five broods (A) and the period between broods (B) in studied groups of daphnids. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) (p < 0.05) after Tukey’s post-hoc multiple comparison tests (A) or Kruskal–Wallis test and Dunn’s post-hoc test (B). For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.
Figure 3.
Average brood size in the first five broods (A) and the period between broods (B) in studied groups of daphnids. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) (p < 0.05) after Tukey’s post-hoc multiple comparison tests (A) or Kruskal–Wallis test and Dunn’s post-hoc test (B). For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.

Figure 4.
The body length (A), carapace height (B), and caudal spine length (C) in parental females. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) after Tukey’s post-hoc multiple comparison tests. For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.
Figure 4.
The body length (A), carapace height (B), and caudal spine length (C) in parental females. Values are means ± standard deviation. Different letters indicate significant differences among groups (p < 0.05) after Tukey’s post-hoc multiple comparison tests. For all variables with the same letters, the difference between the groups is not statistically significant. Significantly different groups have different letters.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Krylov, V.V.; Sizova, A.A.; Sizov, D.A. Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna. Water 2022, 14, 3955. https://doi.org/10.3390/w14233955
AMA Style
Krylov VV, Sizova AA, Sizov DA. Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna. Water. 2022; 14(23):3955. https://doi.org/10.3390/w14233955
Chicago/Turabian StyleKrylov, Viacheslav V., Anastasia A. Sizova, and Daniil A. Sizov. 2022. "Effects of Hypoxia and Hypomagnetic Field on Morphometric and Life-History Traits in Freshwater Cladoceran Daphnia magna" Water 14, no. 23: 3955. https://doi.org/10.3390/w14233955
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.