
A Virological Perspective on the Use of Bacteriophages as Hydrological Tracers


1
Environmental Research and Innovation Department (ERIN), Luxembourg Institute of Science and Technology (LIST), 4422 Belvaux, Luxembourg
2
Faculté des Sciences, de la Technologie et de la Communication (FSTC), Doctoral School in Science and Engineering (DSSE), University of Luxembourg, 4365 Esch-sur-Alzette, Luxembourg
*
Author to whom correspondence should be addressed.
Water 2022, 14(24), 3991; https://doi.org/10.3390/w14243991
Submission received: 8 November 2022
/
Revised: 29 November 2022
/
Accepted: 1 December 2022
/
Published: 7 December 2022
(This article belongs to the Section Hydrology)
Abstract
:Hydrological tracers, commonly used for characterizing water flow paths and sources, do not often meet all the requirements of an ideal tracer. Trans-disciplinary approaches are advocated as the way forward to enlarge the number of tracers available for investigating hydrological processes. Since the 19th century, hydrological tracers have been increasingly used, particularly in underground areas. The tracer toolbox at hand includes a large variety of options, including fluorescent dyes, isotopes, salts or bacteriophages, with each tracer offering specific qualities and complementarities. While their potential for hydrological studies has been studied in karstic environments since the 1960s, bacteriophages remain insufficiently understood. According to the selection methodology used in this review, more than thirty experiments have been listed, involving in total around seventeen different bacteriophages. These have facilitated the investigation of groundwater, surface water (i.e., river, lake and marine water), wetland and wastewater hydrological processes. The tracing experiments have also highlighted the possible interaction between bacteriophages and the surrounding environments. Bacteriophages have successfully helped researchers to understand the water flow within watersheds. Certain advantages, such as the sensitivity of detection, the ease of producing high concentrations of bacteriophages to be injected, their specificity for a host and their non-pathogenicity for human and animal cells, make bacteriophages appreciable tracer candidates for tracing experiments. However, the adsorption process or environmental factors such as temperature, pH and UV light considerably impact the fate of bacteriophages, thereby leading to an attenuation of the phage signal. Considering both the flaws and the qualities of bacteriophages, their use as hydrological tracers requires new insight and further discussions regarding experimental tracing conditions.
1. Introduction
Hydrological tracers have been used for more than a century and have allowed the study of water in all its phases in a wide range of contrasting environments [1]. They also contribute to addressing fundamental questions on hydrological processes, such as the sources of water and matter, flow paths and transit times [2,3], velocities and travel times, hydrodynamic dispersion, groundwater recharge and discharge generation [1,4]. The first known use of tracers dates from the late 1800s, when they were used to investigate groundwater recharge and velocity in European karst aquifers [1,5].
Currently, the tracing tools at hand can be divided into two categories, namely tracers that are naturally or accidentally present in the environment (i.e., environmental tracers) and those that are artificially and intentionally introduced into the environment (i.e., artificial tracers). For example, stable (e.g., hydrogen or oxygen) and radioactive (e.g., tritium) isotopes are environmental tracers [6,7], while salts or dyes are artificial tracers [1,8].
In experimental hydrology, the most commonly used tracers, defined as conventional tracers, are isotopes and fluorescent dyes. In parallel to the use of the previous tracers, additional tracers can be employed, either by considering available natural resources such as diatoms or by introducing colloidal particles such as bacteriophages and microspheres [7,9]. However, the use of these tracers is less widespread than that of conventional tracers. To choose the right tracer among a large set of options, eight criteria are usually considered: (i) low sorptivity and sorption, (ii) good chemical and biological stability, (iii) good solubility, (iv) low background concentration, (v) insensitive to changes (light, pH, temperature), (vi) efficient detection limit, (vii) low toxicity and environmental impact and (viii) low cost. Above all, the perfect tracer should be conservative, i.e., stable and non-reactive with the surrounding environment [7]. Unfortunately, such an ideal tracer does not exist. Although hydrogen or oxygen-stable isotopes are often considered as such, since they are the basic constituents of the water molecule [4,10], recent studies have refuted this fundamental statement [11]. For instance, the initial enthusiasm for the use of stable oxygen and hydrogen isotopes for hydrograph separation has been gradually replaced by substantial concerns about their limitations in, for example, determining flow paths to reach streams during storms [12]. While the analytical techniques used to measure the different types of tracers are generally robust and reliable, the interpretation of tracer signals is far from straightforward. The non-conservative behaviour of most tracers is a major concern in this respect [13]. Fluorescent dyes are best suited to sufficiently transparent waters, but they are quickly attenuated by sunlight and, like rhodamine B, are easily adsorbed on the surrounding materials (e.g., soil constituents) [4,6,14]. In addition, the determination of dye mixtures and dye residuals from the initial spiking can be challenging [15]. Salts, such as bromide or chloride, have also shown their limitations when it comes to differentiating their presence from the natural background [14]. Geochemical elements and isotopes of hydrogen and oxygen in water are cardinal tools for investigating the time source (i.e., event vs. pre-event water), runoff generation mechanisms (e.g., saturation overland flow, groundwater flow, perched saturated flow) and geographic origins (i.e., location of the water before it arrived at the stream) of storm runoff components [16]. Despite considerable knowledge having been gained from countless studies in small experimental catchment areas over the past four decades, theoretical research has gradually replaced field observations and empirical understanding, mainly due to budgetary constraints on the one hand and difficulties in identifying suitable sampling sites in larger catchment areas on the other [17,18]. When considering larger catchment areas, runoff generation mechanisms have also been increasing and form a highly heterogeneous mixture of water types [19]. Consequently, isotope tracers have been preferentially used in small catchment areas [12], and their application to larger scales remains highly challenging [20]. For overcoming this status quo, complementary tools have been proposed, such as distributed fibre-optic temperature sensing [21,22], thermal infrared (IR) imagery [23,24], the use of terrestrial diatoms as environmental biological tracers of hydrological connectivity across multiple scales [25,26], artificial DNA tracers [27,28] or the injection of bacteriophages [14,29]. Bacteriophages are viruses that specifically infect bacterial hosts. They are almost exclusively composed of a nucleic acid molecule (DNA or RNA) carrying the genetic information, which is protected by a protein capsid [30,31]. They exhibit a wide range of shapes, sizes (20 to 200 nm) and structures. Bacteriophages were introduced as hydrological tracers in the late 1960s, and their suitability for such applications has been investigated since then. Three main reasons have motivated hydrologists in the use of bacteriophages as hydrological tracers: (i) the sensitivity of the detection method, (ii) their host specificity and (iii) their so-called eco-friendly properties [32,33,34,35,36]. Yet, the reasons mentioned above must be carefully considered because, if not properly investigated before using bacteriophages as tracers, these aspects may unknowingly hinder the efficient conduct of the tracing experiments.
The objective of this review was to provide a synthetic overview of past and current uses of bacteriophages as hydrological tracers to identify the advantages and limitations of such an approach from an original virological perspective. To achieve this objective, all facets of the use of bacteriophages in the previously reported tracing experiments were reviewed according to the methodology set up and described in the first part of the review. A description of the used bacteriophages and their effectiveness was then carried out to further highlight the limitations encountered when using them. Finally, we provide solutions, improvements, and guidance for future applications, based on our understanding of the fate and transportation of viruses in the environment.
2. Methodology
2.1. Search Strategy
To identify all relevant scientific studies concerning the use of bacteriophages as a hydrological tracer over the years, the literature review was conducted using six different online bibliographic databases, namely Web of Science (https://mjl.clarivate.com/search-results, accessed on 7 November 2022), Science Direct (https://www.sciencedirect.com/, accessed on 7 November 2022), PubMed (https://pubmed.ncbi.nlm.nih.gov/, accessed on 7 November 2022), Wiley Online Library (https://onlinelibrary.wiley.com/, accessed on 7 November 2022), Google Scholar (https://scholar.google.com/, accessed on 7 November 2022) and SpringerLink (https://link.springer.com/, accessed on 7 November 2022). The literature review was conducted until June 2022. For each bibliographic database, the first phase of the search consisted of identifying and counting the number of articles matching the keywords (1) “Phage*” or “Bacteriophage*” and (2) “Hydrological tracer*” or “tracer*”, with the linguistic annotation required by each database. All articles meeting the above-mentioned keyword combination were retained for the second selection phase. It should be noted that book chapters and conference abstracts were also included.
After the initial screening of the bibliographic database using the above-mentioned keywords, all duplicates were removed, and an additional manual screening based on titles and abstracts was performed in order to select only publications using bacteriophages as hydrological tracers and to eliminate studies which conducted tracing experiments for any purpose other than hydrology (e.g., microbial source tracking). No restrictions were placed on the type of experiments conducted, whether in the laboratory, in the field or using modelling, if the bacteriophages were used in the context of understanding water transportation in ecosystems. All experiments using bacteriophages for other purposes, such as microbial source tracking, studies on the behaviour of bacteriophages in the environment or environmental pollution, were eliminated.
2.2. Data Extraction and Analysis
All selected manuscripts were downloaded and sorted using Mendeley v1.19.8 as the bibliographic reference manager software (http://www.mendeley.com, accessed on 7 November 2022). For each scientific paper selected, various data were extracted: the year of experiments, the country where the experiments were conducted, the bacteriophage species used, the environmental compartments investigated (e.g., groundwater, surface water), the environmental characteristics of the compartment investigated and the types of experiments performed (i.e., laboratory or field). In addition, for the most used bacteriophages, technical details of the experiments were also recorded where available: the initial concentrations of the bacteriophage solutions, the location of the injection point(s), the distance travelled by the injected bacteriophages between the injection site and downstream sampling site, the time and speed of bacteriophage transport, the parallel uses of other tracers and the percentage recovery of bacteriophages. All figures produced from bibliographic data were generated using the ‘ggplot2′ package with RStudio v1.4.1106 and R v4.0.4 [37].
3. Bacteriophages as Hydrological Tracers: An Overview
According to our eligibility criteria, thirty-two hydrological tracing experiments using bacteriophages were chosen. They were all conducted between 1968 and the present day (Figure 1A). Kinnunen and his team were the pioneers, with seven experiments performed between 1968 and 1976 [38]. Usually working with conventional tracers, they started working with bacteriophages to study the transportation of groundwater, surface water and wastewater (Figure 1B). At the same time, other researchers turned to the use of bacteriophages for their tracing experiments, albeit scarcely [39,40,41]. All types of water have been subject to experiments using bacteriophages, with groundwater and surface waters being the most investigated compartments. While only a small number of works were carried out in the 1980s [14,15,32,35], a significant increase in this type of experimentation was noticed in the late 1990s [42,43,44,45]. In the 2000s, tracing experiments using bacteriophage became less frequent to the point that they were no longer reported in 2015 [46]. In the 2000s until today, more and more scientific papers using bacteriophages for hydrological tracing purposes have turned to laboratory analyses with the aim of better understanding the fate of bacteriophages in the different compartments of the ecosystem. Indeed, as their properties were not well known at the time, their use as hydrological tracers could have been a limitation.
3.1. First Hydrological Tracing Experiments Using Bacteriophages
In October 1968, the first hydrological tracing experiments using the Escherichia coli bacteriophage F52 (or F52 for short) were performed by Kinnunen and Niemelä [38]. The experiment was the first of a series of tracing experiments conducted until 1975, where the objective was to study the water travel speed of a Kymikoki river (Finland), which extends from a power plant. During these experiments, four to five litres of F52 phage suspensions (1010 to 1011 plaque-forming unit [pfu]/mL) were injected directly into the river, where the main sewer is discharged, and then water was sampled at 17 km further down the stream. The bacteriophages travelled 8700 m in 7.7 h on average with a velocity ranging from 839 to 1872 m per hour, and an average discharge of 345 m3/s was determined. The objectives of this first experiment were not fully achieved, but the results were considered promising enough to warrant further experiments with bacteriophages.
3.2. Objectives of Hydrological Tracing Experiments Using Bacteriophages
Bacteriophages have been used primarily to study water flow velocity in various aquatic environments, such as surface waters (i.e., marine and fresh) [15,35,44] and groundwater [38,41,47,48], and secondly to investigate the connectivity between different aquatic compartments [44,49]. Paul et al. were able to determine the movement of domestic wastewater in the surface waters of the Florida reef using Enterobacteria phage PRD1 (Tectiviridae) and Listonella phage phiHSIC (Siphoviridae) bacteriophages. Overall, the use of bacteriophages has proven to be effective in hydrological tracing experiments, with a better detection sensitivity than conventional tracers, particularly in karst aquifers and saturated porous environments [42,45,48]. The use of bacteriophages was also efficient in studying water flow in fractured strata when used in conjunction with fluorescent dyes [47,49,50]. Despite the strong attenuation of the bacteriophage signal due to the rapid movement of groundwater, the potential of such an artificial tracer in tracing and identifying the water source has been demonstrated for short-term experiments [41,47,48]. In the application of wastewater tracing, bacteriophages have been used to explore potential deficiencies within sewage treatment plants [15,38,51,52]. Bacteriophage Escherichia virus T7 (Autographiviridae), used for leak detection in wastewater treatment systems, was found to be sensitive to changing conditions in sewage treatment plants and was therefore not a suitable tracer for this application. In contrast, the use of the bacteriophage F52 successfully detected the shortcut in a sewage lagoon and allowed for its repair [38]. In 1988, another sewer leak was successfully investigated using Emesvirus zideri (Fiersviridae), formerly known as the bacteriophage MS2 (Leviviridae), where the fluorescent dyes initially used to detect the leak had not given conclusive results [15]. All the compartments studied in such tracing experiments were aquatic environments, and no experiments have been conducted in the soil to potentially investigate the infiltration processes or the preferential water paths in soil pores.
4. Bacteriophages Commonly Used as Hydrological Tracers
4.1. Description of the Bacteriophages Used in the Tracing Experiments
In total, seventeen different bacteriophage strains have been used as hydrological tracers in our set of selected publications. The most commonly used bacteriophages belong to the families Fiersviridae, Siphoviridae and Autographiviridae (Figure 2A). At the species level, the two most widely used are Escherichia virus T7 (Autographiviridae) and the marine bacteriophage Pseudoalteromonas phage vB_PspS-H40/1 (H40/1 for short, Siphoviridae). In equal third place are Serratia marescens phages (unknown families), Emesvirus zideri (Fiersviridae) (former MS2, Leviviridae) and the bacteriophage F52 (unknown family) (Figure 2B). However, the bacteriophage F52 was specifically used in a single study conducted in surface water and wastewater in the very first series of tracing experiments [38].
The viral taxonomy built by the International Committee on Taxonomy of Viruses (ICTV) has recently been updated based on genetic similarities between bacteriophages, thus involving a reclassification of the entire order Caudovirales. For the bacteriophages of this review whose strain and family names have been changed, both the old and new names are mentioned. However, it should be noted that only the names employed in the articles have been retained for convenience.
The detailed characteristics of all bacteriophages used as hydrological tracers are summarised in Table 1. Briefly, most of them have a DNA genome with a head-tail structure, except PRD1, phiX174 and MS2, which all have a single capsid. In addition, MS2 is the only one with an RNA genome. Of all the bacteriophages used in the tracing experiments, MS2, T7, PRD1 and phiX174 are the best known and the most widely described in the literature (i.e., genome, structure, pI, host). In particular, the MS2 phage is used as a study model in many experiments in virology, and a large amount of information is therefore available on it [53,54,55]. F-specific RNA bacteriophages, including MS2, have also been proposed as a microbial source tracking tool or as indicators of faecal and viral pollution of waters [56,57,58], which has led to numerous studies on its behaviour and fate in aquatic environments [59,60]. In contrast, very little is known about the bacteriophages F52, F46 and F137 apart from their bacterial host Escherichia coli. This was isolated from a Finnish river and was used exclusively in the very first tracing experiments [38]. Similarly, for Serratia phages (Figure 2B), the strain was not specified in three of the five experiments reported, while the remaining bacteriophage, S24VA (NCIMB 10645), infecting the bacterial strain Serratia marcescens subsp. marcescens (NCIMB 10644), was used as a tracer. Up until 2020, 25 different genomes of Serratia phages had been described. All described Serratia phages are virulent bacteriophages, except for two of them, bacteriophages Eta and Parlo, which are temperate bacteriophages [61,62]. Little is known about the bacteriophages infecting Serratia marcescens, almost all of which were isolated from human and/or animal specimens (e.g., faeces) [63,64].
4.2. Effectiveness of Bacteriophages as Hydrological Tracers
All the characteristics regarding the transportation of bacteriophages observed during the various tracing experiments selected in this review are reported in Table 2. It should be noted that all ranges in this table have been purely reported from the literature taking into account the lowest and highest values recorded for each characteristic. The bacteriophage T7 was successfully used to study groundwater flow conditions within more or less permeable aquifers, over distances of hundreds of meters [36,42,45]. Marine bacteriophages are particularly appreciated when investigating freshwater flow since they do not display any background concentrations in this type of water. The bacteriophage H40/1 was used to study groundwater and surface water (i.e., lake and river) flow. It was noticed during surface water tracing experiments that both bacteriophages T7 and H40/1 were found to reach the sampling well faster or in a much lower concentration than the conventional tracer (i.e., uranine) [45,79]. It has been suggested that these processes may be due to the use of preferential pathways through the aquifers by the bacteriophages before reaching the sampling wells. While T7 and H40/1 were transported faster through porous materials, they were on the contrary found as being attenuated in lower permeable media, such as in clay and silt aquifers [42,45]. In karst aquifers, all marine bacteriophages were shown to follow shallow flow systems, while at the same time, the bacteriophage H6/1 displayed lower recovery rates than H40/1 due to a higher adsorption on clay [42,45]. In addition, a multi-tracer experiment using the three marine bacteriophages was recognised in an investigation of water circulation in lakes, especially to overcome the detection issue caused by the dilution [80]. Listonella phage ΦHSIC was successfully used from 1995 to 2000 to investigate the wastewater movement from a septic system to surface waters and groundwater. The bacteriophage ΦHSIC, isolated from marine waters, showed particularly high stability in marine subsurface water, where it travelled over long distances (hundreds of meters), and exhibited a notably higher stability than the bacteriophages MS2 and PRD1 that were used as complements [44,51].
While investigating groundwater flow, MS2 concentrations were found to be strongly attenuated, which could be explained by the adsorption of the bacteriophage onto the fracture walls or other material of the subsurface environments [44,46,48]. In experiments using PRD1 as a complement to MS2 to investigate water migration through fractured saprolite aquifer, PRD1 was detected over longer distances than MS2 (18 m vs. 13.5 m) [81]. Both bacteriophages remained viable for hundreds of days after injection, and PRD1 even reappeared in the sampling wells several months after injection, suggesting the remobilization of PRD1 after its adsorption to the clay particles (i.e., saprolite) in the aquifer. This adsorption–desorption process with geological media has indeed been demonstrated for PRD1, with a potential impact on its persistence during its transportation in aquifers, either by slowing it down or accelerating it [29]. Enterobacter cloacae phage showed similar behaviour to other bacteriophages in wetlands and aquifers, with a strong attenuation of its concentration in the aquifer, but better recovery rates than the chemical tracer (i.e., rhodamine WT) in the constructed wetlands. P22H5 also underwent adsorption and dilution events during its transportation through groundwater, despite its efficiency in determining channels within aquifers [50], while Aerobacter aerogenose 243 phages were successfully used without displaying adsorption on clay particles [41].
For the investigation of groundwater flow, the bacteriophage F137, like bromide, was shown to have successfully travelled within the whole groundwater system to determine the velocity of the water travel, while the other used tracers used (i.e., the bacteriophage T7, rhodamine and salts) were lost during transport. In addition, in contrast to the bacteriophage T7, the bacteriophage f1 was shown to be suitable for use in permeable aquifers [36]. Despite a higher attenuation of the bacteriophage phiX174 compared to dyes in a karstic aquifer, it showed less adsorption and more stability than the two other bacteriophages used in the same tracing experiment, Serratia phage and Enterobacter cloacae [47]. The bacteriophage Psf2 also showed its effectiveness when used to complement marine bacteriophages and the bacteriophage T7 and helped explore groundwater conduits [42]. However, the article did not specify the detailed results obtained after the tracing experiments, such as the velocity, the recovery rate or even the distance travelled. Finally, while determining the pathways into a porous limestone aquifer, Serratia phage managed to highlight the potential presence of fractures, notably because its concentration was found to be attenuated during transportation [47].
4.3. Combination of Hydrological Tracers in Multi-Injection Experiments
The specificity of bacteriophages for their bacterial hosts is characteristic that is well-known and appreciated by hydrologists since several bacteriophages can be used simultaneously without interfering with the detection of the others. For instance, MS2 phage, various strains of Serratia phage or even T7 phage are commonly deemed complementary to other tracers and can be used simultaneously with other bacteriophages [15,42,75] or conventional tracers (e.g., salts, fluorescein, rhodamine WT, microspheres) [33,38,46,80]. Experiments relying on the marine bacteriophage H40/1 always additionally included either other marine bacteriophages (i.e., H6/1, H4/4) [42,45,79], non-marine bacteriophages (i.e., T7, MS2) [42,45] or fluorescent dyes (i.e., uranine, fluorescein) [45,78].
Multi-injection experiments helped researchers compare the fate of multiple tracers and then put forward their best properties, depending on the objectives and the studied ecosystems. In the studies selected for this review, MS2, PRD1 and T7 were preferably used for short-scale experiments where the travelled distances ranged from 1 to 15 m, 3 to 100 m and up to 160 m [33,36,48,75]. Studies using MS2 demonstrated that its travel times varied from 1 h to several days in surface treatment wetlands and it was found to be no longer detectable after spending 17 h in the aquifers [44,75]. Other works compared MS2 to conventional tracers such as bromide, fluorescein and rhodamine WT. In most studies, MS2 phage reached the sampling stations with a delay compared to conventional tracers. The authors assumed that this behaviour was due to adsorption–desorption processes and concluded that this may hamper MS2-based estimations of water travel velocities [15,48]. Moreover, the recovered concentration of MS2 phage in bedrock fractures was ten times lower than that of PRD1, suggesting that narrower pathways were preferentially used by MS2 due to its smaller size (MS2—26 nm < PRD1—62 nm), and thus MS2 was not recovered in the sampling site. On the contrary, small bacteriophages (i.e., 20–30 nm diameter) have a velocity similar to that of water, better fitting the flow of water in fractured aquifers [48]. Moreover, when the permeability of the aquifer increases, the bacteriophage moves faster [36]. F137 and bromide had a similar velocity (0.125 m/h) and were the only tracers detectable during the entire experiment, while the signal of rhodamine, chloride and T7 were lost [38].
The contrasting performances of tracers were also noticeable in non-hydrological tracing experiments that were not reported in this review but might offer transferable explanations regarding the potential factors impacting these differences. In studies which aim at tracing microbial contamination or the fate of bacteriophages in water, marine bacteriophages (e.g., H40/1) have shown a higher transportation rate [81,82], a lower degree of dispersion [45,83] and a more sensitive detection limit than conventional tracers such as bromide.
5. Towards a More Adequate Use of Bacteriophages in Hydrological Tracing Experiments
In the selected studies, bacteriophages were valued by hydrologists in particular for their specificity toward their bacterial host, their eco-friendly property and the sensitivity of their detection methods. However, in regard to the ecological equilibrium of the ecosystem of interest, the eco-friendly property remains controversial, and the sensitivity of their detection methods requires further discussion from a virological point of view. In addition, the interaction between bacteriophages and the environment studied is not yet sufficiently understood. This important aspect needs to be further explored, even before the implementation of tracing experiments. The transportation of bacteriophages in the aquatic environment can be strongly influenced by the physicochemical characteristics of the ecosystem as well as their intrinsic properties (e.g., morphology, isoelectric point, size) [30]. Therefore, bacteriophages to be used as tracers must be properly selected according to the studied watershed to ensure their efficiency as hydrological tracers (Table 3).
5.1. Methodological Optimisation for Bacteriophage Detection
5.1.1. Limitations of the Current Detection Methods
In most studies, the detection and quantification of bacteriophages are carried out using the plaque assay. Briefly, the plaque assay is based on the incorporation of host bacterial cells in log-phase growth into agar medium. This creates a dense and turbid layer of bacteria able to sustain viral growth. A bacteriophage suspension can subsequently infect, replicate within and lyse cells. With each lysed cell, multiple adjacent ones become immediately infected by newly produced bacteriophages. After several cycles, a clear zone (a plaque) can be observed in the turbid plaque, indicating the presence of what was initially a single bacteriophage particle. The number of plaque-forming units per volume (i.e., PFU/mL) of a sample can thus be determined from the number of plaques generated on the agar layer [84,85,86]. The plaque assay is one of the easiest, fastest, most cost-effective and most reliable methods for the direct quantification of infectious bacteriophages and therefore the only detection method used in tracing experiments [87]. The sensitivity is appreciable, as one plaque can be detected in a sample volume of 1 to 5 mL [14,36,48,88]. However, only infectious bacteriophages can be detected. As previously mentioned, bacteriophages can be altered and/or inactivated during transportation in the environment, leaving the particles defective or even resulting in them becoming free nucleic acid. Defective particles are no longer infectious, which makes it impossible to detect them by plaque assay. Therefore, the disappearance of bacteriophages described in certain tracing experiments may be due to the use of an inappropriate detection method, rather than the actual loss of the tracer. Furthermore, by using such a methodological approach (plaque assay), only culturable bacteriophages for which the bacterial host is known and cultivable can be used as hydrological tracers for injection, which greatly limits the number of bacteriophages available.
5.1.2. The Interest in Using Several Complementary Methods for Bacteriophage Detection
To overcome the limitations of the plaque assay, the detection of the nucleic acid of a bacteriophage using molecular biology tools should also be considered (Figure 3). In this way, both infectious and non-infectious (damaged, inactivated and free DNA/RNA) bacteriophages can be detected simultaneously. Nowadays, molecular methods are well-developed, not excessively expensive (i.e., qPCR is about 5–10 €/sample) and widely implemented in all microbiology laboratories. A quantitative (real-time) polymerase chain reaction (qPCR) is a genomic amplification coupled with a fluorescent detection system. During the course of the DNA amplification, the fluorescence is detected and recorded in real-time [58,88,89,90]. In the case of RNA-viruses detection, a quantitative reverse transcription PCR (RT-qPCR) is used which transcribes RNA into complementary DNA (cDNA) by reverse transcriptase before carrying out the qPCR [91,92,93]. Beyond detection, such methods also allow for the quantification of the viral nucleic acids originally present in the analysed sample [94]. Moreover, using this molecular technique can promote multi-tracer experiments since the simultaneous detection of different bacteriophage species from an environmental sample is possible thanks to the high specificity of the primers and probes used in such an approach [95,96]. Among the articles selected in this review, only one tracing experiment involved the detection of bacteriophages (F-specific RNA bacteriophages) in stream water samples by applying RT-qPCR [46]. The detection method was successfully used to reveal several patterns such as the better performance of one bacteriophage strain (i.e., GA-phage), a dilution effect caused by precipitations and the loss of all bacteriophages, most likely due to a preferential pathway or bacteriophage behaviour (e.g., adsorption). Given the new information provided by molecular techniques, certain conclusions drawn from hydrological tracing experiments exclusively using the plaque assay would certainly be questionable. Indeed, the loss of bacteriophages described in these studies would most likely be the result of the specific detection of infectious bacteriophages, thus omitting the detection of the non-infectious ones. The two methods should therefore be considered complementary to each other to provide a complete picture of the concentration of bacteriophages recovered during a tracing experiment.
The detection of bacteriophages from water samples can be improved by including a viral concentration step before the analytical step. The purpose of this step is to artificially increase the concentration of the bacteriophage of interest in a water sample by reducing the volume of the sample. Thus, it allows for the detection of small amounts of bacteriophage which would not be quantifiable without the concentration step. The viral concentration of water samples can be achieved by using either adsorption/elution methods (e.g., electronegative or electropositive filters), ultrafiltration with polyethylene glycol (PEG) precipitation, centrifugation or centrifugal ultrafiltration, methods that are well-described in the vast number of reviews available [46,97,98,99].
5.2. “Eco-Friendly” Property of Bacteriophages
5.2.1. The Controversy of This Safe Biological Entity
The main advantage of using bacteriophages as hydrological tracers reported in the literature is their harmless effect on humans and environments since they specifically infect bacteria. Bacteriophages are considered safe for the environment, which has been proven by standard ecological toxicity tests [33,34,35,49]. Bacteriophages were therefore considered suitable for applications where non-polluting tracers were needed [41]. However, considering the ecosystem equilibrium, the claim that bacteriophages are environmentally friendly is partly flawed. Each tracing experiment requires the injection of large volumes (10–20 L) of highly concentrated bacteriophage solutions (104 to 1017 particles per litre) into the environment [33,45,48]. Although bacteriophages are natural biological entities, the introduction of a compound into an environment that is not their natural habitat can cause biological contamination by disturbing the equilibrium of ecosystems. Little is known about the impact of injected bacteriophages on existing bacterial communities, so it is reasonable to assume that these exogenous entities can disrupt the ecological diversity of bacteria [100,101]. Bacterial populations are naturally able to adapt to bacteriophage infection by developing resistance mechanisms mainly through genetic mutations but also through adaptative immunity or surface receptor modification [102,103]. The addition of exogenous bacteriophages in large volumes and concentrations can therefore increase either the mortality or the mutation rate of natural bacteria. The ecological functions of bacteria may also be impacted. Indeed, bacteriophages play a role in the biogeochemical cycles of bacteria, including the global carbon and nitrogen cycles [104,105]. Altering the natural bacterial community could cause disruptions or interruptions in biogeochemical cycles, which could impact nutrient availability (i.e., carbon, nitrogen) and generate faulty processes on a larger scale (i.e., the water cycle) [106,107].
One of the arguments often put forward to support the eco-friendly property of bacteriophages is their rapid elimination from the environment into which they have been introduced. However, although many environmental factors (e.g., temperature, radiation, pH) may be responsible for the inactivation (the loss of their ability to infect bacteria) of bacteriophages in the environment [30], their genetic material (DNA or RNA) can be released and persist for a long period in the environment, particularly their DNA, which is more stable than RNA (up to several years for the DNA and several days for the RNA). Some species of bacteria can perform a genetic transformation, meaning that they can directly take up and integrate free exogenous genetic material from their environment into the host genome through the cell membrane(s) [108,109,110]. Specifically, DNA may be involved in horizontal transfers of antibiotic resistance genes (ARGs), thus promoting the spread of antimicrobial resistance [111]. These transfers of ARGs were found to be significantly mediated by virulent or/and temperate bacteriophages through transduction mechanisms [112]. The transfer occurs either by the introduction of the bacteriophage DNA into the host genomes (temperate bacteriophages) or by pseudo-lysogeny, where the bacteriophage genome is integrated as a plasmid into the cytoplasm of the host bacteria (virulent bacteriophages) [113]. Mainly isolated from wastewater, the dissemination of these ARGs was also encountered in diverse environments, such as freshwater or urban rivers [114,115,116].
5.2.2. The Interest in the Natural Populations of Bacteriophages
The natural population of bacteriophages is large and extremely diverse in all environments. In soil, their concentration reputedly varies from 103 particles per gram of dry soil in cold deserts to 109 particles per gram of dry soil in grasslands and forests [117,118,119]. In aquatic environments, their concentration varies between 103 and 107 particles per millilitre of water depending on the type of water (i.e., groundwater, wastewater, marine and freshwater) [120,121]. Given this high diversity, natural populations of bacteriophages could be interesting potential candidates for hydrological tracing to overcome the limitations of the injection of exogenous bacteriophages, as has already been the case for diatoms [9,26,122]. Nowadays, advances in sequencing technologies and bioinformatics tools allow for the easier characterization of the viral diversity for a given habitat [123,124,125]. This characterization will allow for the determination of the most abundant species and/or genera that can subsequently be selected as potential hydrological tracers [9,126]. Similarly, by detecting and tracing specific sequences extracted from the genetic material of bacteriophages, the speed of the water as well as the preferential water pathways could be determined, especially in terrestrial compartments where bacteriophages are abundant [127].
Finally, bacteriophages introduced at the beginning of tracing experiments may be inactivated as soon as they enter the environment, whereas natural populations of bacteriophages have the advantage of already being adapted to the environment under study. However, for both natural and injected bacteriophages, their fate becomes uncertain once they have left the compartment of origin, and therefore the consideration of the environment of interest is also ideally required.
5.3. The Properties of the Catchment of Interest
5.3.1. Flaws in Considering the Surrounding Environments
Prior to any tracer injection, the characteristics of the ecosystems of interest and the environmental conditions have to be taken into account, as these aspects may be responsible for the loss of the bacteriophage signal during the experiment and its failure. Viral inactivation is the main phenomenon that can occur when bacteriophages are released into the environment, especially when the latter is not in its natural environment. Inactivation is defined as the loss of the ability of bacteriophages to infect host bacteria. It is then no longer possible to detect them in water samples using the plaque assay. Viral inactivation is highly dependent on environmental factors, such as temperatures, UV radiations and pH [128,129]. Temperature is the main factor in viral inactivation in the environment. In particular, high temperatures are responsible for the loss of the ability of bacteriophages to bind to their host cell through heat-induced changes in protein structure, as demonstrated for the MS2 bacteriophage [130]. These transformations can cause inactivation by disrupting the specific structures required for recognition and binding to host cells. Damage to the viral genome was described as negligible following heat treatment [131]. However, DNA bacteriophages, particularly φX174 and PRD1, were revealed as being highly resistant to a wide range of temperatures (>50 °C) and in various environmental matrices (e.g., surface water, seawater and wastewater) [132]. The optimal temperature for the survival of bacteriophages in the aquatic environment ranges between 4 and 10 °C [30,54,132,133]. A study led by Hodgson et al. (2002) highlighted differences in the bacteriophage travel time and recovery rate according to the season (i.e., winter season vs. summer season). During the cold season, the low light and the high river discharge allowed for a higher recovery rate of the bacteriophage MS2 (98%) than during the summer season (23%) [75]. The negative impact of temperature on MS2 bacteriophage detection was also reported by Gitis et al. (2011) and Cheng et al. (2006) for temperatures between 25 °C and 40 °C [134,135]. In the same vein, UV radiation also increases the inactivation rate of bacteriophages during warm seasons, especially in surface waters and wastewater [32,38]. UV radiation can cause substantial damage to viral nucleic acids, while modest protein damage occurred following a UV treatment on the MS2 bacteriophage [131,136,137,138,139]. Among solar radiations, UVB (i.e., 320 < λ < 400 nm) is the most inactivating radiation for bacteriophages [140]. In water loaded with particles such as river or seawater, water turbidity plays an indirect role, since, in turbid waters, radiation penetrates the water to a lesser extent, which limits the inactivation of the bacteriophage [35]. In both aquatic and terrestrial compartments, pH has a key role in the stability of bacteriophages [30,54,141,142]. In alkaline and acidic pH, bacteriophage inactivation was shown to be caused by the deformation, removal or denaturation of the protein capsid [54]. Regarding the bacteriophage T7, a continuous reduction in its concentration was noticed in porous aquifers when a simultaneous increase in the ionic strength and decrease in pH occurred, while the bacteriophage MS2 was found to be intact at a pH = 6.9 in quartz sandy soil columns [45].
In addition to inactivation, the signal of bacteriophages can also be lost during the tracing experiment due to adsorption phenomena. This is defined as the attachment of bacteriophages to other molecules, driven either by van der Waals electrostatic forces or hydrophobic interactions [143,144]. Their adsorption through electrostatic forces is influenced by the pH of the surrounding environment and by charges present on the surface of the molecule [145]. These interaction forces are reversible and mainly occur on mineral surfaces, such as saprolite [80,146] or ferric oxides [147,148]. Hydrophobic interactions are the aggregation of hydrophobic molecules in a hydrophilic media [143,149]. Bacteriophages display different degrees of hydrophobicity, where, for example, the bacteriophage Qb was found to be more hydrophobic than MS2, and, thus, bacteriophages will interact differently depending on their hydrophobicity [150]. This process was studied by Pieper et al. (1997) through the investigation of the effect of organic matter (OM) content on the adsorption of PRD1 during its transportation through sand and gravel aquifers contaminated by sewage. In contaminated aquifers, i.e., those with a high proportion of OM, the adsorption rate in terms of PRD1 on the aquifer particles was reduced, as organic matter was preferentially attached instead of PRD1. This hydrophobic adsorption is reversible and was described as a rapid attachment and slow detachment process [43,80,151]. The possible remobilization of PRD1 bacteriophages after adsorption on aquifer particles has been described. A high pH pulse caused a release of PRD1 into the water body during an experiment through the detachment of PRD1 from the aquifer surface [43,152]. Bacteriophages adsorb on surfaces when their respective electrical charges oppose each other. However, a change in the pH can cause charge repulsion, resulting in the detachment of bacteriophages. in 2013, Sadeghi et al. studied the effect of calcium ions (Ca2+) on PRD1 transport.
Finally, the loss of the bacteriophage signal can also be related to the intrinsic properties of bacteriophages (e.g., their size, structure and shape), especially in porous media aquifers (i.e., gravel and sand). Bacteriophages were shown to be more dispersed into the fracture or fissure zones through infiltration or percolation processes [45,153,154]. Smaller bacteriophages (<77 nm) are better transported due to their low collision efficiency [82,155]. For instance, H40/1, a small bacteriophage (45 nm diameter), was well distributed through an aquifer [45,148]. However, it was not only its size but also its shape that impacted on its transportation during tracing experiments. Indeed, short-tailed and spherical bacteriophages are adsorbed onto particles to a lesser extent than non-contractile-tailed and rod-shaped bacteriophages, due to their lower contact surface [82]. Not only does the morphology of bacteriophages have an impact on their transportation by causing their loss by adsorption onto particles but it can also be responsible for their inactivation. For instance, physical stresses and interactions with surfaces or particles can break certain structural parts of the viral particle, such as the tail of tailed bacteriophages, leading to their inactivation. In addition, bacteriophages with non-contractile tails are more susceptible to inactivation than those with flexible tails since the rigidity of the non-contractile tail makes it more susceptible to breaking [82,153,155].
5.3.2. Awareness of the Interactions between Bacteriophages and the Surrounding Environment
Whether the bacteriophage tracers used are injected or naturally present in an environment, a consideration of the properties of the environment in which they are used is essential to ensure relevant results from hydrological tracing experiments. As discussed above, the results of a hydrological tracing experiment can be strongly affected by the inactivation of the bacteriophages and their disappearance from the studied environment. As a reminder, the main factors to be controlled to limit the rate of inactivation of bacteriophages are temperature and solar radiation, while pH, size and shape will play an important role in the adsorption of bacteriophages onto soil or water particles, causing their avoidance of detection. Temperatures and solar radiation fluctuate with seasonality and are correlated with each other [156,157]. Surface water temperatures range from 0 °C in the coldest regions of the globe to 35 °C in the warmest, with a range of 9 °C to 24 °C in central Europe (data available daily at https://www.seatemperature.org/, accessed on 27 July 2022). The ground temperature was reported to be about 2°C higher than the air temperature, ranging from −19 °C to 37 °C [158,159]. In addition to seasonal variations, soil and water temperatures depend on depth, with temperatures decreasing with increasing depth [160,161]. The combination of cold weather and low solar radiation will therefore provide better environmental conditions for conducting a tracing experiment using bacteriophages. However, water and soil temperatures are not the main obstacles to tracing experiments, as most of the time tracing experiments are short, which limits the impact of these inactivation and disappearance phenomena to a certain extent.
Suspended sediments, colloidal particles and clay were all identified as being responsible for the disappearance of bacteriophages during hydrological tracing experiments as a result of the adsorption of the colloids on the surface of such environmental components [67,144,146]. To make the best use of bacteriophages, their hydrophobicity and electric charge should be determined before the experiment to predict the hydrophobic and electric interactions, respectively. In the case of H40/1, the bacteriophage was defined as being a moderately hydrophobic (contact angle ≈ 53–61°) and negatively charged (zeta potential = −22 to −24 mM) bacteriophage [82]. The bacteriophage H40/1 would appear to adsorb through hydrophobic interactions rather than using electrostatic forces. A study triggering changes in ionic forces, responsible for electrostatic interaction, did not reveal a reduction in the bacteriophage concentration during its transportation, suggesting that the bacteriophage did not adsorb [83]. PRD1 has a high hydrophobic surface due to its lipid-protein membrane located within the protein capsid [162], favouring hydrophobic interactions with the OM [163,164]. On the contrary, the adsorption of the MS2 bacteriophage on minerals is mostly driven by electrostatic forces. As explained previously, these forces are mainly driven by the environmental pH and the bacteriophage pI, particularly in soil. Each bacteriophage has its isoelectric point (pI). This is defined as the pH at which a virus carries no net electrical charge and is a characteristic parameter of the viral particle in equilibrium with the surrounding water chemistry. Knowing the pI of a virus enables the prediction of the virus surface charge in the environment. Virus pIs are found in a pH range from 1.9 to 8.4, where they are most frequently measured between 3.5 and 7 [165]. However, only a small number of pIs of viruses are known, and among the bacteriophages used as hydrological tracers, only those of MS2, PRD1, T7, PhiX174 and H6/1 have been measured (pI = 3.9, 4.0, 4.85, 6.6 and <4 respectively). Schematically, when the pH of the surrounding environment is inferior to the virus pI, the overall viral particle charge will be positive. At pH values above 7, the net surface charge of most viruses, including bacteriophages, is negative, resulting in attractive forces between the bacteriophages and certain positively charged particles of the environment [165,166]. The bacteriophage PhiX174, with a pI between 6.6 and 7, will therefore have weak positive charges on its surface, or no charges at all, at a pH of around 7 (roughly the pH of water), whereas the other bacteriophages, with pIs around 4, will be strongly positively charged, leading to a stronger attraction for negative charges. This process allows for the prevention of potential attachments to environmental matrices or molecules [129]. At a pH of above 4, Kvitsand et al. (2015) and Mayotte et al. (2017) demonstrated the adsorption of MS2 (negatively charged; pI = 3.9) on ferric-type surfaces with a net positive charge [60,147]. MS2 cannot only bind to free mineral and metal ions (i.e., Ca2+, Fe2+, Fe3+, Mn2+) in the medium but also those present on the clay surface [167,168,169,170]. According to Rossi and Aragno (1999), attapulgite caused 20% of T7 adsorption within 10 min, reaching 79% at the end of the experiment (total duration of experiments: 180 min). In another experiment, with a pI being almost equal to the pH of the environment, the bacteriophage T7 was reported to recognise both minerals (positive charges) and clay surface (negative charges), as it had both positive and negative charges on its surface [38,67]. Therefore, clay, particularly montmorillonite and attapulgite, induced the inactivation and concentration attenuation of the bacteriophage T7 sediment columns or colloidal particle batch [67,153]. However, in seawater and lake solutions seeded with Φ11M15, T2 and T7 in laboratory experiments, the inactivation of these three bacteriophages was lower for bacteriophages adsorbed on clay surfaces (specifically kaolinite) than for free bacteriophages [171,172,173]. Moreover, the bacteriophage T7 was still infectious after its adsorption on clay particles, which convinced the authors to assume a protective property of the clay against viral inactivation [67].
In the case of a study involving the injection of bacteriophages, it is important to consider the pI in the selection process of the candidate. If bacteriophages are inappropriately selected, the results of the tracing study may be inconclusive and unusable due to the possible loss of the viral tracer by adsorption to the environmental matrices. It is interesting to mention that during their transportation in the environment, bacteriophages can pass through various ecosystems with different pH values. These changes in environmental conditions during the journeys of bacteriophages should be investigated in advance of the tracing experiment to prevent the possible inactivation of both natural and artificial bacteriophages. It was reported that natural viral communities were less sensitive to UV radiations than lab strains [136]. However, as mentioned previously, it should be noted that the pI is not the only selection criterion regarding adsorption and that the shape or even the size of the virus also matters. As already explained, the tail makes bacteriophages sensitive to inactivation, especially when the tail is not contractile. Therefore, it would be interesting to consider using non-tailed bacteriophages, such as the bacteriophages MS2 or PhiX174, or a tail-head bacteriophage with a contractile tail, such as bacteriophage species belonging to the families Siphoviridae or Autographiviridae (e.g., H40/1 and T7, respectively) over non-contractile-tailed bacteriophages, such as species belonging to the family Myoviridae.
6. Conclusions and Perspectives
Although bacteriophages have been studied and used as hydrological tracers for almost sixty years, their real potential for this application remains poorly understood. They have been widely used in groundwater studies, but their relevance remains underexplored for hydrological investigations. Due to their small size, they successfully contributed to improving our understanding of subsurface water movements in karstic aquifers and in fractured saprolite [45,80,174], and they have finally emerged as good candidates for exploring hydrological processes. While their potential as hydrological tracers remains to be fully characterised, the combined use of bacteriophage and conventional tracers, such as isotopes, may lead to a better understanding of eco-hydro systems [38,42].
As each bacteriophage has its own behaviour, the transportation of each bacteriophage will be different depending on its intrinsic characteristics and the influence of external factors, which makes it impossible to find a tracer model to use in a specific type of ecosystem [45]. A bacteriophage’s size and shape serve as primary criteria. For fractured and fissured media, small bacteriophages will tend to move further in pores, which will reduce the collision efficiency. Adsorption is a common process and is most often encountered during bacteriophage transportation. Taking the hydrophobicity and the electric charge of bacteriophages into account will prevent a high rate of removal. To conclude, the choice of the right bacteriophage for a tracing experiment must be driven by an accurate pre-analysis of both the characteristics of the media and the intrinsic characteristics of bacteriophages.
The different advantages highlighted in the various articles in this review included the sensitivity of the detection method, the possibility to produce a high concentration of bacteriophages for a low volume and their specificity for their hosts. Indeed, bacteriophages were detected before conventional tracers, and they remained detected despite the dilution effect of the experiments [48,82]. Furthermore, several of them were used at the same time without interfering with the detection [35,81,153]. However, the initial high concentration of bacteriophages can be seen as a new type of pollution, as the introduction of an exogenous entity can generate a modification in the natural processes of a habitat. Moreover, the detection method is sensitive but limited to infectious bacteriophages. Complementary molecular techniques need to be considered to detect the damaged bacteriophages still occurring in tracing experiments. Finally, their transportation is influenced by different factors and, more specifically, their adsorption on the surrounding matrices, which has caused the attenuation of bacteriophages during tracing experiments. In addition, other environmental characteristics, such as pH, temperature or radiation, can affect their fate through the inactivation of bacteriophages [29,43,51]. Therefore, the local environment is important to consider when choosing the most appropriate bacteriophage [89].
It is important to note that the design of a tracing experiment using either one or multiple tracers does not differ from a traditional tracing experiment using conventional tracers nor from previous tracing experiments using bacteriophages. The recommendations therefore concern the selection of an appropriate candidate prior to the launch of any experiments in the case of using an artificial tracer. When using natural populations of bacteriophages as a more eco-friendly alternative, no injections are required and only the development of detection methods are needed. In addition, it is recommended to better evaluate the surrounding environment to prevent the loss of the signal of the tracer and to use complementary methods to optimize the obtained results.
Author Contributions
Conceptualization, P.F., L.O. and H.-M.C.; methodology, P.F. and L.O.; validation, P.F., L.O. and H.-M.C.; writing—original draft preparation, P.F.; writing—review and editing, P.F., L.O. and H.-M.C.; visualization, P.F.; supervision, L.O. and H.-M.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Luxembourg National Research Fund (FNR) for grant-ing and supporting the PRIDE program containing the Doctoral Training Unit Hydro-CSI project (PRIDE15/10623093—HYDRO-CSI).
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to acknowledge the Luxembourg National Research Fund (FNR) for its financial support of the Doctoral Training Unit Hydro-CSI (ref: PRIDE15/10623093—HYDRO-CSI). Moreover, the authors would like to thank Laurent Pfister (coordinator of the Doctoral Training Unit Hydro-CSI at the Luxembourg Institute of Science and Technology) for his comments on successive iterations of the manuscript and his expertise in hydrology.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses or interpretation of data; in the writing of the manuscript or in the decision to publish the results.
References
- Leibundgut, C.; Seibert, J. Tracer Hydrology. Treatise Water Sci. 2010, 2, 215–236. [Google Scholar] [CrossRef]
- Mcguire, K.J.; Mcdonnell, J.J. Tracer Advances in Catchment Hydrology. Hydrol. Process. 2015, 29, 5135–5138. [Google Scholar] [CrossRef]
- McGuire, K.J.; McDonnell, J.J. A Review and Evaluation of Catchment Transit Time Modeling. J. Hydrol. 2006, 330, 543–563. [Google Scholar] [CrossRef]
- Flury, M.; Wai, N.N. Dyes as Tracers for Vadose Zone Hydrology. Rev. Geophys. 2003, 41, 1–37. [Google Scholar] [CrossRef]
- Harvey, R.W. Microorganisms as Tracers in Groundwater Injection and Recovery Experiments: A Review. FEMS Microbiol. Rev. 1997, 20, 461–472. [Google Scholar] [CrossRef]
- Behrens, H.; Beims, U.; Dieter, H.; Dietze, G.; Eikmann, T.; Grummt, T.; Hanisch, H.; Henseling, H.; Käss, W.; Kerndorff, H.; et al. Toxicological and Ecotoxicological Assessment of Water Tracers. Hydrogeol. J. 2001, 9, 321–325. [Google Scholar] [CrossRef]
- Schudel, B.; Biaggi, D.; Dervey, T.; Kozel, R.; Müller, I.; Ross, J.H.; Schindler, U. Utilisation Des Traceurs Artificiels En Hydrogéologie—Guide Pratique; Série Géologique; Office fédéral des eaux et de la géologie: Washington, DC, USA, 2002; p. 77. [Google Scholar]
- Berryman, C.J. Tracer Tests for Investigating Flow and Transport in the Hyporheic Zone; Environment Agency: Bristol, UK, 2005; ISBN 9781844326983.
- Coles, A.E.; Wetzel, C.E.; Martínez-Carreras, N.; Ector, L.; Mcdonnell, J.J.; Frentress, J.; Klaus, J.; Hoffmann, L.; Pfister, L. Diatoms as a Tracer of Hydrological Connectivity: Are They Supply Limited? Ecohydrology 2016, 9, 631–645. [Google Scholar] [CrossRef]
- Kirchner, J.W.; Tetzlaff, D.; Soulsby, C. Comparing Chloride and Water Isotopes as Hydrological Tracers in Two Scottish Catchments. Hydrol. Process. 2010, 24, 1631–1645. [Google Scholar] [CrossRef]
- Klaus, J.; McDonnell, J.J. Hydrograph Separation Using Stable Isotopes: Review and Evaluation. J. Hydrol. 2013, 505, 47–64. [Google Scholar] [CrossRef]
- Burns, D.A. Stormflow-Hydrograph Separation Based on Isotopes: The Thrill Is Gone-What’s Next? Hydrol. Process. 2002, 16, 1515–1517. [Google Scholar] [CrossRef]
- Cartwright, I.; Cendón, D.; Currell, M.; Meredith, K. A Review of Radioactive Isotopes and Other Residence Time Tracers in Understanding Groundwater Recharge: Possibilities, Challenges, and Limitations. J. Hydrol. 2017, 555, 797–811. [Google Scholar] [CrossRef]
- Skilton, H.; Wheeler, D. Bacteriophage Tracer Experiments in Groundwater. J. Appl. Bacteriol. 1988, 65, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Martin, C. The Application of Bacteriophage Tracer Techniques in South West Water. Water Environ. J. 1988, 2, 638–642. [Google Scholar] [CrossRef]
- Sklash, M.G.; Farvolden, R.N.; Fritz, P. A Conceptual Model of Watershed Response to Rainfall, Developed through the Use of Oxygen-18 as a Natural Tracer. Can. J. Earth Sci. 1976, 13, 271–283. [Google Scholar] [CrossRef]
- McGlynn, B.; McDonnell, J.; Stewart, M.; Seibert, J. On the Relationships between Catchment Scale and Streamwater Mean Residence Time. Hydrol. Process. 2003, 17, 175–181. [Google Scholar] [CrossRef]
- Burt, T.P.; McDonnell, J.J. Whither Field Hydrology? The Need for Discovery Science and Outrageous Hydrological Hypotheses. Water Resour. Res. 2015, 51, 5919–5928. [Google Scholar] [CrossRef] [Green Version]
- Vitvar, T.; Aggarwal, P.K.; McDonnell, J.J. A Review of Isotope Applications in Catchment Hydrology. In Isotopes in the Water Cycle: Past, Present and Future of a Developing Science; Springer: Dordrecht, The Netherlands, 2005; pp. 151–169. ISBN 140203010X. [Google Scholar]
- Gibson, J.J.; Aggarwal, P.; Hogan, J.; Kendall, C.; Martinelli, L.A.; Stichler, W.; Rank, D.; Goni, I.; Choudhry, M.; Gat, J.; et al. Isotope Studies in Large River Basins: A New Global Research Focus. Eos Trans. Am. Geophys. Union 2002, 83, 613–620. [Google Scholar] [CrossRef]
- Selker, J.S.; Thévenaz, L.; Huwald, H.; Mallet, A.; Luxemburg, W.; Van De Giesen, N.; Stejskal, M.; Zeman, J.; Westhoff, M.; Parlange, M.B. Distributed Fiber-Optic Temperature Sensing for Hydrologic Systems. Water Resour. Res. 2006, 42, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Westhoff, M.C.; Savenjie, H.H.G.; Luxemburg, W.M.J.; Stelling, G.S.; Van De Giesen, N.C.; Selker, J.S.; Pfister, L.; Uhlenbrook, S. A Distributed Stream Temperature Model Using High Resolution Temperature Observations. Hydrol. Earth Syst. Sci. 2007, 11, 1469–1480. [Google Scholar] [CrossRef] [Green Version]
- Pfister, L.; McDonnell, J.J.; Hissler, C.; Hoffmann, L. Ground-Based Thermal Imagery as a Simple, Practical Tool for Mapping Saturated Area Connectivity and Dynamics. Hydrol. Process. 2010, 24, 3123–3132. [Google Scholar] [CrossRef]
- Glaser, B.; Antonelli, M.; Chini, M.; Pfister, L.; Klaus, J. Technical Note: Mapping Surface-Saturation Dynamics with Thermal Infrared Imagery. Hydrol. Earth Syst. Sci. 2018, 22, 5987–6003. [Google Scholar] [CrossRef] [Green Version]
- Pfister, L.; McDonnell, J.J.; Wrede, S.; Hlúbiková, D.; Matgen, P.; Fenicia, F.; Ector, L.; Hoffmann, L. The Rivers Are Alive: On the Potential for Diatoms as a Tracer of Water Source and Hydrological Connectivity. Hydrol. Process. 2009, 23, 2841–2845. [Google Scholar] [CrossRef]
- Klaus, J.; Wetzel, C.E.; Martínez-Carreras, N.; Ector, L.; Pfister, L. A Tracer to Bridge the Scales: On the Value of Diatoms for Tracing Fast Flow Path Connectivity from Headwaters to Meso-Scale Catchments. Hydrol. Process. 2015, 29, 5275–5289. [Google Scholar] [CrossRef]
- Foppen, J.W.; Orup, C.; Adell, R.; Poulalion, V.; Uhlenbrook, S. Using Multiple Artificial DNA Tracers in Hydrology. Hydrol. Process. 2011, 25, 3101–3106. [Google Scholar] [CrossRef]
- Tang, Y.; Foppen, J.W.; Bogaard, T.A. Transport of Silica Encapsulated DNA Microparticles in Controlled Instantaneous Injection Open Channel Experiments. J. Contam. Hydrol. 2021, 242, 103880. [Google Scholar] [CrossRef]
- Harvey, R.W.; Ryan, J.N. Use of PRD1 Bacteriophage in Groundwater Viral Transport, Inactivation, and Attachment Studies. FEMS Microbiol. Ecol. 2004, 49, 3–16. [Google Scholar] [CrossRef]
- Jończyk, E.; Kłak, M.; Miedzybrodzki, R.; Górski, A. The Influence of External Factors on Bacteriophages-Review. Folia Microbiol. 2011, 56, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Wommack, K.E.; Hill, R.T.; Kessel, M.; Russek-Cohen, E.; Colwell, R.R. Distribution of Viruses in the Chesapeake Bay. Appl. Environ. Microbiol. 1992, 58, 2965–2970. [Google Scholar] [CrossRef] [Green Version]
- Sinton, L.W.; Ching, S.B. An Evaluation of Two Bacteriophages as Sewage Tracers. Water Air Soil Pollut. 1987, 35, 347–356. [Google Scholar] [CrossRef]
- Harden, H.S.; Chanton, J.P.; Rose, J.B.; John, D.E.; Hooks, M.E. Comparison of Sulfur Hexafluoride, Fluorescein and Rhodamine Dyes and the Bacteriophage PRD-1 in Tracing Subsurface Flow. J. Hydrol. 2003, 277, 100–115. [Google Scholar] [CrossRef]
- Hodgson, C.J.; Perkins, J.; Labadz, J.C. Evaluation of Biotracers to Monitor Effluent Retention Time in Constructed Wetlands. Lett. Appl. Microbiol. 2003, 36, 362–371. [Google Scholar] [CrossRef]
- Drury, D.F.; Wheeler, D.C. Applications of a Serratia Marcescens Bacteriophage as a New Microbial Tracer of Aqueous Environments. J. Appl. Bacteriol. 1982, 53, 137–142. [Google Scholar] [CrossRef]
- Rossi, P.; de Carvalho-Dill, A.; Müller, I.; Aragno, M. Comparative Tracing Experiments in a Porous Aquifer Using Bacteriophages and Fluorescent Dye on a Test Field Located at Wilerwald (Switzerland) and Simultaneously Surveyed in Detail on a Local Scale by Radio-Magneto-Tellury (12–240 KHz). Environ. Geol. 1994, 23, 192–200. [Google Scholar] [CrossRef]
- Rstudio Team. RStudio: Integrated Development for R; RStudio, PBC.: Boston, MA, USA, 2020. [Google Scholar]
- Kinnunen, K. Tracing Water Movement by Means of Eschirichia Coli Bacteriophages; The water research institute: Helsinki, Finland, 1978; ISBN 951463344X. [Google Scholar]
- Wimpenny, J.W.T.; Cotton, N.; Statham, M. Microbes as Tracers of Water Movement. Water Res. 1972, 6, 731–739. [Google Scholar] [CrossRef]
- Schaub, S.A.; Sorber, C.A. Virus and Bacteria Removal from Wastewater by Rapid Infiltration through Soil. Appl. Environ. Microbiol. 1977, 33, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, R.; Thomas, A. An Example of the Use of Bacteriophage as a Groundwater Tracer. J. Hydrol. 1974, 23, 73–78. [Google Scholar] [CrossRef]
- Drew, D.; Doerfliger, N.; Formentin, K. The Use of Bacteriophages for Multi-Tracing in a Lowland Karst Aquifer in Western Ireland. Tracer Hydrol. 1997, 97, 33–37. [Google Scholar]
- Bales, R.C.; Li, S.; Yeh, T.J.; Lenczewski, M.E.; Gerba, P. Bacteriophage and Microsphere Transport in Saturated Porous Media: Forced-Gradient Experiment at Borden, Ontario. Water Resour. Res. 1997, 33, 639–648. [Google Scholar] [CrossRef] [Green Version]
- Paul, J.H.; Rose, J.B.; Jiang, S.C.; Zhou, X.; Cochran, P.; Kellogg, C.; Kang, J.B.; Griffin, D.; Farrah, S.; Lukasik, J. Evidence for Groundwater and Surface Marine Water Contamination by Waste Disposal Wells in the Florida Keys. Water Res. 1997, 31, 1448–1454. [Google Scholar] [CrossRef]
- Rossi, P.; Doerfliger, N.; Kennedy, K.; Müller, I.; Aragno, M. Bacteriophages as Surface and Ground Water Tracers. Hydrol. Earth Syst. Sci. 1998, 2, 101–110. [Google Scholar] [CrossRef]
- Antonelli, M.; Balasubramanian, M.N.; Ogorzaly, L.; Pfister, L. QPCR (Quantitative Polymerase Chain Reaction) for the Quantification of Bacteriophages in Stream Water Samples to Investigate Hydrological Processes: A Proof-of-Concept Study in the Huewelerbach Experimental Catchment (Luxembourg). In Proceedings of the EGU General assembly, Vienna, Austria, 17–22 April 2016; Volume 18, p. 1. [Google Scholar]
- Maurice, L.; Atkinson, T.C.; Williams, A.T.; Barker, J.A.; Farrant, A.R. Catchment Scale Tracer Testing from Karstic Features in a Porous Limestone. J. Hydrol. 2010, 389, 31–41. [Google Scholar] [CrossRef]
- McKay, L.D.; Cherry, J.A.; Bales, R.C.; Yahya, M.T.; Gerba, C.P. A Field Example of Bacteriophage as Tracers of Fracture Flow. Environ. Sci. Technol. 1993, 27, 1075–1079. [Google Scholar] [CrossRef]
- Bricelj, M. Microbial Tracers in Groundwater Research. RMZ—Mater. Geoenviron. 2003, 1, 67–70. [Google Scholar]
- Curk, B.C.; Stichler, W. Deuterium Tracer Experiment in the Unsaturated Zone of Fractured Karst Aquifers. In Proceedings of the International Symposium On Advances In Isotope Hydrology, Vienna, Austria, 21–25 May 2007; Volume 1, pp. 597–604. [Google Scholar]
- Paul, J.H.; McLaughlin, M.R.; Griffin, D.W.; Lipp, E.K.; Stokes, R.; Rose, J.B. Rapid Movement of Wastewater from On-Site Disposal Systems into Surface Waters in the Lower Florida Keys. Estuaries Coasts 2000, 23, 662–668. [Google Scholar] [CrossRef]
- Horan, N.J.; Parr, J.; Naylor, P.J. Evaluation of Tracers for the Determination of the Mixing Characteristics of Activated Sludge Reactors. Environ. Technol. 1991, 12, 603–608. [Google Scholar] [CrossRef]
- Langlet, J.; Gaboriaud, F.; Gantzer, C. Effects of PH on Plaque Forming Unit Counts and Aggregation of MS2 Bacteriophage. J. Appl. Microbiol. 2007, 103, 1632–1638. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Y.; Ong, S.L.; Hu, J.Y.; Tan, X.L.; Ng, W.J. Effects of PH and Temperature on the Survival of Coliphages MS2 and Qβ. J. Ind. Microbiol. Biotechnol. 2003, 30, 549–552. [Google Scholar] [CrossRef]
- Elhadidy, A.M.; Peldszus, S.; van Dyke, M.I. An Evaluation of Virus Removal Mechanisms by Ultrafiltration Membranes Using MS2 and ΦX174 Bacteriophage. Sep. Purif. Technol. 2013, 120, 215–223. [Google Scholar] [CrossRef]
- Ogorzaly, L.; Tissier, A.; Bertrand, I.; Maul, A.; Gantzer, C. Relationship between F-Specific RNA Phage Genogroups, Faecal Pollution Indicators and Human Adenoviruses in River Water. Water Res. 2009, 43, 1257–1264. [Google Scholar] [CrossRef]
- Ogorzaly, L.; Bertrand, I.; Paris, M.; Maul, A.; Gantzer, C. Occurrence, Survival, and Persistence of Human Adenoviruses and F-Specific RNA Phages in Raw Groundwater. Appl. Environ. Microbiol. 2010, 76, 8019–8025. [Google Scholar] [CrossRef] [Green Version]
- Ogorzaly, L.; Gantzer, C. Development of Real-Time RT-PCR Methods for Specific Detection of F-Specific RNA Bacteriophage Genogroups: Application to Urban Raw Wastewater. J. Virol. Methods 2006, 138, 131–139. [Google Scholar] [CrossRef]
- Jain, R.; Srivastava, R. Metabolic Investigation of Host/Pathogen Interaction Using MS2-Infected Escherichia Coli. BMC Syst. Biol. 2009, 3, 121. [Google Scholar] [CrossRef] [PubMed]
- Mayotte, J.M.; Hölting, L.; Bishop, K. Reduced Removal of Bacteriophage MS2 in during Basin Infiltration Managed Aquifer Recharge as Basin Sand Is Exposed to Infiltration Water. Hydrol. Process. 2017, 31, 1690–1701. [Google Scholar] [CrossRef]
- Bockoven, R.; Gutierrez, J.; Newkirk, H.; Liu, M.; Cahill, J.; Ramsey, J. Complete Genome Sequence of Serratia Marcescens Podophage Parlo. Microbiol. Resour. Announc. 2019, 8, e00569-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denyes, J.M.; Krell, P.J.; Manderville, R.A.; Ackermann, H.W.; She, Y.M.; Kropinski, A.M. The Genome and Proteome of Serratia Bacteriophage η Which Forms Unstable Lysogens. Virol. J. 2014, 11, 6. [Google Scholar] [CrossRef] [Green Version]
- Bujak, K.; Decewicz, P.; Kaminski, J.; Radlinska, M. Identification, Characterization, and Genomic Analysis of Novel Serratia Temperate Phages from a Gold Mine. Int. J. Mol. Sci. 2020, 21, 6709. [Google Scholar] [CrossRef]
- Tian, C.; Zhao, J.; Zhang, Z.; Chen, X.; Wei, X.; Li, H.; Lin, W.; Ke, Y.; Hu, L.; Jiang, A.; et al. Identification and Molecular Characterization of Serratia Marcescens Phages VB_SmaA_2050H1 and VB_SmaM_2050HW. Arch. Virol. 2019, 164, 1085–1094. [Google Scholar] [CrossRef]
- Chan, L.Y.; Kosuri, S.; Endy, D. Refactoring Bacteriophage T7. Mol. Syst. Biol. 2005, 1, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Huss, P.; Meger, A.; Leander, M.; Nishikawa, K.; Raman, S. Mapping the Functional Landscape of the Receptor Binding Domain of T7 Bacteriophage by Deep Mutational Scanning. Elife 2021, 10, e63775. [Google Scholar] [CrossRef]
- Rossi, P.; Aragno, M. Analysis of Bacteriophage Inactivation and Its Attenuation by Adsorption onto Colloidal Particles by Batch Agitation Techniques. Can. J. Microbiol. 1999, 45, 9–17. [Google Scholar] [CrossRef]
- Kallies, R.; Kiesel, B.; Zopfi, J.; Wick, L.Y.; Chatzinotas, A. Complete Genome Sequence of Alteromonas Virus VB_AspP-H4/4. Genome Announc. 2017, 5, e00914-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dedeo, M.T.; Finley, D.T.; Francis, M.B. Viral Capsids as Self-Assembling Templates for New Materials, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; Volume 103, ISBN 9780124159068. [Google Scholar]
- Carr, E.L.; Wilson, M.E.; Adams, S.T.; Arens, D.K.; Ayala, M.; Ayers, H.; Barker, A.; Beecroft, V.; Bishop, E.; Brundage, B.; et al. Genome Sequences of 14 Siphophages That Infect Serratia Marcescens. Microbiol. Resour. Announc. 2022, 11, e0121221. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.C.; Kellogg, C.A.; Paul, J.H. Characterization of Marine Temperate Phage-Host Systems Isolated from Mamala Bay, Oahu, Hawaii. Appl. Environ. Microbiol. 1998, 64, 535–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kallies, R.; Kiesel, B.; Schmidt, M.; Ghanem, N.; Zopfi, J.; Hackermüller, J.; Harms, H.; Wick, L.Y.; Chatzinotas, A. Complete Genome Sequence of Pseudoalteromonas Virus VB_PspP-H6/1 That Infects Pseudoalteromonas Sp. Strain H6. Mar. Genom. 2019, 47, 100667. [Google Scholar] [CrossRef]
- Olsen, R.H.; Siak, J.-S.; Gray, R.H. Characteristics of PRD1, a Plasmid-Dependent Broad Host Range DNA Bacteriophage. J. Virol 1974, 14, 689–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziedaite, G.; Daugelavic, R.; Bamford, J.K.H.; Bamford, D.H. The Holin Protein of Bacteriophage PRD1 Forms a Pore for Small-Molecule and Endolysin Translocation. J. Bacteriol. 2005, 187, 5397–5405. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, C.J.; Perkins, J.; Labadz, J.C. The Use of Microbial Tracers to Monitor Seasonal Variations in Effluent Retention in a Constructed Wetland. Water Res. 2004, 38, 3833–3844. [Google Scholar] [CrossRef]
- Morozova, V.; Jdeed, G.; Kozlova, Y.; Babkin, I.; Tikunov, A.; Tikunova, N. A New Enterobacter Cloacae Bacteriophage Ec151 Encodes the Deazaguanine Dna Modification Pathway and Represents a New Genus within the Siphoviridae Family. Viruses 2021, 13, 1372. [Google Scholar] [CrossRef]
- McKenna, R.; Xia, D.; Willingmann, P.; Ilag, L.L.; Rossmann, M.G. Structure Determination of the Bacteriophage PhiX174. Acta Cryst. B 1992, 48, 499–511. [Google Scholar] [CrossRef]
- Schnegg, P.-A.; Flynn, R. Online Field Fluorometers for Hydrogeological Tracer Tests. Isot. Und Tracer Der Wasserforsch. 2002, 1879, 29–36. [Google Scholar]
- Goldscheider, N.; Haller, L.; Poté, J.; Wildi, W.; Zopfi, J. Characterizing Water Circulation and Contaminant Transport in Lake Geneva Using Bacteriophage Tracer Experiments and Limnological Methods. Environ. Sci. Technol. 2007, 41, 5252–5258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay, L.D.; Sanford, W.E.; Strong, J.M. Field-Scale Migration of Colloidal Tracers in a Fractured Shale Saprolite. Ground Water 2000, 38, 139–147. [Google Scholar] [CrossRef]
- Flynn, R.; Hacini, Y.; Schnegg, P.-A.; Costa, R.; Diomande, K.A. Use of Tracer Tests and Geophysical Logging to Understand Solute and Micro-Organism Tracer Responses in Monitoring Wells with Long Screen Intervals in a Gravel Aquifer. Beiträge Zur. Hydrogeol. 2006, 55, 5–20. [Google Scholar]
- Ghanem, N.; Kiesel, B.; Kallies, R.; Harms, H.; Chatzinotas, A.; Wick, L.Y. Marine Phages as Tracers: Effects of Size, Morphology, and Physico− Chemical Surface Properties on Transport in a Porous Medium. Environ. Sci. Technol. 2016, 50, 12816–12824. [Google Scholar] [CrossRef] [PubMed]
- Flynn, R.M.; Sinreich, M. Characterisation of Virus Transport and Attenuation in Epikarst Using Short Pulse and Prolonged Injection Multi-Tracer Testing. Water Res. 2010, 44, 1138–1149. [Google Scholar] [CrossRef] [PubMed]
- Cooper, P.D. The Plaque Assay of Animal Viruses. Adv. Virus Res. 1961, 8, 319–378. [Google Scholar] [CrossRef]
- Dulbecco, R.; Vogt, M. Some Problems of Animal Virology as Studied by the Plaque Technique. Cold Spring Harb. Symp. Quant. Biol. 1953, 18, 273–279. [Google Scholar] [CrossRef]
- Roux, M. On an Invisible Microbe Antagonistic to Dysentery Bacilli. Note by M. F. d’Herelle, Presented by M. Roux. Comptes Rendus Acad. Des Sci. 1917, 165, 373–375, Erratum in Bacteriophage 2011, 1, 3–5.. [Google Scholar] [CrossRef] [Green Version]
- Shen, C.; Phanikumar, M.S.; Fong, T.T.; Aslam, I.; Mcelmurry, S.P.; Molloy, S.L.; Rose, J.B. Evaluating Bacteriophage P22 as a Tracer in a Complex Surface Water System: The Grand River, Michigan. Environ. Sci. Technol. 2008, 42, 2426–2431. [Google Scholar] [CrossRef]
- Wyer, M.D.; Kay, D.; Watkins, J.; Davies, C.; Kay, C.; Thomas, R.; Porter, J.; Stapleton, C.M.; Moore, H. Evaluating Short-Term Changes in Recreational Water Quality during a Hydrograph Event Using a Combination of Microbial Tracers, Environmental Microbiology, Microbial Source Tracking and Hydrological Techniques: A Case Study in Southwest Wales, UK. Water Res. 2010, 44, 4783–4795. [Google Scholar] [CrossRef]
- Hunt, R.J.; Borchardt, M.A.; Bradbury, K.R. Viruses as Groundwater Tracers: Using Ecohydrology to Characterize Short Travel Times in Aquifers. Groundwater 2014, 52, 187–193. [Google Scholar] [CrossRef]
- Ács, N.; Gambino, M.; Brøndsted, L. Bacteriophage Enumeration and Detection Methods. Front. Microbiol. 2020, 11, 594868. [Google Scholar] [CrossRef]
- Watzinger, F.; Suda, M.; Preuner, S.; Baumgartinger, R.; Ebner, K.; Baskova, L.; Niesters, H.G.M.; Lawitschka, A.; Lion, T. Real-Time Quantitative PCR Assays for Detection and Monitoring of Pathogenic Human Viruses in Immunosuppressed Pediatric Patients. J. Clin. Microbiol. 2004, 42, 5189. [Google Scholar] [CrossRef] [Green Version]
- Toussaint, J.F.; Sailleau, C.; Breard, E.; Zientara, S.; De Clercq, K. Bluetongue Virus Detection by Two Real-Time RT-QPCRs Targeting Two Different Genomic Segments. J. Virol. Methods 2007, 140, 115–123. [Google Scholar] [CrossRef]
- Vester, D.; Lagoda, A.; Hoffmann, D.; Seitz, C.; Heldt, S.; Bettenbrock, K.; Genzel, Y.; Reichl, U. Real-Time RT-QPCR Assay for the Analysis of Human Influenza A Virus Transcription and Replication Dynamics. J. Virol. Methods 2010, 168, 63–71. [Google Scholar] [CrossRef]
- Arya, M.; Shergill, I.S.; Williamson, M.; Gommersall, L.; Arya, N.; Patel, H.R.H. Basic Principles of Real-Time Quantitative PCR. Expert Rev. Mol. Diagn. 2005, 5, 209–219. [Google Scholar] [CrossRef]
- Klymus, K.E.; Ruiz Ramos, D.V.; Thompson, N.L.; Richter, C.A.; Ramos, R.; Development, C. Development and Testing of Species-Specific Quantitative PCR Assays for Environmental DNA Applications. J. Vis. Exp. 2020, 165, e61825. [Google Scholar] [CrossRef]
- Knudsen, S.W.; Ebert, R.B.; Hesselsøe, M.; Kuntke, F.; Hassingboe, J.; Mortensen, P.B.; Thomsen, P.F.; Sigsgaard, E.E.; Hansen, B.K.; Nielsen, E.E.; et al. Species-Specific Detection and Quantification of Environmental DNA from Marine Fishes in the Baltic Sea. J. Exp. Mar. Biol. Ecol. 2019, 510, 31–45. [Google Scholar] [CrossRef]
- Bofill-Mas, S.; Rusiñol, M. Recent Trends on Methods for the Concentration of Viruses from Water Samples. Curr. Opin. Environ. Sci. Health 2020, 16, 7–13. [Google Scholar] [CrossRef]
- Forés, E.; Rusiñol, M.; Itarte, M.; Martínez-Puchol, S.; Calvo, M.; Bofill-Mas, S. Evaluation of a Virus Concentration Method Based on Ultrafiltration and Wet Foam Elution for Studying Viruses from Large-Volume Water Samples. Sci. Total Environ. 2022, 829, 154431. [Google Scholar] [CrossRef]
- Cashdollar, J.L.; Wymer, L. Methods for Primary Concentration of Viruses from Water Samples: A Review and Meta-Analysis of Recent Studies. J. Appl. Microbiol. 2013, 115, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Chevallereau, A.; Pons, B.J.; van Houte, S.; Westra, E.R. Interactions between Bacterial and Phage Communities in Natural Environments. Nat. Rev. Microbiol. 2022, 20, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Maslov, S.; Sneppen, K. Population Cycles and Species Diversity in Dynamic Kill-the-Winner Model of Microbial Ecosystems. Sci. Rep. 2017, 7, 39642. [Google Scholar] [CrossRef]
- Oechslin, F. Resistance Development to Bacteriophages Occurring during Bacteriophage Therapy. Viruses 2018, 10, 351. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Vos, M. Adaptation in Natural Microbial Populations. Annu Rev. Ecol. Evol. Syst. 2015, 46, 503–522. [Google Scholar] [CrossRef] [Green Version]
- Weitz, J.S.; Wilhelm, S.W. Ocean Viruses and Their Effects on Microbial Communities and Biogeochemical Cycles. F1000 Biol. Rep. 2012, 4, 2–9. [Google Scholar] [CrossRef] [Green Version]
- Clokie, M.R.J.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in Nature. Bacteriophage 2011, 1, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Braga, L.P.P.; Spor, A.; Kot, W.; Breuil, M.C.; Hansen, L.H.; Setubal, J.C.; Philippot, L. Impact of Phages on Soil Bacterial Communities and Nitrogen Availability under Different Assembly Scenarios. Microbiome 2020, 8, 52. [Google Scholar] [CrossRef] [Green Version]
- Roux, S.; Brum, J.R.; Dutilh, B.E.; Sunagawa, S.; Duhaime, M.B.; Loy, A.; Poulos, B.T.; Solonenko, N.; Lara, E.; Poulain, J.; et al. Ecogenomics and Potential Biogeochemical Impacts of Globally Abundant Ocean Viruses. Nature 2016, 537, 689–693. [Google Scholar] [CrossRef] [Green Version]
- Chen, I.; Dubnau, D. DNA Uptake during Bacterial Transformation. Nat. Rev. Microbiol. 2004, 2, 241–249. [Google Scholar] [CrossRef]
- Johnston, C.; Martin, B.; Fichant, G.; Polard, P.; Claverys, J.P. Bacterial Transformation: Distribution, Shared Mechanisms and Divergent Control. Nat. Rev. Microbiol. 2014, 12, 181–196. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, Z.; Sun, M.; Shi, Y. Dynamics, Gene Transfer, and Ecological Function of Intracellular and Extracellular DNA in Environmental Microbiome. iMeta 2022, 1, e34. [Google Scholar] [CrossRef]
- Woegerbauer, M.; Bellanger, X.; Merlin, C. Cell-Free DNA: An Underestimated Source of Antibiotic Resistance Gene Dissemination at the Interface Between Human Activities and Downstream Environments in the Context of Wastewater Reuse. Front. Microbiol. 2020, 11, 671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muniesa, M.; Colomer-Lluch, M.; Jofre, J. Could Bacteriophages Transfer Antibiotic Resistance Genes from Environmental Bacteria to Human-Body Associated Bacterial Populations? Mob. Genet. Elem. 2013, 3, 739–751. [Google Scholar] [CrossRef]
- Colavecchio, A.; Cadieux, B.; Lo, A.; Goodridge, L.D. Bacteriophages Contribute to the Spread of Antibiotic Resistance Genes among Foodborne Pathogens of the Enterobacteriaceae Family—A Review. Front. Microbiol. 2017, 8, 1108. [Google Scholar] [CrossRef] [Green Version]
- Sala-Comorera, L.; Nolan, T.M.; Reynolds, L.J.; Venkatesh, A.; Cheung, L.; Martin, N.A.; Stephens, J.H.; Gitto, A.; O’Hare, G.M.P.; O’Sullivan, J.J.; et al. Bacterial and Bacteriophage Antibiotic Resistance in Marine Bathing Waters in Relation to Rivers and Urban Streams. Front. Microbiol. 2021, 12, 2017. [Google Scholar] [CrossRef]
- Zare, S.; Derakhshandeh, A.; Mohammadi, A.; Noshadi, M. Abundance of Antibiotic Resistance Genes in Bacteria and Bacteriophages Isolated from Wastewater in Shiraz. Mol. Biol. Res. Commun. 2021, 10, 73–83. [Google Scholar] [CrossRef]
- Moon, K.; Jeon, J.H.; Kang, I.; Park, K.S.; Lee, K.; Cha, C.J.; Lee, S.H.; Cho, J.C. Freshwater Viral Metagenome Reveals Novel and Functional Phage-Borne Antibiotic Resistance Genes. Microbiome 2020, 8, 75. [Google Scholar] [CrossRef]
- Sinke, L. Viruses in Soils: What Do We Know and Where Do We Go from Here ? Available online: https://www.labroots.com/webinar/viruses-soils-know-here (accessed on 24 June 2019).
- Williamson, K.E.; Fuhrmann, J.J.; Wommack, K.E.; Radosevich, M. Viruses in Soil Ecosystems: An Unknown Quantity Within an Unexplored Territory. Annu. Rev. Virol. 2017, 4, 201–219. [Google Scholar] [CrossRef]
- Florent, P.; Cauchie, H.-M.; Herold, M.; Jacquet, S.; Ogorzaly, L. Soil PH, Calcium Content and Bacteria as Major Factors Responsible for the Distribution of the Known Fraction of the DNA Bacteriophage Populations in Soils of Luxembourg. Microorganisms 2022, 10, 1458. [Google Scholar] [CrossRef]
- Børsheim, K.Y. Native Marine Bacteriophages. FEMS Microbiol. Ecol. 1993, 102, 141–159. [Google Scholar] [CrossRef]
- Bergh, O.; Børsheim, K.Y.; Bratbak, G.; Heldal, M. High Abundance of Viruses Found in Aquatic Environments. Nature 1989, 340, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Carreras, N.; Wetzel, C.E.; Frentress, J.; Ector, L.; McDonnell, J.J.; Hoffmann, L.; Pfister, L. Hydrological Connectivity Inferred from Diatom Transport through the Riparian-Stream System. Hydrol. Earth Syst. Sci. 2015, 19, 3133–3151. [Google Scholar] [CrossRef] [Green Version]
- Kieft, K.; Zhou, Z.; Anantharaman, K. VIBRANT: Automated Recovery, Annotation and Curation of Microbial Viruses, and Evaluation of Viral Community Function from Genomic Sequences. Microbiome 2020, 8, 90. [Google Scholar] [CrossRef] [PubMed]
- Roux, S.; Enault, F.; Hurwitz, B.L.; Sullivan, M.B. VirSorter: Mining Viral Signal from Microbial Genomic Data. PeerJ 2015, 2015, e985. [Google Scholar] [CrossRef]
- Trubl, G.; Hyman, P.; Roux, S.; Abedon, S.T. Coming-of-Age Characterization of Soil Viruses: A User’s Guide to Virus Isolation, Detection within Metagenomes, and Viromics. Soil Syst. 2020, 4, 23. [Google Scholar] [CrossRef] [Green Version]
- Mächler, E.; Salyani, A.; Walser, J.-C.; Larsen, A.; Schaefli, B.; Altermatt, F.; Ceperley, N. Environmental DNA Simultaneously Informs Hydrological and Biodiversity Characterization of an Alpine Catchment. Hydrol. Earth Syst. Sci. 2021, 25, 735–753. [Google Scholar] [CrossRef]
- Peters, J. Fate and Transport of Synthetic DNA in Surface Water; University of Waterloo: Waterloo, ON, Canada, 2021. [Google Scholar]
- Middelboe, M.; Jacquet, S.; Weinbauer, M. Viruses in Freshwater Ecosystems: An Introduction to the Exploration of Viruses in New Aquatic Habitats. Freshw. Biol. 2008, 53, 1069–1075. [Google Scholar] [CrossRef]
- Hurst, C.J.; Gerba, C.P.; Cech, I. Effects of Environmental Variables and Soil Characteristics on Virus Survival in Soil. Appl. Environ. Microbiol. 1980, 40, 1067–1079. [Google Scholar] [CrossRef] [Green Version]
- Drulis-Kawa, Z.; Majkowska-Skrobek, G.; Maciejewska, B. Bacteriophages and Phage-Derived Proteins—Application Approaches. Curr. Med. Chem. 2015, 22, 1757. [Google Scholar] [CrossRef]
- Wigginton, K.R.; Pecson, B.M.; Sigstam, T.; Bosshard, F.; Kohn, T. Virus Inactivation Mechanisms: Impact of Disinfectants on Virus Function and Structural Integrity. Environ. Sci. Technol. 2012, 46, 12069–12078. [Google Scholar] [CrossRef] [PubMed]
- Bertrand, I.; Schijven, J.F.; Sánchez, G.; Wyn-Jones, P.; Ottoson, J.; Morin, T.; Muscillo, M.; Verani, M.; Nasser, A.; de Roda Husman, A.M.; et al. The Impact of Temperature on the Inactivation of Enteric Viruses in Food and Water: A Review. J. Appl. Microbiol. 2012, 112, 1059–1074. [Google Scholar] [CrossRef] [PubMed]
- Fauvel, B.; Ogorzaly, L.; Cauchie, H.M.; Gantzer, C. Interactions of Infectious F-Specific RNA Bacteriophages with Suspended Matter and Sediment: Towards an Understanding of FRNAPH Distribution in a River Water System. Sci. Total Environ. 2017, 574, 960–968. [Google Scholar] [CrossRef] [Green Version]
- Gitis, V.; Dlugy, C.; Gun, J.; Lev, O. Studies of Inactivation, Retardation and Accumulation of Viruses in Porous Media by a Combination of Dye Labeled and Native Bacteriophage Probes. J. Contam. Hydrol. 2011, 124, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Chetochine, A.S.; Pepper, I.L.; Brusseau, M.L. Influence of DOC on MS-2 Bacteriophage Transport in a Sandy Soil. Water Air Soil Pollut. 2007, 178, 315–322. [Google Scholar] [CrossRef]
- Weinbauer, M.G.; Wilhelm, S.W.; Suttle, C.A.; Pledger, R.J.; Mitchell, D.L. Sunlight-Induced DNA Damage and Resistance in Natural Viral Communities. Aquat. Microb. Ecol. 1999, 17, 111–120. [Google Scholar] [CrossRef]
- Beck, S.E.; Rodriguez, R.A.; Hawkins, M.A.; Hargy, T.M.; Larason, T.C.; Linden, K.G. Comparison of UV-Induced Inactivation and RNA Damage in MS2 Phage across the Germicidal UV Spectrum. Appl. Environ. Microbiol. 2016, 82, 1468. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.I.; Jin, S.G.; Pfeifer, G.P. Formation of Cyclobutane Pyrimidine Dimers at Dipyrimidines Containing 5-Hydroxymethylcytosine. Photochem. Photobiol. Sci. 2013, 12, 1409. [Google Scholar] [CrossRef] [Green Version]
- Rochette, P.J.; Therrien, J.P.; Drouin, R.; Perdiz, D.; Bastien, N.; Drobetsky, E.A.; Sage, E. UVA-Induced Cyclobutane Pyrimidine Dimers Form Predominantly at Thymine–Thymine Dipyrimidines and Correlate with the Mutation Spectrum in Rodent Cells. Nucleic Acids. Res. 2003, 31, 2786. [Google Scholar] [CrossRef] [Green Version]
- Silverman, A.I.; Peterson, B.M.; Boehm, A.B.; McNeill, K.; Nelson, K.L. Sunlight Inactivation of Human Viruses and Bacteriophages in Coastal Waters Containing Natural Photosensitizers. Environ. Sci. Technol. 2013, 47, 1870–1878. [Google Scholar] [CrossRef]
- Taj, M.K.; Ling, J.X.; Bing, L.L.; Qi, Z.; Taj, I.; Hassani, T.M.; Samreen, Z.; Yunlin, W. Effect of Dilution, Temperature and PH on the Lysis Activity of T4 Phage against E. Coli BL21. J. Anim. Plant. Sci. 2014, 24, 1252–1255. [Google Scholar]
- Jurczak-Kurek, A.; Gasior, T.; Nejman-Faleńczyk, B.; Bloch, S.; Dydecka, A.; Topka, G.; Necel, A.; Jakubowska-Deredas, M.; Narajczyk, M.; Richert, M.; et al. Biodiversity of Bacteriophages: Morphological and Biological Properties of a Large Group of Phages Isolated from Urban Sewage. Sci. Rep. 2016, 6, 34338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemb, O.; Manefield, M.; Thomas, F.; Jacquet, S. Phage Adsorption to Bacteria in the Light of the Electrostatics: A Case Study Using E. Coli, T2 and Flow Cytometry. J. Virol. Methods 2013, 189, 283–289. [Google Scholar] [CrossRef]
- Schijven, J.F.; Hassanizadeh, S.M. Removal of Viruses by Soil Passage: Overview of Modeling, Processes, and Parameters. Crit Rev. Environ. Sci. Technol. 2000, 30, 49–127. [Google Scholar] [CrossRef]
- Germann, P.F.; Alaoui, A.; Riesen, D. Drag Force Approach to the Transport of Colloids in Unsaturated Soils. Water Resour. Res. 2002, 38, 18-1–18-15. [Google Scholar] [CrossRef]
- Flynn, R.; Hunkeler, D.; Guerin, C.; Burn, C.; Rossi, P.; Aragno, M. Geochemical Influences on H40/1 Bacteriophage Inactivation in Glaciofluvial Sands. Environ. Geol. 2004, 45, 504–517. [Google Scholar] [CrossRef] [Green Version]
- Kvitsand, H.M.L.; Ilyas, A.; Østerhus, S.W. Rapid Bacteriophage MS2 Transport in an Oxic Sandy Aquifer in Cold Climate: Field Experiments and Modeling. Water Resour. Res. 2015, 51, 9725–9745. [Google Scholar] [CrossRef]
- Flynn, R.M.; Mallèn, G.; Engel, M.; Ahmed, A.; Rossi, P. Characterizing Aquifer Heterogeneity Using Bacterial and Bacteriophage Tracers. J. Environ. Qual. 2015, 44, 1448–1458. [Google Scholar] [CrossRef] [Green Version]
- Chattopadhyay, S.; Puls, R.W. Forces Dictating Colloidal Interactions between Viruses and Soil. Chemosphere 2000, 41, 1279–1286. [Google Scholar] [CrossRef]
- Sautrey, G.; Brié, A.; Gantzer, C.; Walcarius, A. MS2 and Qβ Bacteriophages Reveal the Contribution of Surface Hydrophobicity on the Mobility of Non-Enveloped Icosahedral Viruses in SDS-Based Capillary Zone Electrophoresis. Electrophoresis 2018, 39, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Schijven, J.F.; Hoogenboezem, W.; Hassanizadeh, S.M.; Peters, J.H. Modeling Removal of Bacteriophages MS2 and PRD1 by Dune Recharge at Castricum, Netherlands. Water Resour. Res. 1999, 35, 1101–1111. [Google Scholar] [CrossRef]
- Bales, R.C.; Li, S.; Maguire, K.M.; Yahya, M.T.; Gerba, C.P.; Harvey, R.W. Virus and Bacteria Transport in a Sandy Aquifer, Cape Cod, MA. Groundwater 1995, 33, 653–661. [Google Scholar] [CrossRef]
- Flynn, R.; Cornaton, F.; Hunkeler, D.; Rossi, P. Bacteriophage Transport through a Fining-Upwards Sedimentary Sequence: Laboratory Experiments and Simulation. J. Contam. Hydrol. 2004, 74, 231–252. [Google Scholar] [CrossRef] [PubMed]
- Sinton, L.W.; Noonan, M.J.; Finlay, R.K.; Pang, L.; Close, M.E. Transport and Attenuation of Bacteria and Bacteriophages in an Alluvial Gravel Aquifer. N. Z. J. Mar. Freshw. Res. 2000, 34, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Ghanem, N.; Trost, M.; Sánchez Fontanet, L.; Harms, H.; Chatzinotas, A.; Wick, L.Y. Changes of the Specific Infectivity of Tracer Phages during Transport in Porous Media. Environ. Sci. Technol. 2018, 52, 3486–3492. [Google Scholar] [CrossRef] [PubMed]
- Lisovski, S.; Ramenofsky, M.; Wingfield, J.C. Defining the Degree of Seasonality and Its Significance for Future Research. Integr. Comp. Biol. 2017, 57, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Kwiecien, O.; Braun, T.; Brunello, C.F.; Faulkner, P.; Hausmann, N.; Helle, G.; Hoggarth, J.A.; Ionita, M.; Jazwa, C.S.; Kelmelis, S.; et al. What We Talk about When We Talk about Seasonality—A Transdisciplinary Review. Earth Sci. Rev. 2022, 225, 103843. [Google Scholar] [CrossRef]
- Lembrechts, J.J.; van den Hoogen, J.; Aalto, J.; Ashcroft, M.B.; De Frenne, P.; Kemppinen, J.; Kopecký, M.; Luoto, M.; Maclean, I.M.D.; Crowther, T.W.; et al. Global Maps of Soil Temperature. Glob. Chang. Biol. 2022, 28, 3110–3144. [Google Scholar] [CrossRef]
- Zhan, M.J.; Xia, L.; Zhan, L.; Wang, Y. Recognition of Changes in Air and Soil Temperatures at a Station Typical of China’s Subtropical Monsoon Region (1961–2018). Adv. Meteorol. 2019, 9, 6927045. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, G.N. Madhuri Effect of Water Depth on Daily Yield of the Still. Desalination 1987, 61, 67–75. [Google Scholar] [CrossRef]
- Caldarescu, D.E.; Brey, T.; Abele, D.; Beierlein, L.; Lohmann, G.; Ionita, M. The Influence of Depth-Dependent Seasonal Temperature Variability on Growth Signal in Arctica Islandica. Front. Mar. Sci. 2021, 8, 823. [Google Scholar] [CrossRef]
- Caldentey, J.; Bamford, J.K.H.; Bamford, D.H. Structure and Assembly of Bacteriophage PRD1, an Escherichia Coli Virus with a Membrane. J. Struct. Biol. 1990, 104, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, T.; Bales, R.C.; Maguire, K.M.; Gerba, C.P. Effect of PH on Bacteriophage Transport through Sandy Soils. J. Contam. Hydrol. 1993, 14, 55–70. [Google Scholar] [CrossRef]
- Schijven, J.F.; Hassanizadeh, S.M.; De Bruin, R.H.A.M. Two-Site Kinetic Modeling of Bacteriophages Transport through Columns of Saturated Dune Sand. J. Contam. Hydrol. 2002, 57, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Michen, B.; Graule, T. Isoelectric Points of Viruses. J. Appl. Microbiol. 2010, 109, 388–397. [Google Scholar] [CrossRef]
- Rossi, P.; Käss, W. Phagen. In Geohydrologische Markierungstechnik; Gebruüder Borntraeger: Berlin, Stuttgart, 1998; pp. 209–232. [Google Scholar]
- Hijnen, W.A.M.; Brouwer-Hanzens, A.J.; Charles, K.J.; Medema, G.J. Transport of MS2 Phage, Escherichia Coli, Clostridium Perfringens, Cryptosporidium Parvum and Giardia Intestinalis in a Gravel and a Sandy Soil. Environ. Sci. Technol. 2005, 39, 7860–7868. [Google Scholar] [CrossRef]
- Cao, H.; Tsai, F.T.C.; Rusch, K.A. Salinity and Soluble Organic Matter on Virus Sorption in Sand and Soil Columns. Ground Water 2010, 48, 42–52. [Google Scholar] [CrossRef]
- Chu, Y.; Jin, Y.; Flury, M.; Yates, M.V. Mechanisms of Virus Removal during Transport in Unsaturated Porous Media. Water Resour. Res. 2001, 37, 253–263. [Google Scholar] [CrossRef]
- Chu, Y.; Jin, Y.; Baumann, T.; Yates, M.V. Effect of Soil Properties on Saturated and Unsaturated Virus Transport through Columns. Adsorption 2003, 32, 2017–2025. [Google Scholar] [CrossRef]
- Bitton, G. Adsorption of Viruses Onto Surfaces Soil and Water. Water Res. 1975, 9, 473–484. [Google Scholar] [CrossRef]
- Bitton, G.; Mitchell, R. Effect of Colloids on the Survival of Bacteriophages in Seawater. Water Res. 1974, 8, 227–229. [Google Scholar] [CrossRef]
- Babich, H.; Stotzky, G. Reductions in Inactivation Rates of Bacteriophages by Clay Minerals in Lake Water. Water Res. 1980, 14, 185–187. [Google Scholar] [CrossRef]
- Auckenthaler, A.; Raso, G.; Huggenberger, P. Particle Transport in a Karst Aquifer: Natural and Artificial Tracer Experiments with Bacteria, Bacteriophages and Microspheres. Water Sci. Technol. 2002, 46, 131–138. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Total number of scientific papers reporting the use of bacteriophages during hydrological tracing experiments according to the year of experiments (A) and categorised by the water type investigated (B).
Figure 1.
Total number of scientific papers reporting the use of bacteriophages during hydrological tracing experiments according to the year of experiments (A) and categorised by the water type investigated (B).

Figure 2.
Distribution of bacteriophages commonly used in hydrological tracing experiments, classified according to (A) the bacteriophage families and (B) the bacteriophage species. Bacteriophage families are distinguishable by the background colours of the bar chart, and the type of water studied is represented by the coloured pattern textures. Unknown = bacteriophage strains (Cloacae phage, Serratia phage, Aerog243 phage, F137, F52, and F46) for which the families are not known.
Figure 2.
Distribution of bacteriophages commonly used in hydrological tracing experiments, classified according to (A) the bacteriophage families and (B) the bacteriophage species. Bacteriophage families are distinguishable by the background colours of the bar chart, and the type of water studied is represented by the coloured pattern textures. Unknown = bacteriophage strains (Cloacae phage, Serratia phage, Aerog243 phage, F137, F52, and F46) for which the families are not known.

Figure 3.
Methodology for the detection of bacteriophages from environmental samples. (Dil = dilution).
Figure 3.
Methodology for the detection of bacteriophages from environmental samples. (Dil = dilution).


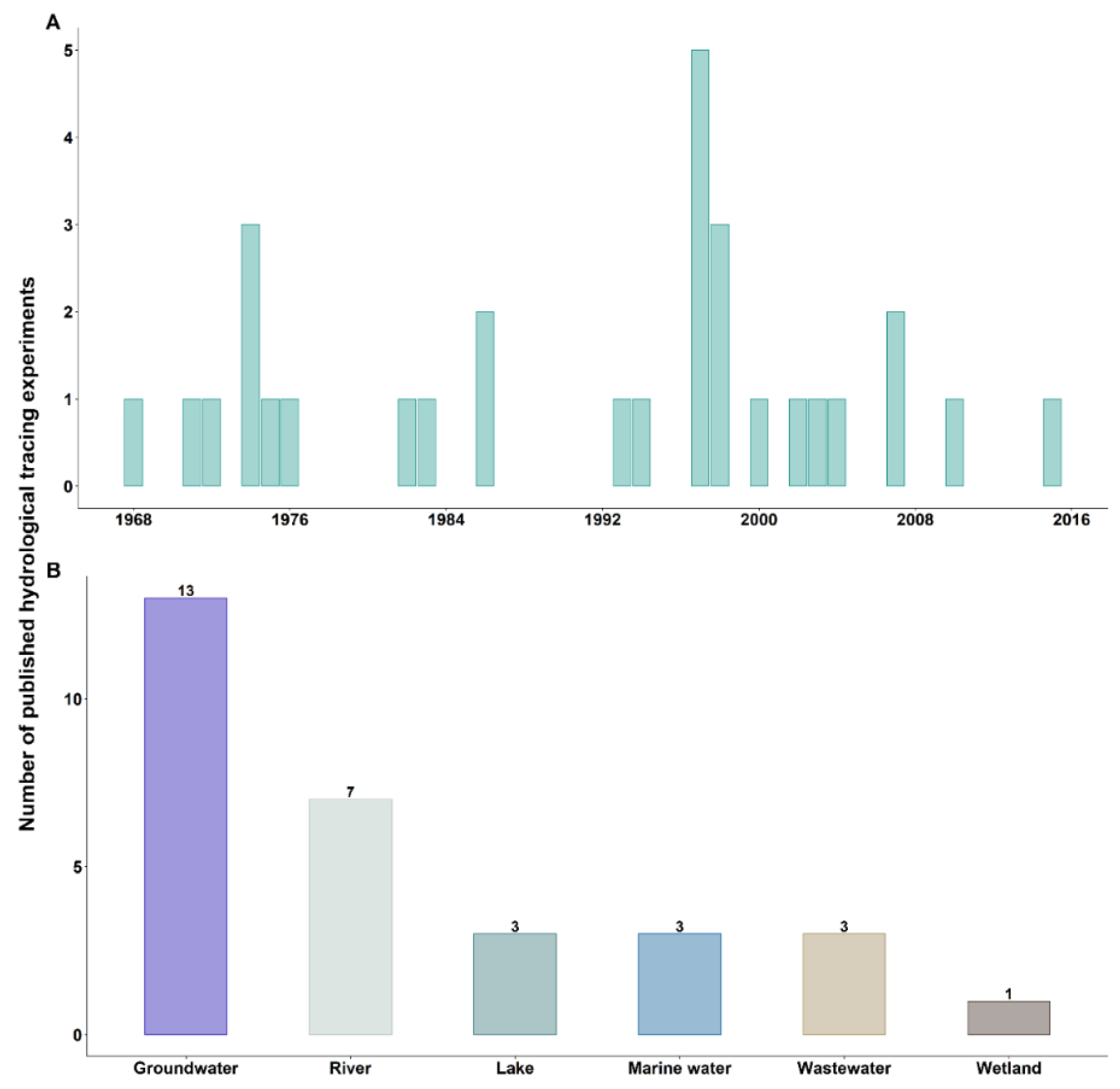{kind=link}
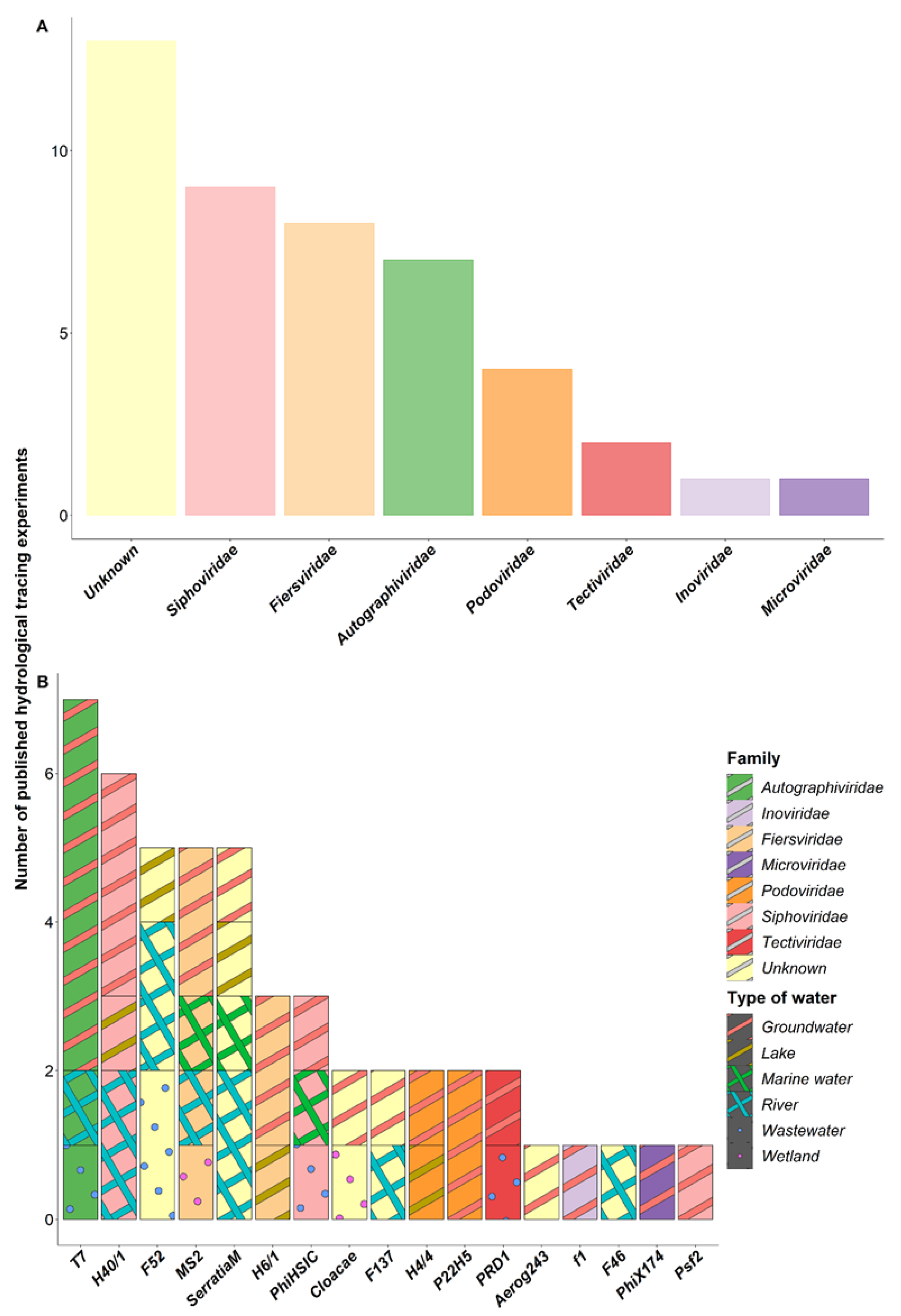{kind=link}
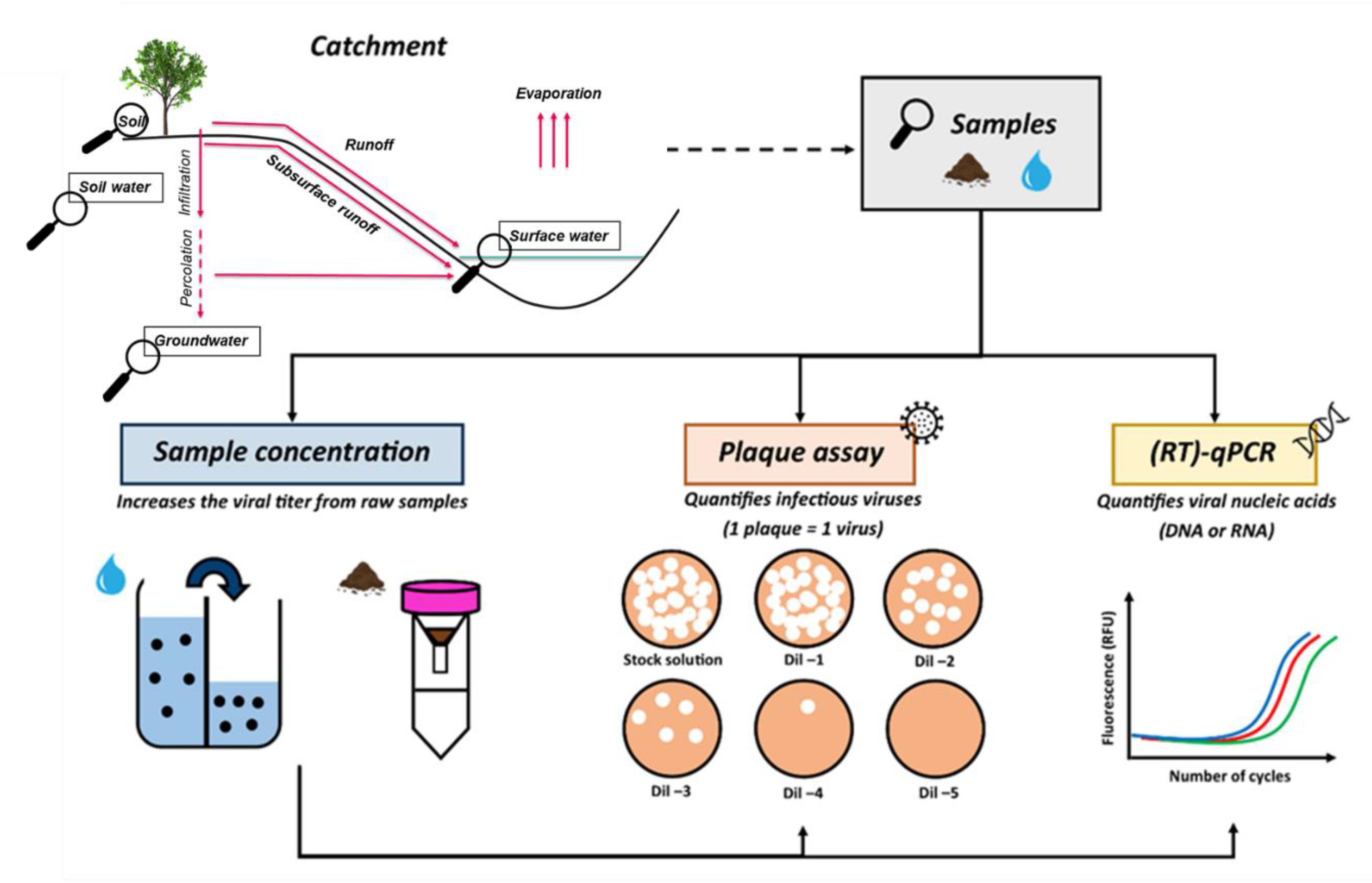{kind=link}
Table 1.
Characteristics of all bacteriophages used as hydrological tracers.
| Bacteriophages | Family | Structure | Genome | Isoelectric Point | Bacterial Host | References |
|---|---|---|---|---|---|---|
| THE FIVE MOST USED BACTERIOPHAGES AS HYDROLOGICAL TRACERS | ||||||
| Enterobacteria phage T7 (Teseptimavirus T7) | Autographiviridae | 55-nm capsid with 19-nm tail | dsDNA of 40 kb | 4.85 | Most strains of Escherichia coli | [65,66,67] |
| Pseudoalteromonas phage vB_PspS-H40/1 | Siphoviridae | Non-contractile 68-nm tail and an icosahedral capsid of 45-nm diameter | dsDNA of 45 kb | Unknown | Pseudoalteromonas sp. | [68] |
| Coliphage F52 | Unknown | Unknown | Unknown | Unknown | Escherichia coli | [38] |
| Lessievirus MS2 (Emesvirus zinderi) | Fiersviridae | Small icosahedral virus (28 nm) | (+) ssRNA of 3 kb | 3.9 | Escherichia coli | [59,69] |
| Serratia phage strains * | Myoviridae, Siphoviridae, Podoviridae, Ackermannviridae | Head-tailed structure | dsDNA of 44–350 kb | Unknown | Strains of Serratia marcescens | [63,70] |
| ALL THE OTHER BACTERIOPHAGES USED AS HYDROLOGICAL TRACERS | ||||||
| Listonella phage phiHSIC | Siphoviridae | 47-nm capsid diameter with a non-contractile tail of 146 nm | dsDNA genome of 37,966 bp | Unknown | Vibrio pelagius | [71] |
| Pseudoalteromonasvirus vB_PspP-H6/1 | Podoviridae | 56-nm diameter icosahedral capsid and a short (15 nm) tail | linear dsDNA genome of a size of 36,753 bp | <4 | Pseudoalteromonas sp. | [72,73] |
| Alteromonasvirus vB_AspP-H4/4 | Podoviridae | Icosahedral capsid of 41 nm with a short tail of 6.6 nm | dsDNA genome of 47,631 bp | Unknown | Alteromonas sp. | [68] |
| Enterobacteria phage PRD1 (Alphatectivirus PRD1) | Tectiviridae | 62-nm capsid | dsDNA genome of 15 kb | 3.8–4.2 | Gram-negative bacterial species (i.e., Escherichia coli, Salmonella enterica, Pseudomonas aeruginosa) | [74,75] |
| Enterobacter cloacae phage strains * | Unknown | Isometric head of about 55–93 nm with a 103 nm long | DNA genomes | Unknown | Enterobacter cloacae | [76,77] |
| Coliphages F137 and F46 | Unknown | Unknown | Unknown | Unknown | Escherichia coli | [38] |
| Salmonella phage P22H5 | Podoviridae | T7-like structure | dsDNA | Unknown | Salmonella typhimurium | [49] |
| Enterobacteria phage f1 | Inoviridae | Filamentous 850 nm long | ssDNA | Unknown | Escherichia coli | [36] |
| Pseudomonas phage Psf2 (Tunavirus Psf2) | Drexlerviridae (former Siphoviridae) | T1-like structure (60-nm head and 151-nm tail) | Circular genome of about 50 kb | Unknown | Pseudomonas fluorescens | [42] |
| Enterobacteria phage phiX174 (Sinsheimervirus phiX174) | Microviridae | Icosahedral capsid | Circular ssDNA | 6.6–7 | Escherichia sp. | [78] |
| Aerobacter aerogenose 243 phages | Unknown | Unknown | Unknown | Unknown | Aerobacter aerogenes NCTC 243 | [41] |
Note: * None of the articles clarified which bacteriophage strain was used. The names in brackets are the latest scientific names updated by the ICTV, and those in bold are the names used in the various papers in which bacteriophages were used as hydrological tracers.
Table 2.
Characteristics of the transportation of bacteriophages during the various tracing experiments.
Table 2.
Characteristics of the transportation of bacteriophages during the various tracing experiments.
| Bacteriophages | Reservoirs | Distance Travelled | Time Travelled | Velocity | Loss at the End of the Experiment | Number of Experiments | Other Tracers Used in Complement | References |
|---|---|---|---|---|---|---|---|---|
| THE FIVE MOST USED BACTERIOPHAGES AS HYDROLOGICAL TRACERS | ||||||||
| Enterobacteria phage T7 (Teseptimavirus T7) | Groundwater and lake | 1–160 m (24 km in one experiment) | 20–70 h | 1–350 m/h | 99.9% | 7 | H40/1, f1, F52, F137, F46, naphthionate | [36,38,45] |
| Pseudoalteromonas phage vB_PspS-H40/1 | Freshwater (especially lakes) | 64 m–6 km | 8–48 h | 8–2000 m/h | 40–99% | 6 | T7, H6/1, uranine | [45] |
| Coliphage F52 | River and lake | 8–62 km | 4–119 h | 900–2000 m/h | - | 5 | T7, F137, F46 | [38] |
| Lessievirus MS2 (Emesvirus zinderi) | Water surface treatment wetlands, aquifers | 4–15 m | 4 h–5 d | 0.2–19 m/h | 10–77% | 5 | PhiHSIC, PRD1, bromide | [44,46,48] |
| Serratia phage strains * | Surface water | 3–10 km | 1–6 h | - | 30–62% | 5 | MS2, fluorescein, Enterobacteria phage, Bacillus spores, Lithium | [15,35] |
| ALL THE OTHER BACTERIOPHAGES USED AS HYDROLOGICAL TRACERS | ||||||||
| Listonella phage phiHSIC | From wastewater to surface water and groundwater | 10 m–4 km | 3–10 h | 0.1–2 m/d 1–140 m/h | - | 3 | PRD1, MS2 | [44,51] |
| Pseudoalteromonasvirus vB_PspP-H6/1 | Freshwater (especially lakes) | 6 km | 5–26 h to 4 d | 23–210 m/h | 75–99.9% | 3 | H40/1, T7, Psf2, H4/4 | [42,45,79] |
| Alteromonasvirus vB_AspP-H4/4 | Freshwater (especially lakes) | - | 1–48 h | - | 99.9% | 2 | H40/1, T7, Psf2, H6/1 | [42,79] |
| Enterobacteria phage PRD1 (Alphatectivirus PRD1) | Low clay-content media | 4 m–4 km | 6 h–5 d | 0.2–5 m/d 1–57 m/h | 50% | 2 | MS2, bromide, PhiHSIC | [48,51] |
| Enterobacter cloacae phage strains * | Constructed wetlands and groundwater | - | 3–14 d | 0.8–4 L/s | 9–64% in wetlands 100% in groundwater | 2 | Photine, fluorescein, Serratia phage | [47] |
| Coliphages F137 and F46 | River and groundwater | 31–46 km | 78–111 h | 300–400 m/h | - | 2 and 1 | T7, F52 | [38] |
| Salmonella phage P22H5 | Karstic aquifer, groundwater | - | 4–28 d | 4–36 m/d | 99.2–99.9% | 2 | Deuterium, bromide, chloride, sulfate, pyranine, naphthionate, uranine, sulforhoramine, microspheres | [49,50] |
| Enterobacteria phage f1 | Permeable aquifers | 11–110 m | 20–70 h | - | 100% | 1 | T7, naphthionate | [36] |
| Pseudomonas phage Psf2 (Tunavirus Psf2) | Groundwater | - | 11 d | - | - | 1 | T7, H6/1, H40/1, H4/4 | [42] |
| Enterobacteria phage phiX174 (Sinsheimervirus phiX174) | Karstic aquifer | - | 10–14 d | - | 99% | 1 | Photine, fluorescein, Serratia phage, Enterobacteria phage | [47] |
| Aerobacter aerogenose 243 phages | Groundwater | 200–700 m | 2–8 d | 1–8 m/h | 99.9% | 1 | - | [41] |
Note: * None of the articles clarified which bacteriophage strain was used. The names in brackets are the latest scientific names updated by the ICTV, and those in bold are the names used in the various papers in which bacteriophages were used as hydrological tracers. h = hours; d = days; s = seconds; m = meters; km = kilometers; L = liters.
Table 3.
The three main issues in the use of bacteriophages as hydrological tracers with their limitations and the proposed solutions.
Table 3.
The three main issues in the use of bacteriophages as hydrological tracers with their limitations and the proposed solutions.
| Status Quo | Encountered Problems | Proposed Solutions | |
|---|---|---|---|
| Detection methods | Use of culture-based methods only | This only detects the infectious bacteriophages, which mean they need to be intact to be detected. | The complementary use of cultured-based methods with molecular techniques, which would allow for the detection of the nucleic acid. Thus, all bacteriophages, intact and damaged, can be detected. |
| Eco-friendly property | Bacteriophages are biological entities and thus are safe for the environment. In addition, they can be rapidly eliminated from the environment. | The injection of large volumes of highly concentrated bacteriophage solutions into the environment. Their genetic material can be released and persist for a long period in the environment. | The natural populations of bacteriophages could be considered since they are numerously abundant. |
| Surrounding environment | Few of the reported tracing experiments using bacteriophages considered the characteristics of the ecosystems of interest and the environmental conditions before tracer injection. | Bacteriophage inactivation is highly dependent on environmental factors, which can lead to their detection using cultured-based methods being missed. | Characterise the catchment of interest before launching the tracing experiment and select the bacteriophage species according to the relationship between the virus and the catchment properties. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Florent, P.; Cauchie, H.-M.; Ogorzaly, L. A Virological Perspective on the Use of Bacteriophages as Hydrological Tracers. Water 2022, 14, 3991. https://doi.org/10.3390/w14243991
AMA Style
Florent P, Cauchie H-M, Ogorzaly L. A Virological Perspective on the Use of Bacteriophages as Hydrological Tracers. Water. 2022; 14(24):3991. https://doi.org/10.3390/w14243991
Chicago/Turabian StyleFlorent, Perrine, Henry-Michel Cauchie, and Leslie Ogorzaly. 2022. "A Virological Perspective on the Use of Bacteriophages as Hydrological Tracers" Water 14, no. 24: 3991. https://doi.org/10.3390/w14243991
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.