
Influence of Anthropogenic Activities on Redox Regulation and Oxidative Stress Responses in Different Phyla of Animals in Coastal Water via Changing in Salinity

 ,
,
Abstract
:1. Introduction
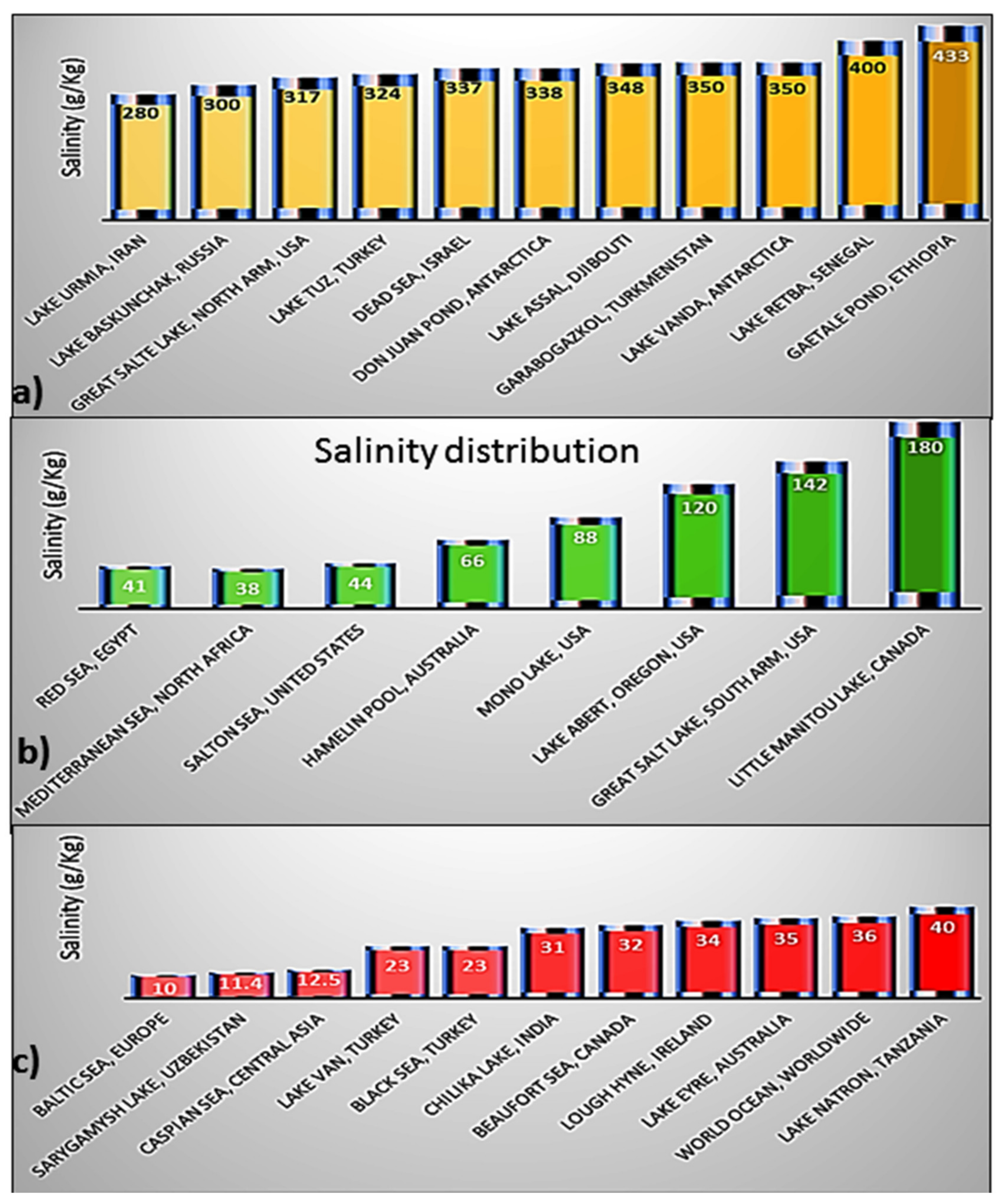
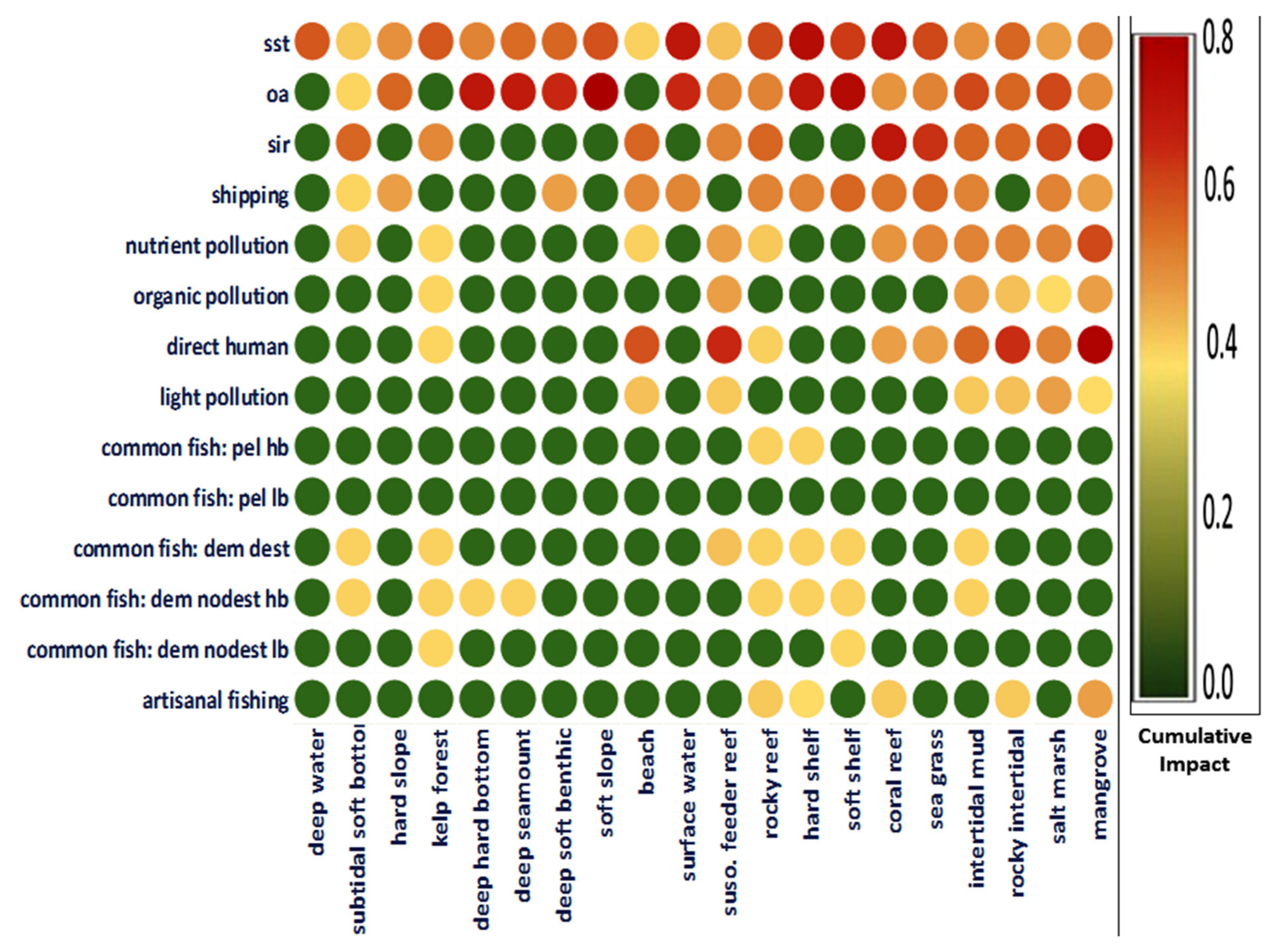
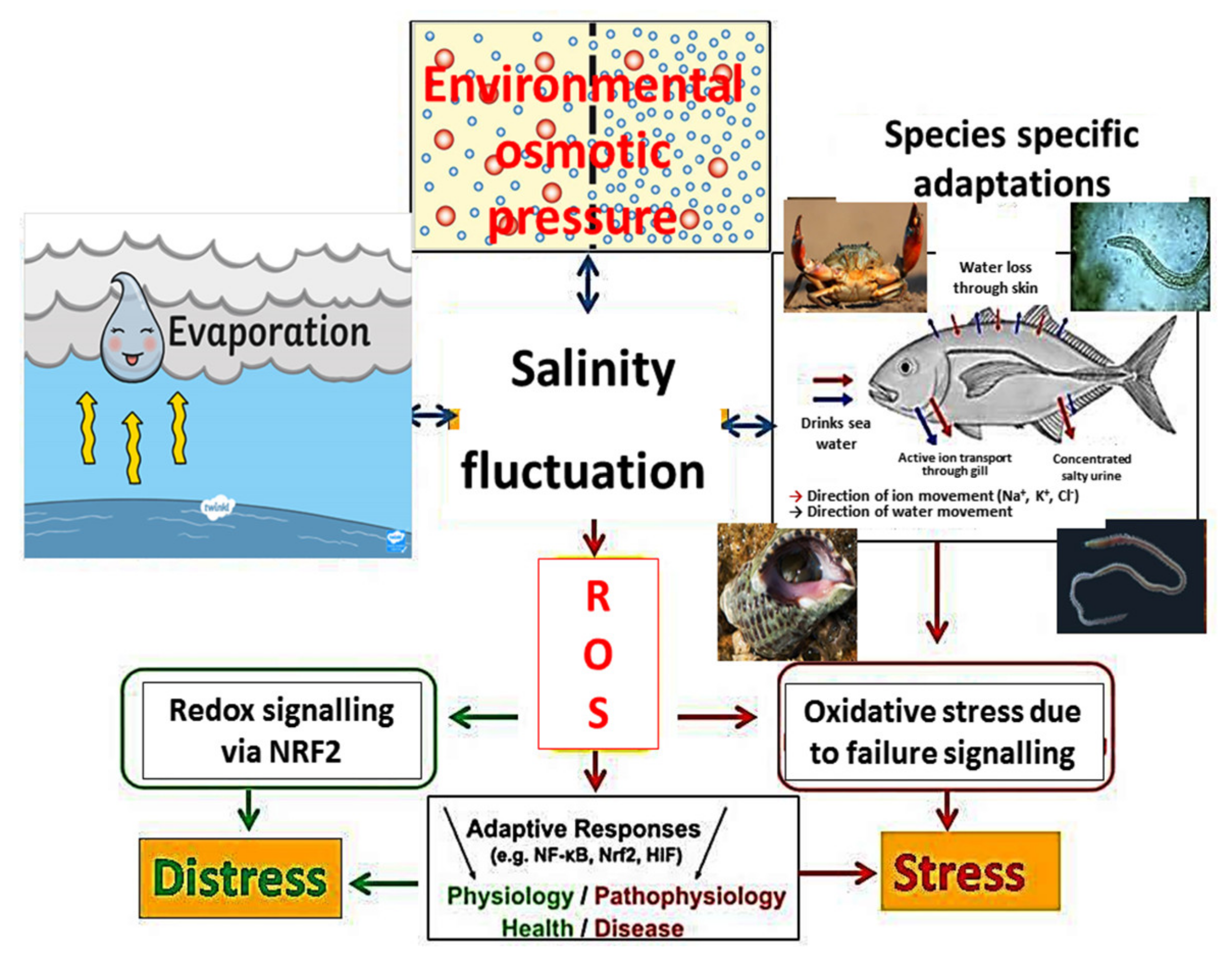
2. Influence of Anthropogenic Activities and Climatic Factors on the Salinity of Coastal Water
3. Salinity and Responses of Animals from Different Phyla
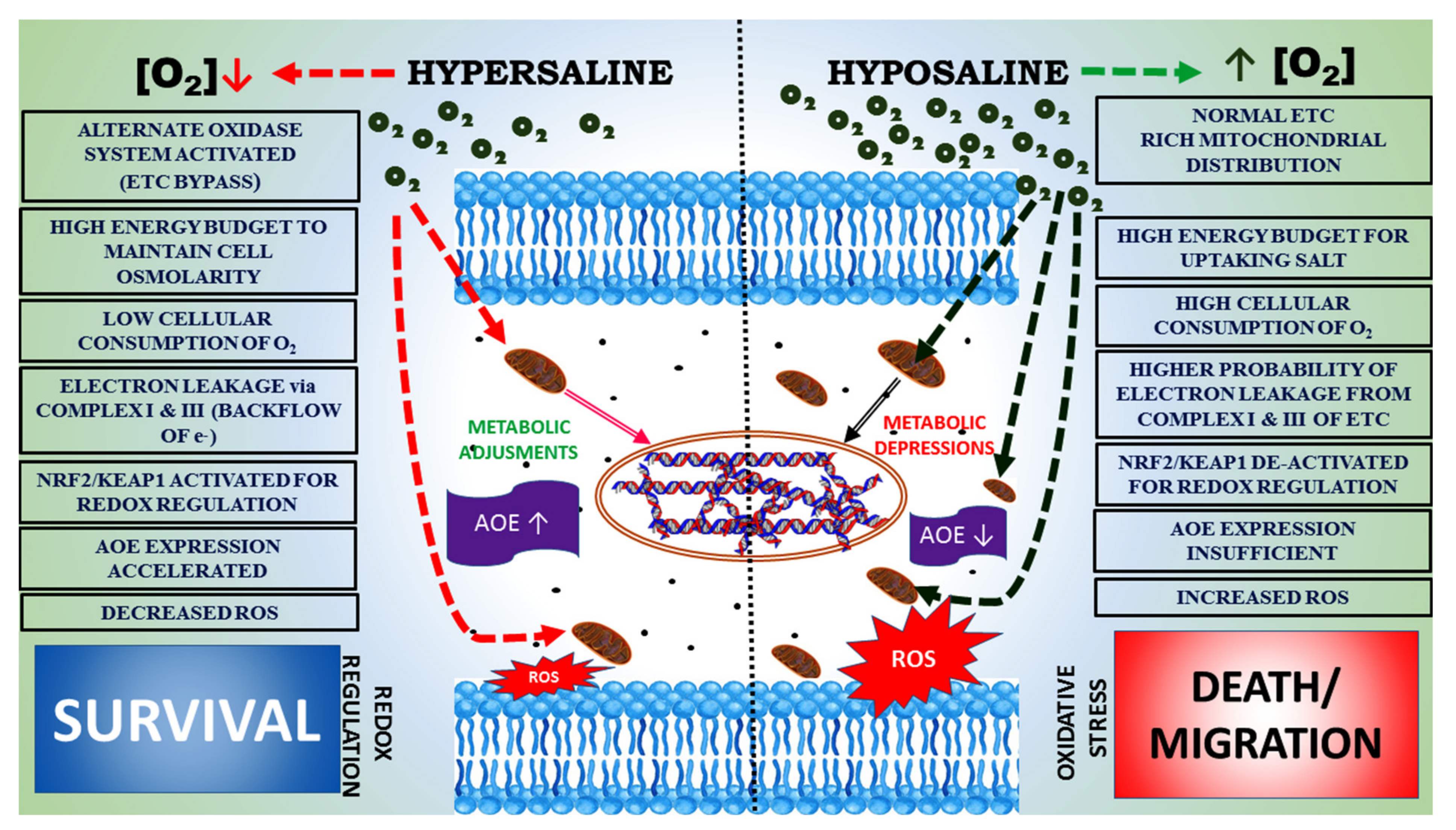
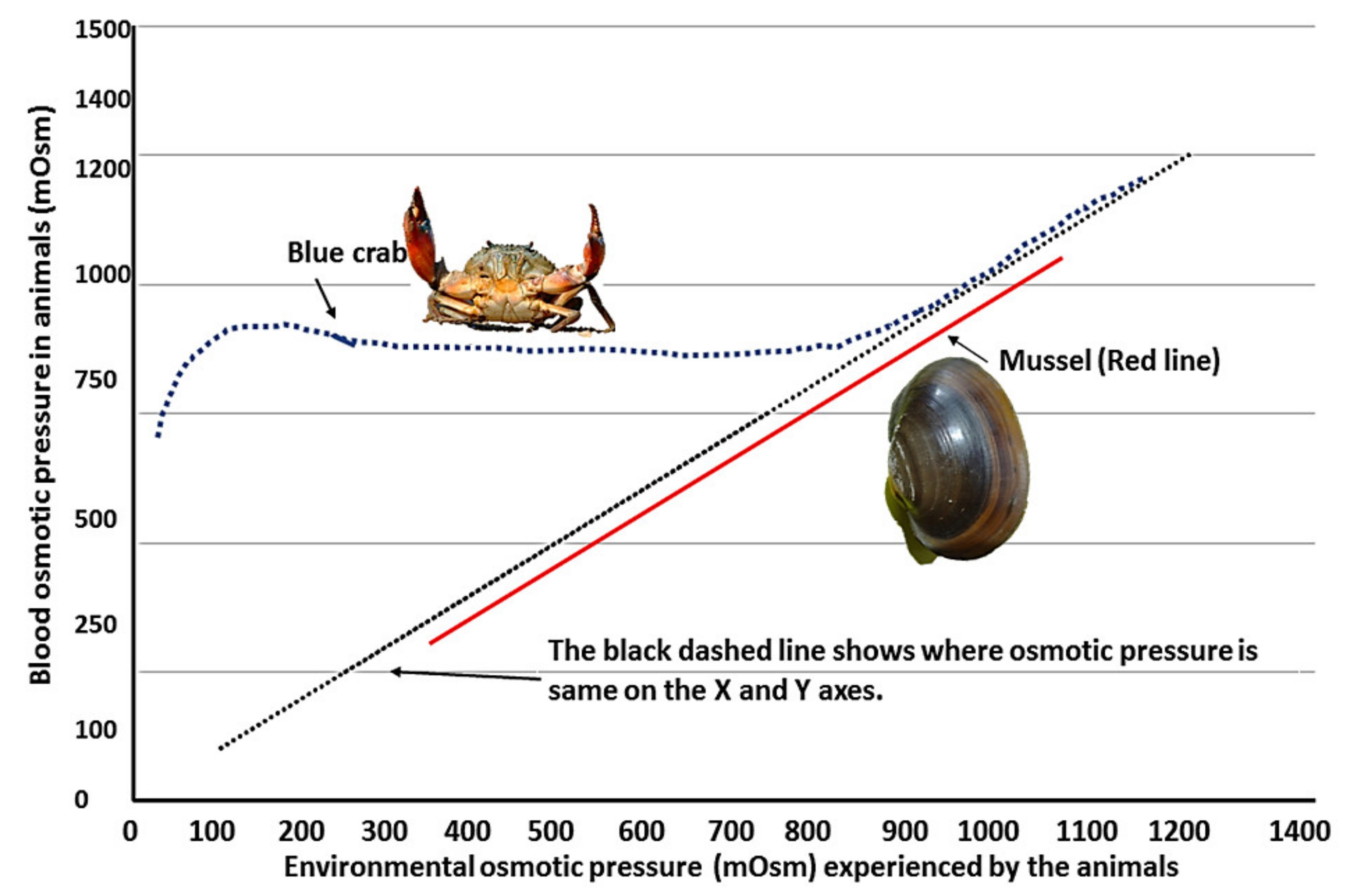
3.1. Physiology of Animals under Changing Salinity
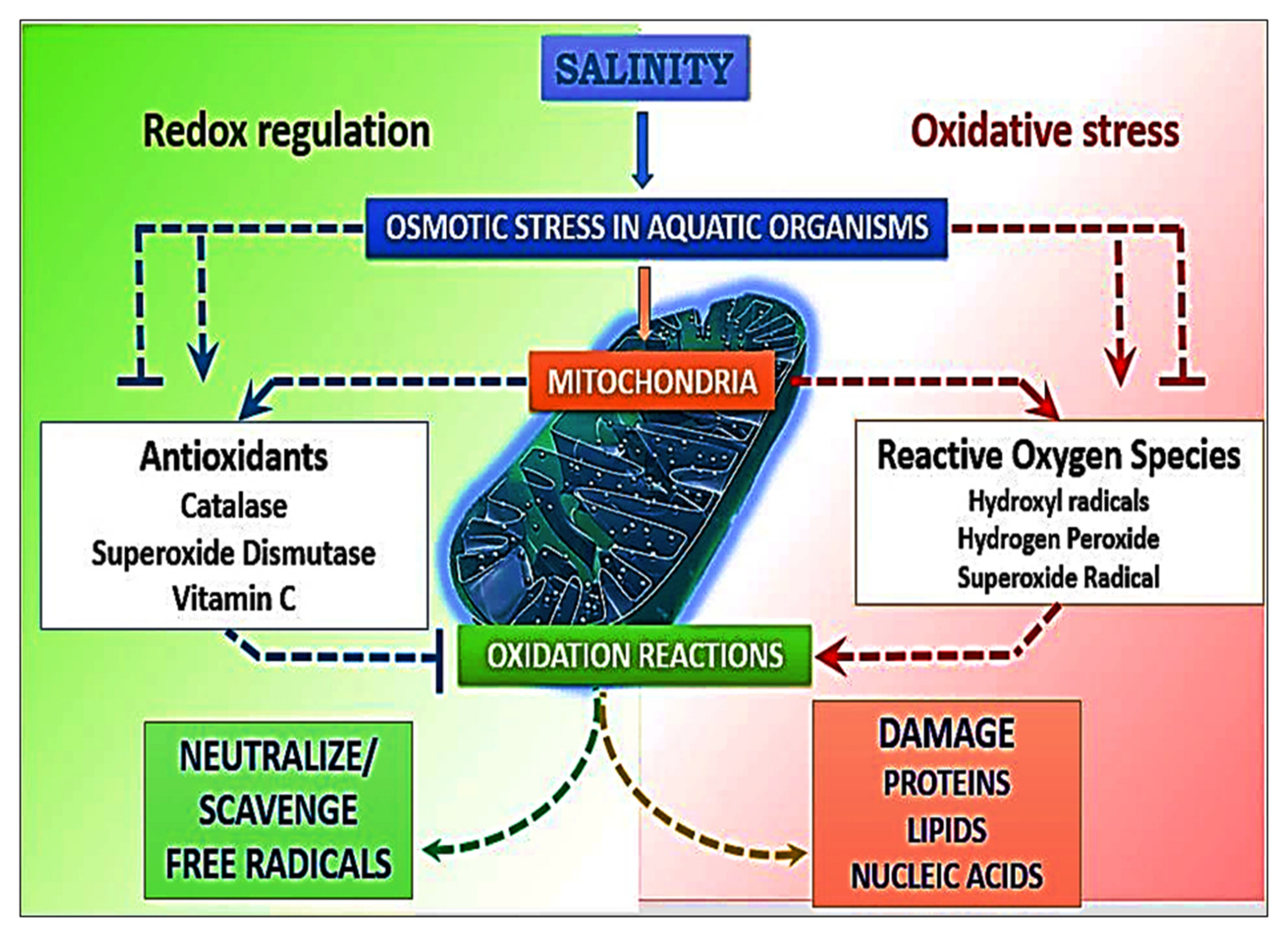
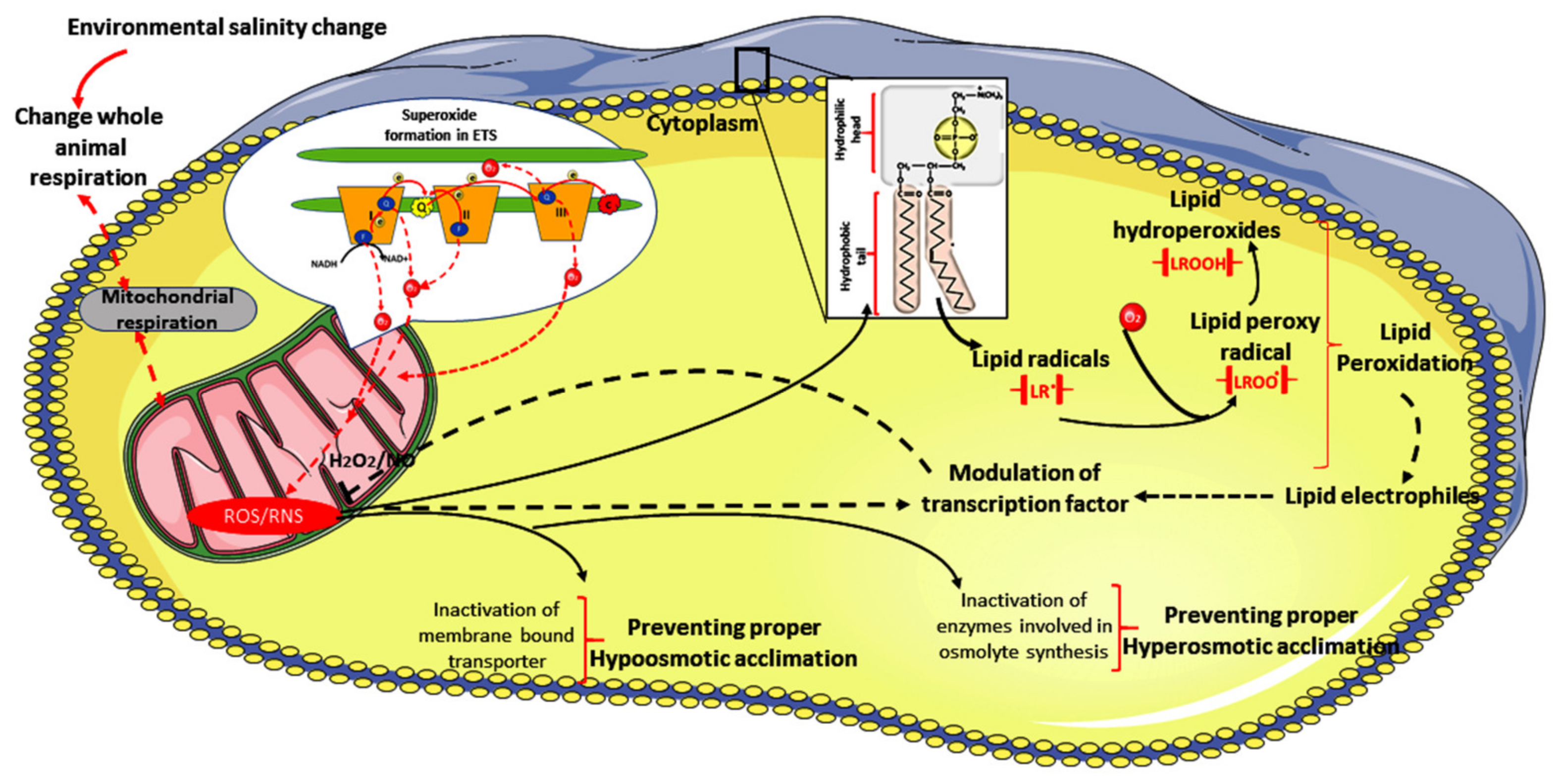
3.2. Stress Incurred in Animals under Salinity Stress
Responses of Animals to Primary and Drastic Salinization
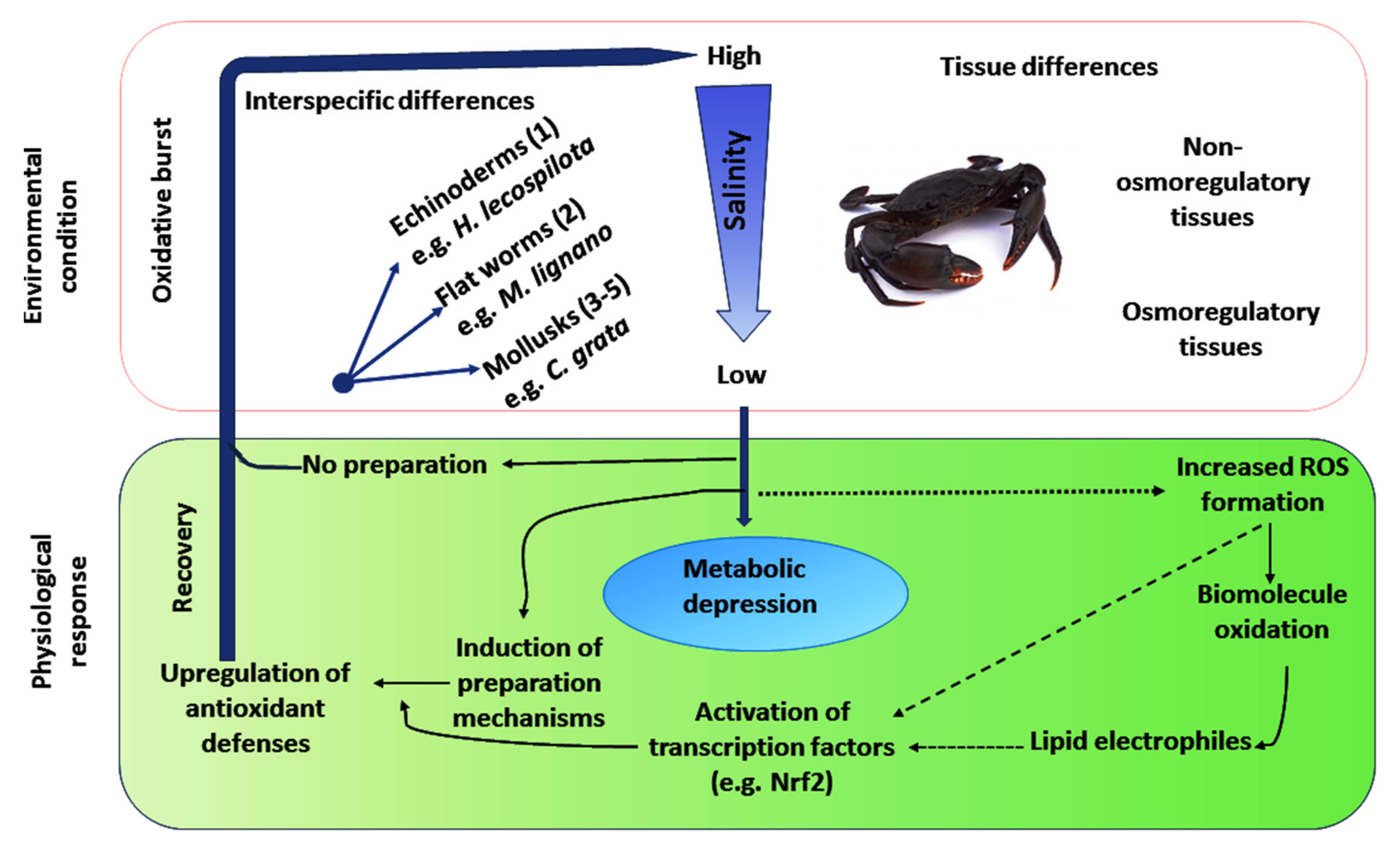
3.3. Inclusive Responses of Organisms with Respect to Alterations in Salinity
3.3.1. Rotifers
3.3.2. Helminthes
3.3.3. Mollusks
3.3.4. Annelids
3.3.5. Arthropods
3.3.6. Echinoderms
3.3.7. Fishes
3.4. Signaling Pathways under Salinity Stress
4. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Williams, W.D. Anthropogenic salinisation of inland waters. Hydrobiologia 2001, 466, 329–337. [Google Scholar] [CrossRef]
- Kefford, B.J.; Hickey, G.; Gasith, A.; Ben-David, E.; Dunlop, J.; Palmer, C.; Allan, K.; Choy, S.; Piscart, C. Global scale variation in the salinity sensitivity of riverine macroinvertebrates: Eastern Australia, France, Israel and South Africa. PLoS ONE 2012, 7, e35224. [Google Scholar] [CrossRef] [PubMed]
- Sowa, A.; Krodkiewska, M.; Halabowski, D.; Lewin, I. Response of the mollusc communities to environmental factors along an anthropogenic salinity gradient. Sci. Nat. 2019, 106, 60. [Google Scholar] [CrossRef] [Green Version]
- Robinson, R.A. The vapour pressure and osmotic equivalence of sea water. J. Mar. Biol. Assoc. UK 1954, 33, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Haumann, F.A.; Gruber, N.; Münnich, M.; Frenger, I.; Kern, S. Sea-ice transport driving southern ocean salinity and its recent trends. Nature 2016, 537, 89–92. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Antioxidant defenses and oxidative stress parameters in tissues of mud crab (Scylla serrata) with reference to changing salinity. Comp. Biochem. Physiol. 2010, 151, 142–151. [Google Scholar] [CrossRef]
- Bal, A.; Panda, F.; Pati, S.G.; Das, K.; Agrawal, P.K.; Paital, B. Modulation of physiological oxidative stress and antioxidant status by abiotic factors especially salinity in aquatic organisms. Comp. Biochem. Physiol. C 2022, 241, 108971. [Google Scholar] [CrossRef]
- Lushchak, V.I.; Bagnyukova, T.V. Temperature increase results in oxidative stress in goldfish tissues. 1. Indices of oxidative stress. Comp. Biochem. Physiol. 2006, 143, 30–35. [Google Scholar] [CrossRef]
- Zeng, Y.; Zhao, C.; Kundzewicz, Z.W.; Lv, G. Distribution pattern of Tugai forests species diversity and their relationship to environmental factors in an arid area of China. PLoS ONE 2020, 15, e0232907. [Google Scholar] [CrossRef]
- Nasrolahi, A.; Havenhand, J.; Wrange, A.L.; Pansch, C. Population and life-stage specific sensitivities to temperature and salinity stress in barnacles. Sci. Rep. 2016, 6, 32263. [Google Scholar] [CrossRef]
- Vélez-Martín, A.; Davy, A.J.; Luque, C.J.; Castellanos, E.M. Disentangling elevation, annual flooding regime and salinity as hydrochemical determinants of halophyte distribution in non-tidal saltmarsh. Ann. Bot. 2020, 126, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Sabat, P.; Narváez, C.; Peña-Villalobos, I.; Contreras, C.; Maldonado, K.; Sanchez-Hernandez, J.C.; Newsome, S.D.; Nespolo, R.; Bozinovic, F. Coping with salt water habitats: Metabolic and oxidative responses to salt intake in the rufous-collared sparrow. Front. Physiol. 2017, 8, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic acid-regulated antioxidant mechanisms and gene expression enhance rosemary performance under saline conditions. Front. Physiol. 2017, 8, 716. [Google Scholar] [CrossRef] [PubMed]
- Hong, M.; Li, N.; Li, J.; Li, W.; Liang, L.; Li, Q.; Wang, R.; Shi, H.; Storey, K.B.; Ding, L. Adenosine monophosphate-activated protein kinase signaling regulates lipid metabolism in response to salinity stress in the red-eared slider turtle Trachemys scripta elegans. Front. Physiol. 2019, 10, 962. [Google Scholar] [CrossRef] [Green Version]
- Cilenti, A.; Provenzano, M.R.; Senesi, N. Characterization of dissolved organic matter from saline soils by fluorescence spectroscopy. Environ. Chem. Lett. 2005, 3, 53–56. [Google Scholar] [CrossRef]
- Munksgaard, N.C.; Wurster, C.M.; Bass, A.; Zagorskis, I.; Bird, M.I. First continuous shipboard δ18O and δD measurements in sea water by diffusion sampling—Cavity ring-down spectrometry. Environ. Chem. Lett. 2012, 10, 301–307. [Google Scholar] [CrossRef]
- Castaño-Sánchez, A.; Hose, G.C.; Reboleira, A.S.P.S. Salinity and temperature increase impact groundwater crustaceans. Sci. Rep. 2020, 10, 12328. [Google Scholar] [CrossRef]
- Mahmoud, E.; El Baroudy, A.; Ali, N.; Sleem, M. Soil amendment with nanoresidues from water treatment increases P adsorption in saline soils. Environ. Chem. Lett. 2020, 18, 171–179. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Effects of salinity on O2 consumption, ROS generation and oxidative stress status of gill mitochondria of the mud crab Scylla serrata. Comp. Biochem. Physiol. C 2012, 155, 228–237. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Modulation of expression of SOD isoenzymes in mud crab (Scylla serrata): Effects of inhibitors, salinity and season. J. Enzym. Inhib. Med. Chem. 2013, 28, 195–204. [Google Scholar] [CrossRef]
- Paital, B.; Chainy, G.B.N. Seasonal variability of antioxidant biomarkers in mud crabs (Scylla serrata). Ecotoxicol. Environ. Saf. 2013, 87, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, N.; Ramalingam, C. Silver nanoparticle antimicrobial activity explained by membrane rupture and reactive oxygen generation. Environ. Chem. Lett. 2016, 14, 477–485. [Google Scholar] [CrossRef]
- Gaikwad, K.K.; Singh, S.; Lee, Y.S. Oxygen scavenging films in food packaging. Environ. Chem. Lett. 2018, 16, 523–538. [Google Scholar] [CrossRef]
- Ranjan, S.; Ramalingam, C. Titanium dioxide nanoparticles induce bacterial membrane rupture by reactive oxygen species generation. Environ. Chem. Lett. 2016, 14, 487–494. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Free Radicals in Biology and Medicine, 8th ed.; Oxford University Press: Oxford, UK, 2015. [Google Scholar] [CrossRef]
- Liu, J.; Jia, H.; Zhu, K.; Zhao, S.; Lichtfouse, E. Formation of environmentally persistent free radicals and reactive oxygen species during the thermal treatment of soils contaminated by polycyclic aromatic hydrocarbons. Environ. Chem. Lett. 2020, 18, 1329–1336. [Google Scholar] [CrossRef]
- Liu, J.; Gao, N.; Wen, X.; Jia, H.; Lichtfouse, E. Plant and algal toxicity of persistent free radicals and reactive oxygen species generated by heating anthracene-contaminated soils from 100 to 600 °C. Environ. Chem. Lett. 2021, 19, 2695–2703. [Google Scholar] [CrossRef]
- Netto, L.E.; Antunes, F. The roles of peroxiredoxin and thioredoxin in hydrogen peroxide sensing and in signal transduction. Mol. Cells 2016, 39, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Turrens, J.F. Mitochondrial formation of reactive oxygen species. J. Physiol. 2003, 552, 335–344. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, W.-N.; Wang, A.-L.; Wang, J.-M.; Sun, R.Y. Effects of dietary vitamin E supplementation on antioxidant enzyme activities in Litopenaeus vannamei (Boone, 1931) exposed to acute salinity changes. Aquaculture 2007, 265, 351–358. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- De Martinez, G.M.; Bianchini, A. Metallothionein-like proteinsin the bluecrab Callinectes sapidus: Effect of water salinity and ions. Comp. Biochem. Physiol. A 2009, 152, 366–371. [Google Scholar] [CrossRef] [PubMed]
- De Zoysa, M.; Whang, I.; Lee, Y.; Lee, S.; Lee, J.-S.; Lee, J. Transcriptional analysis of antioxidant and immune defense genes in disk abalone (Haliotis discus discus) during the thermal, low salinity and hypoxic stress. Comp. Biochem. Physiol. B 2009, 154, 387–395. [Google Scholar] [CrossRef] [PubMed]
- An, M.I.; Choi, C.Y. Activity of antioxidant enzymes and physiological responses in ark shell, Scapharca broughtonii, exposed to thermal and osmotic stress: Effects on hemolymph and biochemical parameters. Comp. Biochem. Physiol. B 2010, 155, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Bayir, A.; Sirkecioglu, A.N.; Bayir, M.; Haliloglu, H.I.; Kocaman, E.M.; Aras, N.M. Metabolic responses to prolonged starvation, food restriction, and refeeding Salmo trutta: Oxidative stress and antioxidant defences. Comp. Biochem. Physiol. B 2011, 159, 191–196. [Google Scholar] [CrossRef]
- Jiang, W.; Fang, J.; Du, M.; Gao, Y.; Fang, J.; Jiang, Z. Integrated transcriptomics and metabolomics analyses reveal benzo[a]pyrene enhances the toxicity of mercury to the Manila clam, Ruditapes philippinarum. Ecotoxicol. Environ. Saf. 2021, 213, 112038. [Google Scholar] [CrossRef]
- Freitas, R.; Pires, A.; Velez, C.; Almeida, Â.; Wrona, F.J.; Soares, A.M.; Figueira, E. The effects of salinity changes on the Polychaete Diopatra neapolitana: Impacts on regenerative capacity and biochemical markers. Aquat. Toxicol. 2015, 163, 167–176. [Google Scholar] [CrossRef]
- Bertrand, C.; Zalouk-Vergnoux, A.; Giambérini, L.; Poirier, L.; Devin, S.; Labille, J.; Perrein-Ettajani, H.; Pagnout, C.; Châtel, A.; Levard, C.; et al. The influence of salinity on the fate and behavior of silver standardized nanomaterial and toxicity effects in the estuarine bivalve Scrobicularia plana. Environ. Toxicol. Chem. 2016, 35, 2550–2561. [Google Scholar] [CrossRef]
- Hossain, M.A.; Aktar, S.; Qin, J.G. Salinity stress response in estuarine fishes from the Murray Estuary and Coorong, South Australia. Fish Physiol. Biochem. 2016, 42, 1571–1580. [Google Scholar] [CrossRef]
- Lee, M.C.; Park, J.C.; Kim, D.H.; Kang, S.; Shin, K.; Park, H.; Han, J.; Lee, J.S. Interrelationship of salinity shift with oxidative stress and lipid metabolism in the monogonont rotifer Brachionus koreanus. Comp. Biochem. Physiol. A 2017, 214, 79–84. [Google Scholar] [CrossRef]
- Moreira, A.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Salinity influences the biochemical response of Crassostrea angulata to Arsenic. Environ. Pollut. 2016, 214, 756–766. [Google Scholar] [CrossRef]
- Rivera-Ingraham, G.A.; Nommick, A.; Blondeau-Bidet, E.; Ladurner, P.; Lignot, J.-H. Salinity stress from the perspective of the energy-redox axis: Lessons from a marine intertidal flatworm. Redox Biol. 2016, 10, 53–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Combined effects of seawater acidification and salinity changes in Ruditapes philippinarum. Aquat. Toxicol. 2016, 176, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Velez, C.; Figueira, E.; Soares, A.M.V.M.; Freitas, R. Native and introduced claims biochemical responses to salinity and pH changes. Sci. Total Environ. 2016, 566–567, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Fuhrmann, M.; Delisle, L.; Petton, B.; Corporeau, C.; Pernet, F. Metabolism of the pacific oyster, Crassostrea gigas, is influenced by salinity and modulates survival to the Ostreid herpes virus OsHV-1. Biol. Open 2018, 7, bio028134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado-Gaytán, M.F.; Gómez-Jiménez, S.; Gámez-Alejo, L.A.; Rosas-Rodríguez, J.A.; Figueroa-Soto, C.G.; Valenzuela-Soto, E.M. Effect of salinity on the synthesis and concentration of glycine betaine in osmoregulatory tissues from juvenile shrimps Litopenaeus vannamei. Comp. Biochem. Physiol. A 2020, 240, 110628. [Google Scholar] [CrossRef]
- Lin, G.; Zheng, M.; Gao, D.; Li, S.; Fang, W.; Huang, J.; Xie, J.; Liu, J.; Liu, Y.; Li, Z.; et al. Hypoosmotic stress induced tissue-specific immune responses of yellowfin seabream (Acanthopagrus latus) revealed by transcriptomic analysis. Fish Shellfish. Immunol. 2020, 99, 473–482. [Google Scholar] [CrossRef]
- Rivera-Ingraham, G.A.; Lignot, J.H. Osmoregulation, bioenergetics and oxidative stress in coastal marine invertebrates: Raising the questions for future research. J. Exp. Biol. 2017, 220, LP-1760. [Google Scholar] [CrossRef] [Green Version]
- Paital, B.; Bal, A.; Rivera-Ingraham, G.A.; Jehan-Herve, L. Increasing frequency of large-scale die-off events in the Bay of Bengal: Reasoning, perspectives and future approaches. Indian J. Mar. Sci. 2018, 47, 2135–2146. [Google Scholar]
- Politis, S.N.; Mazurais, D.; Servili, A.; Zambonino-Infante, J.L.; Miest, J.J.; Tomkiewicz, J.; Butts, I.A.E. Salinity reduction benefits European eel larvae: Insights at the morphological and molecular level. PLoS ONE 2018, 13, e0198294. [Google Scholar] [CrossRef] [Green Version]
- Dominoni, D.M.; Halfwerk, W.; Baird, E.; Buxton, R.T.; Fernandez-Juricic, E.; Fristrup, K.M.; McKenna, M.F.; Mennitt, D.J.; Perkin, E.K.; Seymoure, B.M.; et al. Why conservation biology can benefit from sensory ecology. Nat. Ecol. Evol. 2020, 4, 502–511. [Google Scholar] [CrossRef]
- Chuang, M.F.; Cheng, Y.J.; Andersen, D.; Borzée, A.; Wu, C.S.; Chang, Y.M.; Yang, Y.; Jang, Y.; Kam, Y.C. Increasing salinity stress decreases the thermal tolerance of amphibian tadpoles in coastal areas of Taiwan. Sci. Rep. 2022, 12, 9014. [Google Scholar] [CrossRef] [PubMed]
- Jamei, M.; Karbasi, M.; Malik, A.; Abualigah, L.; Islam, A.R.M.T.; Yaseen, Z.M. Computational assessment of groundwater salinity distribution within coastal multi-aquifers of Bangladesh. Sci. Rep. 2022, 12, 11165. [Google Scholar] [CrossRef] [PubMed]
- Scarponi, D.; Nawrot, R.; Azzarone, M.; Pellegrini, C.; Gamberi, F.; Trincardi, F.; Kowalewski, M. Resilient biotic response to long-term climate change in the Adriatic Sea. Glob. Chang. Biol. Bioenergy 2022, 28, 4041–4053. [Google Scholar] [CrossRef] [PubMed]
- Perkins, S.E.; King, A.D.; Alexander, L. Increased simulated risk of the hot Australian summer of 2012/13 due to anthropogenic activity as measured by heat wave frequency and intensity. Bull. Am. Meteorol. Soc. 2014, 95, S34–S37. [Google Scholar]
- Toan, T.Q. Climate Change and Sea Level Rise in the Mekong Delta: Flood, Tidal Inundation, Salinity Intrusion, and Irrigation Adaptation Methods. In Coastal Disasters and Climate Change in Vietnam; Elsevier: Amsterdam, The Netherlands, 2014; pp. 199–218. [Google Scholar]
- Du, Y.; Zhang, Y.; Shi, J. Relationship between sea surface salinity and ocean circulation and climate change. Sci. China Earth Sci. 2019, 62, 771–782. [Google Scholar] [CrossRef]
- Schmitt, R.W. Salinity and the global water cycle. Oceanography 2008, 21, 12–19. Available online: https://www.jstor.org/stable/24860148 (accessed on 15 October 2022). [CrossRef]
- IPCC. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. In Climate Change 2007: Impacts, Adaptation and Vulnerability; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- IPCC. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. In Summary for Policymakers. Climate Change 2014: Impacts, Adaptation, and Vulnerability; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Erol, A.; Randyr, T.O. Climatic change impacts on the ecohydrology of Mediterranean watersheds. Clim. Chang. 2012, 114, 319–341. [Google Scholar] [CrossRef]
- Al-Shammiri, M. Evaporation rate as a function of water salinity. Desalination 2002, 150, 189–203. [Google Scholar] [CrossRef]
- Obianyo, J.I. Effect of salinity on evaporation and the water cycle. Emerg. Sci. 2019, 3, 255–262. [Google Scholar] [CrossRef]
- Behara, A.; Vinayachandran, P.N.; Shankar, D. Influence of rainfall over eastern Arabian Sea on its salinity. J. Geophys. Res. Oceans 2019, 124, 5003–5020. [Google Scholar] [CrossRef]
- Weatherly, J.W.; Walsh, J.E. The effects of precipitation and river runoff in a coupled ice-ocean model of the Arctic. Clim. Dyn. 1996, 12, 785–798. [Google Scholar] [CrossRef]
- Sciencelearn. Ocean Salinity Science Learning Hub. 2022. Available online: https://www.sciencelearn.org.nz/resources/686-ocean-salinity (accessed on 27 September 2022).
- Murray, J. On the effects of winds on the distribution of temperature in the sea-and fresh-water lochs of the West of Scotland. Scott. Geogr. Mag. 1888, 4, 345–347. [Google Scholar] [CrossRef]
- Skliris, N.; Zika, J.D.; Herold, L.; Josey, S.A.; Marsh, R. Mediterranean Sea water budget long-term trend inferred from salinity observations. Clim. Dyn. 2018, 51, 2857–2876. [Google Scholar] [CrossRef] [Green Version]
- Karsten, U. Research note: Salinity tolerance of Arctic kelps from Spitsbergen. Psychol. Res. 2007, 55, 257–262. [Google Scholar] [CrossRef]
- Velings, N.M.; Mestdagh, M.M. Physico-chemical properties of alginate gel beads. Polym. Gels Netw. 1995, 3, 311–330. [Google Scholar] [CrossRef]
- Dierssen, H.M.; Smith, R.C.; Vernet, M. Glacial meltwater dynamics in coastal waters west of the Antarctic Peninsula. Proc. Natl. Acad. Sci. USA 2002, 99, 1790–1795. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Currell, M.J. Review of drivers and threats to coastal groundwater quality in China. Sci. Total Environ. 2022, 806, 150913. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.; Xie, G.; Deng, J.; Shao, K.; Hu, Y.; He, J.; Zhang, J.; Gao, G. Effects of climate change and anthropogenic activities on lake environmental dynamics: A case study in Lake Bosten Catchment, NW China. J. Environ. Manag. 2022, 319, 115764. [Google Scholar] [CrossRef]
- Thai-Hoang, L.; Thong, T.; Loc, H.T.; Van, P.T.T.; Thuy, P.T.P.; Thuoc, T.L. Influences of anthropogenic activities on water quality in the Saigon River, Ho Chi Minh City. J. Water Health 2022, 20, 491–504. [Google Scholar] [CrossRef]
- Lorrain-Soligon, L.; Bichet, C.; Robin, F.; Brischoux, F. From the field to the lab: Physiological and behavioural consequences of environmental salinity in a coastal frog. Front. Physiol. 2022, 13, 919165. [Google Scholar] [CrossRef]
- Hapsari, K.A.; Jennerjahn, T.; Nugroho, S.H.; Yulianto, E.; Behling, H. Sea level rise and climate change acting as interactive stressors on development and dynamics of tropical peatlands in coastal Sumatra and South Borneo since the Last Glacial Maximum. Glob. Chang. Biol. 2022, 28, 3459–3479. [Google Scholar] [CrossRef] [PubMed]
- Cai, F.; Cao, C.; Qi, H.; Su, X.; Lei, G.; Liu, J.; Zhao, S.; Liu, G.; Zhu, K. Rapid migration of mainland China’s coastal erosion vulnerability due to anthropogenic changes. J. Environ. Manag. 2022, 319, 115632. [Google Scholar] [CrossRef]
- Martínez-Megías, C.; Rico, A. Biodiversity impacts by multiple anthropogenic stressors in Mediterranean coastal wetlands. Sci. Total Environ. 2022, 818, 151712. [Google Scholar] [CrossRef] [PubMed]
- Teunen, L.; De Jonge, M.; Malarvannan, G.; Covaci, A.; Belpaire, C.; Focant, J.F.; Blust, R.; Bervoets, L. Effect of abiotic factors and environmental concentrations on the bioaccumulation of persistent organic and inorganic compounds to freshwater fish and mussels. Sci. Total Environ. 2021, 799, 149448. [Google Scholar] [CrossRef]
- Herczeg, A.L.; Dogramaci, S.; Leaney, F.W.J. Origin of dissolved salts in a large, semi-arid groundwater system: Murray Basin, Australia. Mar. Freshw. Res. 2001, 52, 41–52. [Google Scholar] [CrossRef]
- Herbert, E.R.; Boon, P.; Burgin, A.J.; Neubauer, S.C.; Franklin, R.B.; Ardón, M.; Hopfensperger, K.N.; Lamers, L.P.M.; Gell, P. A global perspective on wetland salinization: Ecological consequences of a growing threat to freshwater wetlands. Ecosphere 2015, 6, art206. [Google Scholar] [CrossRef]
- Palmer, C.G.; Muller, W.J.; Gordon, A.K.; Scherman, P.A.; Davies-Coleman, H.D.; Pakhomova, L.; De Kock, E. The development of a toxicity database using freshwater macroinvertebrates, and its application to the protection of South African water resources. S. Afr. J. Sci. 2004, 100, 643–650. [Google Scholar]
- Cañedo-Argüelles, M.; Rieradevall, M. Disturbance caused by freshwater releases of different magnitude on the aquatic macroinvertebrate communities of two coastal lagoons. Estuar. Coast. Shelf Sci. 2010, 88, 190–198. [Google Scholar] [CrossRef]
- Skjærven, K.H.; Oveland, E.; Mommens, M.; Samori, E.; Saito, T.; Adam, A.-C.; Espe, M. Out-of-season spawning affects the nutritional status and gene expression in both Atlantic salmon female broodstock and their offspring. Comp. Biochem. Physiol. A 2020, 247, 110717. [Google Scholar] [CrossRef]
- Huang, Y.; Wu, D.; Li, Y.; Chen, Q.; Zhao, Y. Characterization and expression of arginine kinase 2 from Macrobrachium nipponense in response to salinity stress. Dev. Comp. Immunol. 2020, 113, 103804. [Google Scholar] [CrossRef]
- Loretz, C.A.; Pollina, C.; Hyodo, S.; Takei, Y.; Chang, W.; Shoback, D. cDNA cloning and functional expression of a Ca2+-sensing receptor with truncated C-terminal tail from the Mozambique Tilapia (Oreochromis mossambicus). J. Biol. Chem. 2004, 279, 53288–53297. [Google Scholar] [CrossRef] [PubMed]
- Marshall, W.S.; Ossum, C.G.; Hoffmann, E.K. Hypotonic shock mediation by pMAPK, JNK, PKC, FAK, OSR1 and SPAK in osmosensing chloride secreting cells of killifish opercular epithelium. J. Exp. Biol. 2005, 208, LP-1077. [Google Scholar] [CrossRef] [PubMed]
- Fiol, D.F.; Chan, S.Y.; Kültz, D. Regulation of osmotic stress transcription factor 1 (Ostf1) in tilapia (Oreochromis mossambicus) gill epithelium during salinity stress. J. Exp. Biol. 2006, 209, 3257. [Google Scholar] [CrossRef] [Green Version]
- Choi, C.Y.; An, K.W. Cloning and expression of Na+/K+- ATPase and osmotic stress transcription factor 1 mRNA in black porgy, Acanthopagrus schlegeli during osmotic stress. Comp. Biochem. Physiol. B 2008, 149, 91–100. [Google Scholar] [CrossRef]
- Tse, W.K.F.; Chow, S.C.; Wong, C.K.C. The cloning of eel osmotic stress transcription factor and the regulation of its expression in primary gill cell culture. J. Exp. Biol. 2008, 211, 1964–1968. [Google Scholar] [CrossRef] [Green Version]
- Chow, S.C.; Ching, L.Y.; Wong, A.M.F.; Wong, C.K.C. Cloning and regulation of expression of the Na+-Cl− taurine transporter in gill cells of freshwater Japanese eels. J. Exp. Biol. 2009, 212, 3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breves, J.P.; Hasegawa, S.; Yoshioka, M.; Fox, B.K.; Davis, L.K.; Vijayan, M.M. Acute salinity challenges in Mozambique and Nile Tilapia: Differential responses of plasma prolactin growth hormone and branchial expression of ion transporters. Gen. Comp. Endocrinol. 2010, 167, 135–142. [Google Scholar] [CrossRef] [PubMed]
- McGuire, A.; Aluru, N.; Takemura, A.; Weil, R.; Wilson, J.M.; Vijayan, M.M. Hyperosmotic shock adaptation by cortisol involves upregulation of branchial osmotic stress transcription factor 1 gene expression in Mozambique Tilapia. Gen. Comp. Endocrinol. 2010, 165, 321–329. [Google Scholar] [CrossRef]
- Tse, W.K.; Lai, K.P.; Takei, Y. Osmotic stress transcription factor 1b (Ostf1b/TSC 222) triggers hyperosmotic responses of different ion transporters in medaka gill and human embryonic kidney cells via the JNK signalling pathway. Int. J. Biochem. Cell Biol. 2011, 43, 1764–1775. [Google Scholar] [CrossRef]
- Hwang, P.P.; Chou, M.Y. Zebrafish as an animal model to study ion homeostasis. Pflug. Arch. 2013, 465, 1233–1247. [Google Scholar] [CrossRef] [Green Version]
- Yancey, P.H. Organic osmolytes as compatible, metabolic and counteracting cytoprotectants in high osmolarity and other stresses. J. Exp. Biol. 2005, 208, 2819–2830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salin, K.; Auer, S.K.; Rey, B.; Selman, C.; Metcalfe, N.B. Variation in the link between oxygen consumption and ATP production, and its relevance for animal performance. Proc. R. Soc. B 2015, 282, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Bradley, T.J. Animal Osmoregulation; Oxford University Press: Oxford, UK, 2008. [Google Scholar] [CrossRef]
- Belli, N.M.; Faleiros, R.O.; Firmino, K.C.; Leone, F.A.; Mc Namara, J.C.; Furriel, R.P. Na+, K+ ATPase activity and epithelial interfaces in gills of the freshwater shrimp Macrobrachium amazonicum (Decapoda, Palaemonidae). Comp. Biochem. Physiol. A 2009, 152, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Mishra, P.; Paital, B.; Jena, S.; Swain, S.S.; Kumar, S.; Yadav, M.K.; Chainy, G.B.N.; Samanta, L. Possible activation of NRF2 by Vitamin E/Curcumin against altered thyroid hormone induced oxidative stress via NFĸB/AKT/mTOR/KEAP1 signalling in rat heart. Sci. Rep. 2019, 9, 7408. [Google Scholar] [CrossRef] [Green Version]
- McGraw, K.J.; Cohen, A.A.; Costantini, D.; Hõrak, P. The ecological significance of antioxidants and oxidative stress: A marriage between mechanistic and functional perspectives. Funct. Ecol. 2010, 24, 947–949. [Google Scholar] [CrossRef]
- Sanz, A.; Trenzado, C.E.; Castro, H.B.; López-Rodríguez, M.J.; de Figueroa, J.T. Relationship between brain and liver oxidative state and maximum lifespan potential of different fish species. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 16, 358–364. [Google Scholar] [CrossRef]
- Zhang, M.; Li, L.; Liu, Y.; Gao, X. Effects of a sudden drop in salinity on Scapharca subcrenata antioxidant defenses and metabolism determined using LC-MS non-targeted metabolomics. Sci. Rep. 2020, 10, 7324. [Google Scholar] [CrossRef]
- Islam, M.J.; Kunzmann, A.; Thiele, R.; Slater, M.J. Effects of extreme ambient temperature in European seabass, Dicentrarchus labrax acclimated at different salinities: Growth performance, metabolic and molecular stress responses. Sci. Total Environ. 2020, 735, 139371. [Google Scholar] [CrossRef]
- Patrick, M.L.; Bradley, T.J. The physiology of salinity tolerance in larvae of two species of Culex mosquitoes: The role of compatible solutes. J. Exp. Biol. 2000, 203, 821–830. [Google Scholar] [CrossRef]
- Xie, Y.; Zhang, P.; Zhang, L. Genome-wide transcriptional responses of marine nematode Litoditis marina to hyposaline and hypersaline stresses. Front. Physiol. 2021, 12, 672099. [Google Scholar] [CrossRef]
- Hawkins, L.J.; Storey, K.B. Advances and applications of environmental stress adaptation research. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2020, 240, 110623. [Google Scholar] [CrossRef]
- Cutler, C.P.; Cramb, G. Branchial expression of an aquaporin 3 (AQP-3) homologue is downregulated in the European eel Anguilla anguilla following seawater acclimation. J. Exp. Biol. 2002, 205, 2643–2651. [Google Scholar] [CrossRef] [PubMed]
- Perry, S.F.; Shahsavarani, A.; Georgalis, T.; Bayaa, M.; Furimsky, M.; Thomas, S.L.Y. Channels, pumps and exchangers in the gill and kidney of freshwater fishes: Their role in ionic and acid-base regulation. J. Exp. Zool. A Comp. Exp. Biol. 2003, 300A, 53–62. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.R.A.; Estêvão, M.D.; Fuentes, J.; Cardoso, J.C.; Fabra, M.; Passos, A.L.; Detmers, F.J.; Deen, P.M.T.; Cerdà, J.; Power, D.M. Isolation of a novel aquaglyceroporin from a marine teleost (Sparus auratus): Function and tissue distribution. J. Exp. Biol. 2004, 207, 1217–1227. [Google Scholar]
- Evans, D.H.; Piermarini, P.M.; Choe, K.P. The multifunctional fish gill: Dominant site of gas exchange, osmoregulation, acid-base regulation, and excretion of nitrogenous waste. Physiol. Rev. 2005, 85, 97–177. [Google Scholar] [CrossRef]
- Ern, R.; Esbaugh, A.J. Effects of salinity and hypoxia-induced hyperventilation on oxygen consumption and cost of osmoregulation in the estuarine red drum (Sciaenops ocellatus). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 222, 52–59. [Google Scholar] [CrossRef]
- Fang, Y.; Chan, V.K.; Hines, C.W.; Stiller, K.T.; Richards, J.G.; Brauner, C.J. The effects of salinity and photoperiod on aerobic scope, hypoxia tolerance and swimming performance of coho salmon (Oncorhynchus kisutch) reared in recirculating aquaculture systems. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 231, 82–90. [Google Scholar] [CrossRef]
- Han, J.; Lee, K.W. Influence of salinity on population growth, oxidative stress and antioxidant defense system in the marine monogonont rotifer Brachionus plicatilis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2020, 250, 110487. [Google Scholar] [CrossRef]
- Pequignat, E. A kinetic autoradiographic study of the direct assimilation of amino acids and glucose by organs of the mussel Mytilus edulis. Mar. Biol. 1973, 19, 227–244. [Google Scholar] [CrossRef]
- Burton, R.S. Incorporation of 14C-bicarbonate into the free amino acid pool during hyperosmotic stress in an intertidal copepod. J. Exp. Zool. 1986, 238, 55–61. [Google Scholar] [CrossRef]
- Bishop, J.S.; Burton, R.S. Amino acid synthesis during hyperosmotic stress in Penaeus aztecus post larvae. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1993, 106, 49–56. [Google Scholar] [CrossRef]
- McAllen, R.; Walker, D.; Taylor, A. The environmental effects of salinity and temperature on the oxygen consumption and total body osmolality of the marine flatworm Procerodes littoralis. J. Exp. Mar. Biol. Ecol. 2002, 268, 103–113. [Google Scholar] [CrossRef]
- Khlebovich, V.V. Levels of Homeostasis, 2nd ed.; Priroda: Moscow, Russia, 2007; pp. 61–65. [Google Scholar]
- Xiao, S.; Wong, N.K.; Li, J.; Lin, Y.; Zhang, Y.; Ma, H.; Mo, R.; Zhang, Y.; Yu, Z. Analysis of in situ transcriptomes reveals divergent adaptive response to hyper- and hypo-salinity in the Hong Kong oyster, Crassostrea hongkongensis. Front. Physiol. 2018, 9, 1491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, Y.; Liu, X. Expression of Na+/K+-ATPase was affected by salinity change in Pacific abalone Haliotis discus hannai. Front. Physiol. 2018, 9, 1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carregosa, V.; Velez, C.; Soares, A.M.; Figueira, E.; Freitas, R. Physiological and biochemical responses of three Veneridae clams exposed to salinity changes. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2014, 177, 1–9. [Google Scholar] [CrossRef]
- Matozzo, V.; Binelli, A.; Parolini, M.; Previato, M.; Masiero, L.; Finos, L.; Bressan, M.; Marin, M.G. Biomarker responses in the clam Ruditapes philippinarum and contamination levels in sediments from seaward and landward sites in the Lagoon of Venice. Ecol. Indic. 2012, 19, 191–205. [Google Scholar] [CrossRef]
- Panda, F.; Pati, S.G.; Bal, A.; Das, K.; Samanta, L.; Paital, B. Control of invasive apple snails and their use as pollutant ecotoxic indicators: A review. Environ. Chem. Lett. 2021, 19, 4627–4653. [Google Scholar] [CrossRef]
- Panda, F.; Pati, S.G.; Bal, A.; Mathur, S.; Nirmaladevi, R.; Paital, B. Temporal morphometric analyses of Pila globosa in India for its use in aquaculture and food industry. J. Basic Appl. Zool. 2021, 82, 17. [Google Scholar] [CrossRef]
- Pati, S.G.; Panda, F.; Samanta, L.; Paital, B. Spatio-temporal changes in oxidative stress physiology parameters in apple snail Pila Globosa as a function of soil Mg, Ca, organic carbon and aquatic physico-chemical factors. Environ. Geochem. Health 2022. [Google Scholar] [CrossRef]
- da Silva, A.Z.; Zanette, J.; Ferreira, J.F.; Guzenski, J.; Marques, M.R.F.; Bainy, A.C.D. Effects of salinity on biomarker responses in Crassostrea rhizophorae (Mollusca, Bivalvia) exposed to diesel oil. Ecotoxicol. Environ. Saf. 2005, 62, 376–382. [Google Scholar] [CrossRef]
- Pati, S.G.; Panda, F.; Jena, S.; Sahoo, D.K.; Paital, B. Effects of soil trace metals, organic carbon load and physicochemical stressors on active oxygen species metabolism in Scylla Serrata sampled along the bay of Bengal in Odisha State, India. Front. Environ. Sci. 2022, 10, 1733. [Google Scholar] [CrossRef]
- Mo, Z.; Li, L.; Ying, L.; Xiaolong, G. Effects of sudden drop in salinity on osmotic pressure regulation and antioxidant defense mechanism of Scapharca subcrenata. Front. Physiol. 2020, 11, 884. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.; Silvestro, S.; Coppola, F.; Meucci, V.; Battaglia, F.; Intorre, L.; Soares, A.M.V.M.; Pretti, C.; Faggio, C. Combined effects of salinity changes and salicylic acid exposure in Mytilus galloprovincialis. Sci. Total Environ. 2020, 715, 136804. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.M.; Gonzalez, C.M. Physiological responses of the Chilean scallop Argopecten purpuratus to decreasing salinities. Aquaculture 1998, 167, 315–327. [Google Scholar] [CrossRef]
- Coughlan, B.M.; Moroney, G.A.; Pelt, F.N.A.M.; O’Brien, N.M.; Davenport, J.; O’Halloran, J. The effects of salinity on the Manila clam (Ruditapes philippinarum) using the neutral red retention assay with adapted physiological saline solutions. Mar. Pollut. Bull. 2009, 58, 1680–1684. [Google Scholar] [CrossRef]
- Bozza, D.C.; Freire, C.A.; Prodocimo, V. Osmo-ionic regulation and carbonic anhydrase, Na+/K+-ATPase and V-H+-ATPase activities in gills of the ancient freshwater crustacean Aegla schmitti (Anomura) exposed to high salinities. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 231, 201–208. [Google Scholar] [CrossRef]
- Liu, Z.M.; Zhu, X.L.; Lu, J.; Cai, W.J.; Ye, Y.P.; Lv, Y.P. Effect of high temperature stress on heat shock protein expression and antioxidant enzyme activity of two morphs of the mud crab Scylla paramamosain. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2018, 223, 10–17. [Google Scholar] [CrossRef]
- Dandapat, J.; Chainy, G.B.N.; Janardhana, R.K. Dietary vitamin-E modulates antioxidant defence system in giant freshwater prawn, Macrobrachium rosenbergii. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2000, 127, 101–115. [Google Scholar] [CrossRef]
- Arun, S.; Subramanian, P. Antioxidant enzymes activity in subcellular fraction of freshwater prawns M. malcolmsonii and M. lamarrei lamarrei. Appl. Biochem. Biotechnol. 1998, 75, 187–192. [Google Scholar] [CrossRef]
- Pan, L.; Zhang, H. Metallothionein, antioxidant enzymes and DNA strand breaks as biomarkers of Cd exposure in a marine crab, Charybdis japonica. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 144, 67–75. [Google Scholar] [CrossRef]
- Maciel, F.E.; Rosa, C.E.; Santos, E.A.; Monserrat, J.M.; Nery, L.E.M. Daily variations in oxygen consumption, antioxidant defenses, and lipid peroxidation in the gills and hepatopancreas of an estuarine crab. Can. J. Zool. 2004, 82, 1871–1877. [Google Scholar] [CrossRef]
- de Oliveira, U.O.; Belló-Klein, A.; Kucharski, L.C. Oxidative balance and immunodetection of antioxidant enzymes in the hepatopancreas of the crab Chasmagnathus granulata subjected to anoxia and reoxygenation. Can. J. Zool. 2006, 84, 677–684. [Google Scholar] [CrossRef]
- Borković, S.S.; Pavlović, S.Z.; Kovačević, T.B.; Štajn, A.Š.; Petrović, V.M.; Saičić, Z.S. Antioxidant defence enzyme activities in hepatopancreas, gills and muscle of Spiny cheek crayfish (Orconectes limosus) from the river Danube. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2008, 147, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, A.P.; Oliveira, P.C.; Guilhermino, L.; Guimarães, L. Effects of salinity stress on neurotransmission, energy metabolism, and anti-oxidant biomarkers of Carcinus maenas from two estuaries of the NW Iberian Peninsula. Mar. Biol. 2012, 159, 2061–2074. [Google Scholar] [CrossRef]
- Binyon, J. Osmotic and hydrostatic permeability of the integument of the starfish Asterias rubens. J. Mar. Biol. Assoc. UK 1980, 60, 627–630. [Google Scholar] [CrossRef]
- Shirley, T.C.; Stickle, W.B. Responses of Leptasterias hexactis (Echinodermata: Asteroidea) to low salinity. Mar. Biol. 1982, 69, 155–163. [Google Scholar] [CrossRef]
- Talbot, T.D.; Lawrence, J.M. The effect of salinity on respiration, excretion, regeneration and production in Ophiophragmus filograneus (Echinodermata: Ophiuroidea). J. Exp. Mar. Biol. Ecol. 2002, 275, 1–14. [Google Scholar] [CrossRef]
- Yuan, X.; Yang, H.; Wang, L.; Zhou, Y.; Gabr, H.R. Effects of salinity on energy budget in pond-cultured sea cucumber Apostichopus japonicus (Selenka) (Echinodermata: Holothuroidea). Aquaculture 2010, 306, 348–351. [Google Scholar] [CrossRef]
- Castellano, G.C.; Santos, I.A.; Freire, C.A. Maintenance of ionic gradients and tissue hydration in the intertidal sea cucumber Holothuria grisea under hypo- and hyper-salinity challenges. J. Mar. Biol. Assoc. UK 2018, 98, 325–332. [Google Scholar] [CrossRef]
- Yu, Z.; Zhou, Y.; Yang, H.; Hu, C. Bottom culture of the sea cucumber Apostichopus japonicus Selenka (Echinodermata: Holothuroidea) in a fish farm, southern China. Aquac. Res. 2014, 45, 1434–1441. [Google Scholar] [CrossRef]
- Paul, B.K.; Rashid, H. (Eds.) Chapter Five—Salinity Intrusion and Impacts. In Climatic Hazards in Coastal Bangladesh; Butterworth-Heinemann: Boston, MA, USA, 2017; pp. 153–182. [Google Scholar] [CrossRef]
- Bray, W.A.; Lawrence, A.L.; Leung-Trujillo, J.R. The effect of salinity on growth and survival of Penaeus vannamei, with observations on the interaction of IHHN virus and salinity. Aquaculture 1994, 122, 133–146. [Google Scholar] [CrossRef]
- Chen, J.C.; Chen, W.C. Temperature tolerance of Haliotis diversicolor supertexta at different salinity and temperature levels. Comp. Biochem. Physiol. A 1999, 124, 73–80. [Google Scholar] [CrossRef]
- Imsland, A.K.; Foss, A.; Gunnarsson, S.; Berntssen, M.H.; FitzGerald, R.; Bonga, S.W.; Ham, E.V.; Nævdal, G.; Stefansson, S.O. The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus). Aquaculture 2001, 198, 353–367. [Google Scholar] [CrossRef]
- Ip, Y.K.; Hiong, K.C.; Wong, S.Z.; Ching, B.; Chen, X.L.; Soh, M.M.; Chng, Y.R.; Ong, J.L.; Wilson, J.M.; Chew, S.F. Branchial Na(+):K(+):2Cl(-) cotransporter 1 and Na(+)/K(+)-ATPase α-subunit in a brackish water-type ionocyte of the euryhaline freshwater white-rimmed stingray, Himantura signifer. Front. Physiol. 2013, 4, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Hou, X.; Xue, X.; Zhu, X.; Chen, Y.; Yang, Z. Interactive effects of temperature and salinity on the survival, oxidative stress, and Na+/K+- ATPase activity of newly hatched obscure puffer (Takifugu obscurus) larvae. Fish Physiol. Biochem. 2019, 45, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Huang, J.J.; Yeh, C.Y.; Tang, C.H.; Hwang, L.Y.; Lee, T.H. Salinity effects on strategies of glycogen utilization in livers of euryhaline milkfish (Chanos chanos) under hypothermal stress. Front. Physiol. 2018, 9, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eissa, N.; Wang, H.P.; Yao, H.; Shen, Z.G.; Shaheen, A.A.; Abou-ElGheit, E.N. Expression of Hsp70, Igf1, and three oxidative stress biomarkers in response to handling and salt treatment at different water temperatures in yellow perch, Perca flavescens. Front. Physiol. 2017, 8, 683. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deane, E.E.; Luk, J.C.; Woo, N.Y. Aquaporin 1a expression in gill, intestine, and kidney of the euryhaline silver sea bream. Front. Physiol. 2011, 2, 39. [Google Scholar] [CrossRef] [Green Version]
- Skomal, G.B.; Mandelman, J.W. The physiological response to anthropogenic stressors in marine elasmobranch fishes: A review with a focus on the secondary response. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 162, 146–155. [Google Scholar] [CrossRef]
- Kim, J.H.; Park, H.J.; Kim, K.W.; Hwang, I.K.; Kim, D.H.; Oh, C.W.; Lee, J.S.; Kang, J.C. Growth performance, oxidative stress and non-specific immune responses in juvenile sablefish Anoplopoma fimbria by changes of water temperature and salinity. Fish Physiol. Biochem. 2017, 43, 1421–1431. [Google Scholar] [CrossRef]
- Wang, L.; Gallagher, E.P. Role of Nrf2 antioxidant defense in mitigating cadmium-induced oxidative stress in the olfactory system of zebrafish. Toxicol. Appl. Pharmacol. 2013, 266, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Yazlak, H.; Orhan, C.; Tuzcu, M.; Akdemir, F.; Sahin, N. The effect of lycopene on antioxidant status in rainbow trout (Oncorhynchus mykiss) reared under high stocking density. Aquaculture 2014, 418, 132–138. [Google Scholar] [CrossRef]
- Katoh, Y.; Iida, K.; Kang, M.I.; Kobayashi, A.; Mizukami, M.; Tong, K.I.; McMahon, M.; Hayes, J.D.; Itoh, K.; Yamamoto, M. Evolutionary conserved N-terminal domain of Nrf2 is essential for the Keap1-mediated degradation of the protein by proteasome. Arch. Biochem. Biophys. 2005, 433, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Ai, C.X.; Wang, Y.H.; Zhang, J.S.; Wu, C.W. Abrupt salinity stress induces oxidative stress via the Nrf2-Keap1 signaling pathway in large yellow croaker Pseudosciaena crocea. Fish Physiol. Biochem. 2017, 43, 955–964. [Google Scholar] [CrossRef]
- Hu, D.; Pan, L.; Zhao, Q.; Ren, Q. Transcriptomic response to low salinity stress in gills of the Pacific white shrimp, Litopenaeus vannamei. Mar. Genom. 2015, 24, 297–304. [Google Scholar] [CrossRef]
- Martínez-Álvarez, R.M.; Hidalgo, M.C.; Domezain, A.; Morales, A.E.; García-Gallego, M.; and Sanz, A. Physiological changes of sturgeon Acipenser naccarii caused by increasing environmental salinity. J. Exp. Biol. 2002, 205, 3699–3706. [Google Scholar] [CrossRef] [PubMed]
- Shukry, M.; Abd EL-Kader, M.F.; Hendam, B.M.; Dawood, M.; Farrag, F.A.; Aboelenin, S.M.; Soliman, M.M.; Abdel-Latif, H.M. Dietary Aspergillus oryzae modulates serum biochemical indices, immune responses, oxidative stress and transcription of HSP 70 and cytokine genes in Nile Tilapia exposed to salinity stress. Animals 2021, 11, 1621. [Google Scholar] [CrossRef]
- Loro, V.L.; Jorge, M.B.; Silva, K.R.; Wood, C.M. Oxidative stress parameters and antioxidant response to sublethal waterborne zinc in a euryhaline teleost Fundulus heteroclitus: Protective effects of salinity. Aqua. Toxicol. 2012, 110, 187–193. [Google Scholar] [CrossRef]
- Blewett, T.A.; Ransberry, V.E.; McClelland, G.B.; Wood, C.M. Investigating the mechanisms of Ni uptake and sub-lethal toxicity in the Atlantic killifish Fundulus heteroclitus in relation to salinity. Environ. Pollut. 2016, 211, 370–381. [Google Scholar] [CrossRef]
- Ransberry, V.E.; Blewett, T.A.; Mc Clelland, G.B. The oxidative stress response in freshwater-acclimated killifish (Fundulus heteroclitus) to acute copper and hypoxia exposure. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 179, 11–18. [Google Scholar] [CrossRef]
- Glover, C.N.; Urbina, M.A.; Harley, R.A.; Lee, J.A. Salinity- dependent mechanisms of copper toxicity in the galaxiid fish, Galaxias maculatus. Aqua. Toxicol. 2016, 174, 199–207. [Google Scholar] [CrossRef]
- Hoffmann, E.K.; Lambert, I.H.; Pedersen, S.F. Physiology of cell volume regulation in vertebrates. Physiol. Rev. 2007, 89, 193–277. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Xu, G.; Ge, X.; Liu, B.; Xu, P.; Song, C.; Zhou, Q.; Zhang, H.; Zhang, W.; Shan, F. The effects of crowding stress on the growth, physiological response, and gene expression of the Nrf2-keapsignaling pathway in blunt snout bream (Megalobrama amblycephala) reared under in-pond raceway conditions. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2019, 231, 19–29. [Google Scholar] [CrossRef]
- Gómez-Angelats, M.; Cidlowski, J.A. Invited review: Cell volume control and signal transduction in apoptosis. J. Toxicol. Pathol. 2002, 30, 541–551. [Google Scholar] [CrossRef] [Green Version]
- Gutierre, S.M.M.; Vitule, J.R.S.; Freire, C.A.; Prodocimo, V. Physiological tools to predict invasiveness and spread via estuarine bridges: Tolerance of Brazilian native and worldwide introduced freshwater fishes to increased salinity. Mar. Freshw. Res. 2014, 65, 425–436. [Google Scholar] [CrossRef]
- Kültz, D. Osmotic, Ionic and Nitrogenous-Waste Balance|Osmosensing. In Encyclopedia of Fish Physiology; Academic Press: San Diego, CA, USA, 2011; pp. 1373–1380. [Google Scholar] [CrossRef]
- Kültz, D. 2—Osmosensing. In Euryhaline Fishes; McCormick, S.D., Farrell, A.P., Brauner, C.J., Eds.; Academic Press: Cambridge, MA, USA, 2012; pp. 45–68. [Google Scholar] [CrossRef]
- Foskett, J.K.; Bern, H.A.; Machen, T.E.; Conner, M. Chloride cells and the hormonal control of teleost fish osmoregulation. J. Exp. Biol. 1983, 106, 255–281. [Google Scholar] [CrossRef]
- Kültz, D.; Avila, K. Mitogen-activated protein kinases are in vivo transducers of osmosensory signals in fish gill cells. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 821–829. [Google Scholar] [CrossRef]
- Behrens, J.W.; van Deurs, M.; Christensen, E.A.F. Evaluating dispersal potential of an invasive fish by the use of aerobic scope and osmoregulation capacity. PLoS ONE 2017, 12, e0176038. [Google Scholar] [CrossRef] [Green Version]
- Laverty, G.; Skadhauge, E. Adaptation of teleosts to very high salinity. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 163, 1–6. [Google Scholar] [CrossRef]
- Karnaky, K.J. Structure and function of the chloride cell of Fundulus heteroclitus and other teleosts. Am. Zool. 1986, 26, 209–224. [Google Scholar] [CrossRef] [Green Version]
- Kültz, D.; Onken, H. Long-term acclimation of the teleost Oreochromis mossambicus to various salinities: Two different strategies in mastering hypertonic stress. Mar. Biol. 1993, 117, 527–533. [Google Scholar] [CrossRef]
- Verlecar, X.N.; Jena, K.B.; Chainy, G.B.N. Biochemical markers of oxidative stress in Perna viridis exposed to mercury and temperature. Chem. Biol. Interact. 2007, 167, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Chainy, G.B.N.; Paital, B.; Dandapat, J. An overview of seasonal changes in oxidative stress and antioxidant defence parameters in some invertebrate and vertebrate species. Scientifica 2016, 2016, 6126570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khamidov, M.; Ishchanov, J.; Hamidov, A.; Donmez, C.; Djumaboev, K. Assessment of soil salinity changes under the climate change in the Khorezm region, Uzbekistan. Int. J. Environ. Res. Public Health 2022, 19, 8794. [Google Scholar] [PubMed]
- Mitra, A.; Banerjee, K.; Sengupta, K.; Gangopadhyay, A. Pulse of climate change in Indian sundarbans: A myth or reality? Natl. Acad. Sci. Lett. 2009, 32, 19–25. [Google Scholar]
- Banerjee, K.; Gatti, R.C.; Mitra, A. Climate change-induced salinity variation impacts on a Stenoecious mangrove species in the Indian sundarbans. Ambio 2017, 46, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Moss, B.; Kosten, S.; Meerhoff, M.; Battarbee, R.W.; Jeppesen, E.; Mazzeo, N.; Havens, K.; Lacerot, G.; Liu, Z.; De Meester, L.; et al. Allied attack: Climate change and eutrophication. Inland Waters 2011, 1, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Guerrero, R.A.; Acha, E.M.; Framiñan, M.B.; Lasta, C.A. Physical oceanography of the Río de la Plata Estuary, Argentina. Cont. Shelf Res. 1997, 17, 727–742. [Google Scholar] [CrossRef]
- Morcillo, M.; Chico, B.; Mariaca, L.; Otero, E. Salinity in marine atmospheric corrosion: Its dependence on the wind regime existing in the site. Corros. Sci. 2000, 42, 91–104. [Google Scholar] [CrossRef]
- Chang, X.; Wang, S.; Chen, H.; Guan, X. Analysis of the main factors affecting salinity in Wuliangsuhai lake. IOP Conf. Ser. Earth Environ. Sci. 2021, 658, 12045. [Google Scholar] [CrossRef]
- Eric, D. Salt weathering: A selective review. Geol. Soc. Lond. Spec. Publ. 2002, 205, 51–64. [Google Scholar] [CrossRef]
- Mol, L.; Viles, H.A. Geoelectric investigations into sandstone moisture regimes: Implications for rock weathering and the deterioration of San Rock Art in the Golden Gate Reserve, South Africa. Geomorphology 2010, 118, 280–287. [Google Scholar] [CrossRef]
- Wedekind, W.; López-Doncel, R.; Dohrmann, R.; Kocher, M.; Siegesmund, S. Weathering of volcanic tuff rocks caused by moisture expansion. Environ. Earth Sci. 2013, 69, 1203–1224. [Google Scholar] [CrossRef] [Green Version]
- Blanke, B.; Arhan, M.; Speich, S. Salinity changes along the upper limb of the Atlantic thermohaline circulation. Geophys. Res. Lett. 2006, 33, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Powell, T.M.; Cloern, J.E.; Huzzey, L.M. Spatial and temporal variability in South San Francisco Bay (USA). I. Horizontal distributions of salinity, suspended sediments, and phytoplankton biomass and productivity. Estuar. Coast. Shelf Sci. 1989, 28, 583–597. [Google Scholar] [CrossRef]
- Liu, C.; Liang, X.; Ponte, R.M.; Vinogradova, N.; Wang, O. Vertical redistribution of salt and layered changes in global ocean salinity. Nat. Commun. 2019, 10, 3445. [Google Scholar]
- Oguchi, C.T.; Yu, S. A review of theoretical salt weathering studies for stone heritage. Prog. Earth Planet Sci. 2021, 8, 32. [Google Scholar]
- Camuffo, D.; Delmonte, M.; Sabbioni, C. Origin and growth mechanisms of the sulfated crusts on urban limestone. Water Air Soil Pollut. 1983, 19, 351–359. [Google Scholar] [CrossRef]
- Knacke, O.; von Erdberg, R. The crystallisation pressure of sodium sulphate decahydrate. Ber. Bunsenges. Phys. Chem. 1975, 79, 653–657. [Google Scholar] [CrossRef]
- Akter, S.; Ahmed, K.R.; Marandi, A.; Schüth, C. Possible factors for increasing water salinity in an embanked coastal island in the Southwest Bengal Delta of Bangladesh. Sci. Total Environ. 2020, 713, 136668. [Google Scholar] [CrossRef]
- Kakeh, J.; Gorji, M.; Mohammadi, M.H.; Asadi, H.; Khormali, F.; Sohrabi, M. Effect of biocrusts on profile distribution of soil water content and salinity at different stages of evaporation. J. Arid Environ. 2021, 191, 104514. [Google Scholar] [CrossRef]
- Badaruddin, S.; Werner, A.D.; Morgan, L.K. Water table salinization due to seawater intrusion. Water Resour. Res. 2015, 51, 8397–8408. [Google Scholar] [CrossRef]
- Song, S.; Li, Z.; Li, C.; Yu, Z. The response of spring phytoplankton assemblage to diluted water and upwelling in the eutrophic Changjiang (Yangtze River) estuary. Acta Oceanol. Sin. 2017, 36, 101–110. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physiological/Biochemical Index | Salinity | Responses ↑ ↓ | Remark |
|---|---|---|---|
| Feed intake | −0.94 | ↓ ↓↓ | Increases metabolic depression |
| Gain in BW | −0.97 | ↓ ↓ ↓ | Modulate circulation, immunity, and blood functions such as O2− and CO2-carrying capacity. |
| Total length | −0.89 | ↓ ↓ ↓ | |
| RBC count | −0.96 | ↓ ↓ ↓ | |
| Hemoglobin | −0.99 | ↓ ↓ ↓ | |
| Thrombocytes | 0.38 | ↑ | |
| Lymphocytes | −0.80 | ↓↓↓ | |
| Monocytes | 0.38 | ↑ | |
| Eosinophils | −0.75 | ↓ ↓ | |
| Basophils | −0.72 | ↓ ↓ | |
| Neutrophils | 0.80 | ↑ ↑ ↑ | |
| LPx in the ARO | 0.93 | ↑ ↑ ↑ | Markers for tissue-level damage |
| LPx in the brain | 0.92 | ↑ ↑ ↑ | |
| LPx in the gills | 0.95 | ↑ ↑ ↑ | |
| LPx in the liver | 0.95 | ↑ ↑ ↑ | |
| LPx in the muscle | 0.95 | ↑↑↑ |
| Kinases in Signaling Pathways | Species in Which the Kinase Response is Observed |
|---|---|
| Mitogen-activated protein kinase | Tilapia (Oreochromis mossambicus) |
| Killifish (Fundulus heteroclitus) | |
| Turbot (Scophthalmus maximus) | |
| Myosin light-chain kinase | Japanese eel (Anguilla japonica) |
| Focal adhesion kinase | Killifish (Fundulus heteroclitus) |
| Osmotic stress transcription factor 1 | Mozambique tilapia (Oreochromis mossambicus) |
| Blackhead seabream (Acanthopagrus schlegelii) | |
| Japanese eel (Anguilla japonica) | |
| Mozambique tilapia (Oreochromis mossambicus) | |
| Nile tilapia (Oreochromis niloticus) | |
| Medaka, the Japanese rice fish (Oryzias latipes) | |
| Zebra fish (Danio rerio) |
| Response | Species | Osmoregulation Hypo/Hyper/Iso | Salinity | Respiration Rate | ROS Responses | Redox Enzyme Level |
|---|---|---|---|---|---|---|
| Extracellular responses | M. ligano | Hyper/iso | Low salinity | ↑ | ↑ (O2−) H2O2 other ROS ↓ | ↑ |
| C. aestuarii | Hyper/iso | High salinity | ↑ | ↑ | SOD ↑ | |
| N. granulata | Hyper/hypo | High salinity | ↓ | -- | ↑ | |
| S. serrata | Hyper/hypo | Salinity | SOD ↑ CAT ↑ | |||
| B. koreanus | ND | High salinity | -- | ↑ | GST activity ↑ | |
| A. naccarii | ND | High salinity | -- | ↑ | SOD ↑ CAT ↑ GPx ↑ | |
| Intracellular responses | A. microstoma | ND | Low salinity | -- | ↑ | CAT ↑ SOD insignificant |
| A. tamarensis | ND | High salinity | -- | ↑ | SOD ↑ CAT insignificant | |
| C. gigas | Osmoconformer | Wide range | No effect | NA | SOD not affected CAT ↑ at high salinity | |
| D. labrax | ND | Low salinity | -- | ↑ | CAT↑ | |
| L. vannamei | ND | Both high and low salinity | -- | ↑ | SOD ↑ CAT ↑ | |
| A. schlegeli | ND | Low salinity | -- | -- | SOD ↑ CAT ↑ | |
| S. broughtonii | ND | Low salinity | -- | -- | SOD ↑ CAT ↑ | |
| S. plana | Osmoconformer | Low salinity | -- | TBARS ↑ | GPx ↑ GST↑ CAT ↑ | |
| D. neopolitana | Osmoconformer | Low salinity | -- | -- | CAT ↑ | |
| V. decussate | Osmoconformer | Wide range of salinity | -- | LPO high | GSH ↑ SOD high at low salinity for all | |
| V. corrugata | ND | Change in salinity | LPO high | GSH ↑ CAT high at low salinity for all | ||
| V. philippinarum | ND | Change in salinity | LPO high | GSH high | ||
| H. discus discus | Osmoconformer | Lower salinity | -- | -- | ↑ | |
| C. angulata | Osmoconformer | Lower salinity | -- | -- | No change | |
| R. philippinarum | Osmoconformer | Lower salinity | -- | -- | SOD ↑ | |
| C. gigas | Osmoconformer | Lower salinity | -- | -- | No change | |
| R. decussates | Osmoconformer | Higher salinity | -- | High LPO at extreme salinity | SOD ↑ CAT ↓ GST ↓ | |
| R. philippinarum | ND | Changing salinity | -- | SOD ↑ CAT ↑ GST no change |
| Anthropogenic or Climatic Factor | Area of Investigation | Impact on Salinity | Reasons for Salinity Alteration | Reference |
|---|---|---|---|---|
| Temperature and rainfall | Khorezm, Uzbekistan | Increased salinity | Decreased groundwater resources | [184] |
| Glacial melting | Indian Sundarbans | Decreased salinity in the Hooghly estuary | Freshwater input from Himalayan glaciers | [185] |
| Regional warming | Central Gangetic Delta, India | Increasing salinity | High rate of evaporation | [186] |
| Global warming | NA | Enhanced salinization | Nutrient loading, evaporative up-concentration of nutrients in reduced water volumes | [187] |
| Wind direction | Coast of Tarragona (Spain), Uruguayan coast | Increases atmospheric salinity | Entrainment of marine aerosols from the sea | [187,188,189] |
| Precipitation | Wuliangsuhai Lake, China | High salinity | Concentrations of salts increase with low precipitation | [190] |
| Moisture | South Africa, Mexico, Germany, and Hungary | Increases salinity | Rock weathering releases mineral salts | [191,192,193] |
| Thermohaline circulation | Across the Atlantic to 47° N | Salinity increases of 0.38 psu | Cooling results in a density increase | [194] |
| Horizontal distribution | Equator (between 20–30 degrees north) | 34 ppt salinity | High humidity and less precipitation | [188,195] |
| Vertical distribution | 300–500 m depth | 33–37 ppt salinity | NA | [196] |
| Gaseous pollutants | Madrid | Increased salinity | Crystallization and hydration pressure; salt weathering | [197] |
| Sulfate/sulfuric acid | Madrid | Increased salinity | Crystallization and hydration pressure; carbonate and silicate rock | [197,198,199] |
| Nitrate/nitric acid | NA | Increased salinity | Decreased nitrate reduction, organic waste, and deicing salts | [197] |
| Ca2+, Mg2+, SO42−, HCO3−, Na+, and Cl− | Southwest Bengal Delta of Bangladesh | High salinity in both ground and surface water (7.5 to 8 ppt) | Saltwater shrimp cultivation, embankments, and excessiveirrigation | [200,201] |
| Biocrusts | Alagol Lake, Northern Iran | Increased salinity up to 50% | Lack of biocrust induces high rates of evaporation | [202] |
| Manmade/natural | Yangtze River, East China Sea | Groundwater salinization | Seawater intrusion | [202,203] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bal, A.; Panda, F.; Pati, S.G.; Anwar, T.N.; Das, K.; Paital, B. Influence of Anthropogenic Activities on Redox Regulation and Oxidative Stress Responses in Different Phyla of Animals in Coastal Water via Changing in Salinity. Water 2022, 14, 4026. https://doi.org/10.3390/w14244026
Bal A, Panda F, Pati SG, Anwar TN, Das K, Paital B. Influence of Anthropogenic Activities on Redox Regulation and Oxidative Stress Responses in Different Phyla of Animals in Coastal Water via Changing in Salinity. Water. 2022; 14(24):4026. https://doi.org/10.3390/w14244026
Chicago/Turabian StyleBal, Abhipsa, Falguni Panda, Samar Gourav Pati, Taslima Nasim Anwar, Kajari Das, and Biswaranjan Paital. 2022. "Influence of Anthropogenic Activities on Redox Regulation and Oxidative Stress Responses in Different Phyla of Animals in Coastal Water via Changing in Salinity" Water 14, no. 24: 4026. https://doi.org/10.3390/w14244026