
Variability of the Carbon Isotope Composition of Peat-Forming Plants during the Biochemical Transformation


Institute of Monitoring of Climatic and Ecological Systems SB RAS, 634055 Tomsk, Russia
*
Author to whom correspondence should be addressed.
Water 2022, 14(24), 4035; https://doi.org/10.3390/w14244035
Submission received: 10 November 2022
/
Revised: 7 December 2022
/
Accepted: 8 December 2022
/
Published: 10 December 2022
(This article belongs to the Special Issue Climate Change and Anthropogenic Impacts on Wetland Ecosystems in Siberia: Past, Present, and Future)
Abstract
:In this study, we describe the variation in δ13C value in the litter of two species of peat-forming plants: Sphagnum fuscum and Eriophorum vaginatum, during 3 years of field decomposition in oligotrophic bog ecosystems drained for the purpose of forest melioration and fire affected and at the stage of post-pyrogenic restoration. Litterbags were periodically retrieved in the autumn and the δ13C value in the residual litter was related to mass loss, litter chemistry, and hydrothermal conditions. Sph. fuscum decomposes much more slowly than E. vaginatum. Low rate of transformation for Sph. fuscum is observed in drained and post-pyrogenic sites, while for E. vaginatum minimal rate of transformation is observed in the native site. During the decomposition of Sphagnum residues, 13C enrichment occurs, and during the decomposition of E. vaginatum, we observed 12C enrichment. The changes in the isotope composition of carbon for investigation sites are insignificant for Sphagnum fuscum, but it was observed for E. vaginatum, the largest of 13C depletion is observed in the drained site (−28.3‰) and minimal in the postpyrogenic site (−27.4‰).
1. Introduction
Occupying a small area (about 3–5%) of the land surface, wetland ecosystems play a significant role in the biosphere, including the global carbon cycle, being sources and sinks of greenhouse gases [1,2,3]. On the territory of Western Siberia, the area of bog ecosystems is almost 50%; peat deposits contain about 36% of the total pool of soil carbon in Russia [1,4,5]. The process of carbon accumulation in peat prevails over the process of its emission, due to the slow process of decomposition of plant residues. This causes a constant increase in peat deposits and the development of bog ecosystems. There are relatively few works devoted to the study of the dynamics of decomposition of peat-forming plants in bog ecosystems [6,7,8,9,10,11,12,13,14,15,16]. The rate of decomposition of plant residues depends on the chemical composition of plants and the hydrothermal conditions under which these processes occur [6,17,18,19]. While humidity and temperature play a leading role in the transformation of plant residues [6,7,17,20], changing weather conditions cause an irregular, intermittent peat-forming process, which is typical for raised bogs that receive their main water supply from precipitation. Climatic changes or anthropogenic impact (drainage) influence the hydrothermal regime of bogs. These variations lead to changes in biogeochemical cycles since warming and a decrease in the level of bog waters stimulate the process of decomposition of organic matter and increase the intensity of CO2 release into the atmosphere. In addition, it leads to changes in the composition and structure of the vegetation cover [1,21,22,23,24].
Melioration, especially for agricultural purposes, leads to the stimulation of microbial decomposition of organic matter and to an increase in the heterotrophic carbon flux into the atmosphere [25,26]. Forest melioration is one of the most sparing options for draining peat soils. Oligotrophic bogs of Western Siberia are in very favorable climatic conditions. Due to the predominance of precipitation over evaporation, there is a rapid recovery of bog ecosystems after forest melioration [27]. Moreover, one of the most significant factors influencing the biogeochemical cycles of bog ecosystems is fires [28]. The restoration of ecosystems and their biogeochemical cycles after fires in natural conditions is of considerable scientific and practical interest. Post-pyrogenic succession is characterized by a gradual change in ecosystems, accompanied by a change in species richness and biomass, and, accordingly, the rate of transformation of organic matter and its chemical composition [29,30].
Many biogeochemical processes are accompanied by a change in the stable carbon isotope ratio, due to which different components of ecosystems and different ecosystems differ in their isotope composition [31]. A change in the stable carbon isotope ratio indicates ongoing biochemical transformations and, therefore, makes it possible to obtain information about the functioning of ecosystems in natural or disturbed conditions, as well as to use it to identify the direction and intensity of processes associated with isotope fractionation [32,33,34]. A large number of studies are devoted to terrestrial and aquatic ecosystems [35,36,37,38,39,40,41]. Changes in δ13C value during the decomposition of plant residues depend on many factors, including changes in the ratio of chemical components of the plant tissues themselves [42], as a result of which δ13C value may change non-linearly during transformation. There are many studies devoted to the study of the isotope composition of natural ecosystems. However, data on changes in the isotope composition in bog ecosystems are not so numerous; moreover, these data are often contradictory depending on the study area, types of ecosystems, and plants themselves [35,36,38,41,43,44,45]. Studies conducted in bog ecosystems are mainly related to the study of the isotope composition of peat organic matter [46,47,48,49,50,51]. Very rarely have the plants themselves or the early stages of transformation of plant remains been studied [52,53,54,55,56,57]. On the territory of the southern taiga subzone of Western Siberia, the issue of changes in the isotope composition during the transformation of plant remains has practically not been studied at all [58], especially in bog ecosystems subject to anthropogenic or pyrogenic influence.
The purpose of the study was to assess the change in the carbon isotope composition during the transformation of plant residues Sph. fuscum and Eriophorum vaginatum in oligotrophic bog ecosystems drained for forest melioration and passed through by fire and at the stage of postpyrogenic restoration.
2. Materials and Methods
The measurement site is located in the south of Western Siberia (Russia). The study was carried out on two oligotrophic bogs “Vasuyganskoe” (field station “Vasyuganye” (IMCES SB RAS)) and “Iksinskoe”, belonging to the northeastern spurs of the Great Vasyugan bog and located in the Bakcharsky district of the Tomsk region. Since the 1970s, ameliorative canal network has been operating in these bogs, as a result of which cardinal changes in the natural environment occurred in the bogs, in particular, the drainage of significant bog areas, which caused massive forest and peat fires [59].
Two observation sites were chosen for the study at Vasuyganskoe bog—the native pine-shrub-sphagnum phytocenosis 56°52′31.7″ N 82°48′27.3″ E (VASnat) and the pine-shrub-sphagnum phytocenosis located near the drainage canal 56°53′33.3″ N 82°51′08.0″ E (VASdry). The Iksinskoe bog is located between the Iksa and Shegarka rivers (Figure 1). In 1998, a large part of the territory burned out in the drained area of the Iksinskoe bog. The vegetation cover from the near-surface peat layer in the bogs was completely destroyed [59]. The following two sites with different degrees of pyrogenic succession were selected for the study: a pine-shrub-sphagnum phytocenosis with a well-defined undergrowth 56°51′42.1″ N 83°17′53.0″ E (Iksa2) and a pine-birch-cotton grass-sphagnum phytocenosis with a less pronounced degree of pyrogenic succession 56°52′03.4″ N 83°11′52.1″ E (Iksa1).
The data on weather conditions were obtained from the nearest meteorological station, Bakchar [rp5.ru], located 40 km from the study site. An atmospheric soil measuring complex was used to monitor the soil temperature and the level of bog waters [60]. The peat deposit temperature was measured 5; 10; 20 cm deep into the peat. Soil temperature data were obtained for the period from September 2018 to September 2021.
2.1. Determination of the Decomposition Rate of Peat-Forming Plants
The following two dominant species of the modern plant cover of oligotrophic bogs have been chosen to study the decomposition of peat-forming plants: Sph. fuscum and E. vaginatum. The decomposition rate of plants was determined by the method of partially isolated samples, which is widely used to study the transformation processes of plant material and peat [Golovatskaya, Nikonova, 2017].
In September 2018, we sampled the moss litter of Sph. fuscum (their top 10 cm-long parts) and remains of E. vaginatum on each of the studied bogs. The plant samples were air-dried at a laboratory and placed in nylon bags 15 × 15 cm size. Each plant sample weighed 10 g. The prepared bags with plants were placed into peat deposits at a depth of 10 cm from the moss surface (sphagnum fiber) in September 2018. In total, 96 bags were prepared (48 bags for each type of plant residue)
Mass loss (ML, % of the initial mass of plant sample) was calculated by the following equation:
where M0 is dry mass of the initial sample and Mt is dry mass of sample remained in bag over 12-, 24-, and 36-month-long periods after the experiment start.
To assess the change in the content of total C in the initial samples of the studied plants, we used the method of simultaneous determination of the total C content in peat soils elaborated by Anstett’s method modified by Ponomareva and Nikolaeva [61,62]. The analyses were performed in three replications.
The carbon isotope composition was determined by isotope ratio mass spectrometry [63] using a DELTA V Advantage isotope mass spectrometer combined with a Flash 2000 elemental analyzer (Thermo Fisher Scientific, Bremen, Germany) equipped with a redox reactor. All samples were milled, after which a sample was placed in tin capsules (tin of a high degree of purity). The optimal weight of a sample for carbon isotope analysis was 450–500 µg. Encapsulated samples were placed in an autosampler of elemental analyzer. The capsule fell into an oxidizing reactor heated to 1020 °C and filled with Cr2O3 and Co3O4 granules, burned in a carrier gas flow (helium, 250 mL/min) with a simultaneous supply of pure oxygen (180 mL/min). The resulting oxidation products (CO2) entered the reduction reactor. A magnesium perchlorate trap was used to remove water. Carbon dioxide entered the DELTA V Advantage mass spectrometer via a capillary through a Conflo IV gas distribution system.
The isotope composition was determined by the following formula:
where nX are 13C isotopes (‰); Rsample is the ratio of the heavy to light isotope in the test sample; Rstandard is the ratio of heavy to light isotope in the standard.
Laboratory reference gas (CO2) was calibrated against IAEA Reference Material IAEA-CH-3 Cellulose with a known stable isotope ratio as follows: δ13C VPDB = –24.72 ± 0.04‰. The absolute measurement error for three repetitions of the analyzed samples is no more than 0.2‰.
2.2. Statistical Analysis
The influence of plant species and site ecosystem type on the rate of decomposition and dynamics of carbon isotope composition was tested using two-way analysis of variance. Student’s t-test was used to assess significant differences in the chemical composition of the litter within and between plant species. All statistical analyzes were performed using Statistics for Windows v. 6.0.
3. Results
3.1. Weather and Hydrothermal Conditions
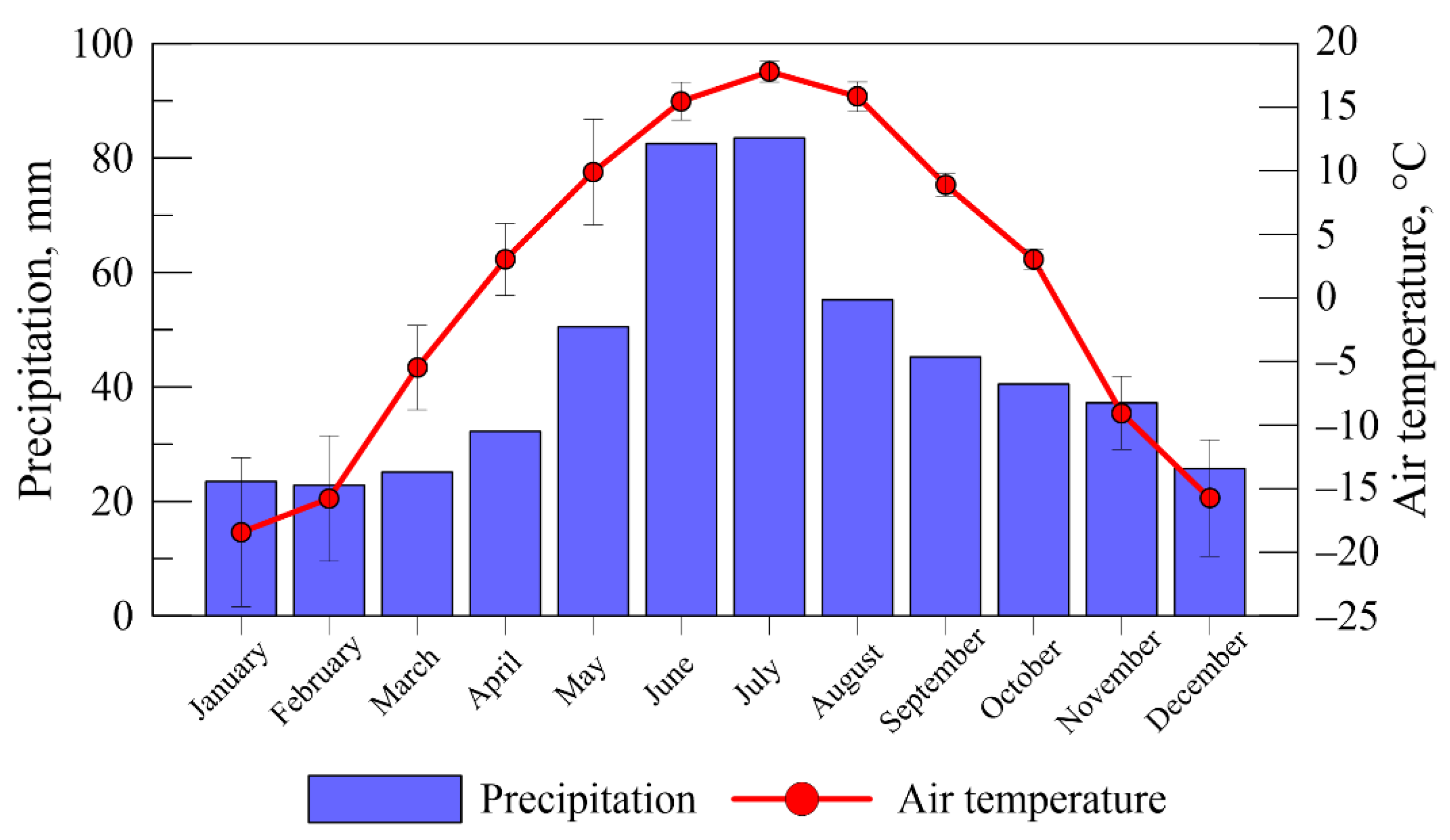
During the study period (2018–2021), the average annual temperature was 0.79 ± 1.47 °C, and the average annual cumulative precipitation was 524 ± 92 mm. The average temperature of the growing season from the beginning of May to the end of September was about 13.9 °C, and the amount of precipitation during the growing season (May–September) was 277 mm (Figure 2).
On average, over the period of the experiment site, VASdry was characterized by the lowest level of bog waters (WTL) −55 cm and the lowest average temperature at a depth of 15 cm during the growing season (+9.5 °C). Iksa1 was the most watered area (WTL = −17 cm) and was characterized by the warmest conditions of the peat deposit (+13.6 °C).
Iksa2 was also quite watered (WTL −27), but the temperature of the peat deposit is much lower (+10.7 °C). VASnat conditions occupied an intermediate position between drained (VASdry) and pyrogenic (Iksa1) peatlands—low WTL (–40 cm), relatively warm conditions of a peat deposit (+12.2 °C).
3.2. Decomposition Rate
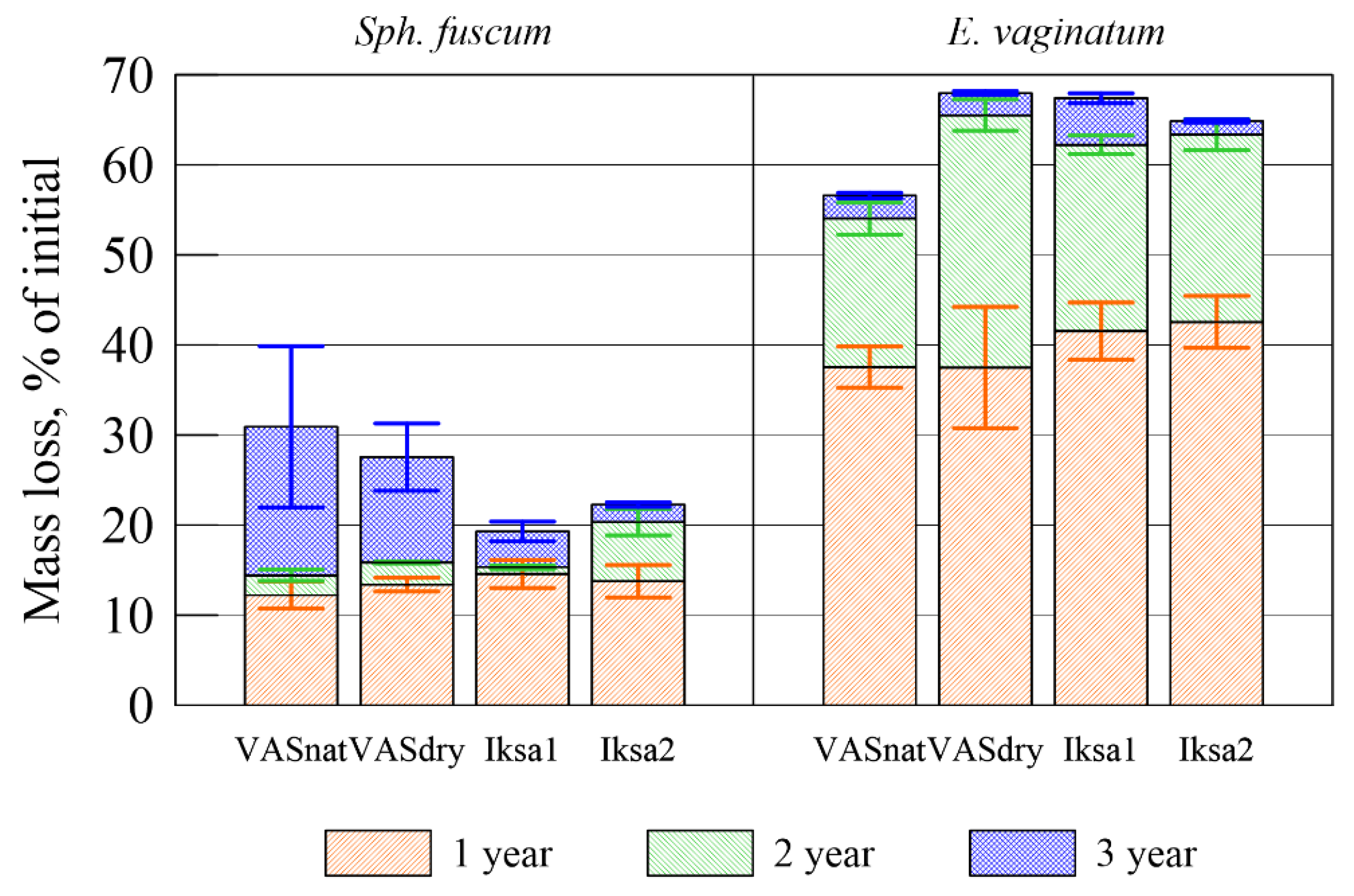
After 3 years of decomposition, the average mass loss for sphagnum litter was 25% (from 19.3 to 30.9% depending on the sites), and E. vaginatum litter was on average 64% (56.6–67.9%) (Table 1).
The most intensive decomposition occurred within 1 year of the experiment, with the exception of fuscum in the natural area, in which the maximum losses were obtained within 3 years. On average, the loss of mass of organic matter during 1 year was 56% and 62% of the total loss for the entire period of the experiment (3 years) for Sph. fuscum and E. vaginatum, respectively (Figure 3). The process of transformation of plant residues of Sph. fuscum occurred more intensively in VASnat and VASdry sites, while the decomposition of moss on Iksa1 and Iksa2 postpyrogenic peatlands was slower. On the VASdry and VASnat sites, the mass loss during the 3 years of the experiment was comparable to (VASdry) or even exceeded (VASnat) the weight loss during the 1-year period. We assume that the increase in the rate of decomposition of Sphagnum fuscum during the third year of the experiment is associated with the activation of microbiological activity. The decomposition of E. vaginatum, on the contrary, proceeded more slowly in natural conditions.
3.3. Variability of the Isotope Composition
The carbon content in the litter decreased after one year of the experiment, for Sph. fuscum by 3–5% and for E. vaginatum by 1.5–4%. By the end of the second year, the content of C had significantly increased in Sph. fuscum in the VASnat and Iksa1 (on average, 5%). In E. vaginatum, the carbon content also increased, but not so significantly. After 3 years, the carbon content returned to its initial values in all samples, with the exception of Sph. fuscum in the VASnat (Figure 4).
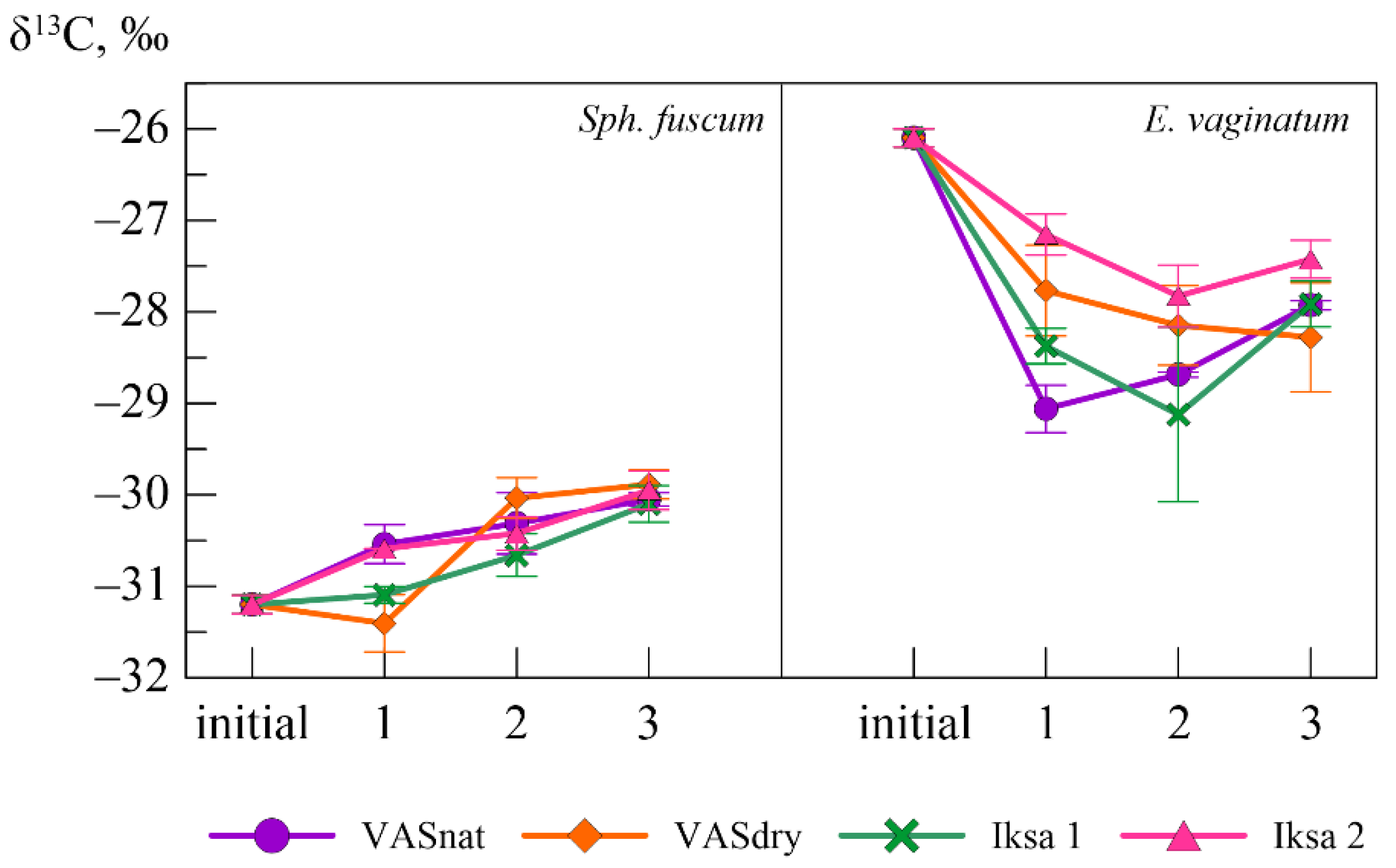
According to the analysis of the isotope composition of plant residuals (Figure 5), during the transformation of Sph. fuscum at all sites, the carbon isotope composition became heavier by an average of 1‰. For Sph. fuscum enrichment in the heavy carbon isotope was observed throughout the period at all sites, except for the VASdry one, where during the 1st year of the experiment a low δ13C value was observed, followed by an increase. For E. vaginatum, the reverse process was observed as follows: the carbon isotope composition changed by an average of −1.8‰. A significant change in the isotope composition of E. vaginatum occurred during 1 year of the experiment, when the average δ13C value was −28‰. Low δ13C values occurred in the E. vaginatum at all sites during the first and second years of incubation. During the third year, there is a slight enrichment by 13C; however, the δ13C value remains significantly lower than in the initial samples. An exception is the E. vaginatum placed at the dried site, where, as a result of the leaching regime, continues despite rather high mass losses.
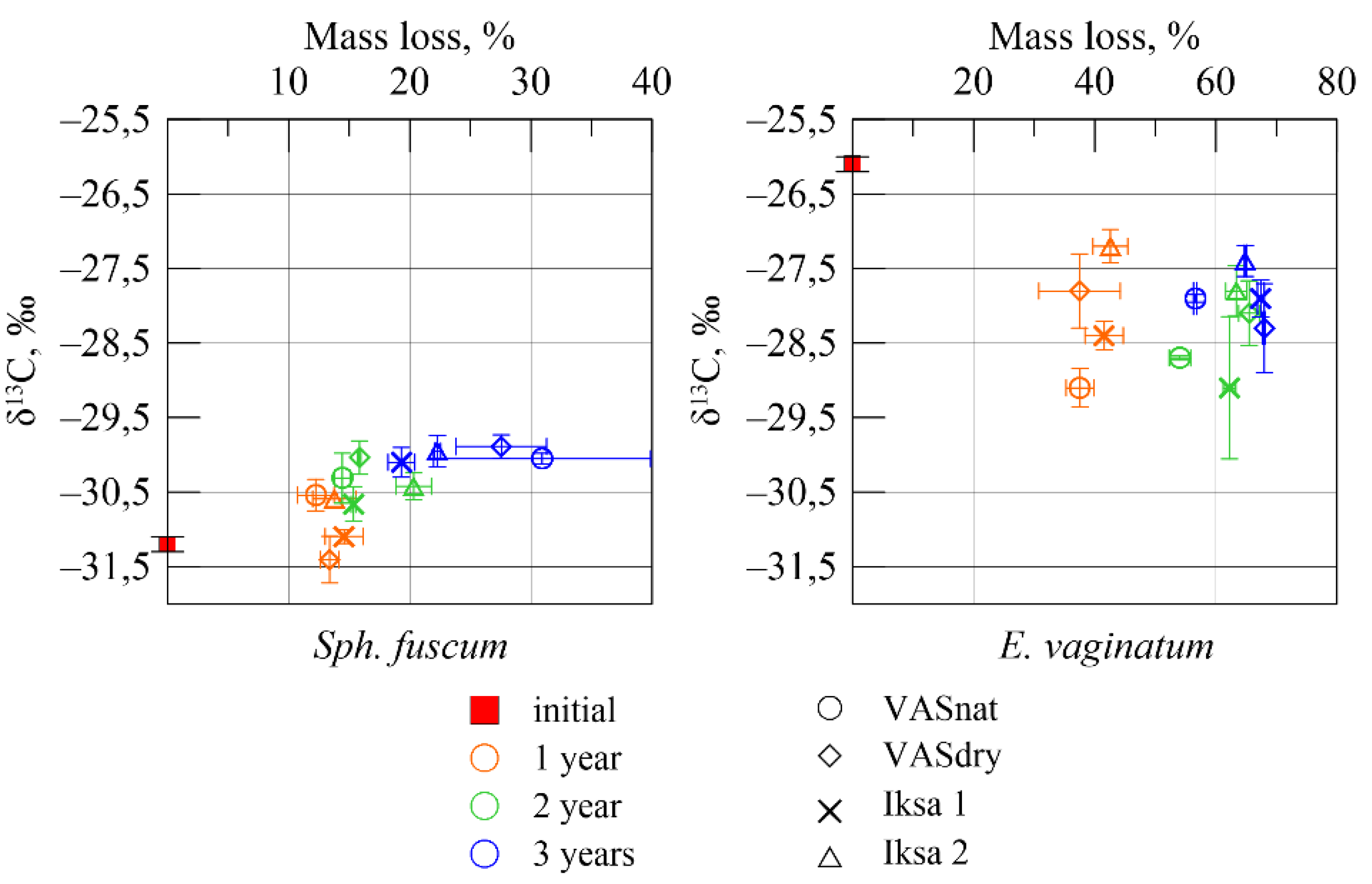
The results of the correlation analysis showed a close relationship between weight loss and changes in the isotope’s composition (Figure 6). For Sph. fuscum, the relationship was positive (correlation coefficient 0.81), and for E. vaginatum, it was negative (−0.79).
According to a two-way analysis of variance, the δ13C value of the residual plants significantly varied depending on the plant species and not depending on the location of the samples (Table 2).
The influence of the location on the δ13C value most clearly appears during the 1st year of the experiment for both Sph. fuscum and E. vaginatum. At the same time, samples of Sph. fuscum incubated in the VASnat and in the Iksa2 area have similar δ13C values throughout the period of investigation, significantly differing from the VASdry and Iksa1. For E. vaginatum, significant differences in the δ13C value were found in the location for all sites, and, unlike sphagnum, the maximum difference was observed between samples on VASnat and Iksa2 during the first year. By the end of the third year, the differences in the location of the samples are not significant.
4. Discussion
The obtained differences in the rate of mass loss of Sph. fuscum and E. vaginatum are significant and are consistent with previously obtained data [12,19]. Sphagnum mosses contain the least amount of bitumen and many easily hydrolyzable and water-soluble compounds of the carbohydrate complex; therefore, sphagnum mosses should be the least resistant to decomposition. However, they also contain a specific phenolic compound, sphagnol, which prevents rapid bacterial decomposition [64]. The low content of lignin can also contribute to slow decomposition. Nutrient-enriched plant residues with a high nitrogen content, a minimal C/N ratio, and a low lignin content usually quickly decompose. For plant residues with a low nitrogen content, a wide C/N ratio, and a high lignin content, the decomposition rate is much lower. In this case, not only the lignin content but also the lignin/nitrogen ratio is an important factor, since it has been shown that the higher the lignin/nitrogen ratio, the slower the decomposition occurs [65,66]. We assume that due to the low nitrogen content (0.4%) with a rather high lignin content (12%), a high lignin/nitrogen ratio (32) was obtained, which apparently contributes to a lower decomposition rate of Sphagnum fuscum. Sphagnum mosses and in some species of herbaceous plants (Carex, Eriophorum, Scheuchzeria) hemicellulose predominates among carbohydrates, which is also more resistant to decomposition [64]. Thus, sphagnum mosses have chemical and mechanochemical immunity, which allows them to remain in deposits. The rapid decomposition of plant residues at the initial stages is associated with the processes of leaching of easily soluble substances from fresh plant litter, as well as the intensive activity of microorganisms [67,68,69,70,71,72]. Microorganisms destroy the easiest available components of the carbohydrate and polypeptide complexes, the content of which decreases with time, and, accordingly, the loss of organic matter decreases [8,73]. Sphagnum organic matter can contain up to 20% labile water-soluble components such as carbohydrates [74]. The loss of these substances explains the large mass loss in the earliest stages of peat formation [75].
The influence of the location on the change of δ13C value can be assessed through such parameters as peat temperature and the water table level. The results of the correlation analysis showed a weak relationship between the temperature of the peat deposit and the δ13C value (r = −0.29, p < 0.05 and −0.41 p < 0.05 for Sph. fuscum and E. vaginatum, respectively). The dependence dynamics of the δ13C value on temperature were noted in the article by Bragazza and Iacumin [15]. They studied seasonal dynamics and observed 13C enrichment during the summer period and 13C depletion in the winter. In addition, studies of litter decomposition under laboratory conditions show changes in the δ13C value depending on temperature [76].
The analysis of the influence of the water table level in the studied areas did not reveal a relationship between the δ13C value and the water table level. Obviously, this is because even at sufficiently low water levels, the moisture content of the upper horizons of the peat deposit remains quite high (80–85%). Therefore, the decrease in the water table level is less critical for the dynamics of the δ13C value in comparison with the change in temperature. At the same time, a decrease in the water table level leads to an increase in the rate of transformation of plant residues under drier conditions, decomposition proceeds more intensively, which is consistent with other studies [10,18,19,53]
The change in δ13C value was different for the two species, the δ13C value of Sph. fuscum litter was enriched, while the δ13C value of E. vaginatum litter decreased. We assume that this also depends on the chemical composition of the plants. Eriophorum has a higher content of cellulose, lignin and lignin-like substances, alcohol-soluble compounds (including aromatic and aliphatic carbohydrates, terpenes, carbolic acids, resins, and fatty acids, essential oils, fats, and phytosterols) [58]. Since soluble elements are more enriched in the heavy carbon isotope than lipids and lignins, the removal of the soluble fraction as a result of leaching will lead to low δ13C values [36,77,78]. Our data are consistent with previous studies in which low δ13C values were noted at the early stages of the decomposition of plant residues [38,41]. Over time, the rate of transformation significantly decreases, probably due to the transformation process proceeds largely due to the activity of microorganisms. Probably, as a result of the activation of microorganisms, the accumulation of a heavy isotope can occur due to the contribution of microbial organic compounds [15]. Earlier studies also noted that enrichment by the heavy isotope can be caused by the inclusion of carbon from soil organic matter by decomposers Wedin et al., [36], in addition, it was found that significant changes occur in microfungal communities [79].
5. Conclusions
Quantitative estimates of the rate of decomposition of two species of peat-forming plants (Sph. fuscum and E. vaginatum) were obtained during 36 months of a field experiment. The highest resistance to decomposition, whose mass loss over the three years of the experiment was 19–31%, characterized the samples of Sph. fuscum. E. vaginatum showed the least resistance to decomposition with the maximum mass loss—56–68% after three years. The most intensive decomposition of plant residuals was observed during the first year of the experiment; further, the rate of decomposition of organic matter decreased.
The rate of decomposition is determined by environmental conditions, but primarily the rate depends on the chemical composition of plants. The next most important factor is the temperature, the higher the temperature. The influence of the water level on the transformation process is unreliable. Our study revealed significant differences in the dynamics of the δ13C value during the decomposition of plant residues. After 3 years, Sph. fuscum showed a high δ13C value, while E. vaginatum showed a low δ13C value relative δ13C values of the initial samples. Due to the peculiarities of the morphological and chemical structure of Sphagnum fuscum, the δ13C value in plant residues is associated with microbiological activity, and the enrichment by the 13C isotope most likely occurs due to microbial biomass. E. vaginatum was characterized by rapid mass loss. This leads to a sharp decrease in the δ13C value and subsequent slight enrichment in the 13C isotope with a decrease in the transformation rate due to a decline in the role of leaching and an increase in the role of microbial decomposition, which leads to an increase in the content of 13C isotope due to microbial biomass. The changes in the isotope composition of carbon at the investigation sites are insignificant for Sphagnum fuscum, but they were observed for E. vaginatum. The largest decrease in the δ13C value is observed in the drained site VASdry (the δ13C value was −28.3‰ for the third year of the experiment relative −26.1‰ for the initial sample) and minimal in the post-pyrogenic site Iksa2 (the difference between the δ13C value in the third year of the experiment and the δ13C value of the initial sample was −1.3‰).
Author Contributions
E.G., L.N.—organized field work and collected plant samples; L.N.—mass loss analysis of plant residues, G.S., D.K.—determination of carbon content and isotopic composition of samples; E.G.—wrote a draft version of the manuscript, which was commented on by all co-authors. All authors have read and agreed to the published version of the manuscript.
Funding
The study was supported by the Russian Science Foundation grant No. 22-27-00363.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank E.A. Dyukarev (IMKES SB RAS, Russia) for providing data on the temperature of the peat deposit and the levels of bog waters. The authors are thankful to the Shared Research Facilities of the Tomsk Scientific Center of the Siberian Branch of the RAS for providing the DELTA V Advantage isotope mass spectrometer.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vompersky, S.E. Role in the carbon cycle bogs. In Biogeocenotic Features of Bogs and Their Rational Use; Nauka: Moscow, Russia, 1994; pp. 5–37. [Google Scholar]
- Gorham, E. Northern peatlands: Role in the carbon cycle and probable responses to climatic warming. Ecol. Appl. 1991, 1, 182–195. [Google Scholar] [CrossRef] [PubMed]
- Succow, M.; Joosten, H. Landschaftsökologische Moorkunde; Schweizerbart Science Publishers: Stuttgart, Germany, 2001; p. 622. [Google Scholar]
- Efremov, S.P.; Efremova, T.T.; Melentieva, N.V. Carbon storages in peatland ecosystems. In Carbon in Forest and Peatland Ecosystems of Russia; Alekseev, V.O., Berdsy, R.A., Eds.; Institute of forest SB RAS: Krasnoyarsk, Russia, 1994; pp. 128–139. [Google Scholar]
- Titlyanova, A.A.; Bulavko, G.I.; Kudryashova, S.Y.; Naumov, A.V.; Smirnov, V.V.; Tanasienko, A.A. The Reserves and Losses of Organic Carbon in the Soils of Siberia. Eur. Soil Sci. 1998, 31, 45–53. [Google Scholar]
- Kozlovskaya, L.S.; Medvedeva, V.M.; Piavchenko, N.I. Dynamics of Organic Matter in the Process of Peat Formation; Nauka LO Publ.: Leningrad, Russia, 1978; p. 171. [Google Scholar]
- Boch, M.S.; Masing, V.V. Wetland Ecosystems of the Soviet Union; Nauka: Leningrad, Russia, 1979; p. 188. [Google Scholar]
- Bambalov, N.N.; Horuzhik, A.V.; Lukoshko, E.S.; Strigutsky, V.P. Transformation of dead plants in marsh biogeocenoses. In Experiment and Mathematical Modeling in the Study of Biogeocenoses of Forests and Bog; Nauka Publ.: Moscow, Russia, 1990; pp. 53–63. [Google Scholar]
- Mironycheva-Tokareva, N.P.; Kosykh, N.P.; Vishnyakova, E.K. Carbon balance components of wetlands in the middle taiga and the forest-tundra. West Siberian Peatlands and Carbon Cycle: Past and present. In Proceedings of the Second International Field Symposium, Khanty-Mansiysk, Russia, 24 August–2 September 2007; pp. 117–118. [Google Scholar]
- Parshina, E.K. Destruction of Plant Matter in Bog Ecosystems of the Taiga and Forest-Tundra Zones of Western Siberia. Ph.D. Dissertation, Tomsk State University, Tomsk, Russia, 2009. [Google Scholar]
- Vishnyakova, E.K.; Mironycheva-Tokareva, N.P.; Kosykh, N.P. Decomposition Dynamics of Plants of Bogs (Data: Vasyugan Bog). Tomsk. State Pedagog. Univ. Bull. 2012, 7, 88–93. [Google Scholar]
- Golovatskaya, E.A.; Nikonova, L.G. Decomposition of plant residues in peat soils of oligotrophic peatlands. Vestn. Tomsk. Gos. Univ. Biol. 2013, 3, 137–151. [Google Scholar]
- Verhoeven, J.T.A.; Arts, H.H.M. Decomposition of Carex litter in fens with different water chemistry. Aquat. Bot. 1992, 43, 365–377. [Google Scholar] [CrossRef]
- Bragazza, L.; Siffi, C.; Iacumin, P.; Gerdol, R. Mass loss and nutrient release during litter decay in peatland: The role of microbial adaptability to litter chemistry. Soil Biol. Biochem. 2007, 39, 257–267. [Google Scholar] [CrossRef]
- Bragazza, L.; Buttler, A.; Siegenthaler, A.; Mitchel, E.A. Plant litter decomposition and nutrient release in peatlands. Carbon Cycl. North. Peatl. 2009, 184, 99–100. [Google Scholar]
- Leroy, F.; Gogo, S.; Buttler, A.; Bragazza, L.; Laggoun-Défarge, F. Litter decomposition in peatlands is promoted by mixed plants. J. Soils Sediments 2018, 18, 739–749. [Google Scholar] [CrossRef] [Green Version]
- Denisenkov, V.P. Basics of Swamp Science; St. Petersburg University Publ.: St. Petersburg, Russia, 2000; p. 224. [Google Scholar]
- Peltoniemi, K.; Strakova, P.; Fritze, H.; Iraizoz, P.A.; Pennanen, T.; Laiho, R. How water-level drawdown modified litter-decomposing fungal and actinobacterial communities in boreal peatlands. Soil Biol. Biochem. 2012, 51, 20–34. [Google Scholar] [CrossRef]
- Nikonova, L.G.; Golovatskaya, E.A.; Kur’ina, I.V.; Kurganova, I.N. Decomposition rate of peat-forming plants in oligotrophic bogs of the southern taiga subzone of Western Siberia: Assessment of the effect of water table level and peat deposit temperature. Eurasian Soil Sci. 2019, 52, 1101–1111. [Google Scholar] [CrossRef]
- Nikonova, L.G.; Kurganova, I.N.; Lopes de Gerenyu, V.O.; Zhmurin, V.A.; Golovatskaya, E.A. Impact of abiotic factors on the decomposition of litter of peat-forming plants in the incubation experiment. Tomsk. State Univ. J. Biol. 2019, 46, 148–170. [Google Scholar] [CrossRef] [Green Version]
- Karelin, D.V.; Zamolodchikov, D.G. Carbon Exchange in Cryogenic Ecosystems; Nauka: Moscow, Russia, 2008; p. 344. [Google Scholar]
- Kudeyarov, V.N.; Zavarzin, G.A.; Blagodatskii, S.A.; Borisov, A.V.; Voronin, P.Y.; Demkin, V.A.; Demkina, T.S.; Yevdokimov, I.V.; Zamolodchikov, D.G.; Karelin ; et al. Pools and Fluxes of Carbon in Terrestrial Ecosystems of Russia; Nauka: Moscow, Russia, 2007; p. 315. [Google Scholar]
- Leadley, P.; Pereira, H.M.; Alkemade, R.; Fernandez-Manjarrés, J.F.; Proença, V.; Scharlemann, J.P.W.; Walpole, M.J. Biodiversity Scenarios: Projections of 21st Century Change in Biodiversity and Associated Ecosystem Services; Secretariat of the Convention on Biological Diversity: Montreal, Canada, 2010; p. 132. [Google Scholar]
- Gifford, R.M.; Howden, M. Vegetation thickening in an ecological perspective: Significance to national greenhouse gas inventories. Environ. Sci. Policy 2001, 4, 59–72. [Google Scholar] [CrossRef]
- Bambalov, N.N. Balance of the Organic Matter of Peat Soils and Methods of Its Study; Nauka Publ.: Minsk, Belarus, 1984; p. 175. [Google Scholar]
- Efimov, V.N. Peat Soils and Their Fertility; Agropromizdat Publ.: Leningrad, Russia, 1986; p. 264. [Google Scholar]
- Golovatskaya, E.A. Carbon fluxes in bog ecosystems of Western Siberia. Doctoral Dissertation, Tomsk State University, Tomsk, Russia, 2013. [Google Scholar]
- Parish, F.; Sirin, A.A.; Charman, D.; Joosten, H.; Minaeva, T.Y.; Silvius, M. Assessment on Peatlands, Biodiversity and Climate Change; Wetlands International: Kuala Lumpur, Wageningen, 2008; p. 20. [Google Scholar]
- Titlyanova, A.A.; Sambuu, A.D. Succession in Grasslands; Publishing House SB RAS: Novosibirsk, Russia, 2016; p. 191. [Google Scholar]
- Mironycheva-Tokareva, N.P.; Mikhailova, E.V.; Vishnyakova, E.K. Transformation of the vegetation cover of marsh ecosystems in the construction of the roadway (on the example of the taiga zone of Western Siberia). Vestn. Orenb. Gos. Univ. 2017, 12, 55–59. [Google Scholar]
- Tiunov, A.V. Structure and Functions of Soil Communities of a Monsoon Tropical Forest (Cat Tien National Park, Southern Vietnam); KMK Scientific Press: Moscow, Russia, 2011; p. 277. [Google Scholar]
- Tiunov, A.V. Stable isotopes of carbon and nitrogen in soil ecological studies. Biol Bull. Russ. Acad. Sci. 2007, 34, 395–407. [Google Scholar] [CrossRef]
- Robinson, D. δ15N as an integrator of the nitrogen cycle. Trends Ecol. Evol. 2001, 16, 153–162. [Google Scholar] [CrossRef]
- Fry, B. Stable Isotope Ecology; Springer: New York, NY, USA, 2006; Volume 521, p. 318. [Google Scholar]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and nitrogen dynamics along the decay continuum: Plant litter to soil organic matter. Plant. Soil 1989, 115, 189–198. [Google Scholar] [CrossRef]
- Wedin, D.A.; Tieszen, L.L.; Dewey, B.; Pastor, J. Carbon isotope dynamics during grass decomposition and soil organic matter formation. Ecology 1995, 76, 1383–1392. [Google Scholar] [CrossRef]
- Amundson, R.; Baisden, W.T. Stable isotope tracers and mathematical models in soil organic matter studies. In Methods in ecosystem science; Springer: New York, NY, USA, 2000; pp. 117–137. [Google Scholar]
- Connin, S.L.; Feng, X.; Virginia, R.A. Isotopic discrimination during long-term decomposition in an arid land ecosystem. Soil Biol. Biochem. 2001, 33, 41–51. [Google Scholar] [CrossRef]
- Lehmann, M.F.; Bernasconi, S.M.; Barbieri, A.; McKenzie, J.A. Preservation of organic matter and alteration of its carbon and nitrogen isotope composition during simulated and in situ early sedimentary diagenesis. Geochim. Cosmochim. Acta 2002, 66, 3573–3584. [Google Scholar] [CrossRef]
- Fellerhoff, C.; Voss, M.; Wantzen, K.M. Stable carbon and nitrogen isotope signatures of decomposing tropical macrophytes. Aquatic Ecol. 2003, 37, 361–375. [Google Scholar] [CrossRef]
- Fernandez, L.; Mahieu, N.; Cadisch, G. Carbon isotopic fractionation during decomposition of plant materials of different quality. Global Biogeochem. Cycles 2003, 17, 1075. [Google Scholar] [CrossRef]
- Ågren, G.I.; Bosatta, E.; Balesdent, J. Isotope discrimination during decomposition of organic matter: A theoretical analysis. Soil Sci. Soc. Am. J. 1996, 60, 1121–1126. [Google Scholar] [CrossRef]
- Handley, L.L.; Raven, J.A. The use of natural abundance of nitrogen isotopes in plant physiology and ecology. Plant. Cell Environ. 1992, 15, 965–985. [Google Scholar] [CrossRef]
- Högberg, P. Tansley review no. 95 15N natural abundance in soil–plant systems. New Phytol. 1997, 137, 179–203. [Google Scholar] [CrossRef]
- Kendall, C. Tracing nitrogen sources and cycling in catchments. In Isotope Tracers in Catchment Hydrology; Kendall, C., McDonnell, J.J., Eds.; Elsevier Science, B.V.: Amsterdam, The Netherlands, 1998; pp. 519–576. [Google Scholar]
- Bunting, M.J.; Warner, B.G.; Aravena, R. Late Quaternary vegetation dynamics and hydroseral development in a Thuja occidentals swamp in southern Ontario. Can. J. Earth Sci. 1996, 33, 1439–1456. [Google Scholar] [CrossRef]
- Lowe, J.J.; Walker, M.J.C. Reconstructing Quaternary Environments, 2nd ed.; Addison Wesley Longman: Harlow, UK, 1997; p. 446. [Google Scholar]
- Vardy, S.R.; Warner, B.G.; Aravena, R. Holocene Climate Effects on the Development of a Peatland on the Tuktoyaktuk Peninsula, Northwest Territories. Quat Res. 1997, 47, 90–104. [Google Scholar] [CrossRef]
- Jasinski, J.P.P.; Warner, B.G.; Andreev, A.A.; Aravena, R.; Gilbert, S.E.; Zeeb, B.A.; Smol, J.P.; Velichko, A.A. Holocene environmental history of a peatland in the Lena River valley, Siberia. Can. J. Earth Sci. 1998, 35, 637–648. [Google Scholar] [CrossRef]
- Wolfe, B.B.; Edwards, T.W.D.; Aravena, R.; Forman, S.L.; Warner, B.G.; Velichko, A.A.; MacDonald, G.M. Holocene paleohydrology and paleoclimate at treeline, north-central Russia, inferred from oxygen isotope records in lake sediment cellulose. Quat. Res. 2000, 53, 319–329. [Google Scholar] [CrossRef]
- Earle, L.; Warner, B.G.; Aravena, R. Rapid development of an unusual peat-accumulation ecosystem in the Chilean Altiplano. Quat. Res. 2003, 59, 2–11. [Google Scholar] [CrossRef]
- Asada, T.; Warner, B.; Aravena, R. Effects of early stage of decomposition on change in carbon and nitrogen isotopes in Sphagnum litter. J. Plant. Interact. 2005, 1, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Golovatskaya, E.A.; Nikonova, L.G. Influence of the level of bog waters on the processes of transformation of sphagnum mosses in peat soil of oligotrophic bogs. Eurasian Soil Sci. 2017, 5, 580–588. [Google Scholar] [CrossRef]
- Drollinger, S.; Kuzyakov, Y.; Glatzel, S. Effects of peat decomposition on δ13C and δ15N depth profiles of Alpine bogs. Catena 2019, 178, 1–10. [Google Scholar] [CrossRef]
- Drollinger, S.; Knorr, K.H.; Knierzinger, W.; Glatzel, S. Peat decomposition proxies of Alpine bogs along a degradation gradient. Geoderma 2020, 369, 114331. [Google Scholar] [CrossRef]
- Bird, J.A.; Kleber, M.; Torn, M.S. 13C and 15N stabilization dynamics in soil organic matter fractions during needle and fine root decomposition. Org. Geochem. 2008, 39, 465–477. [Google Scholar] [CrossRef]
- Simonova, G.; Volkov, Y.; Markelova, A.; Kalashnikova, D. Isotope monitoring of forest ecosystems. In Proceedings of the International Multidisciplinary Scientific GeoConference: SGEM, Albena, Bulgaria, 29 June–5 July 2017; pp. 885–890. [Google Scholar]
- Nikonova, L.G. Production and Destruction Processes in Oligotrophic Bog Ecosystems of the Southern Taiga Subzone of Western Siberia. Ph.D. Dissertation, Siberian Federal University, Krasnoyarsk, Russia, 2020. [Google Scholar]
- Bazanov, V.A.; Egorov, B.A.; L’gotin, V.A.; Skugarev, A.A. Modern spatial dynamics of the Great Vasyugan Mire (the case of the Iksa-Shegarka interfluve). In The Great Vasyugan Mire Current status and development processes; Kabanov, M.V., Ed.; Institute of Atmospheric Optics SB RAN: Tomsk, Russia, 2002; pp. 190–196. [Google Scholar]
- Kurakov, S.A.; Krutikov, V.A.; Ushakov, V.G. Autonomous meter of the temperature profile of the AIPT. Appar. Exp. Tech. 2008, 5, 166–167. [Google Scholar]
- Ponomareva, V.V.; Plotnikova, T.A. Simultaneous Determination of Overall C and N Content in Peat Soils Using the Anstet Method, Modification Ponomareva and Nikolaeva. Methods of Determination of Humus Content and Composition in Soils (Mineral and Peat) in Russia; Federal Academy of Agricultural Sciences VI Lenin; Central Pedology Museum VV Dokuchaev: Leningrad, Russia, 1975; pp. 79–83. (in Russian) [Google Scholar]
- Vorobyeva, L.A. Theory and Practice of Chemical Analysis of Soils; GEOS: Moscow, Russia, 2006; p. 400. [Google Scholar]
- Lebedev, A.T. Mass Spectrometry for the Analysis of the Environment; Tekhnosfer publ: Moscow, Russia, 2013; p. 632. [Google Scholar]
- Dobrovolskaya, T.G.; Golovchenko, A.V.; Zvyagintsev, D.G. Functions of Microbial Complexes of the High-Moor Peatbogs: Reasons for Slow Peat Destruction; Chernov, I.Y., Ed.; KMK: Moscow, Russia, 2013; p. 131. [Google Scholar]
- Melillo, J.M.; Aber, J.D.; Muratore, J.F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology 1982, 63, 621–626. [Google Scholar] [CrossRef]
- Sariyildiz, T.; Anderson, J.M. Variation in the chemical composition of green leaves and leaf litters from three deciduous tree species growing on different soil types. For. Ecol. Manag. 2005, 210, 303–319. [Google Scholar] [CrossRef]
- Scheffer, R.A.; van Logtestijn, R.S.P.; Verhoeven, J.T.A. Decomposition of Carex and Sphagnum litter in two mesotrophic fens differing in dominant plant species. Oikos 2001, 92, 44–54. [Google Scholar] [CrossRef]
- Heal, O.W.; French, D.D. Decomposition of organic matter in tundra. In Soil Organisms and Decomposition in Tundra; IBP Tundra Biome: Stockholm, Sweden, 1974; pp. 279–311. [Google Scholar]
- Pereverzev, V.N.; Evdokimova, G.A. Biochemistry of Humus and Nitrogen Soils of the Kola Peninsula; Nauka: Leningrad, 1987; p. 303. [Google Scholar]
- Billett, M.F.; Palmer, S.M.; Hope, D.; Deacon, C.; Storeton-West, R.; Hargreaves, K.J.; Flechard, C.; Fowler, D. Linking land-atmosphere-stream carbon fluxes in a lowland peatland system. Glob. Biogeochem. Cycles 2004, 18, GB1024. [Google Scholar] [CrossRef]
- Rochefort, L.; Vitt, D.H.; Bayley, S.E. Growth, production, and decomposition dynamics of Sphagnum under natural and experimentally acidified conditions. Ecology 1990, 71, 1986–2000. [Google Scholar] [CrossRef]
- Szumigalski, A.R.; Bayley, S.E. Decomposition along a bog to rich fen gradient in central Alberta, Canada. Can. J. Botany 1996, 74, 573–581. [Google Scholar] [CrossRef]
- Hajek, T. Habitat and species controls on Sphagnum production and decomposition in a mountain bog. Boreal Environ. Res. 2009, 14, 947–958. [Google Scholar]
- Maass, W.S.G.; Craigie, J.S. Examinations of some soluble constituents of Sphagnum gametophytes. Can. J. Botany 1964, 42, 805–813. [Google Scholar] [CrossRef]
- Kracht, O.; Gleixner, G. Isotope analysis of pyrolysis products from Sphagnum peat and dissolved organic matter from bog water. Org. Geochem. 2000, 3, 645–654. [Google Scholar] [CrossRef]
- Andrews, J.A.; Matamala, R.; Westover, K.M.; Schlesinger, W.H. Temperature effects on the diversity of soil heterotrophs and the δ13C of soil-respired CO2. Soil Biol. Biochem. 2000, 32, 699–706. [Google Scholar] [CrossRef]
- Benner, R.; Fogel, M.L.; Sprague, E.K.; Hodson, R.E. Depletion of 13C in lignin and its implications for stable carbon isotope studies. Nature 1987, 329, 708–710. [Google Scholar] [CrossRef]
- Adams, M.A.; Grierson, P.F. Stable isotopes at natural abundance in terrestrial plant ecology and ecophysiology: An update. Plant. Biol. 2001, 3, 299–310. [Google Scholar] [CrossRef]
- Thormann, M.N.; Bayley, S.E. Decomposition along a moderate-rich fen-marsh peatland gradient in boreal Alberta, Canada. Wetlands 1997, 17, 123–137. [Google Scholar] [CrossRef]
Figure 1.
Site locations of studied peatlands, Western Siberia (Tomsk region).

Figure 2.
Weather conditions: average monthly air temperature and precipitation, average for 2019–2021.
Figure 2.
Weather conditions: average monthly air temperature and precipitation, average for 2019–2021.

Figure 3.
Dynamics of mass loss of Sph. fuscum and E. vaginatum at different observation points.

Figure 4.
Dynamics of carbon content in the process of transformation of plant residues of Sph. fuscum and E. vaginatum.
Figure 4.
Dynamics of carbon content in the process of transformation of plant residues of Sph. fuscum and E. vaginatum.

Figure 5.
Variability of the carbon isotope composition of peat-forming plants during the biochemical transformation.
Figure 5.
Variability of the carbon isotope composition of peat-forming plants during the biochemical transformation.

Figure 6.
Dependence of the carbon isotope composition on mass loss.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Percentage of mass loss and chemical composition in initial samples and after 3 years of field decomposition.
Table 1.
Percentage of mass loss and chemical composition in initial samples and after 3 years of field decomposition.
| Plant Species | Site | Mass Loss, % | C, % | δ13C, ‰ | N, % | C/N | Lig | Lig/N |
|---|---|---|---|---|---|---|---|---|
| Sph. fuscum | Initial sample | 45.1 ± 0.6 | −31.2 ± 0.2 | 0.4 ± 0.0 | 118 ± 1 | 12.2 | 32 | |
| VASnat | 30.9 ± 8.9 | 47.3 ± 0.5 | −30.1 ± 0.1 | - | - | - | - | |
| VASdry | 27.6 ± 3.7 | 45.9 ± 1.4 | −29.9 ± 0.2 | - | - | - | - | |
| Iksa1 | 19.3 ± 1.1 | 44.2 ± 0.8 | −30.1 ± 0.2 | - | - | - | - | |
| Iksa2 | 22.3 ± 5.3 | 45.5 ± 0.1 | −30.0 ± 0.2 | - | - | - | - | |
| E. vaginatum | Initial sample | 47.0 ± 0.4 | −26.1 ± 0.2 | 1.0 ± 0.0 | 49 ± 0.1 | 20.3 | 22 | |
| VASnat | 56.6 ± 5.8 | 47.4 ± 1.9 | −27.9 ± 0.1 | - | - | - | - | |
| VASdry | 67.9 ± 1.4 | 47.8 ± 0.1 | −28.3 ± 0.6 | - | - | - | - | |
| Iksa1 | 67.4 ± 3.4 | 47.6 ± 0.6 | −27.9 ± 0.2 | - | - | - | - | |
| Iksa2 | 64.9 ± 5.4 | 47.5 ± 0.6 | −27.4 ± 0.2 | - | - | - | - |
Table 2.
Summary of the ANOVA for the effect of plant species and location of the samples on value δ13C in plant residuals.
Table 2.
Summary of the ANOVA for the effect of plant species and location of the samples on value δ13C in plant residuals.
| Factor | Df | F Value | p |
|---|---|---|---|
| Plant species | 1 | 168.238 | <0.05 |
| Sites | 3 | 1.3 | <0.05 |
| Plant species × Sites | 3 | 0.892 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Golovatskaya, E.; Nikonova, L.; Simonova, G.; Kalashnikova, D. Variability of the Carbon Isotope Composition of Peat-Forming Plants during the Biochemical Transformation. Water 2022, 14, 4035. https://doi.org/10.3390/w14244035
AMA Style
Golovatskaya E, Nikonova L, Simonova G, Kalashnikova D. Variability of the Carbon Isotope Composition of Peat-Forming Plants during the Biochemical Transformation. Water. 2022; 14(24):4035. https://doi.org/10.3390/w14244035
Chicago/Turabian StyleGolovatskaya, Evgeniya, Liliya Nikonova, Galina Simonova, and Daria Kalashnikova. 2022. "Variability of the Carbon Isotope Composition of Peat-Forming Plants during the Biochemical Transformation" Water 14, no. 24: 4035. https://doi.org/10.3390/w14244035
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.