
Composition of Colloidal Organic Matter in Phytoplankton Exudates


1
Marine Biology Station, National Institute of Biology, Fornace 41, 6330 Piran, Slovenia
2
Slovenian NMR Centre, National Institute of Chemistry, Hajdrihova 19, 1000 Ljubljana, Slovenia
3
National Institute of Oceanography and Applied Geophysics, Via A. Piccard 54, 34151 Trieste, Italy
4
Department of Eviron. Sciences, Jozef Stean Institute, Jamova 39, 1000 Ljubljana, Slovenia
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 111; https://doi.org/10.3390/w15010111
Submission received: 30 November 2022
/
Revised: 15 December 2022
/
Accepted: 26 December 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Characterization of Organic Matter in Marine and Freshwater Environment)
Abstract
:The colloidal organic matter (COM) was isolated from the exudates of three cultured phytoplonkters, namely the chlophyte nanoflagellate Tetraselmis sp., the diatom Chaetoceros socialis and the dinoflagellate Prorocentrum minimum, from the Gulf of Trieste (northern Adriatic Sea). The isolation of COM was performed by ultrafiltration with molecular weight cut-off membranes of 5 kDa and final desalinisation by dialysis. The composition of the COM was characterised using C elemental analysis and 1H NMR spectroscopy and compared with COM isolated from a marine sample from the same area (Gulf of Trieste). By using 1H NMR spectroscopy, it was possible to semi-quantitatively determine the concentrations of the main biochemical constituents present in the COM samples. The results showed that the phytoplankton COM was predominantly composed of polysaccharides, with minor contributions from proteins and especially lipids. Therefore, the phytoplankton COM mainly contributes to the marine COM pool in the polysaccharide fraction and less in the protein and lipid fractions.
1. Introduction
Dissolved organic matter (DOM) is an important component of the marine carbon cycle [1] operationally defined as a fraction that passes through a filter of a 0.22 (0.45) μm nominal pore size and which also encompasses colloidal organic matter (COM). COM (from 1 kDa (5 kDa) to 0.22 (0.45) μm) is composed of important biochemical constituents such as lipids, carbohydrates, proteins [2] and humates [1]. Colloidal organic matter (COM) represents approximately 25% of dissolved organic matter (DOM) [3,4]. Recent research has shown the various biochemical functions of COM, especially the role of its biological degradable component in the marine microbial loop [5,6]. Marine COM can act as a precursor in the formation of aggregates and macroaggregates [3,7,8] (i.e., macrogels, presumably of phytoplankton origin, which have periodically occurred in the northern Adriatic Sea in the past, usually in the period of late spring–early summer) [9]. In this context, the composition of phytoplankton exudates could be an important step in decoding the origin of marine COM, most of which has not yet been chemically characterised. It is well known that due to the complexity of marine COM’s composition, it is difficult to elucidate its chemical composition and structure [1,4].
Nuclear magnetic resonance (NMR) spectroscopy has been shown to be a crucial, powerful method for the compositional and structural characterisation of aquatic COM and DOM at the molecular level [4,10,11,12,13]. NMR is viewed as highly complementary and not overlapping with mass spectrometry (MS), which has become the dominant technique in this research field due to its greater sensitivity [14]. However, NMR is simple, non-destructive, quantitative and reproducible for the analysis of DOM in various aquatic media [10], including those in the exudates of phytoplankton and bacterial cultures [14]. As the high salt content interferes with NMR as well as MS, and due to the low DOM concentration, ultrafiltration isolation or solid phase extraction are used for NMR analysis [2].
The aim of this study is to provide a better understanding of the origin and chemical composition of COM (and DOM) in coastal waters, decoding the composition of COM released by some cultured phytoplankters isolated from the coastal area of the Gulf of Trieste (northern Adriatic Sea) in terms of organic C and 1H NMR spectroscopic analyses. Comparisons are made with the marine COM and the macroaggregate compositional data, which are thought to be a product of the COM agglomeration in this area [13,15,16].
2. Materials and Methods
2.1. Culture Methods
The phytoplankton species of the chlorophyte nanoflagellate Tetraselmis sp., the diatom Chaetoceros socialis and the dinoflagellate Prorocentrum minimum isolated from the Gulf of Trieste were grown in batch cultures with enriched seawater (medium ESAW [17]) under 75 µEinstein m−2 s−1 of fluorescent light in a cycle of 12 h of light and 12 h of dark at a constant temperature of 16 °C. The cultures were not axenic. They were first preconditioned in 100 mL of ESAW medium and inoculated in 1 l of ESAW medium after 2 weeks.
In the stationary phase of growth, the cultures, with cell densities ranging between 3.5 × 105 and 1.5 × 106 cells mL−1, were filtered through preignited 0.7 µm pore size Whatman GF/F glass-fibre filters. The filtrates were then filtered through 0.22 µm Nucleopore filters. In order to isolate the COM, the filtrates were ultrafiltered through membranes with a nominal molecular weight cut-off (MWCO) of 5 kDa using a Vivascience Vivaflow 200 unit (Sartorius) with a Masterflex S/L membrane pump (Cole-Palmer) at a flow rate of 300 mL min−1 at 2.5 bar and 20 °C. The average concentration factor (CF) was 6. The retentates were freeze-dried and desalted by dialysis [18] in Milli-Q water for 18 h at 4 °C using MWCO 1 kDa RC membranes (Spectrapor 7, Spectrum Lab). The surrounding milliQ water was exchanged twice after 2 and 4 h to reduce the salinity from PSU 37 to PSU < 0.2.
2.2. Analyses
DOC Analysis
Analyses of the DOC in the filtrates, retentates and permeates were performed with a high-temperature catalytic method using a Shimadzu TOC-L analyser after acidifying the samples with 6 M HCl to eliminate CO2 [19]. The reproducibility was between 1.5 and 3.0%.
2.3. NMR Spectroscopy
The 1H-NMR spectra of the dialysed COM samples were obtained using an Agilent Technologies VNMRS 800 MHz NMR spectrometer in D2O at a temperature of 298 K using a cold probe. Standard 1D 1H-NMR spectra were acquired with the use of DPFGSE solvent suppression. The number of scans was 256, the pulse width was 7.7 ms, the spectral width was 16,000 Hz, the acquisition time was 1 s, and the relaxation delay was 1.5 s.
The 1H-NMR spectroscopy enabled quantitative analysis because the signal intensity was proportional to the amount of compounds present in the sample. In this study, semi-quantitative analysis was performed by integrating (“trajectory lines” in Figure 1, Figure 2, Figure 3 and Figure 4) specific areas in the 1H-NMR spectra that could be assigned to the main biochemical groups. The sum of all 4 integrals was 100%. The integrals were expressed as the relative abundance (in%) of the main biochemical groups (lipids, proteins and CRAM, polysaccharides and formate) in each sample.
3. Results and Discussion
3.1. COM Concentration
The total extracellular Corg. (DOC) content results in the exudates of the three phytoplankton cultures (Table 1) showed that the highest concentration was found in the diatom Chaetoceros socialis, followed by the nanoflagellate Tetraselmis sp. and the dinoflagellate Prorocentrum minimum. This was the opposite of the previously published Corg. elemental composition of cultured T. marina (60% dry weight) and C. socialis (25% dry weight) [20]. The isolated colloidal organic carbon (COC) (calculated as % COC) also showed the opposite order: 39.8% in the nanoflagellate, 35.1% in the dinoflagellate and 23%, in the diatoms, releasing the lowest percentage of extracellular colloidal organic matter. The latter value was similar to the generally reported value that COM represents on average (about 25% of dissolved organic matter (DOM)) [4]. Previous studies showed that in summer 2012, the COM began to accumulate in the Gulf of Trieste, when the percentage of COC in the bulk DOC rose twofold from 16.3 to 32.4% [13]. The COM accumulation from late spring to early summer indicated an increase in refractory COM in the water column and the possibility of the formation of aggregates leading to the episodic (periodic) occurrence of macroaggregates in the northern Adriatic Sea, as has occurred in the past [15,16]. This is in accordance with the progressive increase in DOC concentrations in the Gulf of Trieste towards summer [13,21,22] after the early spring (primary) productive period, while the bacterial production reached its peak in summer [23]. Similar temporal dynamics were reported for the total dissolved carbohydrates and particularly for dissolved polysaccharides in the water column of the Gulf of Trieste, which were hypothesised to be the macroaggregate precursors [24].
3.2. COM Composition
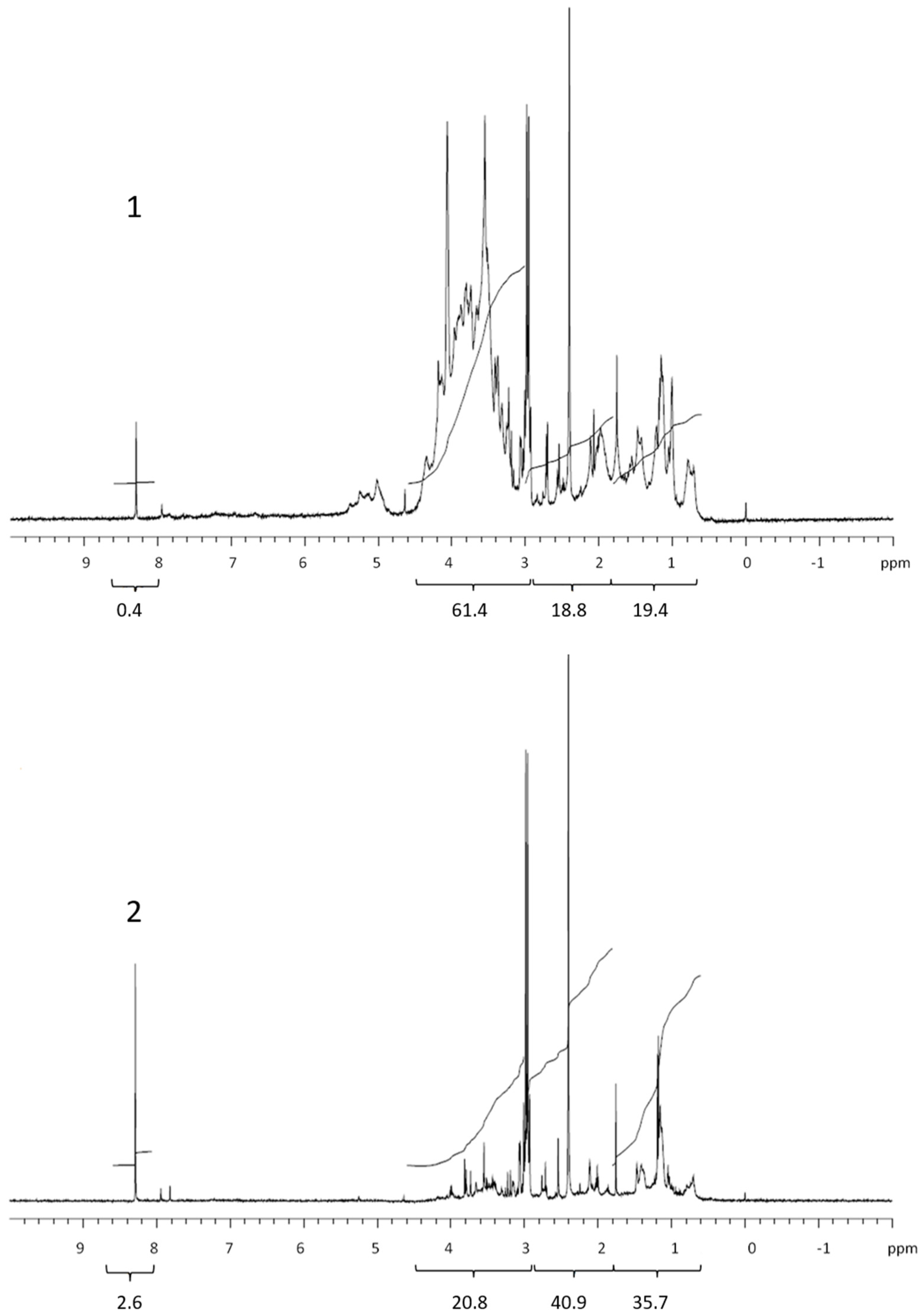
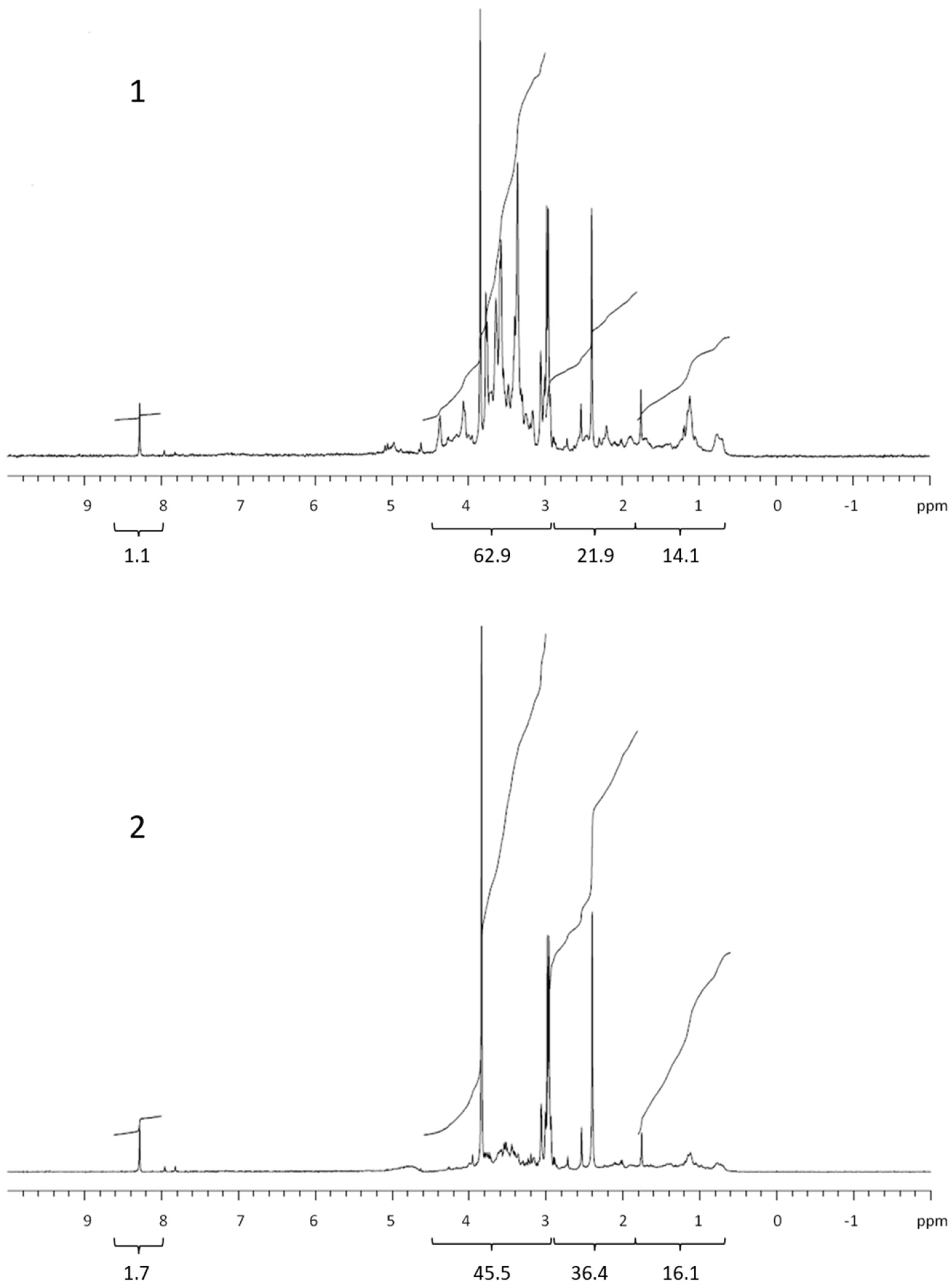
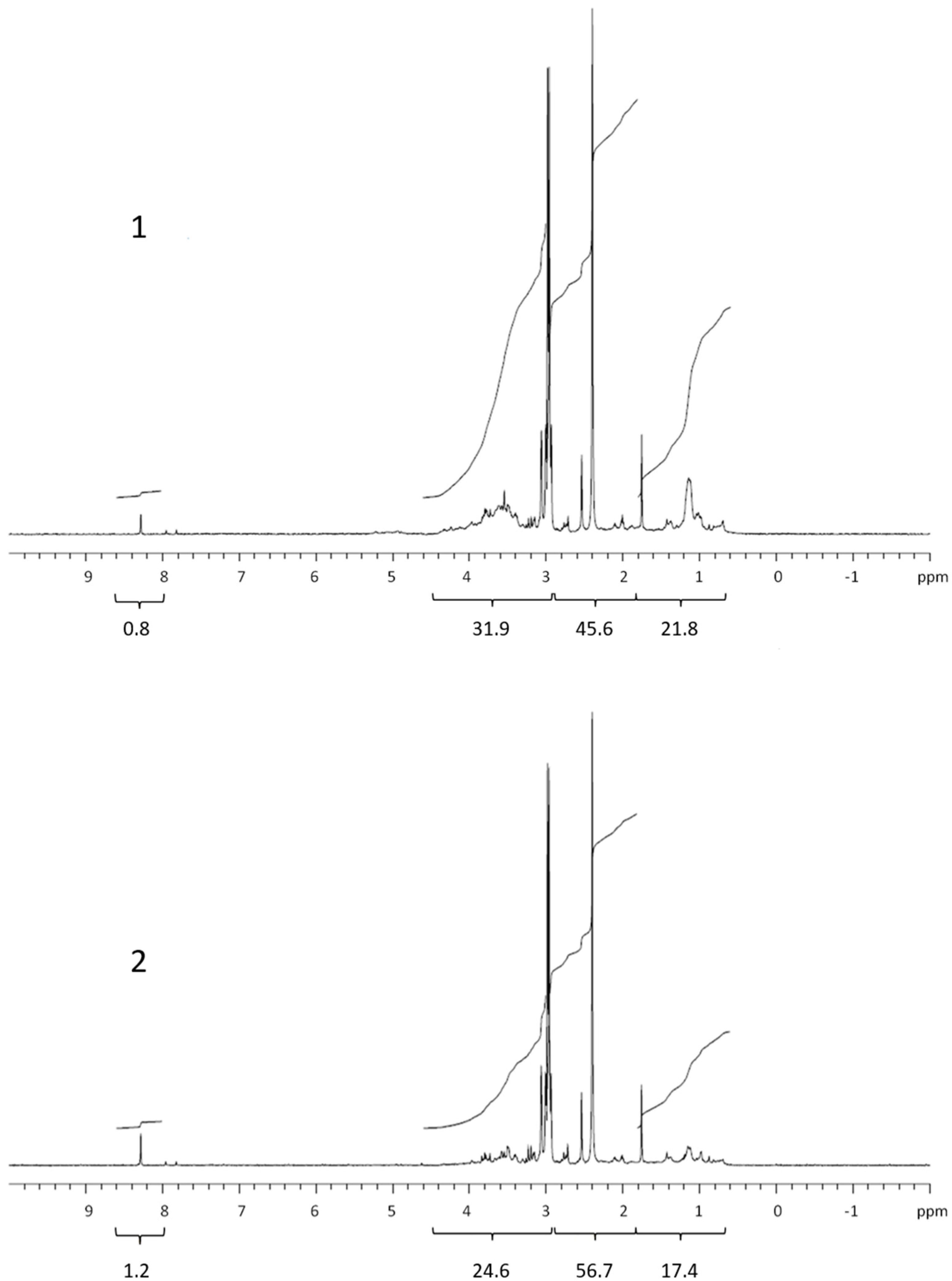
The 1H NMR spectra of the dialysed marine COM (Figure 1, Figure 2, Figure 3 and Figure 4) can be divided into the main groups of proton resonance, which can be assigned to major biochemical components present in the marine environment: protons bonded on or near carbon in alcohol or ether functional groups (polysaccharides; HC-OH, HC-O-C-: δ = 3–4.6 ppm), protons near the amide, ketone and carboxyl functional groups in alicyclic organic molecules (CRAM) (proteins, CRAM; HC-HCOR: δ =1.8–3 ppm) and aliphatic protons (lipids; HCH2-CH2-: δ = 0.6–1.8 ppm) [10,13,15,25,26]. The spectra of all samples exhibited a peak in formate (δ = 8.3 ppm) [27], with higher abundances in the permeates compared with the retentates, which probably originated from the formate esters in the DOM. The spectra, corresponding to the studied phytoplankton exudates, showed peaks for all the main COM components (Figure 1, Figure 2 and Figure 3). Integration revealed that the main difference was represented by polysaccharides and proteins, while lipids and especially formate represented the minor fractions (Table 1). When comparing the samples of cultured phytoplankton species, the polysaccharide fraction was the highest in the retentate of the nanoflagellate Tetraselmis sp. (61.4%), while lipids and proteins accounted for 19.4 and 18.8%, respectively (Table 1). The composition of permeate was different, as proteins represented the highest portion (40.9%). The diatom Chaetoceros socialis also contained the highest polysaccharide level for retentates (62.9%) and permeates (45.5%), followed by proteins and lipids. The retentates and permeates of the dinoflagellate Prorocentrum minimum were quite different, exhibiting the highest levels of proteins and the lowest levels of lipids.
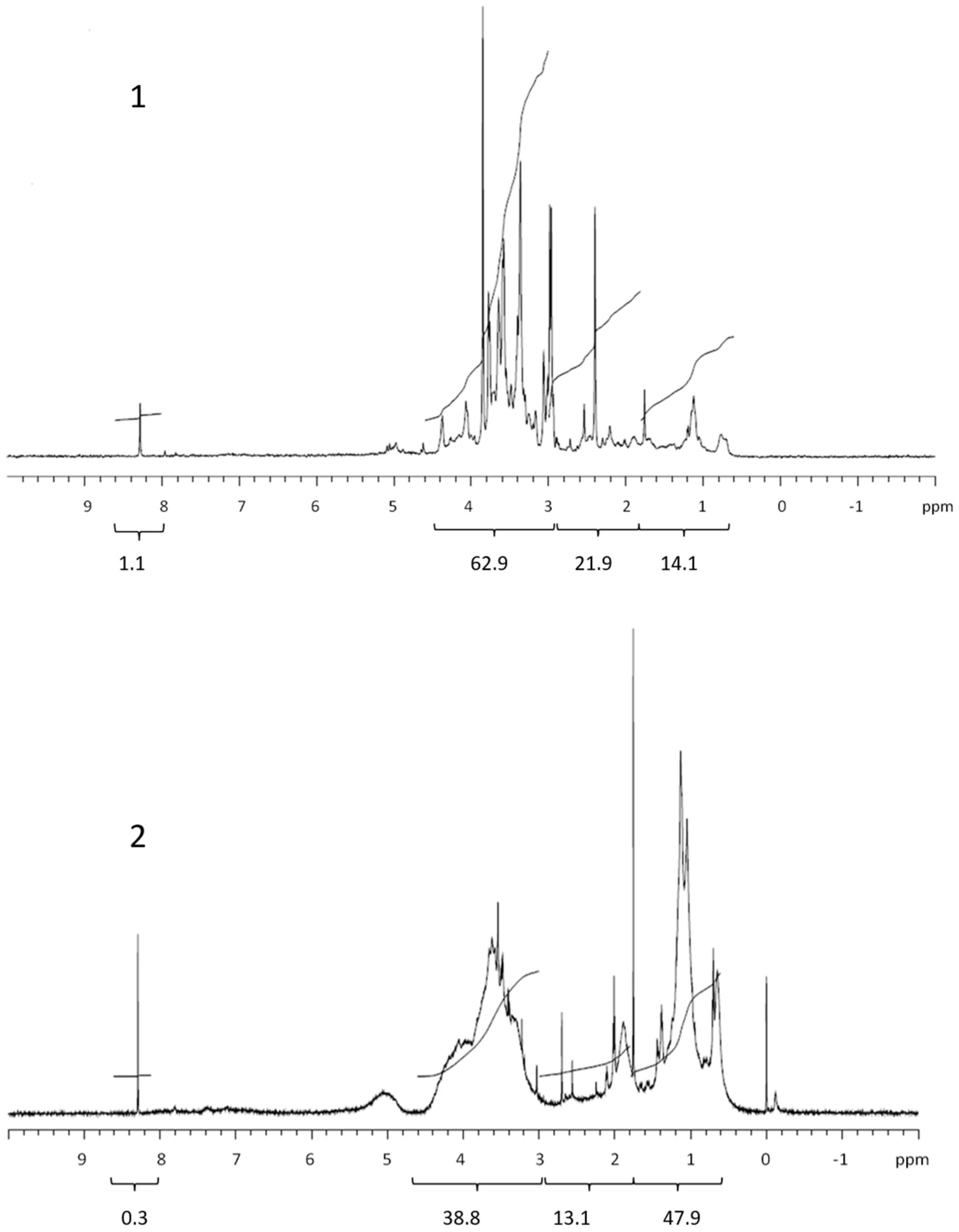
The quantity and composition of the phytoplankton extracellular DOM depend on the organism, its physiological status, the physical and chemical environmental properties and the presence of other organisms [28]. The Corg. levels, 1H NMR spectra and their integrals differed among the studied phytoplankton species. The colloids from the nanoflagellates and diatoms contained mainly polysaccharides, in accordance with the high carbohydrate concentrations in the cell [20], supporting previous conclusions that carbohydrates are major components of phytoplankton exudates [29]. Conversely, the colloids from the dinoflagellates contained mainly proteins. The COM of all phytoplankton cultures exhibited lower lipid contents in comparison with marine COM [13]. Kovac et al. (2002) [15] reported on the presence of lipids (aliphatic) followed by polysaccharide fractions as the major constituents of macroaggregates of phytoplankton origin, with lower concentrations of proteins. The compositional differences between COM in phytoplankton cultures and marine COM and macroaggregates indicate the refractory nature of lipids in the marine environment [30]. However, the differences observed could also be due to different solvents used in 1H NMR analysis of COM and macroaggregates. Comparing the 1H NMR spectrum of the nanoflagellate Tetraselmis sp. COM with marine COM from the Gulf of Trieste [13] revealed a similarity, since nanoflagellates together with diatoms represent the majority of the phytoplankton found in the Gulf of Trieste [31].
4. Conclusions
The COM isolated from the exudates of three cultured phytoplankters, namely the nanoflagellate Tetraselmis sp., the diatom Chaetoceros socialis and the dinoflagellate Prorocentrum minimum, from the Gulf of Trieste is predominantly composed of polysaccharides, with minor contributions from proteins and particularly lipids. The phytoplankton COM contributes to the composition of marine COM, particularly through an increase in the polysaccharide fraction during COM agglomeration and possible macroaggreagate formation, as observed in the past in the northern Adriatic Sea, while the accumulation of lipids is due to their refractory nature and resistance to microbial and chemical degradation.
Author Contributions
J.F. designed the study and drafted the manuscript; K.K. drafted and submitted the manuscript and obtained and analysed the data; P.Š. performed 1H NMR analysis and revised the manuscript; I.F. helped with the study design and revised the manuscript; A.B. provided the phytoplankton cultures and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Slovenian Research Agency (research core funding no. P1-0143, P4-0432 and P1-0242).
Data Availability Statement
During this study no additional data were created.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hedges, J.I. Why dissolved organic matter? In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Academic Press: Boston, MA, USA, 2002; pp. 1–33. [Google Scholar]
- Fillela, M. Environmental colloids and particles (behavior, separation and characterisation). In Environmental Colloids and Particles; Wilkinson, K.J., Lead, J.R., Eds.; John Wiley & Sons: Chichester, UK, 2007; pp. 17–94. [Google Scholar]
- Verdugo, P.; Alldredge, A.L.; Azam, F.; Kirchman, D.L.; Passow, U.; Santschi, P.H. The oceanic gel phase: A bridge in the DOM-POM continuum. Mar. Chem. 2004, 92, 67–85. [Google Scholar] [CrossRef]
- Mopper, K.; Stubbins, A.M.; Ritchie, J.D.; Bialk, H.M.; Hatcher, P.G. Advanced instrumental approaches for characterization of marine dissolved matter: Extraction techniques, mass spectrometry, and nuclear magnetic resonance spectroscopy. Chem. Rev. 2007, 107, 419–442. [Google Scholar]
- Benner, R. Chemical composition and reactivity. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Elsevier: San Diego, CA, USA, 2002; pp. 59–90. [Google Scholar]
- Moran, M.A.; Kujawinski, E.B.; Schroer, W.F.; Amin, S.A.; Bates, N.R.; Bertrand, E.M.; Braakman, R.; Brown, R.; Covert, C.T.; Doney, S.C.; et al. Microbial metabolites in the marine carbon cycle. Nat. Microbiol. 2022, 7, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Chin, W.C.; Orellana, M.V.; Verdugo, P. Spontaneous assembly of marine dissolved organic matter into polymer gels. Nature 1998, 391, 568–572. [Google Scholar] [CrossRef]
- Verdugo, P.; Santschi, P.H. Polymer dynamics of DOC network and gel formation in seawater. Deep Sea Res. II 2010, 57, 1486–1493. [Google Scholar] [CrossRef]
- Giani, M.; Degobbis, D.; Rinaldi, A. Mucilages in the Adriatic and Tyrrhenian Seas. Sci. Total Environ. 2005, 353, 1–2. [Google Scholar] [CrossRef]
- Lam, B.; Simpson, A.J. Direct 1H NMR spectroscopy of dissolved organic matter in natural waters. Analyst 2008, 133, 263–269. [Google Scholar] [CrossRef]
- Hertkorn, N.; Benner, R.; Frommberger, M.; Schmitt-Kopplin, P.; Witt, M.; Kaiser, K.; Kettrup, A.; Hedges, J.I. Characterizationn of a major refractory component of marine dissolved organic matter. Geochim. Cosmochim. Acta 2006, 70, 2990–3010. [Google Scholar] [CrossRef]
- Hertkorn, N.; Harir, M.; Koch, B.P.; Michalke, B.; Schmitt-Kopplin, P. High-field NMR spectroscopy and FTICR mass spectrometry: Powerful discovery tools for the molecular level characterization of marine dissolved organic matter. Biogeosciences 2013, 10, 1583–1624. [Google Scholar] [CrossRef] [Green Version]
- Klun, K.; Šket, P.; Falnoga, I.; Faganeli, J. Variation of colloidal organic matter composition in coastal waters (Gulf of Trieste, northern Adriatic Sea). Geomicrobiol. J. 2015, 32, 609–615. [Google Scholar] [CrossRef]
- Edison, A.S.; Colonna, M.; Gouvenia, G.J.; Holderman, N.R.; Judge, M.Z.; Shen, X.; Zhang, S. NMR: Unique strengths that enhance modern metabolomics research. Anal. Chem. 2021, 93, 478–499. [Google Scholar] [CrossRef] [PubMed]
- Kovac, N.; Bajt, O.; Faganeli, J.; Sket, B.; Orel, B. Study of macroaggregate composition using FT-IR and 1H-NMR spectroscopy. Mar. Chem. 2002, 78, 205–215. [Google Scholar] [CrossRef]
- Kovac, N.; Faganeli, J.; Bajt, O.; Sket, B.; Orel, B.; Penna, N. Chemical composition of macroaggregates in the northern Adriatic Sea. Org. Geochem. 2004, 35, 1095–1104. [Google Scholar] [CrossRef]
- Harrison, P.J.; Waters, R.E.; Taylor, F.J.R. A broad spectrum artificial seawater medium for coastal and open ocean phytoplankton. J. Phycol. 1980, 16, 28–35. [Google Scholar]
- Engel, A.; Handel, N. A novel protocol for determining the concentration and composition of sugars and particulate and high molecular dissolved organic matter (HMW DOM) in sea water. Mar. Chem. 2011, 127, 180–191. [Google Scholar] [CrossRef] [Green Version]
- Sugimura, Y.; Suzuki, Y. A high temperature catalytic oxidation method for the determination of non-volatile dissolved organic carbon in seawater by direct injection of liquid sample. Mar. Chem. 1988, 24, 105–131. [Google Scholar] [CrossRef]
- Faganeli, J.; Pezdič, J.; Fanuko, N. Chemical and isotopic composition of some phytoplankton species. Biol. Vestn. 1989, 37, 9–20. [Google Scholar]
- Faganeli, J.; Herndl, G.J. Dissolved organic matter in the waters of the Gulf of Trieste (Northern Adriatic). Thalassia Jugosl. 1991, 23, 51–63. [Google Scholar]
- De Vittor, C.; Paoli, A.; Fonda Umani, S. Dissolved organic carbon variability in a shallow coastal marine system (Gulf of Trieste, northern Adriatic Sea). Estuar. Coast. Shelf Sci. 2008, 78, 280–290. [Google Scholar] [CrossRef]
- Fonda Umani, S.; Del Negro, P.; Larato, C.; De Vittor, C.; Cabrini, M.; Celio, M.; Falconi, C.; Tamberich, F.; Azam, F. Major inter-annual variations in microbial dynamics in the Gulf of Trieste (northern Adriatic Sea) and their ecosystem implications. Aquat. Microb. Ecol. 2007, 46, 163–175. [Google Scholar] [CrossRef]
- Penna, N.; Kovac, N.; Ricci, F.; Penna, A.; Capellacci, S.; Faganeli, J. The role of dissolved carbohydrates in the northern Adriatic macroaggregate formation. Acta Chim. Slov. 2009, 56, 305–314. [Google Scholar]
- Aluwihare, L.; Repeta, D.; Chen, R. A major biopolymeric component to dissolved organic carbon in surface sea water. Nature 1997, 387, 166–169. [Google Scholar] [CrossRef]
- Lam, B.; Baer, A.; Alaee, M.; Lefebvre, B.; Moser, A.; Williams, A.; Simpson, A.J. Major structural components in freshwater dissolved organic matter. Environ. Sci. Technol. 2007, 41, 8240–8246. [Google Scholar] [CrossRef] [PubMed]
- Kuhl, K.P.; Cave, E.R.; Abram, D.N.; Jaramillo, T.F. New insights into the electrochemical reduction of carbon dioxide on metallic copper surfaces. Energy Environ. Sci 2012, 5, 7050–7059. [Google Scholar] [CrossRef]
- Becker, J.W.; Berube, P.M.; Follett, C.L.; Waterbury, J.B.; Chisholm, S.W.; DeLong, E.F.; Repeta, D.J. Closely related phytoplankton species produce similar suites of dissolved organic matter. Front. Microbiol. 2014, 5, 1–14. [Google Scholar] [CrossRef]
- Biersmith, A.; Benner, R. Carbohydrates in phytoplankton and freshly produced dissolved organic matter. Mar. Chem. 1998, 63, 131–144. [Google Scholar] [CrossRef]
- Repeta, D.J. Chemical characterisation and cycling of dissolved organic matter. In Biogeochemistry of Marine Dissolved Organic Matter; Hansell, D.A., Carlson, C.A., Eds.; Academic Press: London, UK, 2015; pp. 21–63. [Google Scholar]
- Mozetič, P.; France, J.; Kogovšek, T.; Talaber, I.; Malej, A. Plankton trends and community changes in a coastal sea (northern Adriatic): Bottom-up vs. top-down control on relation to environmental drivers. Estuar. Coast. Shelf Sci. 2012, 115, 138–148. [Google Scholar] [CrossRef]
Figure 1.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the nanoflagellate Teratselmis marina. Values below brackets are integrated values.
Figure 1.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the nanoflagellate Teratselmis marina. Values below brackets are integrated values.

Figure 2.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the diatom Chaetoceros socialis. Values below brackets are integrated values.
Figure 2.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the diatom Chaetoceros socialis. Values below brackets are integrated values.

Figure 3.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the dinoflagellate Prorocentrum minimum. Values below brackets are integrated values.
Figure 3.
1H NMR spectra of retentate (1) and permeate (2) isolated from exudate of the dinoflagellate Prorocentrum minimum. Values below brackets are integrated values.

Figure 4.
Comparison between 1H NMR spectra of marine COM from July 2012 (1) and COM isolated from exudate of Chaetoceros socialis (2). Values below brackets are integrated values.
Figure 4.
Comparison between 1H NMR spectra of marine COM from July 2012 (1) and COM isolated from exudate of Chaetoceros socialis (2). Values below brackets are integrated values.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Distribution of the integrated main groups of proton resonances (lipids, proteins and CRAM, polysaccharides and formate) in 1H MNR spectra (δ/ppm), concentrations of Corg. (µmol L−1) in retentates and permeates and % COC from exudates of cultured phytoplankters. Protons that resonated in certain chemical shift range of integrated main groups are in bold (second row).
Table 1.
Distribution of the integrated main groups of proton resonances (lipids, proteins and CRAM, polysaccharides and formate) in 1H MNR spectra (δ/ppm), concentrations of Corg. (µmol L−1) in retentates and permeates and % COC from exudates of cultured phytoplankters. Protons that resonated in certain chemical shift range of integrated main groups are in bold (second row).
| Lipids | Proteins and CRAM | Polysaccharides | Formate | Corg. | % COC * | |
|---|---|---|---|---|---|---|
| HCH2-CH2- | HC-HCOR | HC-OH HC-O-C | HCOO | |||
| δ **/ppm | 0–1.8 | 1.8–3.0 | 3.0–4.6 | 8.0–9.0 | µmol L−1 | % |
| Teratselmis marina | ||||||
| 0.2 µm filtrate | 915.1 | |||||
| Retentate | 19.4 | 18.8 | 61.4 | 0.4 | 364 | 39.8 |
| Permeate | 35.7 | 40.9 | 20.8 | 2.6 | 507 | |
| Chaetoceros socialis | ||||||
| 0.2 µm filtrate | 2285 | |||||
| Retentate | 14.4 | 21.9 | 62.9 | 1.1 | 526 | 23.0 |
| Permeate | 16.4 | 36.4 | 45.5 | 1.7 | 1765 | |
| Prorocentrum minimum | ||||||
| 0.2 µm filtrate | 439.3 | |||||
| Retentate | 21.8 | 45.6 | 31.9 | 0,8 | 154 | 35.1 |
| Permeate | 17.4 | 56.7 | 24.6 | 1.2 | 418 | |
Note(s): * Colloidal organic carbon percentage: %COC = Corg. (retentate) × 100%/Corg. (0.2 µm filtrate). ** Chemical shift range in 1H NMR spectra for each group of compounds.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Klun, K.; Šket, P.; Beran, A.; Falnoga, I.; Faganeli, J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water 2023, 15, 111. https://doi.org/10.3390/w15010111
AMA Style
Klun K, Šket P, Beran A, Falnoga I, Faganeli J. Composition of Colloidal Organic Matter in Phytoplankton Exudates. Water. 2023; 15(1):111. https://doi.org/10.3390/w15010111
Chicago/Turabian StyleKlun, Katja, Primož Šket, Alfred Beran, Ingrid Falnoga, and Jadran Faganeli. 2023. "Composition of Colloidal Organic Matter in Phytoplankton Exudates" Water 15, no. 1: 111. https://doi.org/10.3390/w15010111
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.