
Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters


1
Department of Ecology and Animal Biology, University of Vigo, 36310 Vigo, Spain
2
Department of Plant Biology and Soil Science, Faculty of Biology, University of Vigo, 36310 Vigo, Spain
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 115; https://doi.org/10.3390/w15010115
Submission received: 15 November 2022
/
Revised: 23 December 2022
/
Accepted: 24 December 2022
/
Published: 29 December 2022
(This article belongs to the Special Issue Freshwater and/or Brackish Diatoms: Ecology and Bioindication)
Abstract
:Despite the reported allelopathic nature of the Eucalyptus genus, eucalypt leachates have unknown effects on the trophic base of stream green food webs. Eucalypt plantations have increased worldwide, including riparian ecosystems. We aimed to test whether short-term eucalypt leachates might alter water chemistry, periphytic algal biomass and diatoms, and herbivorous invertebrates’ (i.e., scrapers). We studied two oligotrophic and well-preserved headwaters from NW Spain. The experiment followed a before-after control-impact paired (BACIp) design, with weekly sampling before and after eucalypt leaves addition to streams. In the stream with lower discharge, the eucalypt treatments seemed to affect increases in biomass accrual (Chl-a) and diatom assemblages, disfavouring sensitive species (e.g., Eunotia minor and Achnanthidium pyrenaicum). Therefore, the ecological status was reduced from high to good. In the stream with higher discharge, invertebrate assemblages changed with scrapers having their abundances modified in comparison with the control (e.g., Habrophlebia sp. and Elmidae). Results suggest that eucalypt leaves leachates exert toxic effects on periphyton and on diatom and invertebrate assemblages, and this effect might be mediated by discharge. Alterations at the base of stream food webs may compromise their good ecological status. Further studies are necessary to identify whether this toxicity is due to allelopathic processes.
1. Introduction
Stream ecosystems receive energy from two different sources, terrestrial carbon produced in the basin and imported into the stream and the carbon produced by the primary producers inhabiting the stream [1]. These two energy sources correspond to the “green” and “brown” aquatic food webs [2]. A green food web is based on energy obtained from plants or algae (i.e., by photosynthesis and subsequent consumption by herbivores); while a brown food web is based on the consumption of decomposing biomass by detritivores [2,3]. The allochthonous carbon in the form of leaves, fruits, flowers, and wood has been recognised as the main energy source to forested headwaters as their dense riparian vegetation limits light penetration and consequently instream autochthonous primary production [4,5]. Agricultural and forestry activities in basins can significantly alter the sources of carbon entering stream ecosystems and, at the same time, alter light entrance to streams, affecting carbon production by primary producers [6,7]. Moreover, changes in the native tree riparian vegetation affect the quantity and quality of organic matter inputs reaching streams that may have consequences on the functioning of these ecosystems [8,9].
Forest plantations are considered monospecific crops used to increase biomass production [10]. These plantations generally imply the introduction of tree species outside their natural distribution area, affecting the quality of the organic matter that enters streams and rivers [11]. Eucalyptus globulus Labill. is considered a very productive species by the timber industry due to its very high-growth rate and its remarkable adaptability. For this reason, the area covered by eucalypt plantations has grown enormously in recent decades throughout the world, especially in temperate and tropical climates [12,13]. Since this species arrived at the Iberian Peninsula in 1829 [14], large areas have been transformed into exotic eucalypt monocultures [15,16], generating concern among limnologists [17,18] on the ecological impacts of these large-scale changes in the landscape [19].
Eucalyptus leaves are considered of poor quality because of their lower nutrient contents compared with other riparian deciduous leaves [20,21]. It has been seen that eucalypt plantations in northern Spain located near streams might reduce nitrogen and phosphorus input to them by 67% and 65%, respectively, compared to oak forest streams [11,22], and even more if compared with alder riparian native nitrogen-fixing tree with the greatest N and P contents in leaves [21]. Eucalypt plantations at industrial scales cause high demand for fertilisers and water, and intensive management has the potential for a release of allelopathic substances to flowing waters. These plantations severely influence the soil structure, causing an increase in water consumption, soil erosion [23], loss of nutrients [14] and biodiversity [16], pests and diseases [13], and alterations in soil quality and hydrology [13,24]. Evidence of allelopathic activity and chemical composition of different species of the Eucalyptus genus has been widely reported (e.g., [25,26]. Although the flowers, fruits, bark and roots have allelochemicals in their composition, the recognised allelopathic activity of E. globulus is mainly attributed to its leaves [27,28]. Water-soluble and volatile biologically active compounds, especially essential oils rich in terpenoids, makes eucalypt a potential pesticide (e.g., [28,29], and references therein). Its effect on terrestrial ecosystems has already been studied. Phytotoxic effects have been reported on the germination and root growth of numerous wild plant species, crops, and agricultural weeds (see, e.g., [28,30,31].
The EU water legislation (WFD; 2000/60/EC [32]) aims for rivers and streams to achieve good ecological status. Therefore, it requires the assessment of anthropogenic impacts on water bodies. To meet the targets of the WFD, it is necessary to know the impact of forestry operations on stream water quality and develop methods to estimate the increase in nutrient export to watercourses [33]. Forestry activities are commonly reported to increase erosion and fine sediment deposition in stream headwaters, worsening water quality [34] and decreasing biodiversity and abundance of benthic invertebrates and fish in riparian ecosystems [35]. Diatom indices and multimetrics are used in assessing the ecological status of streams and rivers, being good indicators in flowing waters, as their fast life cycles allow them to respond quickly to nutrients [36]. In addition, diatoms are considered good indicators of herbicidal compounds toxicity [37,38]. However, despite the extensive study of the herbicidal effect of eucalypt, current information on how this toxicity can affect diatom assemblages inhabiting rivers and streams running through eucalypt plantations is still very scarce (but see [39,40]. Many studies have been conducted on the effects that eucalypt leaves have on invertebrates colonising leaves. As expected, with adverse effects on detritivore invertebrates consuming leaves [18,41] but also on fish [17] and scrapers colonising eucalypt leaf litter [42]. However, the effects that eucalypt leachates may have on the green food webs of stream ecosystems are still unknown.
The effect of eucalypt allelopathic compounds on terrestrial plants due to their herbicidal effect has been reported. Therefore, our working hypothesis is that eucalypt leachates also affect diatoms, altering the base of green food webs in headwaters and, in turn, influencing herbivores feeding on them. To answer this hypothesis, we conducted an in situ experiment that simulated a short-term pulse of eucalypt leaves recently fallen from a riparian eucalypt plantation in two well-preserved streams that had not previously been affected by eucalypt plantations. We analysed whether these short-term leachates generate changes in water chemistry, algal biomass (chlorophyll-a concentrations), the composition of diatom assemblages and, consequently, the diatom indices used to assess the ecological status of the streams. Moreover, those changes in the green base of food webs can alter the composition of benthic invertebrates by influencing the herbivores’ assemblages feeding on it.
2. Material and Methods
2.1. Studied Region
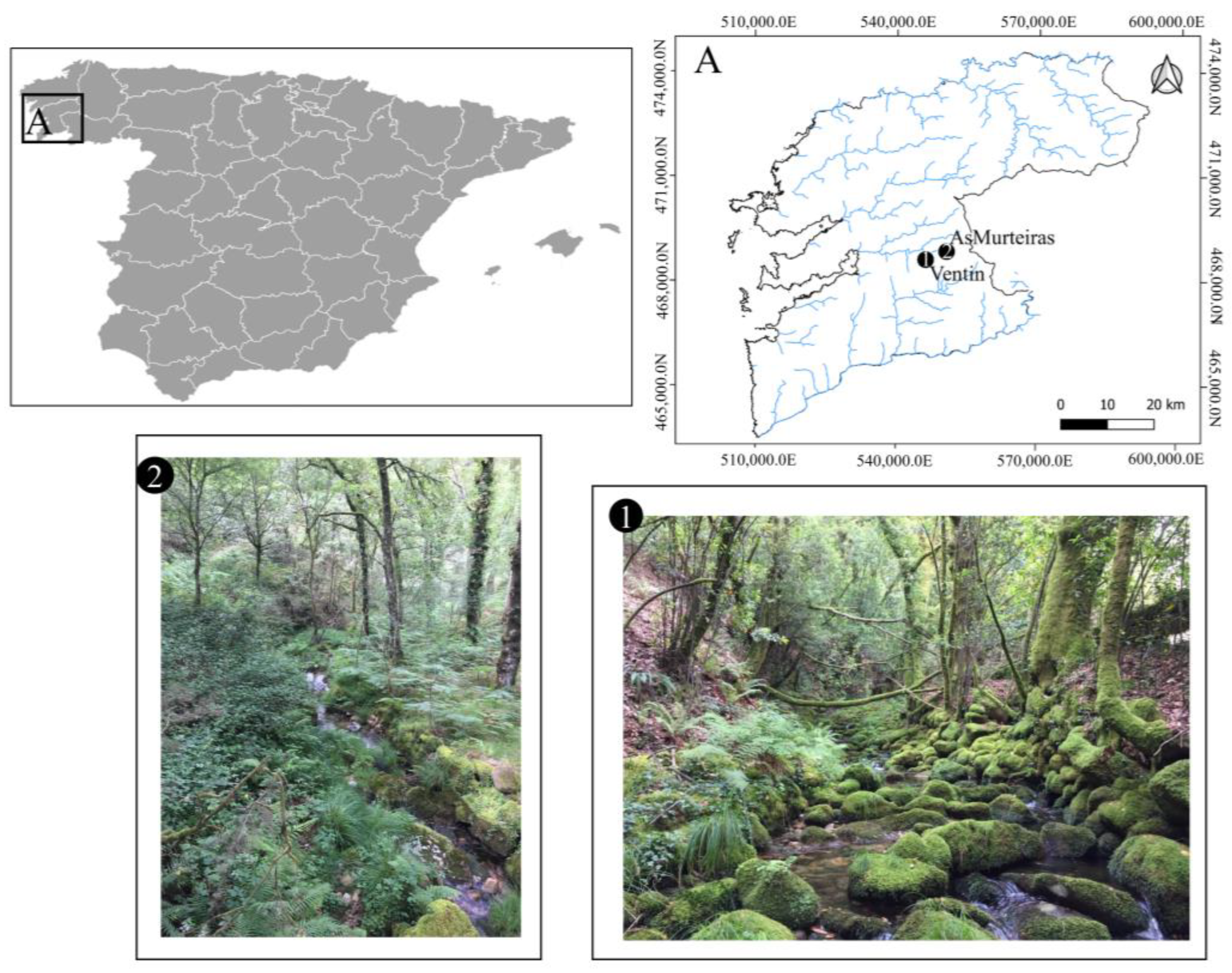
This study was conducted in two second order streams minimally impaired by human pressures located in the Pontevedra province (Galicia, NW Spain) during summer 2020: As Murteiras (42°20′43.8″ N, 8°23′6.4″ W) and Ventín streams (42°19′29″ N, 8°26′21.1″ W) (Figure 1). Both streams were selected among the headwaters of the Oitavén River, basins of Atlantic forest with streams having riparian and basin vegetation dominated by birch (Betula alba L.), oak (Quercus robur L.), accompanied in the forest floor with blueberries (Vaccinium myrtillus L.). This location in the northwest corner of the Iberian Peninsula exposes Galicia to the direct influence of the Atlantic Ocean. Therefore, the area is characterised by rainy weather (annual precipitation around 1500 mm) of relatively constant year-round precipitation frequency, with mild temperatures (around 8.5 °C in winter and 19 °C in summer). The most abundant bedrock materials are granite and mica schist [43], being the studied streams of slightly acid waters, with low salt and nutrient content [44,45].
2.2. Experimental Design
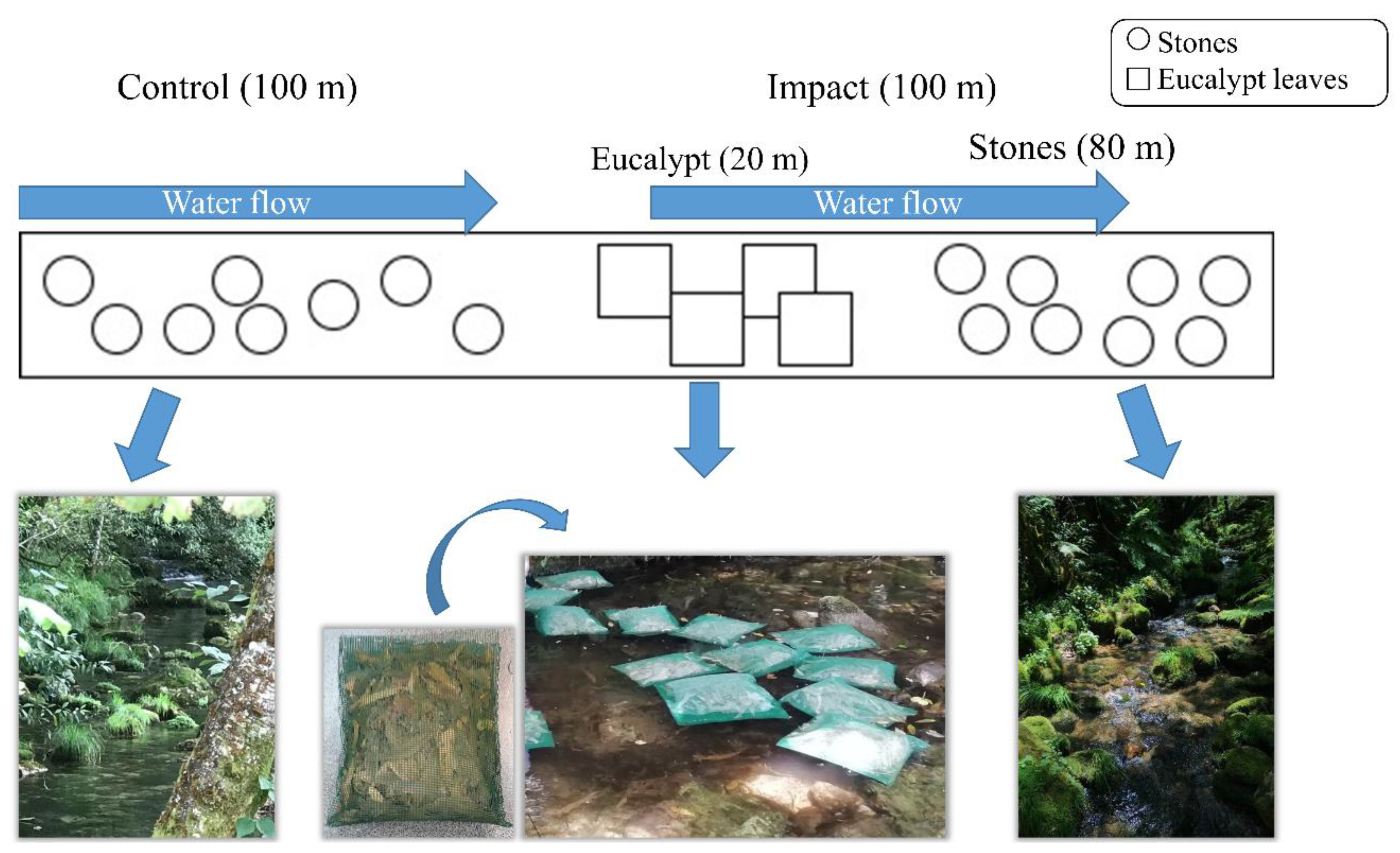
The in-stream experiment followed a Before–After, Control–Impact paired (BACIp) design (Figure 2). In each stream, a 200-m length experimental reach was selected for the study. The first 100 m upstream constituted the control reach, and the last 100 m downstream constituted the impact reach. The mean widths were 1.0 ± 0.2 m and 1.5 ± 0.3 m and the depths were 0.19 ± 0.02 m and 0.27 ± 0.04 m for As Murteiras and Ventín, respectively. In the Ventín stream, there is a small levelling used to divert water to an old mill (8 m long), not in use actually, where water retention is slightly higher than at the control reach; however, it does not prevent the continuous flow between both reaches. Therefore, the experiment consisted of the parallel sampling of both control and impact reaches, before and after the placement, at the upstream end of the impacted reaches, of bags containing known amounts of eucalypt (Eucalyptus globulus L.) leaves.
To elaborate the bags containing eucalypt leaves, freshly fallen eucalypt leaves from a plantation located 40 km from the study area were collected from the most superficial layer of the soil. In the laboratory, they were dried at room temperature and in the dark, to avoid phenol degradation [46], and then we estimated the leaves’ wet weight and Ash Free Dry Mass (AFDM). The percentage of humidity of several fractions of eucalypt leaves (i.e., 14.2–15.3%) was estimated to discount it in the estimation of dry weights to be used in each stream. The quantity of eucalypt leaves used to build the bags deposited on each stream was derived from existing values of benthic amounts of eucalypt leaves in streams affected by eucalypt forestry under low-flow conditions (267.0 g AFDM m−2; [11], multiplied by the area of each impacted reach (mean channel width of the streams throughout the experiment was 1.0 m2 at As Murteiras and 1.5 m2 at Ventín), this area being influenced by the eucalypt leachates from the bags located in the upstream end. Next, the eucalypt leaves were wrapped in mesh net bags (5 mm mesh size) of 1 kg and deposited on the stream bed within the first 20 m of the upstream end of the impacted reach of both streams (Figure 2). The impacted area comprised thus 80 m2 in both streams, which corresponded to 26 kg of eucalypt leaves wet weight in As Murteiras and to 40 kg wet weight in Ventín stream. According to the calculations based on Canhoto and Laranjeira [41], the quantity of phenols from eucalypt leachates released during 7 days was 2328.3 mg mL−1 in As Murteiras and 3582.0 mg mL−1 in the Ventín stream. Although in the case of this study, the leachates are diluted with the discharge.
The study was conducted during the summer of 2020. Following this design, the frequency of samples collection included two dates “before” (28 July and 3 August) and two other dates “after” (10 August and 17 August) the incorporation of the eucalypt bags to the impacted reaches of the studied streams. The net bags with the eucalypt leaves were placed in the impact reaches bed on 6 August 2020.
Stone colonisation. On 21 June 2020, 128 stones of similar size (upper mean area of 193 ± 24 cm2) were collected from a nearby stream and transported to the laboratory. There, they were duly cleaned with bleach and brushed, being finally marked with two white paint dots on their upper face. On 30 June, these stones were left in the streams under current and shallow depth conditions along the 200 m of each stream for the natural colonisation process. Altogether, 32 stones were placed at each control and impact reach at a regular distance of approximately 2 m. At the impact reach, stones were left regularly in the following 80 m downstream (Figure 2). We assumed that the stones should be fully colonised after one month of in-stream submersion [47] and allowed the estimation of benthic chlorophyll-a concentration and of diatom and invertebrate assemblages.
2.3. Field and Laboratory Work
On each sampling date, eight colonised stones were randomly collected per reach and stream (n = 32; Figure 2). These eight stones were first used for the collection of invertebrates; then, six of these eight stones were used for the measurement of chlorophyll-a, and two of them for the sampling of benthic diatoms. In addition, on the same dates, 3–5 natural stones from the streams beds were collected to obtain diatom composition.
In parallel, several physical-chemical variables were measured in situ in the stream with portable probes, and water samples were taken and transported for further analyses to the laboratory.
Physical-chemical variables. Onsite measurements were done at control and impact reaches on every sampling date for water temperature (°C) and dissolved oxygen (mg O2 L–1) and saturation (%) (Hach HQ30d Portable Dissolved Oxygen Meter), electric conductivity (µS cm–1) (Orion 3-Star, ThermoScientific, Waltham, MA, USA), and pH (Hach SensION+ PH1 Portable). Water speed was measured three times in a fixed transect in each stream (at the downstream end of the impact reach) with a portable current meter (Flow probe, model FP101; Global water instrumentation, Gold River, California), and the discharge value per stream was derived by multiplying the wetted area by mean velocity on each sampling date.
Two water samples were collected per reach and stream on each sampling date, kept on ice in the field and then frozen in the laboratory until analyses (n = 32). In the laboratory, water chemical analyses followed the American Public Health Association methods [48]. Phosphate [P-PO4−3 µg L−1], nitrate [N-NO3− µg L−1], nitrite [N-NO2− µg L−1] and ammonium [N-NH4+ µg L−1] were measured with an Auto-Analyzer 3 (Bran + Luebbe, Germany) and analysed in the CACTI (Centro de Apoio Ciéntifico e Tecnolóxico á Investigación, University of Vigo, Spain). Luminosity (lum ft−2) levels and water temperature (°C) were measured at 15 min intervals with a data logger (HOBO onset) placed in each reach and stream.
Chlorophyll-a. To assess algal accrual biomass between dates, chlorophyll-a (Chl-a) was measured on each sampling date from six stone replicates collected per reach and date. In total, 80 Chl-a samples were collected from the two streams. Samples were taken by brushing 32 cm2 from the top surface of each stone with a toothbrush. These samples were washed with distilled water and frozen until analyses. Then, Chl-a samples were filtered through Whatman GF/C glass fibre-filters. Chl-a was extracted in acetone (90%) in the dark at 4 °C for 48 h and was measured spectrophotometrically (Hitachi Model U-2001 UV/Visible Spectrophotometer). Values were determined using Lorenzen [49] equations.
Benthic diatom and invertebrate composition. Two stone replicates per reach and date were collected to assess the composition of benthic diatom assemblages at every sampling date (n = 32). Samples were taken by brushing 32 cm2 from the top surface of each stone with a toothbrush. Moreover, to assess the ecological status of the stream using diatoms, we used the standardized collection protocol [50] by taking 3 to 5 stones collection per sampling date from each reach in each stream. The upper surface of those 3–5 stones was brushed with a toothbrush, then rinsed with stream water in a container for one combined sample per time and site (following the European standards [50,51,52]). Therefore, the 3–5 stones collected per reach and stream on each sampling date constituted a single sample; thus n = 16. All the samples (32 + 16) were preserved with formaldehyde solution (37%) immediately after collection. In the laboratory, an aliquot of 1–3 mL from each sample was treated with a 65% solution of nitric acid (HNO3) and potassium dichromate (K2Cr2O7) at room temperature for 24–48 h to remove the organic content. The acid residues were removed by centrifugation (1500 rpm) and then rinsed with distilled water. Permanent slides were mounted in Naphrax®. Four hundred diatom valves per sample were identified to the lowest feasible taxonomic level under the light microscope (Olympus BX40). Taxonomic identification was based on [53,54,55,56,57,58,59,60,61,62,63].
Benthic invertebrates from previously placed stones were collected on each sampling date using a hand net (mesh size 100 µm) by following the sampling collection technique reported in Death & Winterbourn [64] and Matthaei et al. [65]. During the sample collection, we moved upstream to avoid disturbing the nearby stones and the stream bed itself (the hand net operator collected stones from the stream side or placing the feet on stones). The net was held downstream of the stone, and the stone was quickly introduced into the net. Eight replicates were taken per reach before and after the eucalypt addition in bags. Altogether, 128 benthic invertebrate samples were collected from the two studied streams. Samples were kept under cold and dark conditions, and once in the laboratory, they were identified under a stereomicroscope (Nikon SMZ645) and preserved in alcohol (70%). Identification was at the lowest possible taxonomic level, mainly genus based on Tachet et al. [66].
2.4. Statistical Analyses
Physico-chemical variables. The environmental matrix was constituted of 16 samples (means of two replicates) and eight environmental variables: electric conductivity (µS cm–1), pH, dissolved oxygen (mg O2 L–1), oxygen saturation (%), discharge (m3 s−1), water temperature (°C), N-DIN (µg L−1; summation of N-NH4+ µg L−1, N-NO2− µg L−1 and N-NO3− µg L−1) and P-PO43− µg L−1. Wilcoxon signed-rank test was performed to test for changes in the environmental variables measured before and after and between control and impact reaches.
Diatom and invertebrate composition. To assess changes in diatom and invertebrate composition between streams (As Murteiras and Ventín), reaches (control-impact) and periods (before-after), we performed multivariate techniques. Benthic diatom relative abundances (n = 32) and invertebrate abundances (n = 128) were log (x + 1) transformed. To assess diatom and invertebrate assemblages’ composition non-Metric Multidimensional Scaling (nMDS) ordination was performed. A permutation analysis of variance (PERMANOVA [67] was used on Bray–Curtis similarity matrix with 9999 permutations to test whether diatom and invertebrate assemblages varied across streams, periods and reaches. In addition, to determine the contribution of individual taxa to the overall differences between streams, dates, periods and reaches a SIMilarity PERcentage analysis (SIMPER; 90% cut-off) was used. To identify the set of environmental variables that best explained diatom distribution among samples, a Distance-based linear model (DistLM) [68,69] in combination with distance-based redundancy analysis (dbRDA; [70] were used. Environmental variables were fourth-root transformed, whereas diatom abundances were log (x + 1) transformed before Bray-Curtis similarity matrices were calculated. The best-fit model, based on Akaike’s Corrected Information Criterion (AICc; [71] was selected and visualised using dbRDA ordination. All multivariate analyses were conducted in PRIMER7 v.7 software [72] with the PERMANOVA+ add-on package [70].
Diatom indices were calculated to explore their response to the experimental short-term eucalypt pulse exposure. The Specific Polluosensitivity Index (IPS, [73]) and the Shannon–Wiener Diversity Index (H) were calculated using the Omnidia software v. 5.3 [74]. The MDIAT multimetric was calculated with Excel spreadsheets by averaging six metrics previously calculated with the Omnidia software: IDG, IPS, L&M, SHE, SLAD, and TDI indices with the percentage of the richness of reference taxa (PFSS) and the percentage of the abundance of sensitive taxa (PABSS) [75]. Finally, Ecological Quality Ratios (EQR), defined as the “observed” value of a candidate metric divided by that value “expected” at reference condition [76](Kelly et al., 2012), were calculated for the IPS and MDIAT of each sample. Both streams belong to the river type R-T21 Siliceous Cantabrian-Atlantic rivers, with high and good ecological status boundaries for the IPS being: High (H) > 0.920; Good (G) = 0.920 − 0.690; and for the MDIAT being: High (H) > 0.930; Good (G) = 0.930 − 0.700. We used the Wilcoxon signed-rank test to test for changes between reaches (control and impact) and periods (before and after) in index values. The level of significance for the analysis was set as p < 0.05.
Finally, the effects of eucalypt leachates over time on Chl-a concentrations and percentage of scrapers were tested per each stream and reach using Wilcoxon signed-rank test. All statistical analyses were considered at a significance level of 0.05.
3. Results
3.1. Physico-Chemical Variables
The stream water chemistry was similar in the two streams, characterised by low electric conductivities (mean range 18.1–21.9 µs cm−1), slightly acidic pH (mean range 6.4–6.7), and very low nutrient contents (mean range 5.3–12.9 µg P-PO43− L−1; Table 1), although most variables measured showed lower values in As Murteiras stream. In As Murteiras, conductivity decreased over time, and dissolved oxygen and oxygen saturation showed slightly higher values at the impact reach before the eucalypt addition (p < 0.05; Table 1). Water temperature showed minimal although significant variations between periods, being slightly higher in the ‘before’ period. Luminosity was low throughout the experiment. Because of the failure of one logger, there was no data for As Murteiras control reach. However, values were similar at the impacted reaches of both streams in the ‘before’ period. In Ventín, the luminosity was higher at the impacted reach and in the ‘before’ period (p < 0.05; Table 1). In the Ventín stream, higher N-DIN at the impact reach in the ‘before’ period significantly decreased after eucalypt addition (p < 0.05; Table 1), and P-PO43- concentrations were higher in the period ‘after’ (p < 0.05; Table 1). Discharge was four times higher in Ventín than in As Murteiras throughout the experiment (Table 1). During the ‘before’ period precipitation was absent, however, uninterrupted precipitation events took place during the two weeks after eucalypt addition (precipitation 5.6 ± 3.2 mm, data provided by the closest pluviometric station of the Spanish Meteorological Agency, AEMET), slightly increasing discharge at both streams (Table 1).
3.2. Chlorophyll a
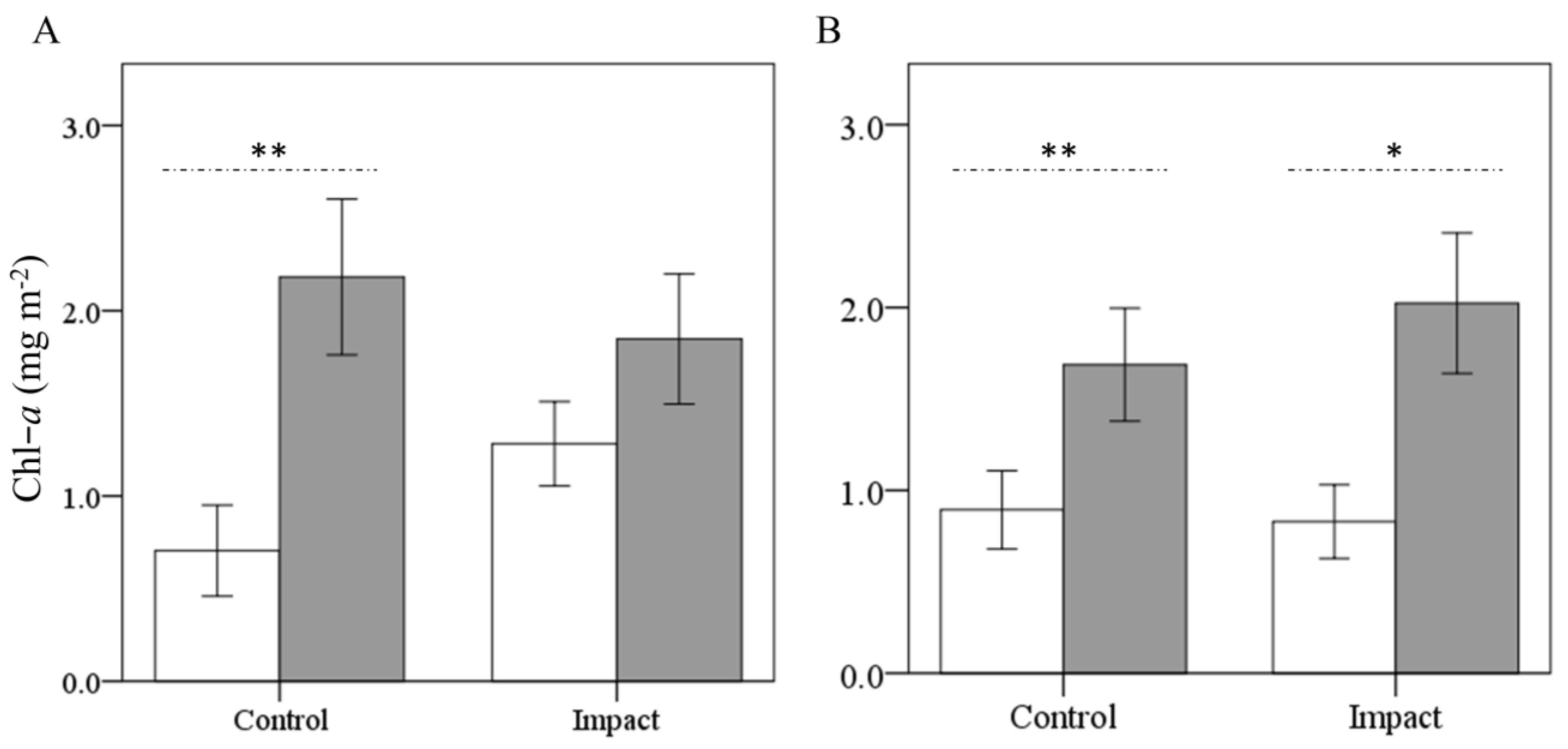
In the before period, Chl-a concentrations were initially similar in both streams (Figure 3). In As Murteiras, mean Chl-a levels significantly tripled their value at the control reach in the ‘after’ period (0.7 to 2.2 mg m−2; Wilcoxon test, p = 0.01), while at the impact reach the increase was low and non-significant (1.3 to 1.8 mg m−2; p = 0.14) despite having higher initial concentrations (Figure 3A). However, in Ventín Chl-a significantly increased at control (0.9 to 1.7 mg m−2; p = 0.01) and impact reach (0.8 to 2.0 mg m−2; p = 0.04) (Figure 3B). In Ventín, significant differences were already found in Chl-a levels between the second and third week at the control reach (Wilcoxon test, p = 0.04).
3.3. Diatom Assemblages
3.3.1. Diatom Composition
A total of 105 diatom taxa were identified in both streams in this study. The most frequent species in As Murteiras stream were: Eunotia intermedia, Psammothidium daonense, Platessa oblongella and Achnanthidium minutissimum; whereas in Ventín stream were P. daonense, E. intermedia, Surirella roba and Humidophila schmassmannii. The within-group average similarities were 77.3% and 67.6% for As Murteiras and Ventín stone samples, respectively. Significant differences were found between the diatom assemblages of both streams (PERMANOVA, p < 0.05). SIMPER analysis showed that ~90% of the cumulative dissimilarity between both streams was owed to 26 taxa, the average dissimilarity between streams being 44.0%.
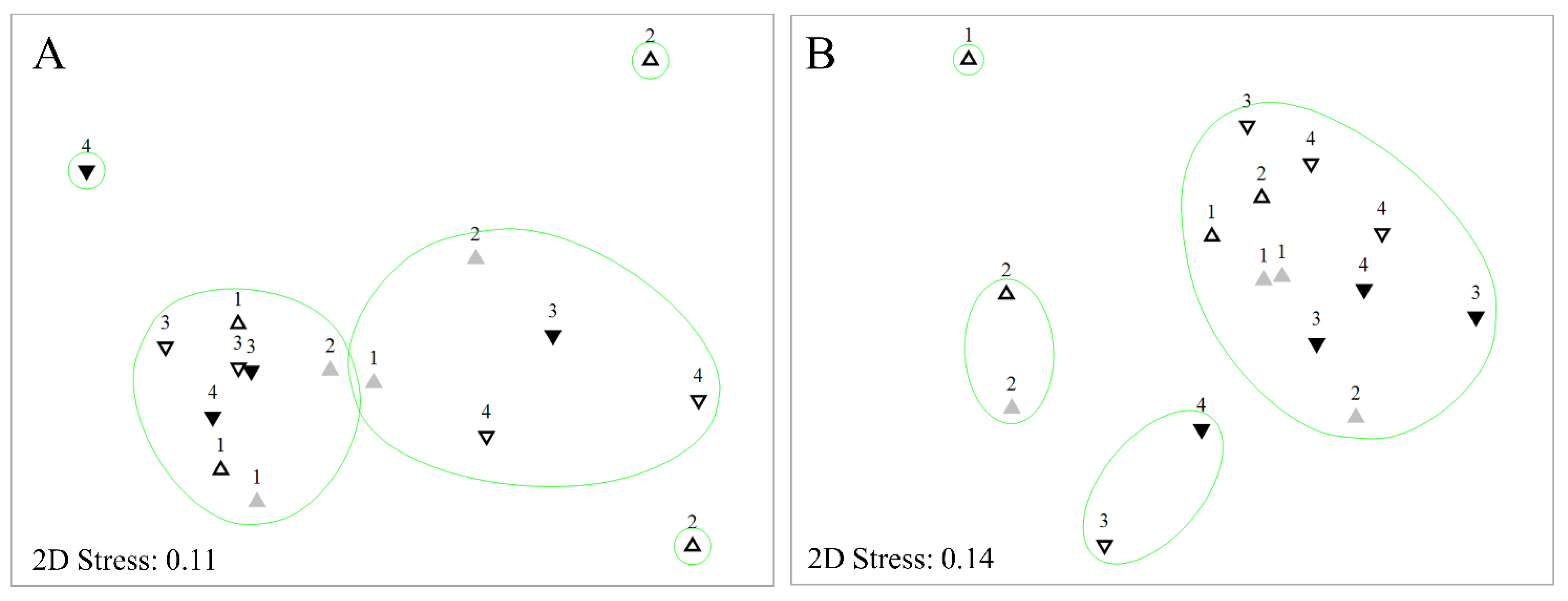
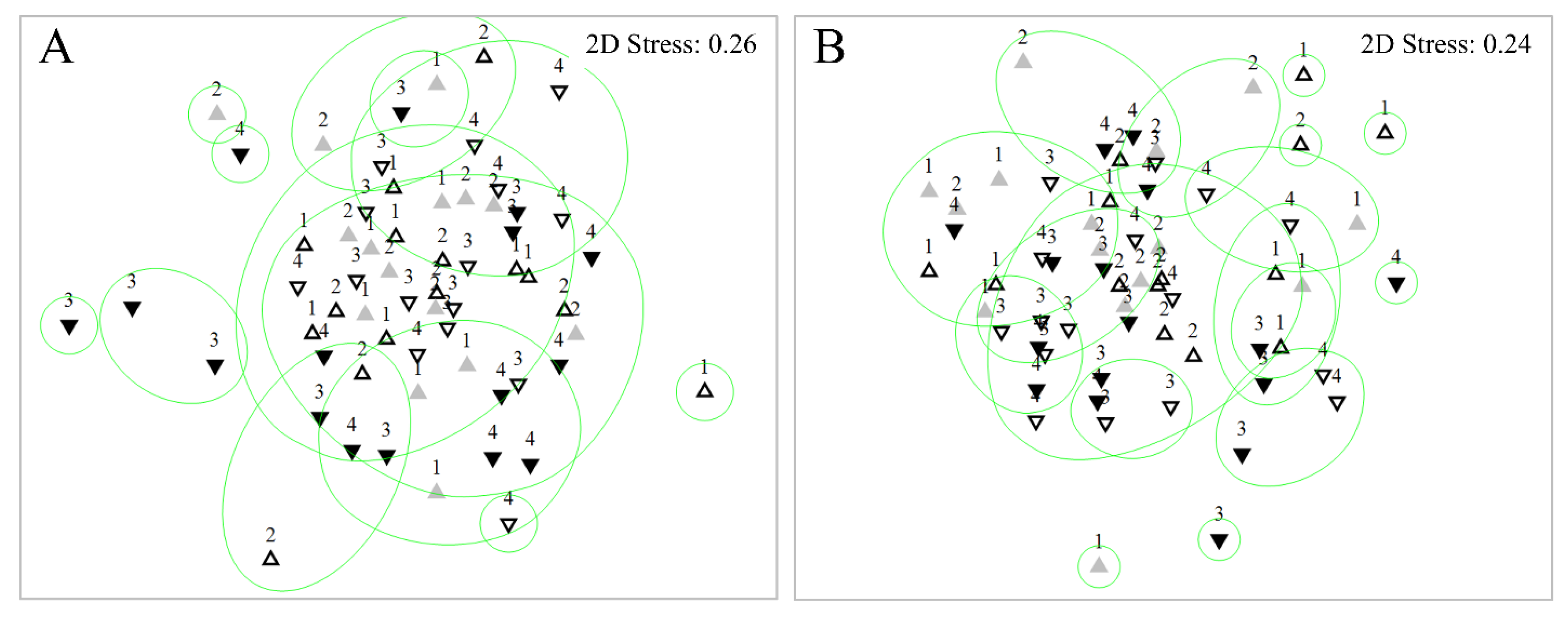
Most of the samples exposed to eucalypt from As Murteiras were plotted towards the left end of the X axis in the nMDS (Figure 4A). This was statistically confirmed by PERMANOVA when analysing the interaction Reach x Period (Pseudo-F = 2.173, p = 0.03). Similarly, those samples affected by the eucalypt in Ventín stream were plotted together in the nMDS (Figure 4B). However, in Ventín, PERMANOVA did not result in significant differences between factors (Reach x period) (Pseudo-F = 1.884, p > 0.05). No significant differences were found among dates in any of the streams (PERMANOVA p > 0.05). SIMPER average dissimilarities were higher at the control reach over time in both streams (Tables S1 and S2). The species decreasing at As Murteiras impact reach but increasing or remaining stable at the control one were: Achnanthidium pyrenaicum, Eunotia minor, Cocconeis euglypta, Gomphonema gracile and Navicula angusta (Table S1). In Ventín stream, Platessa oblongella decreased from before to after at control reach, whereas its abundance increased from before to after at the impact one (Table S2). The DistLM analysis indicated that the best-fit model explaining diatom distribution in As Murteiras was composed by water temperature, electric conductivity and P-PO43− (multiple R2 = 0.29; Figure S1A). The best-fit model explaining diatom distribution in Ventín was composed by N-DIN and P-PO43− (multiple R2 = 0.22; Figure S1B).
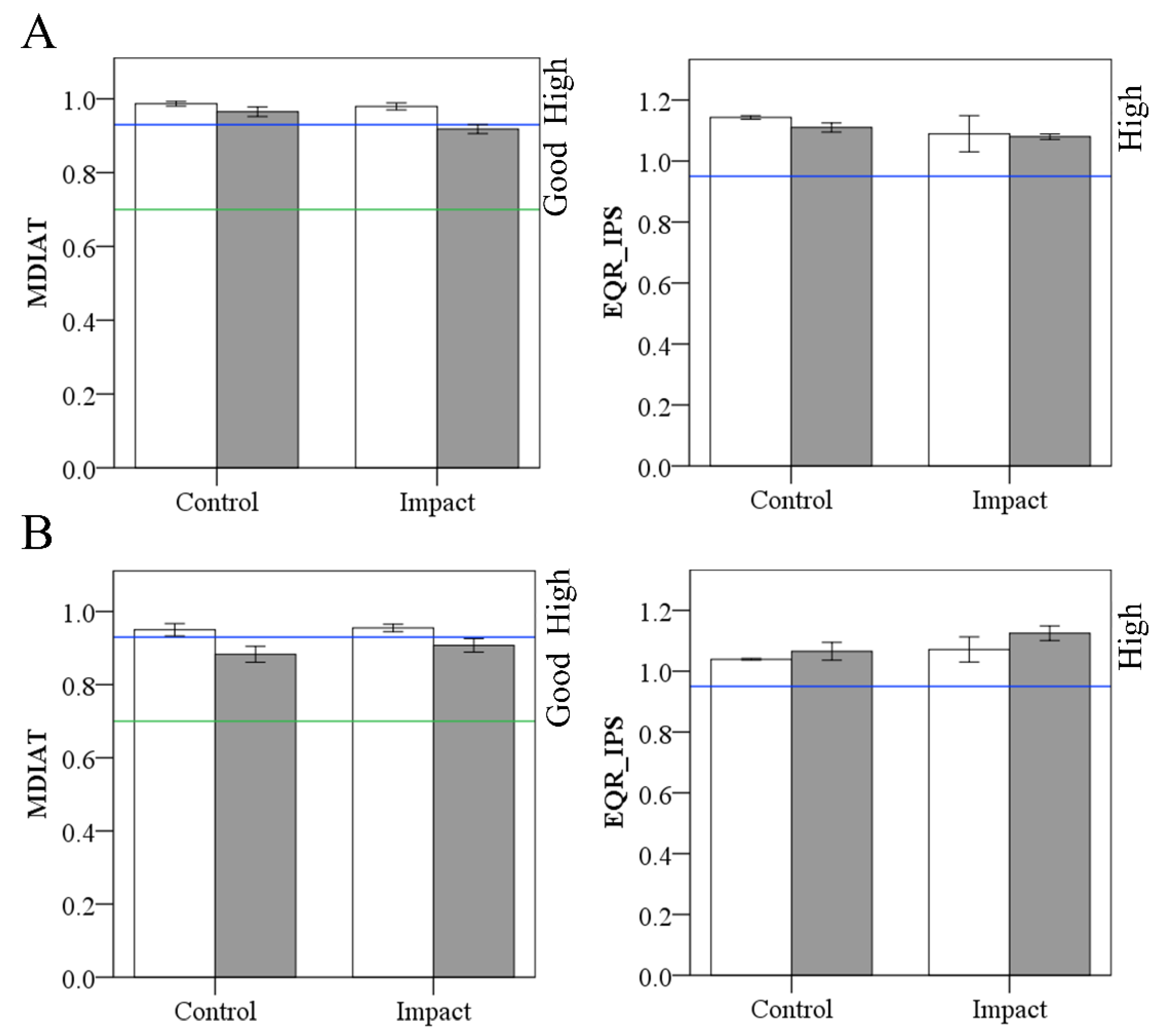
3.3.2. Ecological Status
Diatom indices did not differ between streams, reaches and periods (p > 0.05). In As Murteiras, in the control (before-after) and impact (before) reaches, diatom samples results reached the high ecological status with the multimetric MDIAT. However, samples affected by the eucalypt treatment in the impact reach during the after period were classified as in good status due to the reduction in the abundance of species such as Eunotia minor or Navicula angusta (Figure 5). In Ventín, the MDIAT classified the before samples from control and impact reaches in high status, and those from the “after” period in good ecological status, regardless of the reach (Figure 5). The IPS classified all the samples in high ecological status (Figure 5). The average Shannon–Wiener Diversity index values increased over time at both streams and reaches (Table S3).
3.4. Invertebrate Assemblages
3.4.1. Invertebrate Composition
A total of 51 invertebrate taxa colonised the 128 stones placed in both streams. The most abundant taxa in As Murteiras stream were Habrophlebia sp., Leuctra sp., and Baetis sp.; whereas in Ventín stream were Habrophlebia sp., Leuctra sp., and Hydracarina sp. As for diatoms, invertebrates’ composition was significantly different between streams (PERMANOVA, Pseudo-F = 4.780, p < 0.001). SIMPER ~90% of cumulative dissimilarity was due to 23 taxa, the average dissimilarity between streams being 70.9%. The taxa differing most between streams were Habrophlebia sp. (13.8%), Leuctra sp. (9.0%), and Baetis sp. (6.7%), as they were more abundant in As Murteiras. The within-group average samples similarities were 34.5% and 28.4% for As Murteiras and Ventín streams, respectively.
Those samples from As Murteiras subjected to eucalypt exposure were plotted towards the lower end of the Y axis in the nMDS (Figure 6A). However, no significant differences were found when analysing the interaction Reach x Period (PERMANOVA, Pseudo-F = 0.772, p > 0.05). The PERMANOVA analysis of stones sampled in Ventín stream demonstrated significant differences in the invertebrate composition when analysing the interaction Reach x Period (Pseudo-F = 2.432, p = 0.01). Nevertheless, samples were homogeneously plotted in the nMDS (Figure 6B). No significant differences were found among dates in any of the streams (PERMANOVA p > 0.05). SIMPER average dissimilarities were higher at the impact than at the control reach over time in both streams (Tables S4 and S5). In As Murteiras impact reach, the abundance of the scrapers Habrophlebia sp. Athripsodes sp. Thremma sp. and Hydraenidae decreased, whereas at control reach, it remained stable or decreased slightly (Table S4). Although Habrophlebia sp., Elmidae and Orthocladiinae abundances increased at Ventín impact reach, this increase was more significant at the control. Hydracarina sp. abundance increased at Ventín impact, whereas at control, it remained stable (Table S5).
3.4.2. Scraper’s Abundance
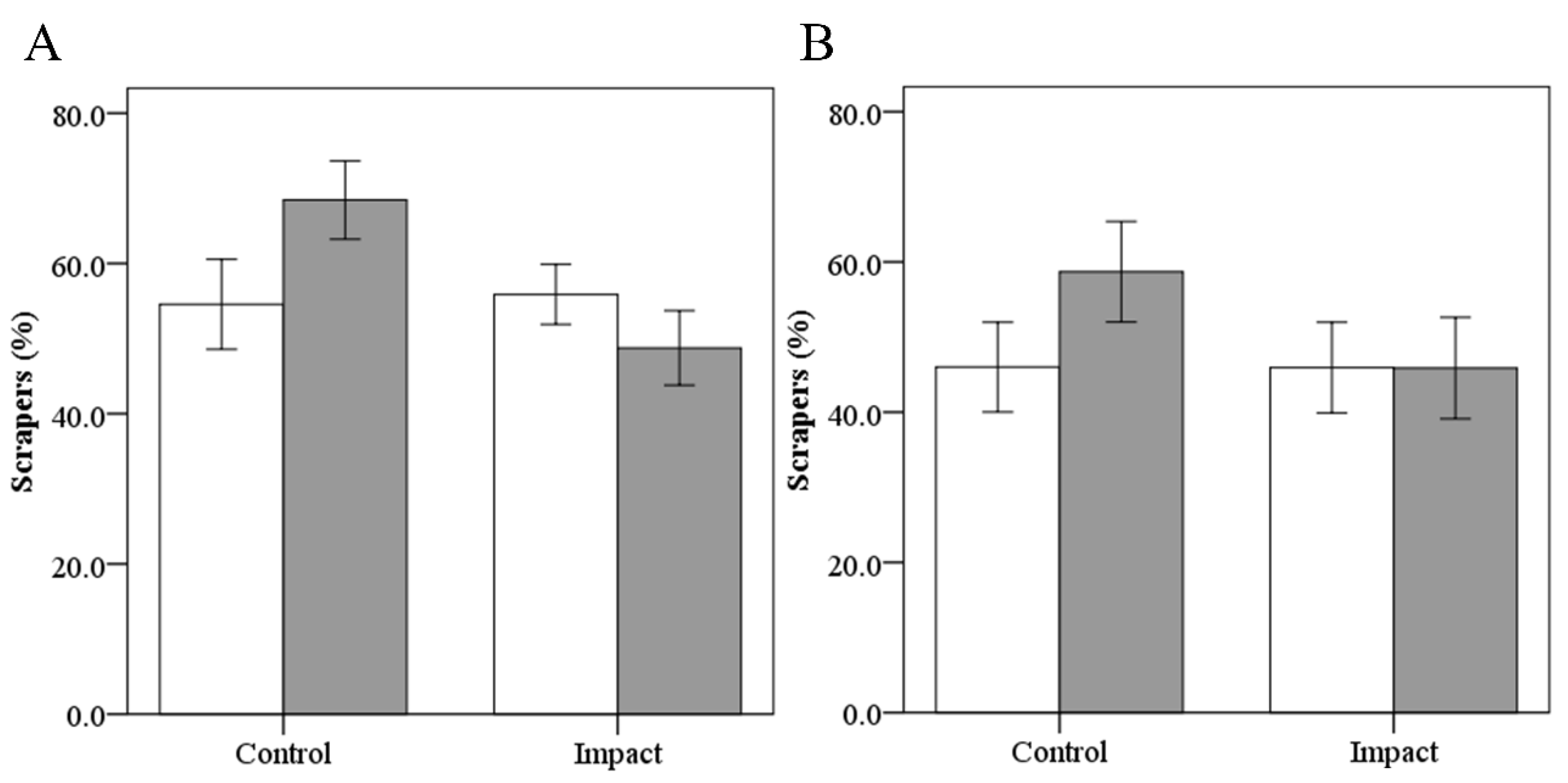
Invertebrate taxa were classified according to their functional feeding group: shredders, collectors (gatherer and filterers), scrapers and predators. The scrapers were the most abundant group from stones sampled in both reaches of both streams throughout the experiment (total abundance > 50%). Scrapers’ mean percentage in samples increased over time at control reach of As Murteiras (mean 54.6% to 68.4%; Wilcoxon test, p = 0.08) and Ventín (46.0% to 58.7%; p = 0.08) (Figure 7). However, this percentage decreased at impact reach of As Murteiras (55.9% to 48.8%; p = 0.78) and it remained stable in Ventín (45.9% to 45.9%; p = 0.23) (Figure 7).
4. Discussion
This study constitutes the first experimental in situ approximation to the effects that eucalypt leachates per se exert on well-preserved headwaters and on their aquatic benthic communities. The study results partially support our hypothesis that green food webs of headwaters could be affected by eucalypt leachates by their influence on the green base. In particular, we found statistically significant changes in diatom (in As Murteiras) and invertebrate (in Ventín) assemblages when exposed to eucalypt leachates. In As Murteiras, this was related to a decrease in the ecological status and a much smaller increase in biomass accrual (Chl-a) at the impacted reach. Invertebrate scrapers seemed to be slightly reduced in representation, even though not significantly, in the after period at the impact reaches of both streams, despite their higher levels of Chl-a. We attribute a lesser effect of eucalypt in Ventín stream because its greater discharge may have caused higher dilution of the allelopathic compounds potentially present in the eucalypt leachates.
4.1. Eucalypt Influence on Physical-Chemical Variables
Eucalypt addition seemed to influence higher conductivity levels at the impacted reaches of both streams, as it has been reported for streams crossing eucalypt plantations in the Iberian Peninsula [77]. Nevertheless, stream conductivities were low throughout the experiment and were characteristic of the siliceous geology of this region [36,44]. There were no changes in pH during this experiment, and oxygen saturation did change with time, even though values were always close to saturation. Our results differ from Canhoto and Laranjeira [41], who reported a drop in water oxygen and pH after eucalypt leaves addition under summer discharge conditions, not comparable to the continuous flowing conditions of this summer study. Nutrient contents before adding the eucalypt were low in both studied streams, having P-PO43− values typical of oligotrophic streams (P-PO43− < 12.5 µg L−1; [78]; however, higher contents in the Ventín stream after period at both control and impact reaches corresponded to mesotrophic streams (P-PO43− 12.5–37.5 µg L−1; [78]. We expected that eucalypt leachates would increase dissolved nutrient contents at impact reaches. Nonetheless, the general P-PO43− increase and N-DIN decrease observed after the eucalypt addition at both reaches and streams seem to follow the controls’ natural tendency, even though changes were more evident in the after period, with the summer more advanced, in the slightly nutrient richer Ventín. The in-stream nutrient uptake has been positively correlated to hydrological retention time [79]; thus overall N-DIN reductions in both streams and reaches can be partly attributed to autotrophs uptake during the summer months [80,81] when their active growing season occurs, as suggested by the negative relationship between Chl-a concentration and N-DIN contents at Ventín impact reach. However, general increases in P-PO43− observed in the streams at control and impact reaches in the ‘after’ period were observed, maybe related to factors other than biota, such as sorption-desorption relationship between stream water and sediments [82].
4.2. Eucalypt Effect on Biomass Accrual (Chl-a)
In forested headwater streams, seasonal changes in light, organic matter inputs, and temperature affect biotic demand for nutrients [83]. Chl-a concentrations were low during the study compared with other Galician second-order streams during summer [84], despite their correspondence with maximum annual values during spring-summer months, according to these authors. Chl-a concentrations increased over time at both streams despite lower luminosity levels observed at Murteiras in the ‘after’ period, being much more pronounced at control than at impact reaches. A higher herbivory pressure (in scrapers abundance) could explain the lower increase in Chl-a concentration at impact vs. control reaches. However, scrapers percentages at both streams’ impact reaches did not increase over time in response to higher autochthonous biomass, but they only slightly increased at control reaches in both streams. Therefore, the observed tendency of lower Chl-a increases at impacted vs. control reaches in the ‘after’ period, could be attributed to the toxicity of eucalypt leachates on stone biofilms. Eucalypt allelopathic compounds are considered to have a strong phytotoxic effect [26] in terrestrial systems, which can have toxic impacts on aquatic phototrophic organisms too. As indicated by Zhou and Yu [85], allelochemical photoinhibition of photosynthesis is typically characterised as a reduction in Chl-a fluorescence, which makes the plant unable to emit the excess energy received from the sun. Thus, this excess energy is transferred to the oxygen through chlorophyll, generating photo-oxidation damage. Excessive damage would lead to membrane destruction and chlorophyll oxidation (see [28]). Eucalypt leaf extracts reduce chlorophyll content in crops, related to possible poor photosynthesis and, as a result, poor plant growth [86]. Some allelopathic compounds from eucalypt have been used as inhibitors of photosynthetic activity in the control of algal blooms [87] and strongly limit the growth of other plant species around them [88]. Exposure to herbicides for long periods and in sunlight can lead to sustained or irreversible effects in photosynthetic organisms [89].
4.3. Eucalypt Leachates Influencing Diatom Species
Changes in the abundance of the most frequent species caused the community to change significantly after exposure to eucalypt leachates. Although changes in the Ventín diatom community were not statistically significant, those samples exposed to eucalypt were plotted together in the nMDS ordination. Therefore, this suggests that allelopathic compounds increase the similarity among those samples by replacing sensitive species. In As Murteiras, several species decreased their abundance in those samples exposed to eucalypt leachates, contributing to significant differences between Reach x Period in this stream. The most remarkable: Achnanthidium pyrenaicum, Eunotia minor, Cocconeis euglypta, Gomphonema gracile and Navicula angusta. E. minor, G. gracile, and N. angusta belong to the Galician diatom reference community [75,90], and its sensitivity to herbicides has already been reported in different studies [91,92]. In the Ventín stream the most notable change was the increase in the abundance of Platessa oblongella at the impact reach, decreasing at the control one, this species has been considered an indicator of agricultural forested streams [93] in agreement with our results. However, fewer species change in abundances at the impact reach, may have influenced the non-significant effect of the Reach x Period interaction in Ventín. Among the most abundant diatom species in the Ventín stream was Humidophila schmassmannii, a relatively rare species that prefers weakly mineralised freshwater oligotrophic environments [94,95]. The Shannon–Wiener diversity index values were higher than in other siliceous streams [96], and increased over the summer as in other studies [97].
The results of the multimetric MDIAT applied to the natural stones collected in the streams indicated that the ecological status class was reduced at the Murteiras’ impact reach after the eucalypt addition, from high to good in only 2 weeks time, evidencing a slight reduction in ecological status but always meeting the good ecological status required by the Water Framework Directive. We attribute this reduction in ecological status to the decrease in abundance of those diatom species belonging to the reference diatom community. The absence of MDIAT response in the Ventín stream might be due to the presence and dominance of Humidophila schmassmannii in this stream (species not contemplated in MDIAT reference community; [75], causing all posterior samples to be classified in good ecological status, and questioning the use of the MDIAT in Ventín samples. The IPS did not change with eucalypt additions, in agreement with its lack of response to disturbance in siliceous rivers [98,99].
4.4. Eucalypt Effect on Benthic Invertebrates
Invertebrate assemblages significantly changed their composition at the impact reach of Ventín stream, and although differences in As Murteiras were not significant, samples exposed to eucalypt leachates were plotted together in the nMDS ordination. Similarly, studies conducted in a Galician stream also identified changes in invertebrate detritivores’ composition feeding on submersed leaves between the native deciduous tree leaves and eucalypt leaves [100]. All the taxa decreasing at Murteiras impact reach but increasing or remaining stable at control one (i.e., Habrophlebia sp. Athripsodes sp. Thremma sp., and Hydraenidae) were scrapers, while Hydracarina sp. (predator) increased at Ventín impact reach. This result agrees with another study showing higher abundance of Hydracarina sp. on eucalypt leaves rather than leaves from native tree species [101]. Although Habrophlebia sp., Elmidae and Orthocladiinae increased slightly at Ventín impact reach, their increase was much greater at the control.
Stream detrital resources from eucalypt influence consumers via feeding [102]. Eucalypt leaves consumption can cause lower growth rates in some invertebrates, which cannot reach pupation, and die within 100 days [41]. Others suffer changes in larvae body stoichiometry and a delay in their development [103]. In this study, green food webs based on periphyton might have their basal resource (algae biomass and composition) affected by toxic eucalypt leachates, and this can have an effect, to be discerned, on consumers [104] that make up the green food chains of these stream ecosystems. We expected a negative effect of eucalypt on algae, which will propagate to herbivores scrapers that consume this resource [105]. Either direct adverse toxic effects affecting scrapers’ abundance or indirect effects caused by the reduction in algae biomass available may lead to higher scrapers’ competition for resources and potentially cause a reduction in scrapers’ abundance [106]. Our results showed a natural tendency at the stream controls, where increases in benthic Chl-a between the studied periods were followed by a slight increase in scrapers’ abundance. In As Murteiras, although the initial abundance of scrapers was similar at both reaches, it increased at the control and decreased at the impact, although not significantly. This fact might be related to the sharp increase experienced by the Chl-a concentrations at As Murteiras control reach. In Ventín, scrapers abundance was low and very similar over time at the impact reach, where higher Chl-a values seemed influenced by N-DIN uptake, as the relationships indicate [107]. Although neither change in scrapers’ abundance was statistically significant, changes in both river controls were close to significance.
We consider this experiment a good first approximation to the effect of eucalypt leachates on the green food web in aquatic ecosystems. The weak significance of some of the results should be attributed to the short duration of the experiment. However, the duration was designed based on the results of previous studies that suggested that eucalypt leaves were degraded by 50% after two weeks of being introduced into the stream [41]. Changes in diatom biofilms in response to this toxic substance were quick, as expected. Diatoms better showed changes in composition when exposed to eucalypt leachates because, due to their short life cycles, they respond more rapidly to environmental and anthropogenic disturbances than multicellular organisms [108]. However, invertebrates have a longer response time to pressure [109]. Thus, we consider that it could have taken more than two weeks after adding the eucalypt for the eucalypt leachates to cause changes in the entire community structure able to colonise of the impact reach, and not only in those individuals colonising the stones. Despite the short duration of the experiment, signs of the toxic effect of eucalypt leachates were detected at the basis of the food chain in these aquatic ecosystems. Future studies are necessary to extend the experiment’s duration, including the simulation of a more realistic chronic disturbance, instead of a single pulse disturbance, to test the actual magnitude of the impact of eucalypt leaves leachates on scrapers.
5. Conclusions
The present in situ simulation of eucalypt litter fall on well-preserved forested streams identified the short-term effects that eucalypt leachates can have on their water chemistry and biological communities. The composition of benthic diatoms and invertebrates was altered under eucalypt exposure by reducing sensitive taxa. Chl-a concentration experienced a lower growth on impact samples affected by eucalypt, although we cannot conclude that the reduction in Chl-a biomass significantly affected scrapers’ abundance. Alterations in diatoms and invertebrates, the first links in the food chains of stream ecosystems, suggest that eucalypt addition may result in altered biodiversity, compromising their ability to reach high and even good ecological status. This study suggests that short-term eucalypt leaf inputs to streams have per se a negative impact on the stream ecosystem. Applying best practices (e.g., respecting a wider margin free of effect, between riverside eucalypt plantations and rivers and streams) is an urgent need to ensure the maintenance and conservation of these minimally disturbed stream ecosystems in the northwest of the Iberian Peninsula. Finally, allelopathic processes potentially underlying such toxicity on green food webs in headwaters deserve to be studied.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15010115/s1, Table S1: Contribution of dominant diatom taxa from As Murteiras to the dissimilarities between periods per reach according to SIMPER analysis. Av. Abund. = average relative abundance; Contrib.% = contribution percentage of diatom taxa. Cum.% = cumulative contribution percentage. Only taxa with a contribution > 2% are represented. Diatom taxa whose changes in abundance over time contributed to differentiate between control and impact are highlighted in bold; Table S2: Contribution of dominant diatom taxa from Ventín to the dissimilarities between periods per reach according to SIMPER analysis. Av. Abund. = average relative abundance; Contrib.% = contribution percentage of diatom taxa. Cum.% = cumulative contribution percentage. Only taxa with a contribution > 2% are represented. Diatom taxa whose changes in abundance over time contributed to differentiate between control and impact are highlighted in bold; Figure S1: Distance-based redundancy analysis on diatom assemblages in (A) As Murteiras and (B) Ventín streams. Numbers representing collection date (d7, d14 before collections and d21, d28 after collections). Samples were differentiated by symbol and colour: Control before (open up-triangle), control after (open down-triangle), impact before (grey up-triangle), and impact after (black down-triangle); Table S3: Average values of the Shannon Diversity Index; Table S4: Contribution of dominant invertebrate taxa from As Murteiras to the dissimilarities between periods per reach according to SIMPER analysis. Av. Abund. = average relative abundance; Contrib.% = contribution percentage of diatom taxa. Cum.% = cumulative contribution percentage. Only taxa with a contribution > 2% are represented. Invertebrate taxa decreasing at the impact and increasing or keeping stable at control are highlighted in bold: Table S5: Contribution of dominant invertebrate taxa from Ventín to the dissimilarities between periods per reach according to SIMPER analysis. Av. Abund. = average relative abundance; Contrib.% = contribution percentage of diatom taxa. Cum.% = cumulative contribution percentage. Only taxa with a contribution > 2% are represented. Invertebrate taxa whose changes in abundance over time contributed to differentiate between control and impact are highlighted in bold.
Author Contributions
Conceptualization, I.P., C.D. and N.P.; methodology, L.G.-P., J.G. and I.P.; software, L.G.-P. and I.P.; validation, I.P., C.D. and N.P.; formal analysis, L.G.-P. and I.P.; investigation, L.G.-P. and I.P.; resources, L.G.-P. and I.P.; data curation, L.G.-P. and I.P.; writing—original draft preparation, L.G.-P., C.D. and I.P.; writing—review and editing, L.G.-P., C.D. and I.P.; visualization, L.G.-P. and I.P.; supervision, I.P.; project administration, I.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are available on request.
Acknowledgments
We are grateful to Alba Castro Guerra for her work and help during fieldwork and the realization of the experiment. Also, thanks to three anonymous referees for many useful comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hagen, E.M.; Mctammany, M.E.; Webster, J.R.; Benfield, E.F. Shifts in allochthonous input and autochthonous production in streams along an agricultural land-use gradient. Hydrobiologia 2010, 655, 61–77. [Google Scholar] [CrossRef]
- Cordone, G.; Salinas, V.; Marina, T.I.; Doyle, S.R.; Pasotti, F.; Saravia, L.A.; Morno, F.R. Green vs brown food web: Effects of habitat type on multidimensional stability proxies for a highly-resolved Antarctic food web. Food Webs 2020, 25, e00166. [Google Scholar] [CrossRef]
- Odum, E.P. The strategy of ecosystem development. Science 1969, 164, 262–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, S.G.; Likens, G.E. Energy Flow in Bear Brook, New Hampshire: An Integrative Approach to Stream Ecosystem Metabolism. Ecol. Monogr. 1973, 43, 421–439. [Google Scholar] [CrossRef]
- Rosas, K.G.; Colón-Gaud, C.; Ramírez, A. Trophic basis of production in tropical headwater streams, Puerto Rico: An assessment of the importance of allochthonous resources in fueling food webs. Hydrobiologia 2020, 847, 1961–1975. [Google Scholar] [CrossRef]
- Gao, Y.; Jia, J.; Lu, Y.; Yang, T.; Lyu, S.; Shi, K.; Zhou, F.; Yu, G. Determining dominating control mechanisms of inland water carbon cycling processes and associated gross primary productivity on regional and global scales. Earth Sci. Rev. 2021, 213, 103497. [Google Scholar] [CrossRef]
- Díaz Villa, M.V.E.; Cristiano, P.M.; De Diego, M.S.; Rodríguez, S.A.; Efron, S.T.; Bucci, S.J.; Scholz, F.; Goldstein, G. Do selective logging and pine plantations in humid subtropical forests affect aboveground primary productivity as well as carbon and nutrients transfer to soil? For. Ecol. Manag. 2022, 503, 119736. [Google Scholar] [CrossRef]
- Pozo, J.; Basaguren, A.; Elósegui, A.; Molinero, J.; Fabre, E.; Chauvet, E. Afforestation with Eucalyptus globulus and leaf litter decomposition in streams of northern Spain. Hydrobiologia 1998, 373–374, 101–110. [Google Scholar] [CrossRef]
- García, L.; Pardo, I. On the way to overcome some ecological riddles of forested headwaters. Hydrobiologia 2012, 696, 123–136. [Google Scholar] [CrossRef]
- Nambiar, E.K.S. Plantation forests: Their scope and a perspective on plantation nutrition. In Nutrition on Plantation Forests; Bowen, G.D., Nambiar, E.K.S., Eds.; Academic Press: London, UK, 1984; pp. 1–15. [Google Scholar]
- Molinero, J.; Pozo, J. Impact of a eucalyptus (Eucalyptus globulus Labill.) plantation on the nutrient content and dynamics of coarse particulate organic matter (CPOM) in a small stream. Hydrobiologia 2004, 528, 143–165. [Google Scholar] [CrossRef]
- Quartucci, F.; Schweier, J.; Jaeger, D. Environmental analysis of Eucalyptus timber production from short rotation forestry in Brazil. Int. J. For. Eng. 2015, 26, 225–239. [Google Scholar] [CrossRef]
- Zaiton, S.; Sheriza, M.R.; Ainishifaa, R.; Alfred, K.; Norfaryanti, K. Eucalyptus in Malaysia: Review on environmental impacts. J. Landsc. Ecol. 2020, 13, 79–94. [Google Scholar] [CrossRef]
- Graça, M.A.S.; Pozo, J.; Canhoto, C.; Elosegi, A. Effects of eucalyptus plantations on detritus, decomposers, and detritivores in streams. Sci. World 2002, 2, 1173–1185. [Google Scholar] [CrossRef] [Green Version]
- Boulet, A.; Prats, S.A.; Malvar, M.C.; González-Pelayo, O.; Coelho, C.O.; Ferreira, A.J.; Keizer, J.J. Surface and Subsurface Flow in Eucalyptus Plantations in North-Central Portugal. J. Hydrol. Hydromech. 2015, 63, 197. [Google Scholar] [CrossRef] [Green Version]
- Goded, S.; Ekroos, J.; Domínguez, J.; Azcárate, J.G.; Guitián, J.A.; Smith, H.G. Effects of eucalyptus plantations on avian and herb species richness and composition in North-West Spain. Glob. Ecol. Conserv. 2019, 19, e00690. [Google Scholar] [CrossRef]
- Oliveira, J.M.; Fernandes, F.; Ferreira, M.T. Effects of forest management on physical habitats and fish assemblages in Iberian eucalypt streams. For. Ecol. Manag. 2016, 363, 1–10. [Google Scholar] [CrossRef]
- Simões, S.; Gonçalves, A.L.; Canhoto, J.M.; Gonçalves, G.; Canhoto, C. Eucalyptus spp. Leaf traits determine litter processing by fungi and invertebrates. Freshw. Biol. 2021, 66, 968–977. [Google Scholar] [CrossRef]
- Bañuelos, R.; Larranaga, S.; Elosegi, A.; Pozo, J. Effects of Eucalyptus Plantations on CPOM Dynamics in Headwater Streams: A Manipulative Approach. Archiv Für Hydrobiologie 2004, 159, 211–228. [Google Scholar] [CrossRef]
- Pozo, J. Leaf litter processing of alder and eucalyptus in the Agüera stream system (North Spain) I. Chemical changes. Arch. Hydrobiol. 1993, 127, 299–317. [Google Scholar] [CrossRef]
- López, E.; Pardo, I.; Felpeto, N. Seasonal differences in green leaf breakdown and nutrient content of deciduous and evergreen tree species and grass in a granitic headwater stream. Hydrobiologia 2001, 464, 51–61. [Google Scholar] [CrossRef]
- Pozo, J.; González, E.; Díez, J.R.; Molinero, J.; Elósegui, A. Inputs of particulate organic matter to streams with different riparian vegetation. J. N. Am. Benthol. Soc. 1997, 16, 602–611. [Google Scholar] [CrossRef]
- Reichert, J.M.; de Deus Junior, J.C.; Borges Junior, N.; Cavalcante, R.B.L. Experimental catchments in the pampa biome: Database on hydrology in grasslands and eucalyptus plantations in subtropical Brazil. Hydrol. Proc. 2021, 35, e14285. [Google Scholar] [CrossRef]
- Martello, F.; de Bello, F.; de Castro Morini, M.S.; Silva, R.R.; Souza-Campana, D.R.D.; Ribeiro, M.C.; Carmona, C.P. Homogenization and impoverishment of taxonomic and functional diversity of ants in eucalyptus plantations. Sci. Rep. 2018, 8, 3266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmigid, H.M.; Morsi, M.M. Cytotoxic and Molecular Impacts of Allelopathic Effects of Leaf Residues of Eucalyptus globulus on Soybean (Glycine max). J. Genet. Eng. Biotechnol. 2017, 15, 297–302. [Google Scholar] [CrossRef]
- Nega, F.; Gudeta, T.B. Allelopathic effect of Eucalyptus globulus Labill. On seed germination and seedling growth of highland teff (Eragrostis tef (Zuccagni) Trotter)) and barley (Hordeum vulgare L.). J. Exp. Agric. Int. 2019, 30, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Morsi, M.M.; Abdelmigid, H.M. Allelopathic activity of ‘Eucalyptus globulus’ leaf aqueous extract on ‘Hordeum vulgare’ growth and cytogenetic behaviour. Aust. J. Crop. Sci. 2016, 11, 1551–1556. [Google Scholar] [CrossRef]
- Puig, C.G.; Reigosa, M.J.; Valentão, P.; Andrade, P.B.; Pedrol, N. Unravelling the bioherbicide potential of Eucalyptus globulus Labill: Biochemistry and effects of its aqueous extract. PloS ONE 2018, 13, e0192872. [Google Scholar] [CrossRef] [Green Version]
- Puig, C.G.; Gonçalves, R.F.; Valentao, P.; Andrade, P.B.; Reigosa, M.J.; Pedrol, N. The consistency between phytotoxic effects and the dynamics of allelochemicals release from Eucalyptus globulus leaves used as biherbicide green manure. J. Chem. Ecol. 2018, 44, 658–670. [Google Scholar] [CrossRef]
- Souto, X.C.; Bolaño, J.C.; González, L.; Reigosa, M.J. Allelopathic effects of tree species on some soil microbial populations and herbaceous plants. Biol. Plan. 2001, 44, 269–275. [Google Scholar] [CrossRef]
- Puig, C.G.; Revilla, P.; Barreal, M.E.; Reigosa, M.J.; Pedrol, N. On the suitability of Eucalyptus globulus green manure for field weed control. Crop. Protection 2019, 121, 57–65. [Google Scholar] [CrossRef]
- European Commission. Directive 2000/60/EC of the European Parliament and of the Council of 23 October 2000 Establishing a Framework for Community Action in the Field of Water Policy (Water Framework Directive). Off. J. Eur. Union 2000, 43, 1–73. Available online: https://eur-lex.europa.eu/eli/dir/2000/60/oj (accessed on 4 October 2021).
- Palviainen, M.; Finér, L.; Laurén, A.; Mattsson, T.; Högbom, L. A method to estimate the impact of clear-cutting on nutrient concentrations in boreal headwater streams. Ambio 2015, 44, 521–531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marttila, H.; Kløve, B. Dynamics of erosion and suspended sediment transport from drained peatland forestry. J. Hydrol. 2010, 388, 414–425. [Google Scholar] [CrossRef]
- Kukuła, K.; Bylak, A. Synergistic impacts of sediment generation and hydrotechnical structures related to forestry on stream fish communities. Sci. Total Environ. 2020, 737, 139751. [Google Scholar] [CrossRef]
- González-Paz, L.; Delgado, C.; Pardo, I. Understanding divergences between ecological status classification systems based on diatoms. Sci. Total Environ. 2020, 734, 139418. [Google Scholar] [CrossRef]
- Debenest, T.; Silvestre, J.; Coste, M.; Delmas, F.; Pinelli, E. Herbicide Effects on Freshwater Benthic Diatoms: Induction of Nucleus Alterations and Silica Cell Wall Abnormalities. Aquat. Toxicol. 2008, 88, 88–94. [Google Scholar] [CrossRef] [Green Version]
- Wood, R.J.; Mitrovic, S.M.; Lim, R.P.; Warne, M.S.J.; Dunlop, J.; Kefford, B.J. Benthic diatoms as indicators of herbicide toxicity in rivers–A new SPEcies at risk (SPEARherbicides) index. Ecol. Indic. 2019, 99, 203–213. [Google Scholar] [CrossRef]
- Friberg, N.; Winterbourn, M.J. Interactions between riparian leaves and algal/microbial activity in streams. Hydrobiologia 1996, 341, 51–56. [Google Scholar] [CrossRef]
- Pinheiro, C.; Oliveira, U.; Rodrigues, T.; Oliva-Teles, L.; Vieira, M.N. Assessing the ecological status of a Mediterranean river: Benthic invertebrates and diatoms as complementary bioindicators. Limnetica 2020, 39, 299–315. [Google Scholar] [CrossRef]
- Canhoto, C.; Laranjeira, C. Leachates of Eucalyptus globulus in Intermittent Streams Affect Water Parameters and Invertebrates. Int. Rev. Hydrobiol. 2007, 92, 173–182. [Google Scholar] [CrossRef] [Green Version]
- Andriotti, J.; Petrucio, M.M.; Silva, A.L.L.D. Exploring the impacts of non-native leaf litter on invertebrate community and leaf decomposition in a Atlantic Forest stream. Acta Limnol. Bras. 2022, 34, 1–11. [Google Scholar] [CrossRef]
- Eimil-Fraga, C.; Álvarez-Rodríguez, E.; Rodríguez-Soalleiro, R.; Fernández-Sanjurjo, M.J. Influence of Parent Material on the Aluminium Fractions in Acidic Soils Under Pinus pinaster in Galicia (NW Spain). Geoderma 2015, 255–256, 50–57. [Google Scholar] [CrossRef]
- Pardo, I. Comparative water quality characterization by PCA of an unperturbed and a polluted stream. Arch. FÜR Hydrobiol. 1994, 132, 95–114. [Google Scholar] [CrossRef]
- Delgado, C.; Pardo, I. Comparison of Benthic Diatoms from Mediterranean and Atlantic Spanish Streams: Community Changes in Relation to Environmental Factors. Aquat. Bot. 2015, 120, 304–314. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential Oils as Potential Alternative Biocontrol Products Against Plant Pathogens and Weeds: A Review. Foods 2020, 9, 365. [Google Scholar] [CrossRef] [Green Version]
- González-Paz, L.; Almeida, S.F.P.; Pardo, I.; Delgado, C. Periphyton colonization and changes in the diatom assemblages of an artificial urban pond. Fundam. Appl. Limnol. 2020, 193, 313–326. [Google Scholar] [CrossRef]
- APHA. Standard Methods for the Examination of Water, Sewage, and Wastewater, 17th ed.; American Public Health Association: New York, NY, USA, 1989. [Google Scholar]
- Lorenzen, C.J. Determination of chlorophyll and pheo-pigments: Spectrophotometric equations 1. Limnol. Oceanogr. 1967, 12, 343–346. [Google Scholar] [CrossRef]
- BS EN 14407:2004; Water Quality. Guidance Standard for the Identification, and Enumeration of Benthic Diatom Samples from Rivers, and Their Interpretation. British Standards Institution: London, UK, 2004.
- Kelly, M.G.; Cazaubon, A.; Coring, E.; Dell’Uomo, A.; Ector, L.; Goldsmith, B.; Guasch, H.; Hürlimann, J.; Jarlman, A.; Kawecka, B.; et al. Recommendations for the routine sampling of diatoms for water quality assessments in Europe. J. Appl. Phycol. 1998, 10, 215–224. [Google Scholar] [CrossRef]
- AFNOR. Qualité de L’eau—Guide Pour L’échantillonnage en Routine et le Prétraitement des Diatomées Benthiques de Rivières. Norme Française NF EN 13946: 2003. Available online: www.boutique.afnor.org/fr-fr/norme/nf-en-13946/qualite-de-leau-guide-pour-lechantillonnage-en-routine-et-le-pretraitement-/fa102567/21845#AreasStoreProductsSummaryView (accessed on 10 June 2022).
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae, Süsswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1988; Volume 2–2, pp. 1–596. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 3. Teil: Centrales, Fragilariaceae, Eunotiaceae. Süsswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1991; Volume 2–3, pp. 1–600. [Google Scholar]
- Krammer, K.; Lange-Bertalot, H. ‘Bacillariophyceae’. 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema. Gesamtliteraturverzeichnis, Susswasserflora von Mitteleuropa; Verlag: Stuttgart, Germany, 1991; Volume 2–4, pp. 1–437. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 1. Allgemeiner und Encyonema part. Bibl. Diatomol. 1997, 36, 1–382. [Google Scholar]
- Krammer, K. Die Cymbelloiden Diatomeen. Teil 2. Encyonema part, Encyonopsis und Cymbellopsis. Bibl. Diatomol. 1997, 37, 1–469. [Google Scholar]
- Krammer, K. Cymbella. In Diatoms of the European Inlands Waters and Comparable Habitats; Lange-Bertalot, H., Ed.; Koeltz Botanical Books: Königstein, Germany, 2002; Volume 3, p. 584. [Google Scholar]
- Lange-Bertalot, H. 85 neue Taxa und über 100 weitere neu definierte Taxa ergänzend zur Süsswasserflora von Mitteleuropa 2/1–4. Biblioth. Diatomol. 1993, 27, 1–454. [Google Scholar]
- Lange-Bertalot, H. (Ed.) Navicula sensu stricto. 10 genera separated from Navicula sensu lato. Frustulia. In Diatoms of Europe 2; Verlag: Stuttgart, Germany, 2001; p. 526. [Google Scholar]
- Prygiel, J.; Coste, M. Guide Méthodologique Pour la Mise en Œuvre de L’INDICE Biologique Diatomées. NF T90-394Ï; Agence de L’Eau Artois-Picardie: Douai, France, 2000; p. 134. [Google Scholar]
- Trobajo, R.; Rovira, L.; Ector, L.; Wetzel, C.E.; Kelly, M.; Mann, D.G. Morphology and identity of some ecologically important small Nitzschia species. Diatom Res. 2013, 28, 37–59. [Google Scholar] [CrossRef]
- Wetzel, C.E.; Ector, L.; Van der Vijver, B.; Compère, P.; Mann, D.G. Morphology, typification and critical analysis of some ecologically important small naviculoid species (Bacillariophyta). Fottea 2015, 15, 203–234. [Google Scholar] [CrossRef] [Green Version]
- Death, R.G.; Winterbourn, M.J. Environmental Stability and Community Persistence: A Multivariate Perspective. J. N. Am. Benthol. Soc. 1994, 13, 125–139. [Google Scholar] [CrossRef]
- Matthaei, C.D.; Arbuckle, C.J.; Townsend, C.R. Stable surface stones as refugia for invertebrates during disturbance in a New Zealand stream. J. N. Am. Benthol. Soc. 2000, 19, 82–93. [Google Scholar] [CrossRef]
- Tachet, H.; Richoux, P.; Bournaud, M.; Usseglio-Polatera, P. Invertébrés d’Eau Douce, 3rd ed.; CNRS: Paris, France, 2010; p. 607. [Google Scholar]
- McCune, B.; Grace, J.B.; Urban, D.L. Analysis of Ecological Communities; MjM Software Design Gleneden Beach: Gleneden Beach, OR, USA, 2002. [Google Scholar]
- Legendre, P.; Anderson, M.J. Distance-Based Redundancy Analysis: Testing Multispecies Responses in Multifactorial Ecological Experiments. Ecol. Monogr. 1999, 69, 1–24. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting Multivariate Models to Community Data: A Comment on Distance-Based Redundancy Analysis. Ecology 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Anderson, M.; Gorley, R.; Clarke, K.P. For PRIMER: Guide to Software and Statistical Methods; Primer-e: Plymouth, UK, 2008. [Google Scholar]
- Akaike, H. Factor Analysis and the AIC. Psychometrika 1987, 52, 317–332. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER V7. PRIMER-E: Plymouth; Plymouth Marine Laboratory: Plymouth, UK, 2015; Volume 20. [Google Scholar]
- Cemagref, M. Etude Des Méthodes Biologiques D’Appréciation Quantitative De La Qualité Des Eaux. In Rapport Cemagref QE Lyon-AF; Bassin Rhône Méditerranée Corse: Lyon, France, 1982. [Google Scholar]
- Lecointe, C.; Coste, M.; Prygiel, J. Omnidia 3.2. Diatom Index Software Including Diatom Database with Taxonomic Names, References and Codes of 11645 Diatom Taxa. Hydrobiologia 2003, 269, 509–513. [Google Scholar]
- Delgado, C.; Pardo, I.; García, L. A multimetric diatom index to assess the ecological status of coastal Galician rivers (NW Spain). Hydrobiologia 2010, 644, 371–384. [Google Scholar] [CrossRef]
- Kelly, M.G.; Gómez-Rodríguez, C.; Kahlert, M.; Almeida, S.F.; Bennett, C.; Bottin, M.; Delmas, F.; Descy, J.P.; Dörflinger, G.; Kennedy, B.; et al. Establishing expectations for pan-european diatom based ecological status assessments. Ecol. Indic. 2012, 20, 177–186. [Google Scholar] [CrossRef]
- Ferreira, V.; Larrañaga, A.; Gulis, V.; Basaguren, A.; Elosegi, A.; Graça, M.A.S.; Pozo, J. The Effects of Eucalypt Plantations on Plant Litter Decomposition and Macroinvertebrate Communities in Iberian Streams. For. Ecol. Manage. 2015, 335, 129–138. [Google Scholar] [CrossRef]
- Dodds, W.K.; Jones, J.R.; Welch, E.B. Suggested Classification of Stream Trophic State: Distributions of Temperate Stream Types by Chlorophyll, Total Nitrogen, and Phosphorus. Water Res. 1998, 32, 1455–1462. [Google Scholar] [CrossRef]
- Drummond, J.D.; Bernal, S.; von Schiller, D.; Martí, E. Linking in-Stream Nutrient Uptake to Hydrologic Retention in Two Headwater Streams. Freshw. Sci. 2016, 35, 1176–1188. [Google Scholar] [CrossRef]
- Hoellein, T.J.; Tank, J.L.; Rosi-Marshall, E.J.; Entrekin, S.A.; Lamberti, G.A. Controls on spatial and temporal variation of nutrient uptake in three Michigan headwater streams. Limnol. Oceanogr. 2007, 52, 1964–1977. [Google Scholar] [CrossRef]
- Pardo, I.; Kuglerová, L.; García, L.; Martí, E. Nutrient availability modulates the effect of water abstraction on the metabolism of 2 lowland forested streams. Freshw. Sci. 2022, 41, 299–314. [Google Scholar] [CrossRef]
- Grimm, N.B.; Fisher, S.G.; Minckley, W.L. Nitrogen and phosphorus dynamics in hot desert streams of southwestern USA. Hydrobiologia 1981, 83, 303–312. [Google Scholar] [CrossRef]
- Hill, W.R.; Mulholland, P.J.; Marzolf, E.R. Stream ecosystem responses to forest leaf emergence in spring. Ecology 2001, 82, 2306–2319. [Google Scholar] [CrossRef]
- Pardo, I.; Álvarez, M. Comparison of resource and consumer dynamics in Atlantic and Mediterranean streams. Limnetica 2006, 25, 271–286. [Google Scholar] [CrossRef]
- Zhou, Y.H.; Yu, J.Q. Allelochemicals and photosynthesis. In Allelopathy: A physiological Process with Ecological Implications; Reigosa, M.J., Pedrol, N., González, L., Eds.; Springer: Dordrecht, The Netherlands, 2006; pp. 127–156. [Google Scholar]
- Jayakumar, M.; Eyini, M. Allelopathic effect of Eucalyptus globulus labil in groundnut and corn. Comp. Physiol. Ecol. 1990, 15, 109–113. [Google Scholar]
- Zhao, W.; Zheng, Z.; Zhang, J.; Roger, S.; Luo, X. Evaluation of the use of eucalyptus to control algae bloom and improve water quality. Sci. Total Environ. 2019, 667, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Sillett, S.C.; Van Pelt, R.; Kramer, R.D.; Carroll, A.L.; Koch, G.W. Biomass and growth potential of Eucalyptus regnans up to 100m tall. For. Ecol. Manage. 2015, 348, 78–91. [Google Scholar] [CrossRef]
- Vallotton, N.; Moser, D.; Eggen, R.I.L.; Junghans, M.; Chèvre, N. S-metolachlor pulse exposure on the alga Scenedesmus vacuolatus: Effects during exposure and the subsequent recovery. Chemosphere 2008, 73, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Pardo, I.; Delgado, C.; Abraín, R.; Gómez-Rodríguez, C.; García-Roselló, E.; García, L.; Reynoldson, T.B. A predictive diatom-based model to assess the ecological status of streams and rivers of Northern Spain. Ecol. Indic. 2018, 90, 519–528. [Google Scholar] [CrossRef]
- Paule, A.; Roubeix, V.; Lauga, B.; Duran, R.; Delmas, F.; Paul, E.; Rols, J.L. Changes in tolerance to herbicide toxicity throughout development stages of phototrophic biofilms. Aquat. Toxicol. 2013, 144–145, 310–321. [Google Scholar] [CrossRef] [Green Version]
- Esteves, S.M.; Keck, F.; Almeida, S.F.P.; Figueira, E.; Bouchez, A.; Rimet, F. Can we Predict Diatoms Herbicide Sensitivities with Phylogeny? Influence of Intraspecific and Interspecific Variability. Ecotoxicology 2017, 26, 1065–1077. [Google Scholar] [CrossRef]
- Turunen, J.; Markkula, J.; Rajakallio, M.; Aroviita, J. Riparian forests mitigate harmful ecological effects of agricultural diffuse pollution in medium-sized streams. Sci. Total Environ. 2019, 649, 495–503. [Google Scholar] [CrossRef]
- Genkal, S.I.; Yarushina, M.I. To morphology, taxonomy and distribution of rare species of the genus Sellaphora (Bacillariophyta) in Russia. Int. J. Algae 2018, 20, 1–12. [Google Scholar] [CrossRef]
- Cantonati, M.; Zorza, R.; Bertoli, M.; Pastorino, P.; Salvi, G.; Platania, G.; Prearo, M.; Pizzul, E. Recent and Subfossil Diatom Assemblages as Indicators of Environmental Change (Including Fish Introduction) in a High-Mountain Lake. Ecol. Ind. 2021, 125, 107603. [Google Scholar] [CrossRef]
- Kazanci, N.; Ekingen, P.; Türkmen, G.; Ertunç, Ö.; Dügel, M.; Gültutan, Y. Assessment of ecological quality of Aksu Stream (Giresun, Turkey) in Eastern Black Sea Region by using Water Framework Directive (WFD) methods based on benthic macroinvertebrates. Rev. Hydrobiol. 2010, 3, 165–184. [Google Scholar]
- Farhadian, O.; Pouladi, M.; Vazirizadeh, A.; Sedaghat, R. A Study of Diatoms Seasonal Distribution and Biodiversity in Helleh River Estuary, Persian Gulf. Environ. Stud. Persian Gulf 2015, 2, 32–44. [Google Scholar]
- Andersen, D.K.; Larsen, S.E.; Johansson, L.S.; Alnøe, A.B.; Baattrup-Pedersen, A. Udvikling Af Biologisk Indeks for Bentiske Alger (Fytobenthos) i Danske Vandløb; Aarhus Universitet, DCE–Nationalt Center for Miljø og Energi: Roskilde, Denmark, 2018; Volume 42. [Google Scholar]
- González-Paz, L.; Delgado, C.; Pardo, I. How good is good ecological status? A test across river typologies, diatom indices and biological elements. Sci. Total Environ. 2022, 815, 152901. [Google Scholar] [CrossRef] [PubMed]
- García, L.; Pardo, I.; Richardson, J. A cross-continental comparison of stream invertebrate community assembly to assess convergence in forested headwater streams. Aquat. Sci. 2014, 76, 29–40. [Google Scholar] [CrossRef]
- Wasserman, R.J.; Sanga, S.; Buxton, M.; Dalu, T.; Cuthbert, R.N. Does invasive river red gum (Eucalyptus camaldulensis) alter leaf litter decomposition dynamics in arid zone temporary rivers? Inland Waters 2020, 11, 104–113. [Google Scholar] [CrossRef]
- Larrañaga, A.; Basaguren, A.; Pozo, J. Impacts of Eucalyptus globulus plantations on physiology and population densities of invertebrates inhabiting Iberian Atlantic streams. Int. Rev. Hydrobiol. 2009, 94, 497–511. [Google Scholar] [CrossRef]
- García, L.; Pardo, I. Food type and temperature constraints on the fitness of a dominant freshwater shredder. Ann. De Limnol. 2015, 51, 227–235. [Google Scholar] [CrossRef] [Green Version]
- García, L.; Richardson, J.S.; Pardo, I. Leaf quality influences invertebrate colonization and drift in a temperate rainforest stream. Can. J. Fish. Aquat. Sci. 2012, 69, 1663–1673. [Google Scholar] [CrossRef]
- Ledger, M.E.; Hildrew, A.G. The ecology of acidification and recovery: Changes in herbivore-algal food web linkages across a stream pH gradient. Environ. Pollut. 2005, 137, 103–118. [Google Scholar] [CrossRef]
- Kidd, K.A.; Paterson, M.J.; Rennie, M.D.; Podemski, C.L.; Findlay, D.L.; Blanchfield, P.J.; Liber, K. Direct and indirect responses of a freshwater food web to a potent synthetic oestrogen. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130578. [Google Scholar] [CrossRef] [Green Version]
- Dodds, W.K.; Smith, V.H. Nitrogen, Phosphorus, and Eutrophication in Streams. Inland Waters 2016, 6, 155–164. [Google Scholar] [CrossRef]
- Pandey, L.K.; Bergey, E.A. Metal toxicity and recovery response of riverine periphytic algae. Sci. Total Environ. 2018, 642, 1020–1031. [Google Scholar] [CrossRef] [PubMed]
- Karaouzas, I.; Smeti, E.; Vourka, A.; Vardakas, L.; Mentzafou, A.; Tornés, E.; Sabater, S.; Muñoz, I.; Skoulikidis, N.T.; Kalogianni, E. Assessing the ecological effects of water stress and pollution in a temporary river-Implications for water management. Sci. Total Environ. 2018, 618, 1591–1604. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the sampling sites Ventín (1) and As Murteiras (2) streams, located in the Galician region.
Figure 1.
Map of the sampling sites Ventín (1) and As Murteiras (2) streams, located in the Galician region.

Figure 2.
Design of the experimental approach of this study, showing control and impact reaches position in the stream. The net bags containing the eucalypt leaves are also represented, as well as their position in the upstream end (first 20 m) of the impact reach.
Figure 2.
Design of the experimental approach of this study, showing control and impact reaches position in the stream. The net bags containing the eucalypt leaves are also represented, as well as their position in the upstream end (first 20 m) of the impact reach.

Figure 3.
Mean Chl-a concentration during the experiment, measured before (white bars) and after (grey bars) the eucalypt addition in As Murteiras (A) and Ventín (B) streams. Wilcoxon test with * significant at the 0.05 level and ** at the 0.01 level. Error bars represent ±1 SE.
Figure 3.
Mean Chl-a concentration during the experiment, measured before (white bars) and after (grey bars) the eucalypt addition in As Murteiras (A) and Ventín (B) streams. Wilcoxon test with * significant at the 0.05 level and ** at the 0.01 level. Error bars represent ±1 SE.

Figure 4.
Non-metric Multi Dimensional Scaling (nMDS) analysis of As Murteiras (A) and Ventín (B) benthic diatom samples during the experiment. Numbers representing collection week (1, 2 before collections and 3, 4 after collections). Samples were differentiated by symbol and colour: Control before (open up-triangle), control after (open down-triangle), impact before (grey up-triangle), and impact after (black down-triangle). Clustering at 60% (green circle).
Figure 4.
Non-metric Multi Dimensional Scaling (nMDS) analysis of As Murteiras (A) and Ventín (B) benthic diatom samples during the experiment. Numbers representing collection week (1, 2 before collections and 3, 4 after collections). Samples were differentiated by symbol and colour: Control before (open up-triangle), control after (open down-triangle), impact before (grey up-triangle), and impact after (black down-triangle). Clustering at 60% (green circle).

Figure 5.
Diatom indices values measured before (white bars) and after (grey bars) the eucalypt addition in As Murteiras (A) and Ventín (B). Lines on Y axis representing the ecological status boundaries: blue line, high ecological status; and green line, good ecological status. Error bars represent ±1 SE.
Figure 5.
Diatom indices values measured before (white bars) and after (grey bars) the eucalypt addition in As Murteiras (A) and Ventín (B). Lines on Y axis representing the ecological status boundaries: blue line, high ecological status; and green line, good ecological status. Error bars represent ±1 SE.

Figure 6.
Cluster analysis of benthic invertebrate community resemblance, as nMDS overlay of As Murteiras (A) and Ventín (B). Numbers representing collection week. Samples were differentiated by symbol and colour: Control before (open up-triangle), control after (open down-triangle), impact before (grey up-triangle), and impact after (black down-triangle). Clustering at 40% (green circle).
Figure 6.
Cluster analysis of benthic invertebrate community resemblance, as nMDS overlay of As Murteiras (A) and Ventín (B). Numbers representing collection week. Samples were differentiated by symbol and colour: Control before (open up-triangle), control after (open down-triangle), impact before (grey up-triangle), and impact after (black down-triangle). Clustering at 40% (green circle).

Figure 7.
Changes in the average percentage of scrapers at control and impact reaches of As Murteiras (A) and Ventín (B) before (white bar) and after (grey bar) the eucalypt addition. Error bars represent ±1 SE.
Figure 7.
Changes in the average percentage of scrapers at control and impact reaches of As Murteiras (A) and Ventín (B) before (white bar) and after (grey bar) the eucalypt addition. Error bars represent ±1 SE.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Average values and standard error of the physico-chemical variables studied during the experiment in control and impact reaches of As Murteiras and Ventín streams. B (Before), A (After). The discharge was measured at a single point per stream at the downstream end of the impact reach. * Wilcoxon test, p < 0.05.
Table 1.
Average values and standard error of the physico-chemical variables studied during the experiment in control and impact reaches of As Murteiras and Ventín streams. B (Before), A (After). The discharge was measured at a single point per stream at the downstream end of the impact reach. * Wilcoxon test, p < 0.05.
| As Murteiras | Ventín | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | Impact | Significance | Control | Impact | Significance | |||||||
| B | Ar | B | A | B/A | C/I | B | A | B | A | B/A | C/I | |
| Conductivity (µs cm−1) | 19.0 ± 0.2 | 19.2 ± 0.1 | 18.1 ± 0.1 | 19.4 ± 0.7 | * | 19.0 ± 0.2 | 19.0 ± 0.6 | 21.2 ± 1.1 | 21.9 ± 1.7 | * | ||
| pH | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.4 ± 0.0 | 6.7 ± 0.3 | 6.4 ± 0.0 | * | |||
| Dissolved oxygen (mg L−1) | 8.9 ± 0.0 | 8.8 ± 0.1 | 9.2 ± 0.1 | 8.9 ± 0.0 | * | * | 8.8 ± 0.3 | 9.1 ± 0.1 | 8.6 ± 0.0 | 9.0 ± 0.2 | * | |
| Oxygen saturation (%) | 97.9 ± 1.5 | 96.7 ± 1.1 | 100.0 ± 0.6 | 97.0 ± 0.6 | * | * | 95.4 ± 1.2 | 97.8 ± 0.7 | 94.6 ± 0.9 | 96.3 ± 1.5 | * | |
| Water temperature (°C) | 17.6 ± 0.8 | 16.5 ± 0.1 | 17.0 ± 0.7 | 16.9 ± 0.7 | 17.6 ± 0.8 | 16.5 ± 1.3 | 18.1 ± 0.4 | 16.5 ± 0.8 | * | |||
| Water temperature (°C) 24 h | 16.1 ± 0.0 | 15.8 ± 0.0 | * | 15.6 ± 0.0 | 15.3 ± 0.0 | 15.5 ± 0.0 | 15.3 ± 0.0 | * | * | |||
| Luminosity (lum ft−2) | 8.7 ± 0.8 | 9.4 ± 0.8 | 5.6 ± 0.5 | 3.7 ± 0.3 | 10.1 ± 0.9 | 7.0 ± 0.6 | * | * | ||||
| N-DIN (µg L−1) | 10.3 ± 2.4 | 9.2 ± 2.6 | 9.3 ± 0.9 | 7.6 ± 0.2 | 9.5 ± 3.7 | 4.5 ± 1.9 | 20.6 ± 1.2 | 15.5 ± 3.4 | * | * | ||
| P-PO43− (µg L−1) | 6.3 ± 0.2 | 10.9 ± 5.6 | 5.5 ± 0.0 | 8.7 ± 3.3 | 5.7 ± 0.7 | 12.9 ± 6.4 | 5.3 ± 0.4 | 12.3 ± 7.5 | * | |||
| Discharge (m3 s−1) | 0.09 ± 0.06 | 0.11 ± 0.01 | 0.40 ± 0.16 | 0.48 ± 0.12 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
González-Paz, L.; Gestido, J.; Delgado, C.; Pedrol, N.; Pardo, I. Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters. Water 2023, 15, 115. https://doi.org/10.3390/w15010115
AMA Style
González-Paz L, Gestido J, Delgado C, Pedrol N, Pardo I. Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters. Water. 2023; 15(1):115. https://doi.org/10.3390/w15010115
Chicago/Turabian StyleGonzález-Paz, Lorena, Josué Gestido, Cristina Delgado, Nuria Pedrol, and Isabel Pardo. 2023. "Short-Term Effect of Eucalyptus Leachates on Green Food Webs in Headwaters" Water 15, no. 1: 115. https://doi.org/10.3390/w15010115
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.