
SiO2 Nanoparticles Suspension Exposures with Marine Invertebrates: Genotoxicity Response
by
 ,
,
Sergey Petrovich Kukla
1,
Victor Pavlovich Chelomin
1,
Andrey Alexandrovich Mazur
1,*,
Valentina Vladimirovna Slobodskova
1 and
Marina Alexandrovna Mazur
2 1
Il’ichev Pacific Oceanological Institute, Far Eastern Branch, Russian Academy of Sciences, Vladivostok 690041, Russia
2
A.V. Zhirmunsky National Scientific Center of Marine Biology, Far Eastern Branch, Russian Academy of Sciences, Vladivostok 690041, Russia
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 162; https://doi.org/10.3390/w15010162
Submission received: 10 November 2022
/
Revised: 28 December 2022
/
Accepted: 29 December 2022
/
Published: 31 December 2022
(This article belongs to the Section Water Quality and Contamination)
Abstract
:The volume of nanoparticles (NPs) used in various sectors of human life is steadily increasing. Due to their unique properties, silicon dioxide nanoparticles (SiO2 NPs) are widely used in industrial production. However, their ecotoxicological effect on marine biota is extremely poorly understood. To highlight this problem, experiments were conducted with short-term SiO2 NPs exposure to the sand dollar Scaphechinus mirabilis sperm, and a 6-day exposure to the adult bivalve mollusk Mytilus trossulus. SiO2 NPs exposure caused an increase in the concentration of Si in the digestive gland of the mollusk, resulting in DNA damage increase. Exposure to all NPs concentrations caused DNA damage to increase but did not affect the sperm fertilization ability. It was found that the DNA is a target in both chronic and acute SiO2 NPs exposure.
1. Introduction
Since the end of the 20th century, nanotechnology products began to enter everyday life. Today, nanoparticles (NPs) are used in almost all fields of human activity from electronics and mechanical engineering to personal hygiene items. As a result, the NPs production and use continue to grow [1,2]. NPs are part of products that come into direct contact with humans, such as medicines and cosmetics [3,4].
Nanoscale materials acquire new physical and chemical properties that are not characteristic of macroscale particles of the same material. This not only explains the increasing number of NP-containing products on the market but also causes increased interest in them by toxicologists and ecotoxicologists. There is evidence that NPs can enter the human body by crossing the tissue barrier, through the mucous membranes of the lung and digestive system, and directly through the skin [5]. In addition, nanoscale particles easily penetrate the placental and blood–brain barriers in the organism. [6,7].
However, in addition to the risk to human health, scientists are interested in the impact of NPs on marine ecosystems. This is because, in the process of production, use, and utilization, NP-containing products inevitably get into the environment. The endpoint of the migration is most often the aquatic environment. At present, different NPs are found in waste, rivers, lakes, and seawater [8,9,10,11]. A wide range of negative effects of exposure to different NPs on aquatic organisms of various systematic groups has been identified [12].
SiO2 NPs are common in nature as a major component of dust storms in arid regions [13]. Moreover, they are among the most mass-produced NPs today. They are used as an additive in concrete, paint, and plastics, as auxiliary material in the rubber and textile industries, and as an anti-caking agent in various pharmaceuticals [2,14]. The production volumes of SiO2 NPs are estimated to be over one million tons per year [1], and the size of the global market is expected to exceed USD 5 billion by 2025 [2].
Under normal conditions, SiO2 is an inert substance [14]. With the great number of papers devoted to the preparation and use of SiO2 NPs, only a small proportion addresses the toxicity [14]. Still, even these few pieces of data are fragmentary and contradictory [2]. Meanwhile, most ecotoxicological research in this area has been completed on classic model freshwater organisms [14].
Given that in the marine environment, NPs show a tendency for slow sedimentation, accumulating in marine sediments [15], sensitive planktonic organisms and organisms of benthic communities are assumed to be the main ecosystem components affected by NPs. Bivalve mollusks, especially members of the Mytilidae family, are a common model organism in the study of chronic pollutants toxicity. Sea urchin sperm, same time, is a very sensitive model for evaluating pollutant short-term exposure. Thus, our study aimed to investigate the genotoxic possibility of SiO2 NPs in concentrations higher than the natural using two models: sperm of sand dollar S. mirabilis and digestive gland cells of the bivalve mollusk M. trossulus.
2. Material and Methods
2.1. Preparation of Working Suspensions
NPs sized 5–20 nm procured by Sigma-Aldrich were used to prepare the test suspensions. (Cat NO 637246) (Darmstadt, Germany). The basic characteristics of NPs were considered in the work of Breznan et al. (2017) [16] and are presented in Table 1. Before applying the stock suspension of NPs to the aquariums, the suspension was ultrasound treated for 20 min.
2.2. Description of the Experiment
The experiment was carried out at the marine experimental station “Popov Island” Il’ichev Pacific Oceanological Institute. Hydrobionts were collected in coastal zone Sea of Japan. The animals were acclimated under controlled conditions for 3 days in seawater purified with a three-fraction gravel filter followed by UV sterilization. Water parameters were pH 8.1; salinity 32.69 ± 0.14 psu; O2 7.4 ± 0.3 mg/L; T 17.1 ± 0.3 °C with a 16 h photoperiod). After acclimation, two experiment series were carried out.
In the first series, M. trossulus were divided into two groups of 45 specimens each: one group without treatment (control group) and the other with SiO2 NPs (10 mg/L) (experimental group). For the control and experimental groups, 15 mussels were placed in three 15 L tanks for each group. The total duration of the experiment was 6 days. During the experiment, a daily change in seawater with the addition of SiO2 NPs was carried out. Intensive aeration was used to maintain the saturation of water with oxygen and prevent the sedimentation of nanoparticles. The aquariums were maintained at a relatively stable temperature. Animals were not fed; mortality was not recorded.
S. mirabilis spawning was stimulated by 0.5 M KCl. The gametes were prepared according to the standard technique described earlier [17]. Sperm cells were collected and diluted (1:20,000) with pure seawater (filtered and sterilized) just before the experiment. After preparation, the gametes were checked for quality by control fertilization (if the fertilization rate was below 95%, such eggs were not used). Next, the sperm were exposed to SiO2 NPs at different concentrations (5, 10, and 100 mg/L) for 1 h. Fertilization was carried out in pure seawater with incubated spermatozoa. After 20 min, the proportion of zygotes was estimated from the resulting fertilization membrane. In the experiment, 3 parental pairs were used, in 4 parallels. At least 100 zygotes were visually assessed in each parallel.
2.3. Silicon Concentration Measurement
Samples of dried (at 85 °C) pieces of digestive gland tissue were mineralized with a mixture of concentrated acids 16 M HNO3 and 11.2 M HClO4 in a 3:1 ratio at 90 °C. The residue was dissolved in 2.5 mL of bidistilled water.
The concentration of silicon was estimated using atomic absorption spectrophotometer (Shimadzu AA-7000, Kyoto, Japan). All the samples were in triplicate.
2.4. Comet Assay
The digestive gland was extracted from the mussels and washed three times with 4 °C calcium-free magnesium-free buffer (CMF). Then, they were carefully cut into small fragments and placed in CMF buffer. After CMF incubation, the isolated cells were used in comet assay. In the case of work on sperm cells, this procedure was not performed.
An amount of 50 μL of proper cell suspension was mixed with low gelling temperature agarose (100 μL), then cell–agarose suspension was stirred and transferred to a slide containing normal gelling temperature agarose and covered with a coverslip. The slides were kept at 4 °C until the agarose solidified. The coverslip was then carefully removed. The slide was then incubated in the lysis solution for 1 h in a light-protected place at 4 °C. After this, the slides were incubated in electrophoresis buffer (pH > 13) for 40 min. This was followed by electrophoresis for 15 min. After neutralization, the slides were stained with SYBR Green.
A fluorescence microscope Axio Imager A1 (Carl Zeiss, Oberkochen, Germany) with an AxioCam MRc digital camera (Carl Zeiss, Oberkochen, Germany) was used to record comets images. The obtained images were processed using the CaspLab computer program (https://casplab.com (accessed on 15 December 2022), University of Wroclaw, Wroclaw, Poland). In the control and experimental groups, the comets were counted for four individuals in three parallels (N = 12) containing not less than 50 comets (n = 600).
2.5. Statistical Analysis
Statistical analysis was carried out using MS Excel and Statistica software v.10 (Software Inc., Palo. Alto, CA, USA). Al data were tested for homogeneity (Shapiro–Wilk’s test). For comet assay data and successful fertilization nonparametric Kruskal–Wallis ANOVA test was conducted. In all tests, p-value < 0.05 represents statistical significance.
3. Results and Discussion
Benthic filter feeders are a unique group to study the ecotoxicological impact of NPs exposure [18]. Their specific feeding method leads to the accumulation and concentration of a whole range of pollutants, including NPs of different origins.
In the first series of experiments, we demonstrated that silicon was not detected in the digestive gland of the control group of M. trossulus, while after 6 days of staying in water containing 10 mg/L SiO2 NPs, the silicon content increased up to 11.04 ± 0.3 µg/g dry weight (Table 2).
According to the most probable viewpoint, during the filtration process, NPs are concentrated on the gills of the mussel, forming aggregates/agglomerates, and then by the movement of cilia are sent to the mouth and then end up in the digestive gland [19].
It is well known that the digestive gland of mussels is the primary accumulating organ of NPs, such as Ag [20], CuO [21], and TiO2 [22]. The literature data on the accumulation of SiO2 NPs in aquatic organisms indicates that they can also accumulate in the digestive system in Daphnia magna [23] and even penetrate through the chorion into the Danio rerio embryo [24].
Considering that digestive gland cells are characterized by high activity of endocytosis processes representing the main way of NPs entry into the cell [25], most researchers consider exactly the digestive gland as the main target of NPs toxic effect.
Among all major cellular components that can be damaged, nuclear DNA is a critical target. To assess the effect of SiO2 NPs accumulation on the genome of M. trossulus digestive gland cells, the comet assay was applied. Due to the high accuracy of nuclear DNA integrity registration at the earliest stages of genome destruction, this method is widely used in ecotoxicological studies [26]. This analysis is assessed to be several times as sensitive as the biomarkers used to assess toxicity at the organismal level [27].
Visual analysis of the comets formed during the experiment showed that in mussel digestive gland cells exposed to SiO2 NPs, nuclear DNA molecules formed well-defined comets, due to the migration of low-polymer DNA fragments. The results showed that in the control group of digestive gland cells the level of DNA damage was 7.41 ± 0.33% DNA in comet tails. In experimental mollusks accumulating SiO2 NPs, this parameter was 19.28 ± 0.86% of DNA in the tail (Table 2), which is 2.5-fold higher than the control values and indicates the genotoxic properties of these NPs.
To date, there are very few data on the toxic effects of SiO2 NPs on mollusks [14]. Si nanotubes had a cytotoxic effect on the mollusk Modiolus modiolus hemocytes, as well as changed the membrane potential of M. modiolus and Arca boucardi hemocytes [28]. In vitro exposure of Mytilus galloprovincialis hemocytes to SiO2 NPs caused an increased release of extracellular free radicals [29]. The research on short-term exposure to Si NPs in vivo showed induction of catalase activity and imbalance of lipid metabolism in M. galloprovincialis digestive cells [18].
Fertilization and development of embryos and larvae of the sand dollar S. mirabilis occur after the release of gametes directly into the seawater. Gametes and larvae are part of the plankton and are unable to evade contaminated water masses. Directly in seawater, the outer membranes and receptors of sea urchin gametes are exposed to various adverse factors, including NPs. Therefore, sea urchin embryos and larvae are among the most common biological models used to study NPs toxicity [30].
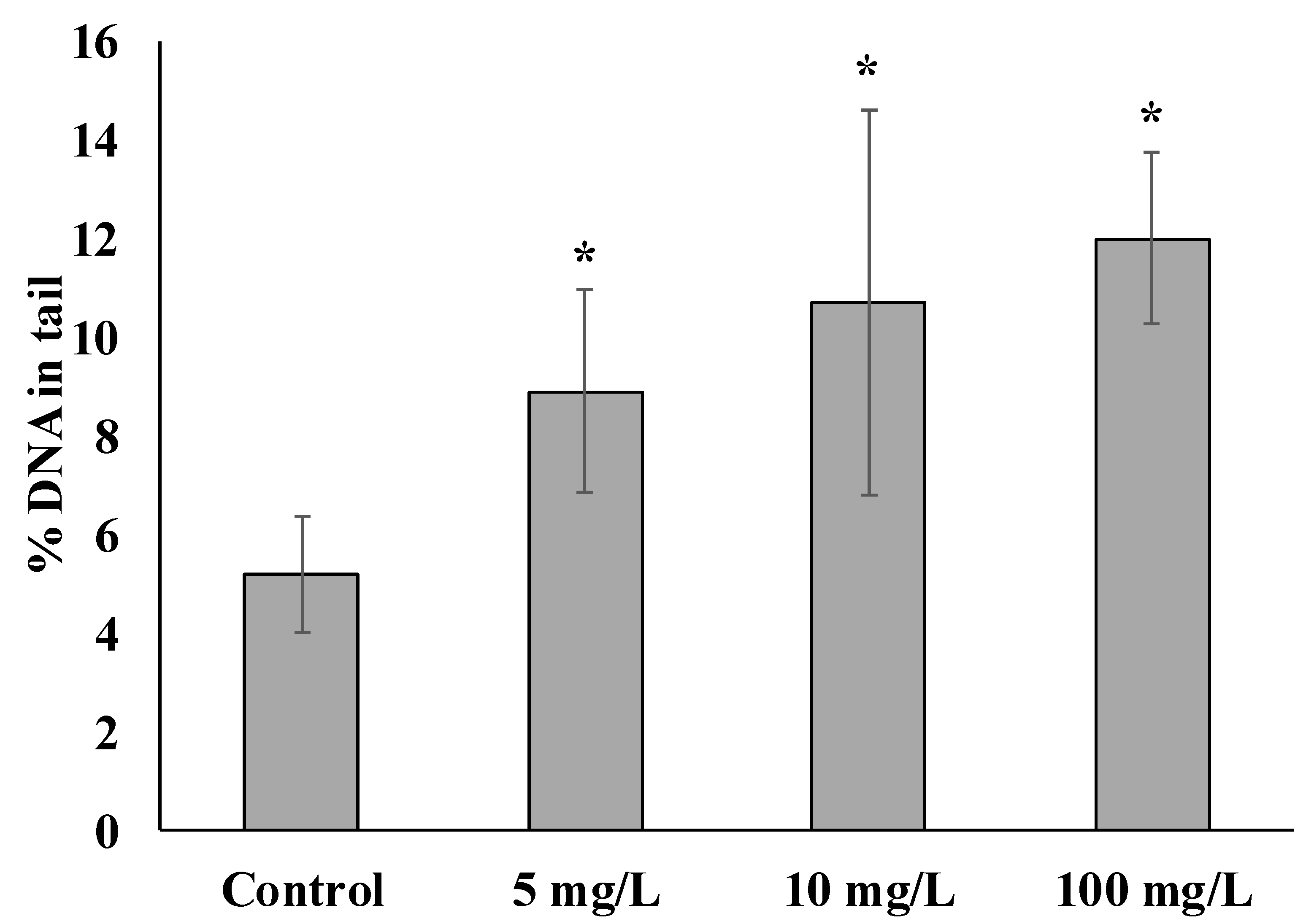
In this respect, laboratory experiments of the interaction of SiO2 NPs with sperm to a certain extent imitate the real situation that occurs when NPs get into the marine environment. Sperm and eggs perform a unique and most important biological function of genome formation for the next generation of development. Therefore, changes in gamete genome integrity when exposed to pollutants represent an ecologically important aspect of toxicity. The diagram shows the results of experiments characterizing the level of genome damage in the S. mirabilis sperm before and after short-term exposure to various concentrations of SiO2 NPs (Figure 1).
In sperm not exposed to NPs, the damage was 5.20 ± 1.18% of DNA in the tail. The presence of slight fragmentation of nuclear DNA in untreated male gametes can be explained by the accumulation of breaks during gametogenesis [31].
Common to all experimental groups is a significant increase in the average level of damaged DNA in the comet tail depending on the concentration of SiO2 NPs. It is of particular interest that even when exposed to the lowest concentration of NPs, the degree of sperm DNA damage increased 1.7-fold relative to the control and amounted to 8.89 ± 2.05% DNA in the tail. When exposed to 10 and 100 mg/L, DNA damage was 10.67 ± 3.9 and 11.99 ± 1.74%, respectively.
Although there are a few experimental works to date showing the genotoxic effects of SiO2 NPs, they have all been carried out in vivo and in vitro on the mammalian model. According to the experimental data, SiO2 NPs increased DNA damage in cells of various organs of mice [32]. In addition, Maser et al., 2015 showed an increase in damage in the DNA molecule in human and rodent cell cultures after 24 h exposure to 15 nm and 55 nm SiO2 NPs (50 μg/mL). At the same time, in vivo exposure of rats to the same particles did not lead to significant DNA damage [33].
Of particular interest are the studies demonstrating changes in gene expression in aquatic organisms after exposure to Si-containing NPs, which indicates the effect of these particles on the functioning of DNA. Thus, exposure to Si-containing NPs caused changes in gene expression in the cells of roundworms, mollusks, echinoderms, and fish [25,34,35,36]. It is important to note that genes responsible for important organism functions were affected. In M. edulis hemocytes exposed to pure Si NPs, changes in gene expression were observed, including those responsible for protein synthesis of the antiradical system and DNA repair system [35]. NPs exposure to adult nematodes Caenohabditis elegans was accompanied by inhibition of the expression of genes responsible for sperm fertilization capacity and normal larval development [25]. Changes in the functioning of such systems pose a serious threat to reproductive potential and normal adult organism development.
Most researchers, when characterizing damage in the DNA molecule, supposerd that it can be caused by both direct and indirect mechanisms. Mechanisms of the first kind are associated with the ability of NPs to bind directly to the DNA molecule and affect its structure [37]. Indirect mechanisms include a whole spectrum of different interactions of NPs with the cell, including NPs dissolution absorbed by the cell, toxic ions formation, and interaction NPs with cell biostructures, with subsequent disturbance of their functions [38].
According to some authors, the genotoxic activity of NPs can be determined by their ability to induce reactive oxygen and nitrogen species (ROS) that damage DNA [39].
Taking into account the characteristic features of the used biomodels, we can assume that SiO2 NPs cause DNA destruction in different ways. In the case of mussels, as we noted above, there is a high probability of SiO2 NPs entering the cells of the digestive gland. Although in the described experiments we did not determine the localization of NPs inside the cells but relied on well-known literature data, we should recognize as the main mechanism of genotoxicity the possibility of the direct effect of SiO2 NPs directly on DNA or components of the DNA repair system. This is confirmed by the work of Chen and Mikecz (2005) [40], who showed that particles can enter not only into the cell but also into the nucleus and interact with DNA.
Given the unique physicochemical properties of NPs in general and SiO2 in particular [2,14], indirect causes of genotoxicity must be considered. The relatively large area of NPs implies finding a large number of atoms with high-energy bonds that easily react with the surrounding molecules to form ROS on the NP’s surface [41].
On the contrary, in the case of sperm cells, the genotoxicity of SiO2 NPs cannot be a consequence of their uptake by cells, since sperm cells do not possess the developed mechanisms of endocytosis, the main pathway of NPs uptake into a cell. In this case, it can be assumed that SiO2 NPs interacted with the receptor-signaling system of the sperm membrane and induced ROS formation, leading to oxidative stress. Sperm are characterized by low DNA repair capacity and weak antioxidant protection in contrast to somatic cells [42,43], which explains their potentially high susceptibility to damage by genotoxic substances. As shown earlier, ROS can be generated directly on the surface of sea urchin sperm [44]. A similar mechanism of ROS formation was likely initiated by exposure of sea urchin sperm Paracentrotus lividus to CuO NPs, which caused significant DNA fragmentation [45]. Significant DNA fragmentation was observed because of CuO NPs exposure to sea urchin sperm of Paracentrotus lividus [45], which was most likely related to ROS formation. As an example, previous studies showed that ZnO NPs [46] as well as chemically inert polystyrene microspheres [47], interacting with the sperm of the sand dollar S. mirabilis, caused DNA damage.
As part of the problem under consideration, to evaluate the functional properties of sperms with different degrees of genome damage, their ability to fertilize eggs was tested. It turned out that at all concentrations of SiO2 NPs used in the experiments and regardless of the level of DNA damage, the sperm retained the ability to fertilize eggs. Fertilization success in all experimental groups did not differ significantly from the control groups and was 95–98% (Figure 2).
Based on the results, we can assume that the NP concentrations used in our experiments are sufficient to cause biochemical changes in the sperm, leading to DNA damage, but not sufficient to affect their fertilization capacity. On the other hand, it is also reasonable to assume that the integrity of the sperm genome is not a critical condition for fertilization in sea urchins.
According to few available literature data, the presented results are characteristic not only for S. mirabilis. Thus, in experiments on the effect of SiO2 NPs on sperm of sea urchin P. lividus, no significant changes in the percentage of fertilized eggs were also observed [48]. A similar result was observed when P. lividus sperm were exposed to amino-modified SiO2 NPs [36].
However, although DNA damage does not affect the fertilization ability of sperm, it can cause serious abnormalities in the further development of embryos and larvae. Thus, in Gambardella et al. (2015) [48], despite the absence of deviations in fertilization, after exposure of sperm to SiO2 NPs, there were observed development delays and abnormalities at the gastrula and pluteus stages. An increase in the anomalies of larval development of sea urchins was also observed after exposure to Ag, Au, ZnO, and CuO NPs [28,49,50,51]. A comparison of the obtained results with the literature data indicates serious preconditions for the development of remote pathological processes after exposure of SiO2 NPs to sperm of sand dollar S. mirabilis.
4. Conclusions
In conclusion, it should be noted that we were among the first to show the genotoxic effect of exposure to SiO2 NPs’ high concentration on marine organisms. It was found that DNA is targeted in chronic NP exposure to benthic filter organisms, as in acute NP exposure to sperm in the water column.
At the same time, the results obtained indicate the need for further ecotoxicological studies of the impact of NPs on marine organisms of different systematic and trophic groups using a multi-marker biochemical approach. Additionally, further research on the SiO2 spermiotoxicity needs to be combined with a larval development study. Identification of the most sensitive targets in the biochemical system of the organism is a necessary step for the transition to the assessment of the toxicity of NPs in a range of concentrations similar to those existing in the environment.
Author Contributions
Conceptualization, S.P.K., V.P.C., and V.V.S.; methodology, S.P.K., V.V.S., and M.A.M.; software, S.P.K.; validation, A.A.M., V.V.S., and M.A.M.; formal analysis, S.P.K., A.A.M., and M.A.M.; investigation, S.P.K., A.A.M., V.V.S., and M.A.M.; resources, V.P.C.; data curation, V.P.C. and V.V.S.; writing—original draft preparation, S.P.K. and V.P.C.; writing—review and editing, S.P.K., A.A.M., and V.V.S.; visualization, S.P.K.; supervision, V.P.C.; project administration, V.P.C.; and funding acquisition, V.P.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the state assignment for research work of V.I. Il’ichev Pacific Oceanological Institute, FEB RAS (No. 121-21500052-9).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors would like to thank Elena Vladimirovna Zhuravel, at the Far Eastern Federal University, for her help with the sand dollar biotesting. L. T. Kovekovdova, a senior researcher at the Pacific branch of the Federal State Budget Scientific Institution “Russian Federal Research Institute of Fisheries and Oceanography” and L.F. Kolosova a leading engineer of the POI FEB RAS, for the determination of silicon content.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Giese, B.; Klaessig, F.; Park, B.; Kaegi, R.; Steinfeldt, M.; Wigger, H.; von Gleich, A.; Gottschalk, F. Risks, release and concentrations of engineered nanomaterial in the environment. Sci. Rep. 2018, 8, 1565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahamed, A.; Liang, L.; Lee, M.Y.; Bobacka, J.; Lisak, G. Too small to matter? Physicochemical transformation and toxicity of engineered nTiO2, nSiO2, nZnO, carbon nanotubes, and nAg. J. Hazard. Mater. 2021, 404 Pt A, 124107. [Google Scholar] [CrossRef]
- D’Mello, S.R.; Cruz, C.N.; Chen, M.L.; Kapoor, M.; Lee, S.L.; Tyne, K.M. The evolving landscape of drug products containing nanomaterials in the United States. Nat. Nanotechnol. 2017, 12, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Fytianos, G.; Rahdar, A.; Kyzas, G.Z. Nanomaterials in cosmetics: Recent updates. Nanomaterials 2020, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Guo, H.; Cui, M.; Huang, R.; Su, R.; Qi, W.; He, Z. Interaction of particles with mucosae and cell membranes. Colloids. Surf. B Biointerfaces 2020, 186, 110657. [Google Scholar] [CrossRef] [PubMed]
- Sawicki, K.; Czajka, M.; Matysiak-Kucharek, M.; Fal, B.; Drop, B.; Meczy’nska-Wielgosz, S.; Sikorska, K.; Kruszewski, M.; Kapka-Skrzypczak, L. Toxicity of metallic nanoparticles in the central nervous system. Nanotechnol. Rev. 2019, 8, 175–200. [Google Scholar] [CrossRef] [Green Version]
- Bongaerts, E.; Nawrot, T.S.; van Pee, T.; Ameloot, M.; Bové, H. Translocation of (ultra) fine particles and nanoparticles across the placenta; a systematic review on the evidence of in vitro, ex vivo, and in vivo studies. Part. Fibre Toxicol. 2020, 17, 56. [Google Scholar] [CrossRef]
- Kaegi, R.; Ulrich, A.; Sinnet, B.; Vonbank, R.; Wichser, A.; Zuleeg, S.; Simmler, H.; Brunner, S.; Vonmont, H.; Burkhardt, M.; et al. Synthetic TiO2 nanoparticle emission from exterior facades into the aquatic environment. Environ. Pollut. 2008, 156, 233–239. [Google Scholar] [CrossRef]
- Gondikas, A.P.; von der Kammer, F.; Reed, R.B.; Wagner, S.; Ranville, J.F.; Hofmann, T. Release of TiO2 nanoparticles from sunscreens into surface waters: A one-year survey at the old Danube recreational Lake. Environ. Sci. Technol. 2014, 48, 5415–5422. [Google Scholar] [CrossRef]
- Markus, A.A.; Krystek, P.; Tromp, P.C.; Parsons, J.R.; Roex, E.W.M.; de Voogt, P.; Laane, R.W.P.M. Determination of metal-based nanoparticles in the river Dommel in the Netherlands via ultrafiltration, HR-ICP-MS and SEM. Sci. Total Environ. 2018, 631–632, 485–495. [Google Scholar] [CrossRef]
- Souza, I.C.; Mendes, V.A.S.; Duarte, I.D.; Rocha, L.D.; Azevedo, V.C.; Matsumoto, S.T.; Elliott, M.; Wunderlin, D.A.; Monferrán, M.V.; Fernandes, M.N. Nanoparticle transport and sequestration: Intracellular titanium dioxide nanoparticles in a neotropical fish. Sci. Total Environ. 2019, 658, 798–808. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.J.; Tyler, C.R.; Galloway, T.S. Impacts of metal and metal oxide nanoparticles on marine organisms. Environ. Pollut. 2014, 186, 257–271. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.; Wei, Y.; Wang, X.; Liu, S.; Zhang, H.; Yuan, S. Mechanistic study of the adsorption and penetration of modified SiO2 nanoparticles on cellular membrane. Chemosphere 2022, 294, 133793. [Google Scholar] [CrossRef]
- Ale, A.; Gutierrez, M.F.; Rossi, A.S.; Bacchetta, C.; Desimone, M.F.; Cazenave, J. Ecotoxicity of silica nanoparticles in aquatic organisms: An updated review. Environ. Toxicol. Pharmacol. 2021, 87, 103689. [Google Scholar] [CrossRef] [PubMed]
- Bundschuh, M.; Filser, J.; Luderwald, S.; McKee, M.S.; Metreveli, G.; Schaumann, G.E.; Schulz, R.; Wagner, S. Nanoparticles in the environment: Where do we come from, where do we go to? Environ. Sci. Eur. 2018, 30, 6. [Google Scholar] [CrossRef] [Green Version]
- Breznan, D.; Das, D.D.; O’Brien, J.S.; MacKinnon-Roy, C.; Nimesh, S.; Vuong, N.Q.; Bernatchez, S.; DeSilva, N.; Hill, M.; Kumarathasan, P.; et al. Differential cytotoxic and inflammatory potency of amorphous silicon dioxide nanoparticles of similar size in multiple cell lines. Nanotoxicology 2017, 11, 223–235. [Google Scholar] [CrossRef] [Green Version]
- Dinnel, P.A.; Stober, Q.J.; Crumley, S.C.; Nakatani, R.E. Development of a sperm cell toxicity test for marine water. Aquat. Toxicol. Haz. Asses. 1982, 1, 82–98. [Google Scholar] [CrossRef]
- Canesi, L.; Fabbri, R.; Gallo, G.; Vallotto, D.; Marcomini, A.; Pojana, G. Biomarkers in Mytilus galloprovincialis exposed to suspensions of selected nanoparticles (Nano carbon black, C60 fullerene, Nano-TiO2, Nano-SiO2). Aquat. Toxicol. 2010, 100, 168–177. [Google Scholar] [CrossRef]
- Browne, M.A.; Dissanayake, A.; Galloway, T.S.; Lowe, D.M.; Thompson, R.C. Ingested microscopic plastic translocates to the circulatory system of the mussel, Mytilus edulis (L.). Environ. Sci. Technol. 2008, 42, 5026–5031. [Google Scholar] [CrossRef]
- Al-Sid-Cheikh, M.; Rouleau, C.; Pelletier, E. Tissue distribution and kinetics of dissolved and nanoparticulate silver in Iceland scallop (Chlamys islandica). Mar. Environ. Res. 2013, 86, 21–28. [Google Scholar] [CrossRef]
- Chelomin, V.P.; Slobodskova, V.V.; Zakhartsev, M.K.; Kukla, S.P. Genotoxic potential of copper oxide nanoparticles in the bivalve mollusk Mytilus trossulus. J. Ocean Univ. China 2017, 16, 339–345. [Google Scholar] [CrossRef]
- Kukla, S.; Slobodskova, V.; Mazur, A.; Chelomin, V.; Kamenev, Y. Genotoxic testing of titanium dioxide nanoparticles in Far Eastern mussels, Mytilus trossulus. Pollution 2021, 7, 129–140. [Google Scholar] [CrossRef]
- Yang, S.; Ye, R.; Han, B.; Wei, C.; Yang, X. Ecotoxicological effect of nano-silicon dioxide particles on Daphnia magna. Integr. Ferroelectr. 2014, 154, 64–72. [Google Scholar] [CrossRef]
- Pham, D.H.; Roo, B.D.; Nguyen, X.B.; Vervaele, M.; Kecskés, A.; Ny, A.; Copmans, D.; Vriens, H.; Locquet, J.P.; Hoet, P.; et al. Use of zebrafish larvae as a multi-endpoint platform to characterize the toxicity profile of silica nanoparticles. Sci. Rep. 2016, 6, 37145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, H.J.; Choi, J. Clathrin-mediated endocytosis is involved in uptake and toxicity of silica nanoparticles in Caenohabditis elegans. Chem. Biol. Interact. 2019, 311, 108774. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.N. Ecotoxicological applications and significance of the comet assay. Mutagenesis 2008, 23, 207–221. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Fernandez-Triana, J.; Roughley, R.; Hebert, D.N. DNA barcode accumulation curves for understudied taxa and areas. Mol. Ecol. Resour. 2009, 9, 208–216. [Google Scholar] [CrossRef]
- Pikula, K.; Chaika, V.; Zakharenko, A.; Savelyeva, A.; Kirsanova, I.; Anisimova, A.; Golokhvast, K. Toxicity of carbon, silicon, and metal-based nanoparticles to the hemocytes of three marine bivalves. Animals 2020, 10, 827. [Google Scholar] [CrossRef]
- Canesi, L.; Ciacci, C.; Vallotto, D.; Gallo, G.; Marcomini, A.; Pojana, G. In Vitro effects of suspensions of selected nanoparticles (C60 fullerene, TiO2, SiO2) on Mytilus hemocytes. Aquat. Toxicol. 2010, 96, 151–158. [Google Scholar] [CrossRef]
- Gambardella, C.; Ferrando, S.; Gatti, A.M.; Cataldi, E.; Ramoino, P.; Aluigi, M.G.; Faimali, M.; Diaspro, A.; Falugi, C. Review: Morphofunctional and biochemical markers of stress in sea urchin life stages exposed to engineered nanoparticles. Environ. Toxicol. 2016, 11, 1552–1562. [Google Scholar] [CrossRef]
- Mahaye, N.; Thwala, M.; Cowan, D.A.; Musee, N. Genotoxicity of metal based engineered nanoparticles in aquatic organisms: A review. Mutat. Res. 2017, 773, 134–160. [Google Scholar] [CrossRef] [PubMed]
- Nemmar, A.; Yuvaraju, P.; Beegam, S.; Yasin, J.; Kazzam, E.E.; Ali, B.H. Oxidative stress, inflammation, and DNA damage in multiple organs of mice acutely exposed to amorphous silica nanoparticles. Int. J. Nanomed. 2016, 11, 919–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maser, E.; Schulz, M.; Sauer, U.G.; Wiemann, M.; Ma-Hock, L.; Wohlleben, W.; Hartwig, A.; Landsiedel, R. In Vitro and in vivo genotoxicity investigations of differently sized amorphous SiO2 nanomaterials. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 794, 57–74. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Li, Q.; Jiang, L.; Zou, Y.; Duan, J.; Sun, Z. Genome-Wide transcriptional analysis of silica nanoparticle-induced toxicity in zebrafish embryos. Toxicol. Res. 2016, 5, 609–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barrick, A.; Mouneyrac, C.; Manier, N.; De Lantivy, L.; Jrad, N.; Châtel, A. Towards the development of a high throughput screening approach for Mytilus edulis hemocytes: A case study on silicon-based nanomaterials. Mar. Environ. Res. 2018, 142, 306–318. [Google Scholar] [CrossRef]
- Tacconi, S.; Augello, S.; Persano, F.; Sbarigia, C.; Carata, E.; Leporatti, S.; Fidaleo, M.; Dini, L. Amino-Functionalized Mesoporous silica nanoparticles (NH 2-MSiNPs) impair the embryonic development of the sea urchin Paracentrotus lividus. Environ. Toxicol. Pharmacol. 2022, 95, 103956. [Google Scholar] [CrossRef] [PubMed]
- Magdolenova, Z.; Collins, A.; Kumar, A.; Dhawan, A.; Stone, V.; Dusinska, M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology 2014, 8, 233–278. [Google Scholar] [CrossRef]
- Chang, Y.N.; Zhang, M.; Xia, L.; Zhang, J.; Xing, G. The toxic effects and mechanisms of CuO and ZnO nanoparticles. Materials 2012, 5, 2850–2871. [Google Scholar] [CrossRef] [Green Version]
- Manke, A.; Wang, L.; Rojanasakul, Y. Mechanisms of nanoparticle-induced oxidative stress and toxicity. Biomed. Res. Int. 2013, 2013, 942916. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; von Mikecz, A. Formation of nucleoplasmic protein aggregates impairs nuclear function in response to SiO2 nanoparticles. Exp. Cell Res. 2005, 305, 51–62. [Google Scholar] [CrossRef]
- Nabeshi, H.; Yoshikawa, T.; Matsuyama, K.; Nakazato, Y.; Tochigi, S.; Kondoh, S.; Hirai, T.; Akase, T.; Nagano, K.; Abe, Y.; et al. Amorphous nanosilica induce endocytosis-dependent ROS generation and DNA damage in human keratinocytes. Part. Fibre Toxicol. 2011, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewis, C.; Galloway, T.S. Genotoxic damage in Polychaetes: A study of species and cell-type sensitivities. Mutat. Res. Genet. Toxicol. Environ. Mutat. 2008, 654, 69–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devaux, A.; Fiat, L.; Gillet, C.; Bony, S. Reproduction impairment following paternal genotoxin exposure in brown trout (Salmo trutta) and Arctic charr (Salvelinus alpinus). Aquat. Toxicol. 2011, 101, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Kazama, M.; Hino, A. Sea urchin spermatozoa generate at least two reactive oxygen species; the type of reactive oxygen species changes under different conditions. Mol. Reprod. Dev. 2012, 79, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Manfra, L.; Boni, R.; Rotini, A.; Migliore, L.; Tosti, E. Cytotoxicity and genotoxicity of CuO nanoparticles in sea urchin spermatozoa through oxidative stress. Environ. Int. 2018, 118, 325–333. [Google Scholar] [CrossRef]
- Kukla, S.P.; Chelomin, V.P.; Mazur, A.A.; Slobodskova, V.V. Zinc oxide nanoparticles induce DNA damage in sand dollar Scaphechinus mirabilis sperm. Toxics 2022, 10, 348. [Google Scholar] [CrossRef]
- Mazur, A.A.; Chelomin, V.P.; Zhuravel, E.V.; Kukla, S.P.; Slobodskova, V.V.; Dovzhenko, N.V. Genotoxicity of polystyrene (PS) microspheres in short-term exposure to gametes of the sand dollar Scaphechinus mirabilis (Agassiz, 1864) (Echinodermata, Echinoidea). J. Mar. Sci. Eng. 2021, 9, 1088. [Google Scholar] [CrossRef]
- Gambardella, C.; Morgana, S.; Bari, G.D.; Ramoino, P.; Bramini, M.; Diaspro, A.; Falugi, C.; Faimali, M. Multidisciplinary screening of toxicity induced by silica nanoparticles during sea urchin development. Chemosphere 2015, 139, 486–495. [Google Scholar] [CrossRef]
- Burić, P.; Jakšić, Ž.; Štajner, L.; Dutour Sikirić, M.; Jurašin, D.; Cascio, C.; Calzolai, L.; Lyons, D.M. Effect of silver nanoparticles on Mediterranean sea urchin embryonal development is species specific and depends on moment of first exposure. Mar. Environ. Res. 2015, 111, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Mazur, A.A.; Zhuravel, E.V.; Slobodskova, V.V.; Mazur, M.A.; Kukla, S.P.; Chelomin, V.P. Waterborne exposure of adult sand dollar, Scaphechinus mirabilis (Agassiz, 1864), to zinc ions and zinc oxide nanoparticles affects early development of its offspring. Water Air Soil Pollut. 2020, 231, 115. [Google Scholar] [CrossRef]
- Kukla, S.P.; Slobodskova, V.V.; Zhuravel, E.V.; Mazur, A.A.; Chelomin, V.P. Exposure of adult sand dollars (Scaphechinus mirabilis) (Agassiz, 1864) to copper oxide nanoparticles induces gamete DNA damage. Environ. Sci. Pollut. Res. Int. 2022, 26, 39451–39460. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
DNA damage of sperm exposed with different concentrations of NPs (mean ± standard deviation, n = 3). * Difference from the control is significant (p < 0.05).
Figure 1.
DNA damage of sperm exposed with different concentrations of NPs (mean ± standard deviation, n = 3). * Difference from the control is significant (p < 0.05).

Figure 2.
Fertilization success of S. mirabilis eggs by sperm exposed by different concentrations of SiO2 NPs (mean ± standard deviation).
Figure 2.
Fertilization success of S. mirabilis eggs by sperm exposed by different concentrations of SiO2 NPs (mean ± standard deviation).


{kind=link}
{kind=link}
Table 1.
The basic properties of SiO2 NPs.
| Size, nm | Hydrodynamic Size, nm | Purity, % | Total Surface Area, m2/g | Zetta Potential, mV |
|---|---|---|---|---|
| 20 ± 4.6 | 576 ± 104 | 95.5 | 613 ± 47 | −11.5 ± 0.7 |
Table 2.
Si concentration and DNA damage level in digestive gland cells of experimental and control mussels (mean ± standard deviation). n.d.—not detected. * Difference from the control is significant (p < 0.05).
Table 2.
Si concentration and DNA damage level in digestive gland cells of experimental and control mussels (mean ± standard deviation). n.d.—not detected. * Difference from the control is significant (p < 0.05).
| Parameter | Control Group | Experimental Group |
|---|---|---|
| Silicon concentration, µg/gdw | n.d. | 11.04 ± 0.3 * |
| DNA damage, %DNA in tail | 7.41 ± 0.33 | 19.28 ± 0.86 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kukla, S.P.; Chelomin, V.P.; Mazur, A.A.; Slobodskova, V.V.; Mazur, M.A. SiO2 Nanoparticles Suspension Exposures with Marine Invertebrates: Genotoxicity Response. Water 2023, 15, 162. https://doi.org/10.3390/w15010162
AMA Style
Kukla SP, Chelomin VP, Mazur AA, Slobodskova VV, Mazur MA. SiO2 Nanoparticles Suspension Exposures with Marine Invertebrates: Genotoxicity Response. Water. 2023; 15(1):162. https://doi.org/10.3390/w15010162
Chicago/Turabian StyleKukla, Sergey Petrovich, Victor Pavlovich Chelomin, Andrey Alexandrovich Mazur, Valentina Vladimirovna Slobodskova, and Marina Alexandrovna Mazur. 2023. "SiO2 Nanoparticles Suspension Exposures with Marine Invertebrates: Genotoxicity Response" Water 15, no. 1: 162. https://doi.org/10.3390/w15010162
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.