
BioH2 Production Using Microalgae: Highlights on Recent Advancements from a Bibliometric Analysis
by
 , , and
, , and
Shirin P. Arimbrathodi
1,
Muhammad Asad Javed
1,† ,
,
Mohamed A. Hamouda
1,2,* ,
,
Ashraf Aly Hassan
1,2 and
Mahmoud E. Ahmed
1
1
Department of Civil and Environmental Engineering, United Arab Emirates University, Al Ain P.O. Box 15551, United Arab Emirates
2
National Water and Energy Center, United Arab Emirates University, Al Ain P.O. Box 15551, United Arab Emirates
*
Author to whom correspondence should be addressed.
†
Equal contribution with the first author.
Water 2023, 15(1), 185; https://doi.org/10.3390/w15010185
Submission received: 28 November 2022
/
Revised: 26 December 2022
/
Accepted: 28 December 2022
/
Published: 2 January 2023
(This article belongs to the Section Wastewater Treatment and Reuse)
Abstract
:Demand for clean energy has increased due to the proliferation of climate change impact from excessive emission of greenhouse gases (GHG) from the combustion of fossil fuels. H2 is a clean energy source since water vapor is the only byproduct after its combustion. Growing microalgae offers a promising low-energy and low-cost approach for bioH2 production. In this study, a bibliometric analysis was performed for the production of H2 using microalgae to evaluate the conceptual, intellectual, and social structure of the dataset. In addition, a scoping review of articles was conducted to highlight recent advancements and identify future research recommendations. A total of 184 relevant publications over 23 years (2000–2022) were retrieved from the Scopus database for analysis. The results demonstrated an exponential increase in citations from 283 to 996 in the last decade, indicating the interest in bioH2 production from microalgae. Results also revealed that the International Journal of Hydrogen Energy accounted for more than 25% of the published articles, of which China contributed almost 28%. Oxygen sensitivity of the H2ase enzyme and sulfur deprivation were highlighted as the main limiting factors of bioH2 production using microalgae. It was also evident that the most widely studied microalgae species were green algae, especially Chlamydomonas and Chlorella. Effective process modifications, particularly hybridizing microalgae with bacteria consortium and implementing oxygen regulating strategies, were shown to give up to a 10-fold increase in H2 yield. This study also discusses recent developments in technologies, strategies, microalgal species, and optimizing controlling factors affecting bioH2 production.
1. Introduction
Increases in environmental pollution, global warming, and climate change are mainly associated with the emission of greenhouse gasses (GHGs) from the combustion of non-renewable fuels. Therefore, alternative energy sources are required to control these emissions. The Sustainable Development Goals and net-zero commitments of the United Nations also urge nations to adopt alternative sustainable energy sources [1]. Hydrogen (H2) is considered a viable alternative energy source because of its high energy density by mass (142 MJ/kg) and pure combustion product, water (H2O) [2,3]. H2 energy can be beneficial in alleviating the environmental problems caused by the fuel crisis and greenhouse gas emissions. However, the high cost of H2 generation is a significant barrier to developing the H2 economy. The primary method for producing H2 is by steam reforming of methane, which uses a sizable amount of fossil fuel. The overall effectiveness might be restricted to 60% in the case of H2 production from electricity based on electrolysis technology [2,4]. BioH2 can be produced using several sources (feedstocks) [5,6]. There is currently a great need for a sustainable source of H2. Microalgae is a promising feedstock and probably a higher efficiency route for the synthesis of H2, mainly because of its greater carbon dioxide (CO2) fixation efficiency, higher growth rate, greater photosynthetic efficiency, higher energy density, high lipid concentration, higher nutrient acquisition, and the capacity to flourish in different circumstances [7,8,9,10].
Diverse microalgae can generate bioH2, including Chlorella sp., Scenedesmus sp., Monoraphidium sp., Platymonas sp., Tetraspora sp., Closterium sp., and Chlamydomonas sp. [11,12,13,14]. Even though green microalgae have high potential as a renewable energy source, only about 70 species from more than 30 genera have been researched so far [15]. Chlorella sp. is the most favorable for a high bioH2 production potential. C. vulgaris var. vulgaris, C. pyrenoidosa, C. sorokiniana, C. fusca, C. lewinii, C. homosphaera, and C. protothecoides are some important Chlorella species [16,17]. Moreover, C. reinhardtii is a common microalga model to investigate for H2 generation. Sulfur-deprived C. reinhardtii could produce H2 continuously, even in the absence of acetate or any other organic substrates in the medium under strictly photoautotrophic conditions [18,19,20]. Blue-green algae, also known as cyanobacteria, can produce H2. Moreover, cyanobacteria are filamentous nitrogen organisms with special cells named heterocysts for nitrogen fixation. The classic nitrogen-fixing genera include species like Anabaena, Calothrix, Nostoc, and Oscillatoria. Anabena has gained attention due to its bioH2 production during nitrogen fixation, but its metabolic models have not yet been utilized to study bioH2 production [14,21].
Different biological techniques can achieve bioH2 production from microalgae. Still, the most distinguished techniques are bio-photochemical (direct or indirect photolysis), fermentation process (light or dark), bio-electrochemical (microbial fuel and electrolysis cells), and lastly, thermochemical conversion (pyrolysis and gasification) [22,23]. The efficient production of biohydrogen yield from microalgae biomass depends on factors such as nutrients, pH, temperature, light intensity, photoreactor configuration, substrate concentration, and cell density. Various strategies can be adopted to improve hydrogen production using microalgae, such as immobilization, pretreatment techniques, and genetic engineering.
Many articles have already discussed the various aspects of bioH2 production using microalgae [24,25]. However, this study is different from the previous works because the main interest of this paper is to conduct a bibliometric analysis on bioH2 production using microalgae. Bibliometric analysis is an important quantitative and statistical tool to determine the growth and development of a specific research field by evaluating the conceptual, intellectual, and social structure of the dataset [26]. It highlights the contribution of authors, nations, journals, and their collaborations by carefully analyzing the networks of keywords, authors, and articles. This allows researchers to understand several aspects, such as key research topics, emerging themes, and novel strategies and to subsequently identify the research gaps. Recently, bibliometric analysis was used to explore progress in the production of bioH2 energy from food waste [26] and bioH2 production from dark fermentation [27]. However, a bibliometric analysis focusing on bioH2 production using microalgae has yet to be performed.
The main focus of this paper is to conduct a bibliometric analysis of the published research, discuss the recent advancements, and identify future research recommendations in bioH2 production from microalgae. A comprehensive bibliometric analysis of the relevant literature published between 2000 and 2022 was conducted to answer the following research questions:
- What trends can be detected when analyzing literature investigating bioH2 production from microalgae?
- Who are the major contributors to this research area?
- What are the recent advancements and research gaps?
The structure of this paper has six sections. Section 2 is a brief background on biohydrogen production using microalgae, while Section 3 provides the methodology employed in this research. Next, Section 4 discusses the results obtained from the bibliometric analysis. Then, Section 5 provides highlights of recent developments and future research opportunities in the field of biohydrogen production using microalgae. Section 6 presents the conclusion.
2. Background
2.1. The Techniques to Produce BioH2 Using Microalgae
Microalgae contain pigment molecules capable of absorbing solar energy and converting it into chemical energy by simultaneously splitting water into oxygen (O2) and protons (H+). The photosynthetic electron transfer constitutes light and dark reactions. The light reaction helps to obtain electrons by splitting water in photosystem II (PSII) and transfer the electrons through an electron transport chain from PSII to Photosystem I (PSI). This results in the generation of Adenosine triphosphate (ATP) and strong reductants (NAD(P)H). Photobiological bioH2 production is associated with photosynthesis, where the final electron acceptor, ferredoxin (Fd), donates electrons to enzymes involved in H2 metabolism [28]. Biophotolysis is the initial step of microalgal bioH2 production. In direct biophotolysis, microalgae convert solar energy into chemical energy, and H2 is derived from the electrons and protons generated by the water splitting at PSII. Nevertheless, some of the restrictions of biophotolysis include O2 generation by the activity of PSII, the requirement for a customized photobioreactor, the sensitivity of H2ase to O2, and low yield [29,30]. For indirect biophotolysis, electrons and protons are mainly supplied by the degradation of intracellular carbon compounds. Indirect photolysis has two stages: first, the carbohydrate biomass is generated from photosynthesis, and in the next stage, H2 and CO2 are produced due to the fermentation of carbohydrate-rich biomass. In these two steps, oxygen and H2 will be separated. This prevents enzyme deactivation and removes CO2 from the H2 and CO2 mixture, making H2 purification easier. Some drawbacks include high H2 selectivity, the restricted effect of O2 on the H2ase, and low yield [31]. Figure 1 shows a schematic diagram of hydrogen production by direct and indirect biophotolysis.
Dark fermentation has gained attention because of its relatively high bioH2 production rates. The complex organic substances (lipids, carbohydrates, and proteins) are subjected to four stages: hydrolysis, acidogenesis, acetogenesis, and methanogenesis. Firstly, lipids, carbohydrates, and proteins are hydrolyzed into sugars, fatty acids, and amino acids. In the acidogenesis stage, the hydrolyzed products are acidified to form H2, CO2, fatty acids, and other intermediates. In the next step, acetogenesis, the fatty acids produced are again fermented to generate H2 and acetate. The final products, methane (CH4) and CO2, are formed from the decarboxylation of acetate by the acetoclastic methanogens. H2-utilizing methanogens consume H2 gas as an electron donor while reducing the carbon dioxide to methane. In dark fermentation, H2 is produced as an intermediate metabolite in the acidogenesis and acetogenesis transformations. Two mechanisms which involve specific coenzymes are responsible for the evolution of H2 gas, either by formic acid catabolic transformation or by re-oxidation of nicotinamide adenine dinucleotide (NADH) catalyzed by H2ase pathway; H2ase is the major enzyme in the process [32,33,34]. Dark fermentation has many advantages: it is capable of continually producing H2 without depending on sunlight, has high energy efficiency, is an eco-friendly and economical process, is easy to commercialize, has a less complicated bioreactor design, and has a wide range of organic acid as byproducts. The major disadvantages are low bioH2 production due to the accumulation of O2, methanogenic bacteria utilizing H2 as an electron donor, more tedious and expensive H2 recovery due to the generation of CO2 and other gaseous products, and low substrate conversion efficiency [24,25,32].
In photo fermentation, H2 is produced by the photosynthetic bacteria by breaking down the organic compounds with the help of nitrogenase (N2ase) enzymes under nitrogen-deprived conditions. Atmospheric nitrogen is converted into ammonium ions used by microorganisms as a nitrogen source through nitrogen (N2) fixation by nitrogenase. They are only found in cyanobacteria and non-sulfur purple and green sulfur bacteria. These bacteria consume acetic acid and use ATP as an energy source. This leads to the transfer of the electrons by ferredoxin to the enzyme nitrogenase and results in N2 fixation. Moreover, nitrogenase protons are converted to H2 in the absence of nitrogen [25,33]. The major advantages are that photosynthetic bacteria use various spectral energy and substrates, can treat effluents from dark fermentation, have higher substrate transformation efficiency, and hence have a high H2 yield. One disadvantage is the requirement of a light source; the photosynthesis efficiency directly depends on the availability of light. Moreover, it requires a large area and an anaerobic bioreactor, which increases costs [24,32].
Thermochemical processes include liquefaction, gasification, and pyrolysis. The wet microalgae biomass is converted into gaseous bioH2 during hydrothermal gasification. The process includes heating biomass at higher temperatures in a compressed water medium. The reaction is quick due to the higher temperatures; the main products obtained are H2, CO2, and methane (CH4). The compressed water medium usually has a low percentage of effluents and can be reused [35,36]. The microalgae should be dried up to low moisture content in conventional thermal gasification for higher efficiency. Nevertheless, the high moisture content of microalgae often results in high energy consumption during thermal gasification. Thus, the main advantage of applying supercritical water gasification (SCWG) is that it can be conducted without the drying process, but rather in an aqueous state. Lastly, pyrolysis occurs in the absence of oxygen at higher temperatures. Dry microalgae are needed to be fed to the reactor, which demands a large amount of energy. The conversion efficiency of microalgae depends on parameters such as reaction temperature, retention time, and the composition of feedstock [9,37].
Bioelectrochemical systems are an alternate method for bioH2 production. Microalgae catalyze the oxidation–reduction at the anode and cathode, respectively, and act as electrochemical catalysts. Microbial fuel cells and microbial electrolysis cells are the two categories of bioelectrochemical systems. Microbial Fuel Cells (MFCs) are eco-friendly bioelectrochemical devices that produce electrical energy from chemical energy obtained from biomass. The H2 forms at the cathodes as a result of the reduction reaction and is collected by an external system. The primary restriction of MFC is membrane fouling, which occurs due to the long-term growth of biofilm in separators. It can also occur due to the accumulation of microbes, which leads to the formation of thick biofilm on the surface of the membrane. This prevents the transfer of H2 ions from the anode to the cathode. Another restriction is the removal of heavy metals, which causes lower efficiency performance of microbes and pH imbalance. The higher cost of the membrane prevents the large-scale expansion of MFC. Microbial Electrolysis Cells (MECs) are a different form of MFC; the bioH2 is produced by the oxidation of organic matter, which is catalyzed by electroactive biofilms. It is yet to be developed in an efficient and scalable design. One major advantage of MEC is the higher efficiency in metal ions removal. The drawbacks are the higher cost as well as the H2 loss and contamination [25,38,39].
2.2. Factors Affecting BioH2 Production Using Microalgae
Efficient production of bioH2 yield from microalgae biomass depends on factors such as nutrients, pH, temperature, light intensity, photoreactor configuration, substrate concentration, and cell density. Compared to a near-neutral pH, higher H2 productivity was observed when the pH was around 6. At a highly acidic pH, H2 yield declines because of the inactivation of the acetate-producing bacteria. Temperature is another parameter that influences the metabolic pathways of H2ase. A temperature range of 15–35 °C is good for microalgal growth. Moreover, the proper configuration of the photoreactor is important as it is critical for the effective use of light and the provision of sufficient surface area for the growth of microalgae. Additive subtracts such as biotin, cyanocobalamin, and thiamine are required to add to the culture to support the maximum cell growth and bioH2 production. For optimal bioH2 production, the culture needs a balance of carbohydrate-based substrate. Moreover, a carbon source is also required for the microalgae to flourish in all conditions except photoautotrophic conditions [15,40].
Furthermore, high bioH2 production and good microalgal growth can be achieved by introducing the proper fraction of nutrients such as nitrogen, phosphorus, and trace elements. The nitrogen element mainly regulates the protein synthesis and growth metabolites of microalgae, while the phosphorus element regulates most of the cell’s activities and metabolism. The trace elements like magnesium, sodium, and zinc are important supplements that play a role in improving microalgal cultivation. The problem associated with the inhibition of H2ase is caused by the presence of oxygen and can be resolved by sulfur deprivation. BioH2 production is relatively low at lower light intensities. Exposure to high light intensity can increase bioH2 production rates by inhibiting photosynthetic O2 [25,41]. In addition, optimal light conditions can reduce the lag period of microalgae and increase H2 yield [25,41]. Cell density controls the amount of light that passes through the microalgal cell, and it depends on the nature of the cultivation process. The low cell concentration will not allow the uptake of the dissolved O2 into the microalgae culture. In contrast, a high cell density may cause the cumulation of starch and hinder the productivity rate. Therefore, to have significant bioH2 production, an active growth phase and cell density for the culture should be maintained [15]. The application of bioH2 production using microalgae is still limited due to the lack of proper distribution, capture, storage, and transformation technologies.
2.3. Strategies to Improve the BioH2 Production from Microalgae
Various strategies can be adopted to improve bioH2 production using microalgae, such as immobilization of microalgae, pretreatment techniques, nanoparticles, and genetic engineering. The pretreatment immediately disrupts the microalgal cell walls and enhances the accessibility of carbohydrates present in the cells. Different pretreatment methods include chemical, thermal, mechanical, enzyme, and combined methods. The best pretreatment method and its optimal conditions are yet to be determined [15,25].
Microalgal immobilization is the mechanism of the entrapment of microalgal cells on or into solid support. It has many advantages, such as high cell density, alleviating manipulation of cultures, and easy microalgae cell harvesting. Further, this approach protects the cells from unwanted contaminations and sudden changes in other culture parameters. This also results in high bioH2 production due to the enhanced permeability of cell walls. In addition, the microalgal cells wash out, get reduced, and cause an overall increase in H2 yields. The major drawbacks are the slow infusion of nutrients from the medium into microalgae and the high sunlight gradient within the cells because of high cell density [12,13]. Nanotechnology is capable of bioH2 production due to its role in intracellular electron transfer, microalgal growths, and enzymes involved in bioH2 generation. Genetic engineering and metabolic engineering can be used to modify specific pathways to increase bioH2 production. The photosynthetic barriers and inhibition factors can be suppressed [24].
3. Methodology
The results presented in this paper are based on a bibliometric analysis of articles published between 2000 and 2022 on bioH2 production from microalgae. The analysis uses information from the published literature to answer the research questions, find research trends, identify research gaps, and identify future research directions. In addition, a scoping review of highly cited articles and recent publications helped outline recent advancements. Figure 2 shows the framework for the bibliometric analysis.
3.1. Data Collection
Scopus was chosen as the database for the present study. Scopus is one of the popular abstracts and citation databases of peer-reviewed literature. It has stringent quality criteria for indexation as well as more inclusive journals. It is the most commonly used database and covers various subjects [42]. The search query targeted publications from January 2000 until November 2022. This time frame was selected because most publications happened during this period, while only three journal articles related to bioH2 production using microalgae were published between 1995 and 1999. The data were extracted on the 22nd of November, 2022. The search string performed in Scopus was based on “title”. The search query was as follows: (“Microalgae” OR “Micro-algae” OR “Green algae” OR “Alga*”) AND (“Hydrogen” OR “Biohydrogen” OR “Bio-hydrogen” OR “BioH2” OR “Bio-H2” OR “H2”). To further limit the search, only articles written in English were considered. The initial search string produced 385 articles. Further screening and filtering to remove non-relevant and duplicate articles resulted in a total of 184 articles (a list of articles included in the analysis is included in Supplementary Materials S1).
3.2. Software Selection
Many software tools such as VOSviewer, CitNetExplorer, CiteSpace, and Bibliometrix support bibliometric analysis. VOSviewer is designed to address the graphical representation of bibliometric maps. These maps illustrate the structural and dynamic aspects of the research in an easy-to-interpret manner [42,43]. Data from Scopus was imported into VOS viewer (Visualization of Similarities) (version 1.6.18.0) and Microsoft Excel (version 16.0.11929.20288) for visualization and quantitative results.
3.3. Data Analysis
The Scopus database was employed to elucidate articles with the highest citations, identify publication trends, and extract the keywords. The evaluation of journals and countries was based on total publications (TP), total citations (TC), and the citations per publication (CPP). TP can indicate a journal or country’s contribution to the research area, while TC can represent the relevance and the quality of the papers published. CPP can assess the consistency of contribution to knowledge. Moreover, for the top authors, the ranking was performed using a normalized citation score. It is the total number of citations of a particular publication in relation to the average number of citations of all the publications in the same year. Leading articles were ranked using local and global citation scores. Global citations are the total number of citations across all indexing databases, whereas local citations are those inside the selected 184 articles. Co-authorship between countries was highlighted by a visualization map to determine which research collaborations are the strongest. Co-citation analysis was performed by a network map. In addition, visualization of keywords and their number of occurrences was also carried out. Furthermore, a scoping review of articles was conducted to highlight recent advancements and identify future research recommendations.
4. Results
The results of the bibliometric study on bioH2 production by microalgae are presented in the following sections, which focus on the trend in publications, major keywords, expressive journals, key authors, most cited articles, and influential countries.
4.1. Publication and Citations Trends
Figure 3 shows insights into the growing interest of bioH2 production using microalgae by presenting the yearly distribution of 184 articles. Only six articles were published between 1990 and 1999. However, starting from 2000, a gradual increase in the number of articles was noted. The publication of articles was relatively low in the first decade, but from 2013 onwards, there was a considerable increase in the number of articles, reaching a peak of 21 in 2022. The increasing number of articles may be due to the increased attention on and advancements in bioH2 production using microalgae in recent years. However, there was a drastic decline between 2019 and 2020. This could be due to the SARS-CoV-2 virus pandemic and related lockdown, which resulted in the temporary closure of universities and institutes. The citation trend of the articles is represented by the line graph. The number of citations increased from 28 in 2004 to 996 in 2022. The results signify the strong interest in bioH2 production using microalgae due to the growing demand for renewable bioenergy.
4.2. Keyword Analysis
4.2.1. Most Used Keywords
Figure 4 shows the keyword network map generated by VOSviewer using co-occurrence analysis. A threshold value of 4 minimum occurrences was considered. It was observed that out of the 433 keywords, only 27 met the threshold. Expected keywords related to bioH2, such as “biohydrogen”, biohydrogen production”, “hydrogen” and “hydrogen production” were removed to focus on unexpected keywords. Five clusters were formed based on their similarities. The primary keyword in each cluster represents a particular area.
The main keyword “microalgae” in the green cluster links with several significant keywords related with techniques to produce bioH2 using microalgae, such as “dark fermentation” and “fermentation”. Other keywords present in the same cluster are “pretreatment” and “methane”, which are related to the fermentation technique. BioH2 production from microalgae by fermentation has considerably increased by applying various pretreatment technologies [25]. Both violet and blue clusters have keywords related to different techniques to produce bioH2 using microalgae, such as “supercritical gasification”, “pyrolysis”, and “gasification”. Similarly, “green algae” in the red cluster links with different themes, such as “H2ase”, “sulfur deprivation”, “photobioreactor”, and “photosynthesis”, which are the other factors affecting bioH2 production using microalgae [15]. Among these, H2ase and sulfur deprivation are critical factors. Several studies focused on optimizing factors and techniques related to bioH2 production using microalgae [44,45]. The red and yellow cluster algae contain keywords related to microalgae strains such as “Platymonas subcordiformis” and “Chlamydomonas reinhardtii”. These two belong to the division Chlorophyta of green algae. Early research was more focused on Chlamydomonas reinhardtii [18,40]. It can be inferred that green, blue, and violet clusters are primarily associated with the techniques used to produce bioH2 using microalgae. The red and yellow clusters focus on the factors affecting bioH2 production and investigate microalgae strains.
The co-occurrence relation between two keywords is denoted by a curved line known as a “link.” The strength of the link is characterized by a number. A higher number indicates a stronger connection. Table 1 shows the above keywords and their total link strength. Microalgae were identified as the most occurring author’s keyword with the strongest link of 22. Biomass, pretreatment, and photobioreactor have similar occurrences and total link strength.
4.2.2. Emerging Keywords
Many keywords emerged in the last decade and have shown an increasing trend, as shown in Figure 5. Nitrogen deprivation had lower occurrences; however, a recent increase indicates it emerged as a critical factor in bioH2 yield. However, phosphorous deprivation has yet to be thoroughly explored. Several studies investigated the effects of different pretreatment methods such as autoclave, ultrasonication, ammonia, microwave, and electrolysis in H2 generation. These methods increase the solubilization and liberation of available organic material such as sugars, proteins, and lipids from algal cells, so H2 production performance is enhanced [15,46,47,48]. Dark fermentation showed a significantly increasing trend of occurrences, indicating the focus on achieving higher bioH2 yield by applying pretreatment techniques. Tetraspora sp. CU2551 had a significant increase even though it only appeared since 2018. Mixed microalgae consortia and co-fermentation showed similar growing behavior, indicating their strong correlation. Hydrothermal gasification has not advanced in bioH2 production compared to supercritical water gasification.
4.3. Major Contributors to the Field
The analysis revealed that the top journals, based on total publications (TP), are the International Journal of Hydrogen Energy (Int. J. Hydrog. Energy), Bioresource Technology, Biotechnology for Biofuels, Energy, and Fuel with a TP of 47, 17, 7, 5, and 4, respectively. Similarly, in terms of total citations (TC), these journals have the highest rankings after Energy and appeared in the top 10, as shown in Table 2. Int. J. Hydrog. Energy had the highest number of publications during the years 2014 and 2021, with 37 articles out of 47 total publications. The most cited articles in Int. J. Hydrog. Energy discuss the capability of new microalgal strains from freshwater and brackish water, such as Chlamydomonas noctigama and Chlamydomonas Euryale. They produced significant bioH2 yield under anaerobic conditions and sulfur deprivation. However, none of the new strains gave higher efficiency than the traditional strain Chlamydomonas reinhardtii. Furthermore, a laboratory bioreactor which successfully studied bioH2 production was developed and implemented. The important consideration during bioreactor construction was selecting the material used to avoid toxic effects [49]. The following article evaluated the impact of different pretreatment methods on the H2 fermentation of microalgae consortia. Methods such as autoclaving, ultrasonication, and electrolysis were applied on mixed microalgae consortia composed of Scenedesmus and Chlorella species [48]. Another article proposed an enhanced integrated system for simultaneous bioH2 production, storage, and power generation [9]. The most cited article in Bioresource Technology covered supercritical water gasification of Chlorella vulgaris, Spirulina platensis, and Saccharina latissima in the presence and absence of sodium hydroxide and nickel supported on alumina. BioH2 production more than doubled in the presence of sodium hydroxide, compared to in its absence [50]. Although the journals Plant Physiology, Planta, and J. Biological Chemistry have high TC and CPP, they had relatively lower TP compared to other journals. The most cited article in Plant Physiology describes the photobiological H2 gas production from green alga Chlamydomonas reinhardtii. This was the first work that showed the possibility to produce and accumulate significant volumes of H2 gas using Chlamydomonas reinhardtii for several days. The study outlines a unique method for sustained photobiological generation of H2 gas by reversible hydrogenase pathway [51]. The most cited article in Planta discusses the biochemical and morphological characteristics of Chlamydomonas reinhardtii [52].
An outline of the most influential countries is presented in Table 3, with a minimum number of 5 publications. China and the USA have the highest number of articles and citations. China is the world’s biggest polluter and its biggest green energy investor. China initially started publishing in 2004, and after that, there was a gradual increase in TP addressing various aspects of bioH2 production using microalgae, such as different techniques, optimization of factors, alternate strains, and pretreatments. Although Thailand is ranked third for TP with 17 publications, it is not ranked in the top ten of TC and CPP. There was a four-year publishing gap over the period from 2010 to 2015. However, from 2017 until 2022, there have been continuous publications. Different microalgae strains with successful bioH2 production rates were isolated [28,53,54]. The most cited article by USA shows the possibility of producing and accumulating significant volumes of H2 gas using Chlamydomonas reinhardtii for several days. The study outlines a unique method for sustained photobiological generation of H2 gas by reversible hydrogenase pathway [51]. Both USA and China established a series of programs supporting the research and development of bioenergy technologies. The diverse reasons behind these programs include energy security and independence, climate change, sustainability, and economic development. Lastly, the most cited work by Germany characterized the interaction between photosynthetic electron transport and bioH2 production in green algae, where it was demonstrated that photosynthetic ferredoxin is essential for efficient electron transfer between PSI and hydrogenase HydA1. The in-between electrostatic interaction process and electron transfer was performed by site-directed mutagenesis [55].
In addition, the analysis of the top authors provides a better understanding of their expertise in specific topics of bioH2 production from algae. VOS viewer was used to determine the top authors from the 184-node network analyzed. A minimum number of publications per author was set to 5, and hence 12 authors out of the 687 met that threshold. The top 10 were filtered based on the normalized citations and are presented in Table 4. From the normalized citation score, the top authors are Jerry D Murphy, Ao Xia, Maria Lucia Ghirardi, Jie Cheng, and Lin Zhengyan. These five authors are considered the most prominent in bioH2 production using microalgae. Table 5 shows the top 10 articles in terms of global and local citations.
The top three impactful articles ranked by global citations are the same as the top three articles ranked by local citations. Kruse et al. [56] developed a new approach to enhance bioH2 production in engineered Chlamydomonas cells by increasing proton (H+) and electron supply to the hydrogenase enzyme. The bioH2 production rates were 5–13-fold higher than those of the wild-type strains. Antal et al. [57] demonstrate that any change in PSII activity of Chlamydomonas reinhardtii deprived of sulfur is characterized by dramatic changes during cell adaptation and nutrient stress. The main factor which controls the photochemical activity of PSII and bioH2 production under sulfur deprivation was the reduced state of the plastoquinone pool. It regulates the remaining water-splitting capacity of PSII and the electron transport to hydrogenase. Onwudili et al. [50] evaluated Chlorella vulgaris, Spirulina platensis, and Saccharina latissimi. These three were processed under specific supercritical water gasification conditions. The bioH2 gas yields were more than two times higher in sodium hydroxide (NaOH) presence than in its absence. Xia et al. [2] assessed the co-fermentation of micro and macroalgae to improve the performance of bioH2 production. Guan et al. [58] demonstrated the photobiological bioH2 production by a marine green alga, Platymonas subcordiformis. The dependence of bioH2 production on sulfur deprivation revealed that bioH2 was greatly enhanced 13-fold when sulfur was deprived from the medium. This result suggests that sulfur plays a critical role in the production of bioH2 evolution. An increase in bioH2 production was obtained when the medium pH was greater than 5. A similar observation was made by Maneeruttanarungroj et al. [28]. A novel unicellular H2-producing green alga belonging to family Tetraspora was isolated from a freshwater pond in Thailand. BioH2 yield was increased with increased pH from 5.75 to 9.30, and using a medium lacking both nitrogen and sulfur resulted in about a 50% increase in the bioH2 yield.
4.4. Nature of Collaboration
Country authorship analysis was conducted with a minimum of five co-authored publications per country. Out of the 47 identified countries, 18 met the criterion, as presented in Figure 6. The round nodes represent the total number of publications for each country. The larger the number of publications, the bigger the size of the bubble. The co-authorship among these countries is depicted by the curved lines that connect one country to another. Moreover, the thickness of the lines illustrates the term recurrence between the countries. Notable collaborations exist between China and Ireland, with a link strength of 8. The collaborative articles between these two countries mainly discuss co-fermentation and novel pretreatment methods. The second highest collaboration was between China and Australia and China and Japan, with a link strength of 5. The rest of the countries exhibit low co-authorship of 3 links or less. Most countries (such as China, the USA, and India) which focus on bioH2 production had total GHG emissions of more than 14 gigatons of CO2 equivalent (GtCO2e), 5 GtCO2e, and 3 GtCO2e in 2020 [63].
Furthermore, a co-authorship map was created between the authors, with the minimum number of documents being 3. This resulted in 53 authors meeting this criterion out of a total of 687. However, 33 authors were excluded from the analysis as no links were found among them. The most significant collaboration was between Mei Zhao (Jiangnan University, China) and Ruan Wunquan (Jiangnan University, China), Maria Lucia Ghirardi (National Renewable Energy Laboratory, USA) and Michael Seibert (National Renewable Energy Laboratory, USA), and Valeria Nagy (Institute of Plant Biology, Hungary) and Szilvia Z Toth (Institute of Plant Biology, Hungary). All the co-authorship was among authors from the same university. This hints that even though collaborations exist between countries, as shown in Figure 6, these are between different authors, not amounting to the threshold of 3 co-authored publications.
4.5. Co-Citation Analysis
Bibliometric citation analysis uses various techniques, such as co-authorship, bibliometric coupling-citation, co-word, and co-occurrence analysis. Co-citation analysis expresses the frequency of two or more articles cited by other articles. It gives different research themes and aids in finding the origin and direction for future research in the research area. The co-citation networks of bioH2 production using microalgae are visualized in Figure 7. It is significant to note that some articles repeat more than once, which features the presence of interconnection between them. The clustering network was created by VOSviewer. A cutoff value of six co-citations was established to select the most influential papers, resulting in the 18 articles shown in Figure 7 [64]. The bubble size represents the number of citations of the articles and the strength of co-citations presented by the thickness of the lines between nodes. Nodes were labeled by the authors’ names and article title. However, the maximum size of the label was restricted to 30 characters. The five most co-cited articles are discussed below.
Zhang et al. [52] evaluated the biochemical and morphological characteristics of Chlamydomonas reinhardtii. At the same time, Melis et al. [51] described a unique method for sustained photobiological generation of H2 gas from the green alga Chlamydomonas reinhardtii by preventing the severe O2 sensitivity of the enzyme H2ase. Melis [65] summarized the recent advances and directions for future development in the bioH2 metabolism of unicellular green algae. Furthermore, potential practical applications of bioH2 and the biochemistry of anaerobic bioH2 photoproduction exist. Kosourov et al. [66] demonstrated the effects of the addition of sulfur to a sulfur-depleted medium in the case of bioH2 by Chlamydomonas reinhardtii cells in a self-made automated photobioreactor system. The bioH2 production was optimized by a controlled amount of sulfur in the medium at the time of sulfur deprivation. However, the addition of excess sulfur delayed the onset of bioH2 production and lowered the final yield of H2. Hallenbeck et al. [67] analyzed the various techniques used in bioH2 production from microalgae and identified critical limiting factors. They also discussed dark fermentation in detail. Gaffron et al. [68] demonstrated the capability of Scenedesmus to liberate H2 slowly in the dark when the surrounding air is replaced by nitrogen. They also showed the increase in bioH2 liberation by the illumination of fermenting algae in the absence of CO2 and H2.
5. Discussion of Recent Developments
Highlighting recent developments in the field could help overcome difficulties in the research area. This would consequently help identify the research opportunities and future research directions. Table 6 groups the advancements into four main themes: (1) techniques to produce bioH2 using microalgae, (2) optimization of factors affecting bioH2 production using microalgae, (3) alternative and potential micro-algal species for bioH2 production, and (4) strategies to improve the bioH2 production from microalgae.
Some researchers have illustrated that bioH2 production can be dramatically enhanced with the presence of metal and metal oxide nanoparticles like nickel oxide (NiO) and iron oxide (Fe2O3) [24,69]. Co-fermentation of pretreated rice residue and microalgae in a mix ratio of 5:1 showed a 10.7-fold higher yield than fermentation of pretreated microalgae alone. The addition of glucose almost doubled the bioH2 yield of Chlorella pyrenoidosa. The exogenic glucose acts as an electron donor for hydrogenases [104]. Co-fermentation of Lyngbya limnetica and glucose for bioH2 production has been explored. The elements such as iron, cobalt, and nickel are the main components of nutrient media. Therefore, the presence of nanocatalysts based on these elements fastens the growth of fermentative microorganisms, subsequently increasing bioH2 production [69]. Future research can focus on optimizing operational factors, like sludge/algae ratio, initial pH, and temperature. Furthermore, magnetite (Fe3O4) was shown to be an effective supplement as it resulted in higher bioH2 production by Clostridium butyricum DSM 10702 under anaerobic fermentation conditions, even when subjected to the inhibitory conditions of mixed sugar composition and various concentrations of toxic materials [73]. Similarly, results of nZVI dosing in a dark co-fermentation system showed that the cumulative H2 increased by 29.20% [74]. Since non-catalytic gasification has a low reaction rate, low efficiency, and high carbon monoxide fractions, recent research investigated the ability of catalytic gasification to decrease the reaction temperature while maintaining the same gasification efficiency. The most prominent methods among that line of research include utilizing SCWG with Nickel (Ni)/zeolite catalysts [78], Ni catalyst and Yttrium oxide Y2O3, catalytic and non-catalytic gasification with a hybrid functional mixture of eggshell-derived calcium oxide (CaO) [79], and catalytic hydrothermal gasification using noble metals platinum (Pt) and ruthenium (Ru) [35].
Optimization of parameters can be acheived by technology, such as modeling of reaction kinetics via an empirical logistic model and Aspen Plus V11, a commercial process simulator, to optimize the essential process parameters such as time, sulfur concentrations, and the pH of the medium [89], and application of Gibbs energy minimization and entropy maximization methodologies using GAMS software for optimization of conditions for supercritical gasification [45]. One recent article in 2022 studied the ability of Parachlorella kessleri RA-002 to generate bioH2 in two different media with and without nitrogen deprivation. The highest H2 yield was found during algae growth under nitrogen-deprived conditions, which was 4–5 times higher than in cells cultivated without nitrogen deprivation [88]; the potassium deprivation condition is also a promising choice to enhance bioH2 production in biological systems [87].
Around 200,000–800,000 species in various algae genera have been discovered so far [25]. Chlamydomonas reinhardtii was the most investigated microalgae for bioH2 production using microalgae [18,40]. After 2017, the researchers focused more on Chlorella sp. [105,106]. Scenedesmus sp. and Tetraspora sp. are the other microalgae strains explored after Chlorella sp. [87,88,107]. Duangjan et al. [108] revealed that under conditions of nutrient deficiency, the order Chlorellales and Volvocales could produce bioH2. The genus Chlorella and Chlamydomonas belong to these orders, respectively. Furthermore, there are studies based on microalgal modifications for enhanced bioH2 yield. For instance, a new PSI-H2ase chimera polypeptide expressed in a Chlamydomonas reinhardtii strain lacking endogenous H2ases drastically diminished CO2 fixation and O2 scavenging and allowed bioH2 production for at least four days [93]; the possibility of H2 generation from the diatoms by culturing it in various solar panel photobioreactors [94] and Tris-acetate-phosphate medium (TAP) with 0.2 mg/L of Co to improve the growth and biomass in C. reinhardtii cultures; and subsequently, biofuel generation [109].
The concentrations of disintegration degree and soluble chemical oxygen demand increased after high-pressure homogenization pretreatment of Taihu lake wet algae. It enhanced bioH2 generation from pre-treated wet algae and food waste by anaerobic co-digestion [3]. Maswanna et al. [12] observed that immobilized cells of Tetraspora sp. CU2551 in an alginate matrix and under aerobic sulfur deprivation could produce maximum bioH2 production. The calcium alginate gels restricted the diffusion of O2 to the H2ase, further enhancing bioH2 production. It could also produce H2 when the medium was refreshed for up to six cycles over 43 days. Another study by the same authors on immobilized cells of Tetraspora sp. CU2551 in an alginate matrix and under anaerobic sulfur deprivation results in enhanced H2 production and shortened incubation time compared to other microalgae such as Cyanobacteria, Chlamydomonas reinhardtii CC-124, and Anabaena PCC 7120 under the same conditions [13]. From this brief discussion, some research opportunities can be identified as follows:
- Designing customized photobioreactors for investigating direct biophotolysis.
- Developing techniques to prevent the inhibition of biophotolysis by limiting high light intensity and O2 formation.
- Assessing the possibility of limiting the amount of accumulated biomass during the growth phase and improving light transformation efficiency in indirect biophotolysis.
- Investigating alternative methods to improve substrate transformation efficiency and H2–CO2 mixture separation and to control O2 accumulation during dark fermentation.
- Improving H2 conversion efficiency, light transformation efficiency, and control inhomogeneity in the light distribution in photo fermentation.
- Exploring different strains of microalgae for higher bioH2 yield.
6. Conclusions and Future Directions
The bibliometric analysis revealed important findings regarding research in the field of bioH2 production from microalgae. Results revealed that the number of publications increased from three in 2000 to 996 in 2022. The identified prominent research themes focused on investigating the factors affecting the process efficiency and on the impact of using different microalgae strains. Moreover, the analysis highlighted key emerging trends, the most compelling of which included “pretreatment”, “supercritical water gasification”, and “dark fermentation”. It was also revealed that China, USA, Thailand, India, and Australia are the leading countries on this research topic. The greatest collaboration was between China and Ireland. The articles from the collaboration between these countries mainly discuss co-fermentation and novel pretreatment methods. All the co-authorship was among authors from the same university/country, which indicates that the number of international collaborations between countries is low.
The identified recent developments and research gaps will aid future research to better advance bioH2 production from microalgae. The following are the key recommendations that can be adopted for future studies:
- More investigations on identifying genetic strategies to reduce the O2 sensitivity of the H2ase enzyme.
- Investigations to find different co-culture methods, such as algae-bacteria consortium for a continuous bioH2 yield.
- Emphasis on reducing the operation cost and realistic pilot studies for scaling up the process.
- Investigations to find effective pretreatment combinations.
- Studying the potential of nanoparticles to enhance bioH2 yield.
- Focusing on Life Cycle Assessments to scale up microalgal bioH2 production.
- Exploring the possibilities to incorporate genetic engineering.
This study might be affected by some limitations since the analysis only included publications after the year 2000. This may have resulted in excluding earlier contributions to the research area. Moreover, as the search focused on the occurrence of the keywords in the “title” rather than “all fields”, this could have resulted in missing some relevant articles from the analysis. Limiting the document type to only journal articles written in English and not including conference papers or book chapters might have eliminated some relevant publications from the analysis; however, novel ideas are usually presented in journal articles.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15010185/s1, S1: List of 184 articles included in the bibliometric analysis.
Author Contributions
Conceptualization, S.P.A., M.A.J., M.A.H., and A.A.H.; methodology, S.P.A., M.A.J., M.A.H., and A.A.H.; software, S.P.A. and M.A.H.; formal analysis, S.P.A., M.A.J., M.A.H., and A.A.H.; investigation, S.P.A., M.A.J., M.A.H., and A.A.H.; resources, M.A.H. and A.A.H.; data curation, S.P.A. and M.A.J.; writing—original draft preparation, S.P.A.; writing—review and editing, S.P.A., M.A.J., M.A.H., and A.A.H.; visualization, S.P.A., M.A.J., M.A.H., A.A.H., and M.E.A.; supervision, M.A.J., M.A.H., and A.A.H.; project administration, M.A.H. and A.A.H.; funding acquisition, M.A.H. and A.A.H. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge the financial support from the National Water and Energy Center at U.A.E. University under Grant Numbers G00003296 and G00003501.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting this study’s findings are available on request from the corresponding author.
Conflicts of Interest
The authors have no conflict of interest to declare.
References
- Ampese, L.C.; Sganzerla, W.G.; Di Domenico Ziero, H.; Mudhoo, A.; Martins, G.; Forster-Carneiro, T. Research Progress, Trends, and Updates on Anaerobic Digestion Technology: A Bibliometric Analysis. J. Clean. Prod. 2022, 331, 130004. [Google Scholar] [CrossRef]
- Xia, A.; Jacob, A.; Tabassum, M.R.; Herrmann, C.; Murphy, J.D. Production of Hydrogen, Ethanol and Volatile Fatty Acids through Co-Fermentation of Macro- and Micro-Algae. Bioresour. Technol. 2016, 205, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Xu, J.; Xue, H.; Li, C.; Liu, H.; Gu, S.; Miao, H.; Ruan, W. Improving Hydrogen Recovery from Anaerobic Co-Digestion of Algae and Food Waste by High-Pressure Homogenisation Pre-Treatment. Environ. Chem. Lett. 2021, 19, 3497–3504. [Google Scholar] [CrossRef]
- Srirangan, K.; Pyne, M.E.; Perry Chou, C. Biochemical and Genetic Engineering Strategies to Enhance Hydrogen Production in Photosynthetic Algae and Cyanobacteria. Bioresour. Technol. 2011, 102, 8589–8604. [Google Scholar] [CrossRef] [PubMed]
- Vanyan, L.; Cenian, A.; Trchounian, K. Biogas and Biohydrogen Production Using Spent Coffee Grounds and Alcohol Production Waste. Energies 2022, 15, 5935. [Google Scholar] [CrossRef]
- Jensen, L.S.; Kaul, C.; Juncker, N.B.; Thomsen, M.H.; Chaturvedi, T. Biohydrogen Production in Microbial Electrolysis Cells Utilizing Organic Residue Feedstock: A Review. Energies 2022, 15, 8396. [Google Scholar] [CrossRef]
- Tamburic, B.; Zemichael, F.W.; Maitland, G.C.; Hellgardt, K. Parameters Affecting the Growth and Hydrogen Production of the Green Alga Chlamydomonas reinhardtii. Int. J. Hydrog. Energy 2011, 36, 7872–7876. [Google Scholar] [CrossRef]
- Gholkar, P.; Shastri, Y.; Tanksale, A. Renewable Hydrogen and Methane Production from Microalgae: A Techno-Economic and Life Cycle Assessment Study. J. Clean. Prod. 2021, 279, 123726. [Google Scholar] [CrossRef]
- Aziz, M. Integrated Hydrogen Production and Power Generation from Microalgae. Int. J. Hydrog. Energy 2016, 41, 104–112. [Google Scholar] [CrossRef]
- Shobana, S.; Saratale, G.D.; Pugazhendhi, A.; Arvindnarayan, S.; Periyasamy, S.; Kumar, G.; Kim, S.-H. Fermentative Hydrogen Production from Mixed and Pure Microalgae Biomass: Key Challenges and Possible Opportunities. Int. J. Hydrog. Energy 2017, 42, 26440–26453. [Google Scholar] [CrossRef]
- Raheem, A.; Ji, G.; Memon, A.; Sivasangar, S.; Wang, W.; Zhao, M.; Taufiq-Yap, Y.H. Catalytic Gasification of Algal Biomass for Hydrogen-Rich Gas Production: Parametric Optimization via Central Composite Design. Energy Convers. Manag. 2018, 158, 235–245. [Google Scholar] [CrossRef]
- Maswanna, T.; Lindblad, P.; Maneeruttanarungroj, C. Improved Biohydrogen Production by Immobilized Cells of the Green Alga Tetraspora sp., CU2551 Incubated under Aerobic Conditions. J. Appl. Phycol. 2020, 32, 2937–2945. [Google Scholar] [CrossRef]
- Maswanna, T.; Phunpruch, S.; Lindblad, P.; Maneeruttanarungroj, C. Enhanced Hydrogen Production by Optimization of Immobilized Cells of the Green Alga Tetraspora sp., CU2551 Grown under Anaerobic Condition. Biomass Bioenergy 2018, 111, 88–95. [Google Scholar] [CrossRef]
- Vargas, S.R.; Santos, P.V.D.; Giraldi, L.A.; Zaiat, M.; Calijuri, M.d.C. Anaerobic Phototrophic Processes of Hydrogen Production by Different Strains of Microalgae Chlamydomonas sp. FEMS Microbiol. Lett. 2018, 365, fny073. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, S.F.; Mofijur, M.; Nahrin, M.; Chowdhury, S.N.; Nuzhat, S.; Alherek, M.; Rafa, N.; Ong, H.C.; Nghiem, L.D.; Mahlia, T.M.I. Biohydrogen Production from Wastewater-Based Microalgae: Progresses and Challenges. Int. J. Hydrog. Energy 2021, 47, 37321–37342. [Google Scholar] [CrossRef]
- Rahmani, A.; Zerrouki, D.; Djafer, L.; Ayral, A. Hydrogen Recovery from the Photovoltaic Electroflocculation-Flotation Process for Harvesting Chlorella pyrenoidosa Microalgae. Int. J. Hydrog. Energy 2017, 42, 19591–19596. [Google Scholar] [CrossRef]
- Alalayah, W.M.; Al-Zahrani, A.; Edris, G.; Demirbas, A. Kinetics of Biological Hydrogen Production from Green Microalgae Chlorella vulgaris using Glucose as Initial Substrate. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 1210–1215. [Google Scholar] [CrossRef]
- Yagi, T.; Yamashita, K.; Okada, N.; Isono, T.; Momose, D.; Mineki, S.; Tokunaga, E. Hydrogen Photoproduction in Green Algae Chlamydomonas reinhardtii Sustainable over 2 Weeks with the Original Cell Culture without Supply of Fresh Cells nor Exchange of the Whole Culture Medium. J. Plant Res. 2016, 129, 771–779. [Google Scholar] [CrossRef]
- Das, A.A.K.; Esfahani, M.M.N.; Velev, O.D.; Pamme, N.; Paunov, V.N. Artificial Leaf Device for Hydrogen Generation from Immobilised C. reinhardtii Microalgae. J. Mater. Chem. A 2015, 3, 20698–20707. [Google Scholar] [CrossRef] [Green Version]
- Oncel, S.S.; Kose, A.; Faraloni, C.; Imamoglu, E.; Elibol, M.; Torzillo, G.; Vardar Sukan, F. Biohydrogen Production from Model Microalgae Chlamydomonas reinhardtii: A Simulation of Environmental Conditions for Outdoor Experiments. Int. J. Hydrog. Energy 2015, 40, 7502–7510. [Google Scholar] [CrossRef]
- Chen, Y. Global Potential of Algae-Based Photobiological Hydrogen Production. Energy Environ. Sci. 2022, 15, 2843–2857. [Google Scholar] [CrossRef]
- Faraji, M.; Saidi, M. Hydrogen-Rich Syngas Production via Integrated Configuration of Pyrolysis and Air Gasification Processes of Various Algal Biomass: Process Simulation and Evaluation Using Aspen Plus Software. Int. J. Hydrog. Energy 2021, 46, 18844–18856. [Google Scholar] [CrossRef]
- Adnan, M.A.; Xiong, Q.; Muraza, O.; Hossain, M.M. Gasification of Wet Microalgae to Produce H2-Rich Syngas and Electricity: A Thermodynamic Study Considering Exergy Analysis. Renew. Energy 2020, 147, 2195–2205. [Google Scholar] [CrossRef]
- Li, S.; Li, F.; Zhu, X.; Liao, Q.; Chang, J.-S.; Ho, S.-H. Biohydrogen Production from Microalgae for Environmental Sustainability. Chemosphere 2022, 291, 132717. [Google Scholar] [CrossRef]
- Musa Ardo, F.; Wei Lim, J.; Ramli, A.; Kee Lam, M.; Kiatkittipong, W.; Alaaeldin Abdelfattah, E.; Kashif Shahid, M.; Usman, A.; Wongsakulphasatch, S.; Tasnim Sahrin, N. A Review in Redressing Challenges to Produce Sustainable Hydrogen from Microalgae for Aviation Industry. Fuel 2022, 330, 125646. [Google Scholar] [CrossRef]
- Sridhar, A.; Ponnuchamy, M.; Senthil Kumar, P.; Kapoor, A.; Xiao, L. Progress in the Production of Hydrogen Energy from Food Waste: A Bibliometric Analysis. Int. J. Hydrog. Energy 2022, 47, 26326–26354. [Google Scholar] [CrossRef]
- Sillero, L.; Sganzerla, W.G.; Forster-Carneiro, T.; Solera, R.; Perez, M. A Bibliometric Analysis of the Hydrogen Production from Dark Fermentation. Int. J. Hydrog. Energy 2022, 47, 27397–27420. [Google Scholar] [CrossRef]
- Maneeruttanarungroj, C.; Lindblad, P.; Incharoensakdi, A. A Newly Isolated Green Alga, Tetraspora sp., CU2551, from Thailand with Efficient Hydrogen Production. Int. J. Hydrog. Energy 2010, 35, 13193–13199. [Google Scholar] [CrossRef]
- Mona, S.; Kumar, S.S.; Kumar, V.; Parveen, K.; Saini, N.; Deepak, B.; Pugazhendhi, A. Green Technology for Sustainable Biohydrogen Production (Waste to Energy): A Review. Sci. Total Environ. 2020, 728, 138481. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Li, Q.; Wang, S.; Wang, L.; Liu, H.; Fan, C. Engineering a Chemoenzymatic Cascade for Sustainable Photobiological Hydrogen Production with Green Algae. Energy Environ. Sci. 2020, 13, 2064–2068. [Google Scholar] [CrossRef]
- Dalena, F.; Senatore, A.; Tursi, A.; Basile, A. Bioenergy Production from Second- and Third-Generation Feedstocks. In Bioenergy Systems for the Future; Elsevier: Amsterdam, The Netherlands, 2017; pp. 559–599. ISBN 978-0-08-101031-0. [Google Scholar]
- Goswami, R.K.; Mehariya, S.; Obulisamy, P.K.; Verma, P. Advanced Microalgae-Based Renewable Biohydrogen Production Systems: A Review. Bioresour. Technol. 2021, 320, 124301. [Google Scholar] [CrossRef] [PubMed]
- Javed, M.A.; Zafar, A.M.; Aly Hassan, A.; Zaidi, A.A.; Farooq, M.; El Badawy, A.; Lundquist, T.; Mohamed, M.M.A.; Al-Zuhair, S. The Role of Oxygen Regulation and Algal Growth Parameters in Hydrogen Production via Biophotolysis. J. Environ. Chem. Eng. 2022, 10, 107003. [Google Scholar] [CrossRef]
- Show, K.-Y.; Yan, Y.; Zong, C.; Guo, N.; Chang, J.-S.; Lee, D.-J. State of the Art and Challenges of Biohydrogen from Microalgae. Bioresour. Technol. 2019, 289, 121747. [Google Scholar] [CrossRef] [PubMed]
- Jiao, J.-L.; Wang, F.; Duan, P.-G.; Xu, Y.-P.; Yan, W.-H. Catalytic Hydrothermal Gasification of Microalgae for Producing Hydrogen and Methane-Rich Gas. Energy Sources Part A Recovery Util. Environ. Eff. 2017, 39, 851–860. [Google Scholar] [CrossRef]
- Kumar, M.; Oyedun, A.O.; Kumar, A. A Parametric Study through the Modelling of Hydrothermal Gasification for Hydrogen Production from Algal Biomass. Can. J. Chem. Eng. 2021, 99, S39–S54. [Google Scholar] [CrossRef]
- Demirbas, A. Hydrogen from Mosses and Algae via Pyrolysis and Steam Gasification. Energy Sources Part A Recovery Util. Environ. Eff. 2009, 32, 172–179. [Google Scholar] [CrossRef]
- Chader, S.; Mahmah, B.; Chetehouna, K.; Amrouche, F.; Abdeladim, K. Biohydrogen Production Using Green Microalgae as an Approach to Operate a Small Proton Exchange Membrane Fuel Cell. Int. J. Hydrog. Energy 2011, 36, 4089–4093. [Google Scholar] [CrossRef] [Green Version]
- Chatzitakis, A.; Nikolakaki, E.; Sotiropoulos, S.; Poulios, I. Hydrogen Production Using an Algae Photoelectrochemical Cell. Appl. Catal. B Environ. 2013, 142–143, 161–168. [Google Scholar] [CrossRef]
- Saifuddin, N.; Ong, M.Y.; Priatharsini, P. Optimization of Photosynthetic Hydrogen Gas Production by Green Alga in Sulfur Deprived Condition. Indian J. Sci. Technol. 2016, 9, 93390. [Google Scholar] [CrossRef]
- Mujalin Pholchan, K.K. Effect of Light Intensities and Atmospheric Gas Conditions on Biohydrogen Production of Microalgae Isolated from Fisheries Wastewater. Environ. Nat. Resour. J. 2017, 15, 21–29. [Google Scholar] [CrossRef]
- Raman, R.; Nair, V.K.; Prakash, V.; Patwardhan, A.; Nedungadi, P. Green-Hydrogen Research: What Have We Achieved, and Where Are We Going? Bibliometrics Analysis. Energy Rep. 2022, 8, 9242–9260. [Google Scholar] [CrossRef]
- Aria, M.; Cuccurullo, C. Bibliometrix: An R-Tool for Comprehensive Science Mapping Analysis. J. Informetr. 2017, 11, 959–975. [Google Scholar] [CrossRef]
- Nazarpour, M.; Taghizadeh-Alisaraei, A.; Asghari, A.; Abbaszadeh-Mayvan, A.; Tatari, A. Optimization of Biohydrogen Production from Microalgae by Response Surface Methodology (RSM). Energy 2022, 253, 124059. [Google Scholar] [CrossRef]
- Gomes, J.G.; Mitoura, J.; Guirardello, R. Thermodynamic Analysis for Hydrogen Production from the Reaction of Subcritical and Supercritical Gasification of the C. vulgaris Microalgae. Energy 2022, 260, 125030. [Google Scholar] [CrossRef]
- Cheng, J.; Yue, L.; Ding, L.; Li, Y.-Y.; Ye, Q.; Zhou, J.; Cen, K.; Lin, R. Improving Fermentative Hydrogen and Methane Production from an Algal Bloom through Hydrothermal/Steam Acid Pretreatment. Int. J. Hydrog. Energy 2019, 44, 5812–5820. [Google Scholar] [CrossRef]
- Wang, Q.; Gong, Y.; Liu, S.; Wang, D.; Liu, R.; Zhou, X.; Nghiem, L.D.; Zhao, Y. Free Ammonia Pretreatment To Improve Bio-Hydrogen Production from Anaerobic Dark Fermentation of Microalgae. ACS Sustain. Chem. Eng. 2019, 7, 1642–1647. [Google Scholar] [CrossRef]
- Kumar, G.; Sivagurunathan, P.; Thi, N.B.D.; Zhen, G.; Kobayashi, T.; Kim, S.-H.; Xu, K. Evaluation of Different Pretreatments on Organic Matter Solubilization and Hydrogen Fermentation of Mixed Microalgae Consortia. Int. J. Hydrog. Energy 2016, 41, 21628–21640. [Google Scholar] [CrossRef]
- Skjånes, K.; Knutsen, G.; Källqvist, T.; Lindblad, P. H2 Production from Marine and Freshwater Species of Green Algae during Sulfur Deprivation and Considerations for Bioreactor Design. Int. J. Hydrog. Energy 2008, 33, 511–521. [Google Scholar] [CrossRef]
- Onwudili, J.A.; Lea-Langton, A.R.; Ross, A.B.; Williams, P.T. Catalytic Hydrothermal Gasification of Algae for Hydrogen Production: Composition of Reaction Products and Potential for Nutrient Recycling. Bioresour. Technol. 2013, 127, 72–80. [Google Scholar] [CrossRef]
- Melis, A.; Zhang, L.; Forestier, M.; Ghirardi, M.L.; Seibert, M. Sustained Photobiological Hydrogen Gas Production upon Reversible Inactivation of Oxygen Evolution in the Green Alga Chlamydomonas reinhardtii. Plant Physiol. 2000, 122, 127–136. [Google Scholar] [CrossRef]
- Zhang, L.; Happe, T.; Melis, A. Biochemical and Morphological Characterization of Sulfur-Deprived and H2-Producing Chlamydomonas reinhardtii (Green Alga). Planta 2002, 214, 552–561. [Google Scholar] [CrossRef] [PubMed]
- Sirawattanamongkol, T.; Maswanna, T.; Maneeruttanarungroj, C. A Newly Isolated Green Alga Chlorella sp. KLSc59: Potential for Biohydrogen Production. J. Appl. Phycol. 2020, 32, 2927–2936. [Google Scholar] [CrossRef]
- Unpaprom, Y. A Newly Isolated Green Alga, Scenedesmus acuminatus, from Thailand with Efficient Hydrogen Production. Chiang Mai J. Sci. 2017, 44, 1270–1278. [Google Scholar]
- Winkler, M.; Kuhlgert, S.; Hippler, M.; Happe, T. Characterization of the Key Step for Light-Driven Hydrogen Evolution in Green Algae. J. Biol. Chem. 2009, 284, 36620–36627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kruse, O.; Rupprecht, J.; Bader, K.-P.; Thomas-Hall, S.; Schenk, P.M.; Finazzi, G.; Hankamer, B. Improved Photobiological H2 Production in Engineered Green Algal Cells. J. Biol. Chem. 2005, 280, 34170–34177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antal, T.K.; Krendeleva, T.E.; Laurinavichene, T.V.; Makarova, V.V.; Ghirardi, M.L.; Rubin, A.B.; Tsygankov, A.A.; Seibert, M. The Dependence of Algal H2 Production on Photosystem II and O2 Consumption Activities in Sulfur-Deprived Chlamydomonas einhardtii Cells. Biochim. Et Biophys. Acta (BBA) Bioenerg. 2003, 1607, 153–160. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Deng, M.; Yu, X.; Zhang, W. Two-Stage Photo-Biological Production of Hydrogen by Marine Green Alga Platymonas subcordiformis. Biochem. Eng. J. 2004, 19, 69–73. [Google Scholar] [CrossRef]
- Lakatos, G.; Deák, Z.; Vass, I.; Rétfalvi, T.; Rozgonyi, S.; Rákhely, G.; Ördög, V.; Kondorosi, É.; Maróti, G. Bacterial Symbionts Enhance Photo-Fermentative Hydrogen Evolution of Chlamydomonas Algae. Green Chem. 2014, 16, 4716–4727. [Google Scholar] [CrossRef]
- Duman, G.; Uddin, M.A.; Yanik, J. Hydrogen Production from Algal Biomass via Steam Gasification. Bioresour. Technol. 2014, 166, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-H.; Kim, H.-C.; Choi, J.-A.; Abou-Shanab, R.A.I.; Dempsey, B.A.; Regan, J.M.; Kim, J.R.; Song, H.; Nam, I.-H.; Kim, S.-N.; et al. Photoautotrophic Hydrogen Production by Eukaryotic Microalgae under Aerobic Conditions. Nat. Commun. 2014, 5, 3234. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, A.V.; Thomas-Hall, S.R.; Malnoë, A.; Timmins, M.; Mussgnug, J.H.; Rupprecht, J.; Kruse, O.; Hankamer, B.; Schenk, P.M. Transcriptome for Photobiological Hydrogen Production Induced by Sulfur Deprivation in the Green Alga Chlamydomonas reinhardtii. Eukaryot. Cell 2008, 7, 1965–1979. [Google Scholar] [CrossRef] [PubMed]
- United Nations Environment Programme. Emissions Gap Report 2022: The Closing Window—Climate Crisis Calls for Rapid Transformation of Societies. Nairobi. 2022. Available online: https://www.unep.org/emissions-Gap-Report-2022 (accessed on 1 November 2022).
- Leung, X.Y.; Sun, J.; Bai, B. Bibliometrics of Social Media Research: A Co-Citation and Co-Word Analysis. Int. J. Hosp. Manag. 2017, 66, 35–45. [Google Scholar] [CrossRef]
- Melis, A. Photosynthetic H2 Metabolism in Chlamydomonas reinhardtii (Unicellular Green Algae). Planta 2007, 226, 1075–1086. [Google Scholar] [CrossRef] [PubMed]
- Kosourov, S.; Tsygankov, A.; Seibert, M.; Ghirardi, M.L. Sustained Hydrogen Photoproduction by Chlamydomonas reinhardtii: Effects of Culture Parameters. Biotechnol. Bioeng. 2002, 78, 731–740. [Google Scholar] [CrossRef]
- Hallenbeck, P. Biological Hydrogen Production; Fundamentals and Limiting Processes. Int. J. Hydrog. Energy 2002, 27, 1185–1193. [Google Scholar] [CrossRef]
- Gaffron, H.; Rubin, J. Fermentative and Photochemical Production of Hydrogen in Algae. J. Gen. Physiol. 1942, 26, 219–240. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, N.; Srivastava, M.; Singh, R.; Syed, A.; Bahadur Pal, D.; Elgorban, A.M.; Kushwaha, D.; Mishra, P.K.; Gupta, V.K. Co-Fermentation of Residual Algal Biomass and Glucose under the Influence of Fe3O4 Nanoparticles to Enhance Biohydrogen Production under Dark Mode. Bioresour. Technol. 2021, 342, 126034. [Google Scholar] [CrossRef]
- Sun, C.; Xia, A.; Liao, Q.; Fu, Q.; Huang, Y.; Zhu, X.; Wei, P.; Lin, R.; Murphy, J.D. Improving Production of Volatile Fatty Acids and Hydrogen from Microalgae and Rice Residue: Effects of Physicochemical Characteristics and Mix Ratios. Appl. Energy 2018, 230, 1082–1092. [Google Scholar] [CrossRef]
- Xu, J.; Upcraft, T.; Tang, Q.; Guo, M.; Huang, Z.; Zhao, M.; Ruan, W. Hydrogen Generation Performance from Taihu Algae and Food Waste by Anaerobic Codigestion. Energy Fuels 2019, 33, 1279–1289. [Google Scholar] [CrossRef]
- Hwang, J.-H.; Lee, M.; Kang, E.H.; Lee, W.H. Renewable Algal Photo H2 Production without S Control Using Acetate Enriched Fermenter Effluents. Int. J. Hydrog. Energy 2021, 46, 1740–1751. [Google Scholar] [CrossRef]
- Kim, D.-H.; Yoon, J.-J.; Kim, S.-H.; Park, J.-H. Effect of Conductive Material for Overcoming Inhibitory Conditions Derived from Red Algae-Based Substrate on Biohydrogen Production. Fuel 2021, 285, 119059. [Google Scholar] [CrossRef]
- Zhao, M.; Liu, Z.; Xu, J.; Liu, H.; Dai, X.; Gu, S.; Ruan, W. Dosing Effect of Nano Zero Valent Iron (nZVI) on the Dark Hydrogen Fermentation Performance via Lake Algae and Food Waste Co-Digestion. Energy Rep. 2020, 6, 3192–3199. [Google Scholar] [CrossRef]
- Chang, K.-L.; Lin, Y.-C.; Shangdiar, S.; Chen, S.-C.; Hsiao, Y.-H. Hydrogen Production from Dry Spirulina Algae with Downstream Feeding in Microwave Plasma Reactor Assisted under Atmospheric Pressure. J. Energy Inst. 2020, 93, 1597–1601. [Google Scholar] [CrossRef]
- Nurdiawati, A.; Zaini, I.N.; Irhamna, A.R.; Sasongko, D.; Aziz, M. Novel Configuration of Supercritical Water Gasification and Chemical Looping for Highly-Efficient Hydrogen Production from Microalgae. Renew. Sustain. Energy Rev. 2019, 112, 369–381. [Google Scholar] [CrossRef]
- Nurdiawati, A.; Zaini, I.N.; Aziz, M. Efficient Hydrogen Production from Algae and Its Conversion to Methylcyclohexane. Chem. Eng. Trans. 2018, 70, 1507–1512. [Google Scholar] [CrossRef]
- Xie, L.-F.; Duan, P.-G.; Jiao, J.-L.; Xu, Y.-P. Hydrothermal Gasification of Microalgae over Nickel Catalysts for Production of Hydrogen-Rich Fuel Gas: Effect of Zeolite Supports. Int. J. Hydrog. Energy 2019, 44, 5114–5124. [Google Scholar] [CrossRef]
- Raheem, A.; Cui, X.; Mangi, F.H.; Memon, A.A.; Ji, G.; Cheng, B.; Dong, W.; Zhao, M. Hydrogen-Rich Energy Recovery from Microalgae (Lipid-Extracted) via Steam Catalytic Gasification. Algal Res. 2020, 52, 102102. [Google Scholar] [CrossRef]
- Phunpruch, S. Biomass and Biohydrogen Production by Unicellular Green Alga Chlorella vulgaris var. vulgaris TISTR 8261 Using Frozen Food Industrial Wastewater. Asia-Pac. J. Sci. Technol. 2022, 27, 9-Jan. [Google Scholar] [CrossRef]
- Maliutina, K.; Tahmasebi, A.; Yu, J. Pressurized Entrained-Flow Pyrolysis of Microalgae: Enhanced Production of Hydrogen and Nitrogen-Containing Compounds. Bioresour. Technol. 2018, 256, 160–169. [Google Scholar] [CrossRef]
- Rosha, P.; Kumar, S.; Vikram, S.; Ibrahim, H.; Al-Muhtaseb, A.H. H2-Enriched Gaseous Fuel Production via Co-Gasification of an Algae-Plastic Waste Mixture Using Aspen PLUS. Int. J. Hydrog. Energy 2022, 47, 26294–26302. [Google Scholar] [CrossRef]
- Ban, S.; Lin, W.; Luo, J. Ca2+ Enhances Algal Photolysis Hydrogen Production by Improving the Direct and Indirect Pathways. Int. J. Hydrog. Energy 2019, 44, 1466–1473. [Google Scholar] [CrossRef]
- Chen, J.; Li, Y.; Li, M.; Shi, J.; Wang, L.; Luo, S.; Liu, H. Chemical Flocculation-Based Green Algae Materials for Photobiological Hydrogen Production. ACS Appl. Bio Mater. 2022, 5, 897–903. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sivaramakrishnaiah, M.; Deepanraj, B.; Saravanan, R.; Reddy, M.V. A Novel Optimization Approach for Biohydrogen Production using Algal Biomass. Int. J. Hydrog. Energy 2022. [Google Scholar] [CrossRef]
- Sittijunda, S.; Sitthikitpanya, N.; Plangklang, P.; Reungsang, A. Two-Stage Anaerobic Codigestion of Crude Glycerol and Micro-Algal Biomass for Biohydrogen and Methane Production by Anaerobic Sludge Consortium. Fermentation 2021, 7, 175. [Google Scholar] [CrossRef]
- Pewnual, T.; Jampapetch, N.; Saladtook, S.; Raksajit, W.; Klinsalee, R.; Maneeruttanarungroj, C. Response of Green Alga Tetraspora sp., CU2551 under Potassium Deprivation: A New Promising Strategy for Hydrogen Production. J. Appl. Phycol. 2022, 34, 811–819. [Google Scholar] [CrossRef]
- Manoyan, J.; Samovich, T.; Kozel, N.; Demidchik, V.; Gabrielyan, L. Growth Characteristics, Biohydrogen Production and Photochemical Activity of Photosystems in Green Microalgae Parachlorella kessleri Exposed to Nitrogen Deprivation. Int. J. Hydrog. Energy 2022, 47, 16815–16823. [Google Scholar] [CrossRef]
- Bechara, R.; Azizi, F.; Boyadjian, C. Process Simulation and Optimization for Enhanced Biophotolytic Hydrogen Production from Green Algae Using the Sulfur Deprivation Method. Int. J. Hydrog. Energy 2021, 46, 14096–14108. [Google Scholar] [CrossRef]
- Laokua, N.; Rittiyan, N.; Kornrawudaphikasama, Y.; Klinsalee, R.; Tonawut, Y.; Preechaphonkul, N.; Raksajit, W.; Khetkorn, W.; Dejtisakdi, W.; Maneeruttanarungroj, C. Optimal Conditions for Maximized H2 Yield from a New Green Algal Strain Chlorella sp., KLSc61. J. Appl. Phycol. 2022, 34, 1909–1919. [Google Scholar] [CrossRef]
- Manoyan, J.; Gabrielyan, L.; Kozel, N.; Trchounian, A. Regulation of Biohydrogen Production by Protonophores in Novel Green Microalgae Parachlorella Kessleri. J. Photochem. Photobiol. B Biol. 2019, 199, 111597. [Google Scholar] [CrossRef]
- Paramesh, K.; Lakshmana Reddy, N.; Shankar, M.V.; Chandrasekhar, T. Enhancement of Biological Hydrogen Production Using Green Alga Chlorococcum minutum. Int. J. Hydrog. Energy 2018, 43, 3957–3966. [Google Scholar] [CrossRef]
- Kanygin, A.; Smith, A.; Nagy, V.; Tóth, S.Z.; Redding, K.E. Interplay between Hydrogen Production and Photosynthesis in a Green Alga Expressing an Active Photosystem I-Hydrogenase Chimera. Int. J. Hydrog. Energy 2022, 47, 21969–21983. [Google Scholar] [CrossRef]
- Rai, A.; Khan, M.J.; Ahirwar, A.; Deka, R.; Singh, N.; Schoefs, B.; Marchand, J.; Varjani, S.; Vinayak, V. Hydrogen Economy and Storage by Nanoporous Microalgae Diatom: Special Emphasis on Designing Photobioreactors. Int. J. Hydrog. Energy 2022, 47, 42099–42121. [Google Scholar] [CrossRef]
- Cheng, J.; Yue, L.; Hua, J.; Dong, H.; Li, Y.-Y.; Zhou, J.; Lin, R. Hydrothermal Heating with Sulphuric Acid Contributes to Improved Fermentative Hydrogen and Methane Co-Generation from Dianchi Lake Algal Bloom. Energy Convers. Manag. 2019, 192, 282–291. [Google Scholar] [CrossRef]
- Homburg, S.V.; Kruse, O.; Patel, A.V. Viability, Growth, and Hydrogen Production of Green Microalgae in Novel Silica Hydrogels. Chem. Ing. Tech. 2018, 90, 1162. [Google Scholar] [CrossRef] [Green Version]
- Ban, S.; Lin, W.; Wu, F.; Luo, J. Algal-Bacterial Cooperation Improves Algal Photolysis-Mediated Hydrogen Production. Bioresour. Technol. 2018, 251, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Hupp, B.; Pap, B.; Farkas, A.; Maróti, G. Development of a Microalgae-Based Continuous Starch-to-Hydrogen Conversion Approach. Fermentation 2022, 8, 294. [Google Scholar] [CrossRef]
- Shetty, P.; Boboescu, I.Z.; Pap, B.; Wirth, R.; Kovács, K.L.; Bíró, T.; Futó, Z.; White, R.A.; Maróti, G. Exploitation of Algal-Bacterial Consortia in Combined Biohydrogen Generation and Wastewater Treatment. Front. Energy Res. 2019, 7, 52. [Google Scholar] [CrossRef]
- Jokel, M.; Nagy, V.; Tóth, S.Z.; Kosourov, S.; Allahverdiyeva, Y. Elimination of the Flavodiiron Electron Sink Facilitates Long-Term H2 Photoproduction in Green Algae. Biotechnol. Biofuels 2019, 12, 280. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhuang, X.; Chen, M.; Zeng, Z.; Cai, X.; Li, H.; Hu, Z. An Endogenous MicroRNA (MiRNA1166.1) Can Regulate Photobio-H2 Production in Eukaryotic Green Alga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2018, 11, 126. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Jiang, X.; Hu, C.; Sun, T.; Zeng, Z.; Cai, X.; Li, H.; Hu, Z. Optogenetic Regulation of Artificial MicroRNA Improves H2 Production in Green Alga Chlamydomonas reinhardtii. Biotechnol. Biofuels 2017, 10, 257. [Google Scholar] [CrossRef] [Green Version]
- Elman, T.; Hoai Ho, T.T.; Milrad, Y.; Hippler, M.; Yacoby, I. Enhanced Chloroplast-Mitochondria Crosstalk Promotes Ambient Algal-H2 Production. Cell Rep. Phys. Sci. 2022, 3, 100828. [Google Scholar] [CrossRef]
- Liu, J.-Z.; Ge, Y.-M.; Sun, J.-Y.; Chen, P.; Addy, M.; Huo, S.-H.; Li, K.; Cheng, P.-F.; Ruan, R. Exogenic Glucose as an Electron Donor for Algal Hydrogenases to Promote Hydrogen Photoproduction by Chlorella pyrenoidosa. Bioresour. Technol. 2019, 289, 121762. [Google Scholar] [CrossRef] [PubMed]
- Phunpruch, S. Effect of Cell Density and Nutrient Deprivation on Hydrogen Production by Unicellular Green Alga Scenedesmus sp., KMITL-OVG1. Asia-Pac. J. Sci. Technol. 2019, 24, 1–9. [Google Scholar] [CrossRef]
- Sengmee, D.; Cheirsilp, B.; Suksaroge, T.T.; Prasertsan, P. Biophotolysis-Based Hydrogen and Lipid Production by Oleaginous Microalgae Using Crude Glycerol as Exogenous Carbon Source. Int. J. Hydrog. Energy 2017, 42, 1970–1976. [Google Scholar] [CrossRef]
- Gholkar, P.; Shastri, Y.; Tanksale, A. Catalytic Reactive Flash Volatilisation of Microalgae to Produce Hydrogen or Methane-Rich Syngas. Appl. Catal. B Environ. 2019, 251, 326–334. [Google Scholar] [CrossRef]
- Duangjan, K.; Nakkhunthod, W.; Pekkoh, J.; Pumas, C. Comparison of Hydrogen Production in Microalgae under Autotrophic and Mixotrophic Media. Bot. Lith. 2017, 23, 169–177. [Google Scholar] [CrossRef] [Green Version]
- Narravula Raga, S.; Duddela, V.; Pallaval Veera, B.; Poda, S.; Thummala, C. Effects of Various Factors on Biomass, Bioethanol, and Biohydrogen Production in Green Alga Chlamydomonas reinhardtii. J. App. Biol. Biotech. 2021, 9, 152–156. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of bioH2 production through (a) direct biophotolysis and (b) indirect biophotolysis.
Figure 1.
Schematic diagram of bioH2 production through (a) direct biophotolysis and (b) indirect biophotolysis.

Figure 2.
Research methodology flowchart. (n: number of articles).

Figure 3.
Number of articles and number of citations for articles published on the topic of bioH2 production from microalgae.
Figure 3.
Number of articles and number of citations for articles published on the topic of bioH2 production from microalgae.

Figure 4.
The network map for the top keywords is subdivided into four clusters based on the similarities. Keywords related to bioH2 were removed. (The size of the node represents the frequency of occurrence, and the curved lines show the co-appearance between the keywords).
Figure 4.
The network map for the top keywords is subdivided into four clusters based on the similarities. Keywords related to bioH2 were removed. (The size of the node represents the frequency of occurrence, and the curved lines show the co-appearance between the keywords).

Figure 5.
Emerging keywords in the field of biohydrogen production using microalgae.

Figure 6.
Co-authorship and countries (the size of the node represents the volume of publications, and the line thickness illustrates the term recurrence between the countries).
Figure 6.
Co-authorship and countries (the size of the node represents the volume of publications, and the line thickness illustrates the term recurrence between the countries).

Figure 7.
Co-citation network map. (The node size represents the number of citations, and the curved lines show the co-citation ties between the references).
Figure 7.
Co-citation network map. (The node size represents the number of citations, and the curved lines show the co-citation ties between the references).


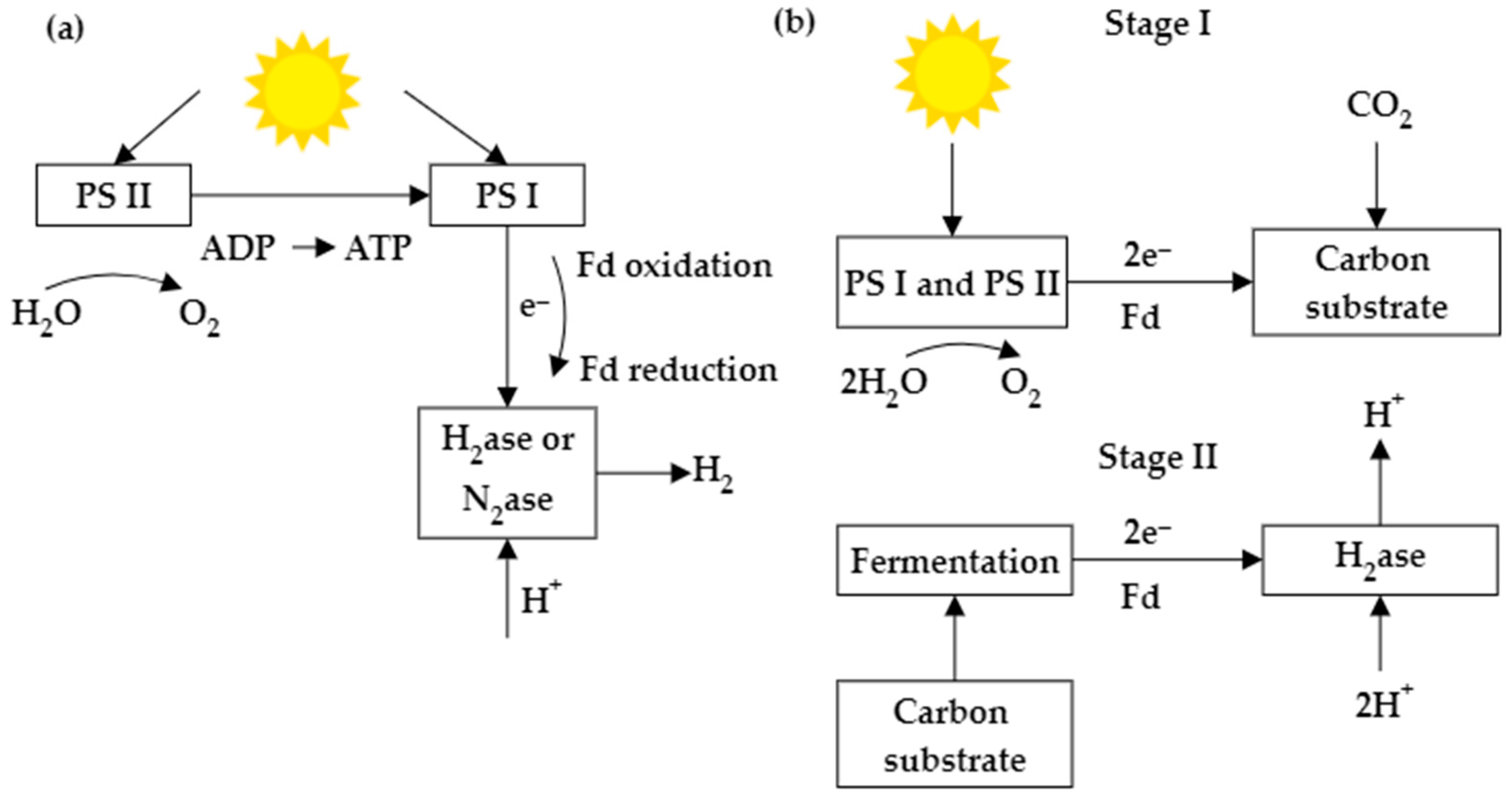{kind=link}
{kind=link}
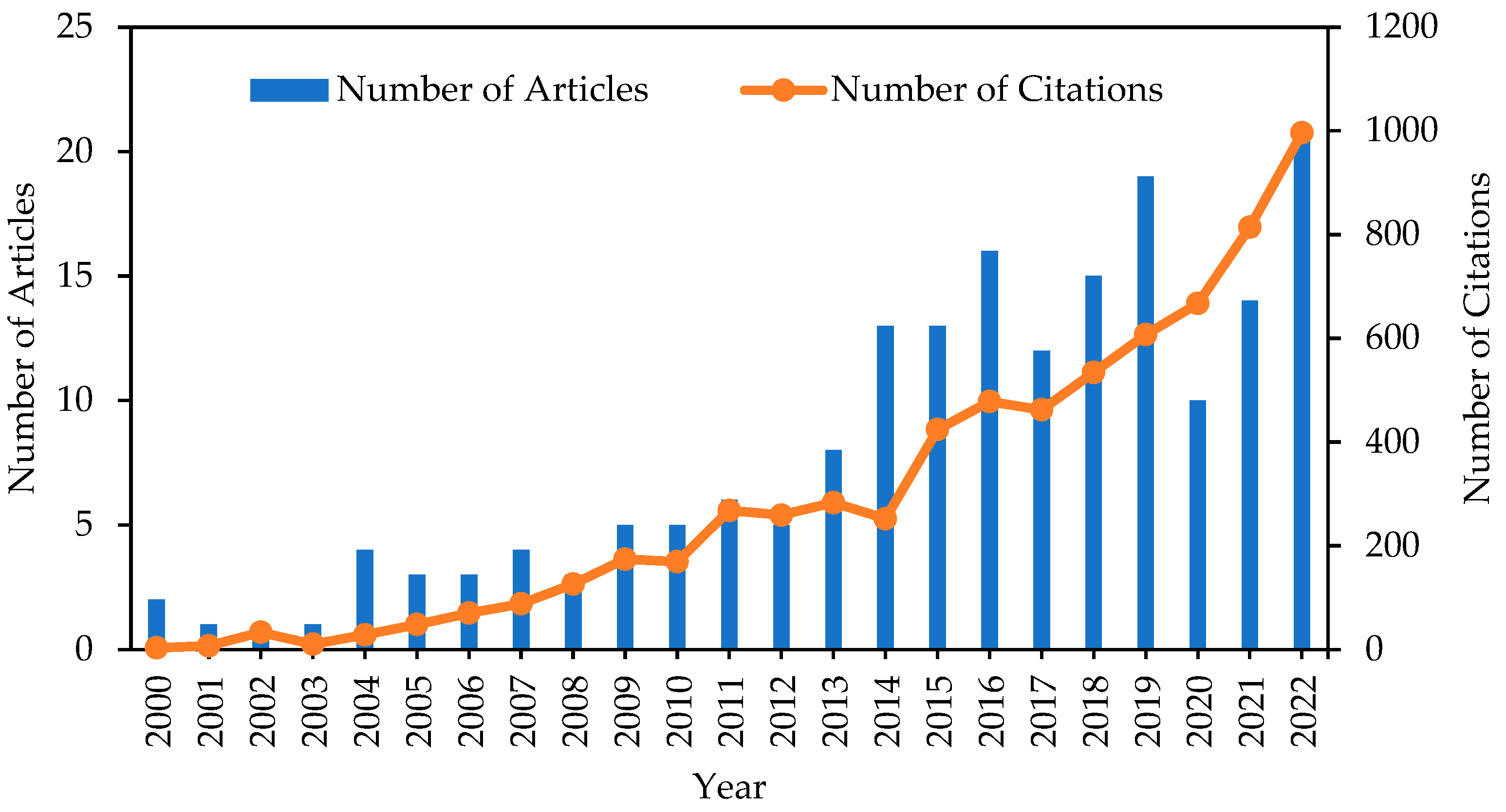{kind=link}
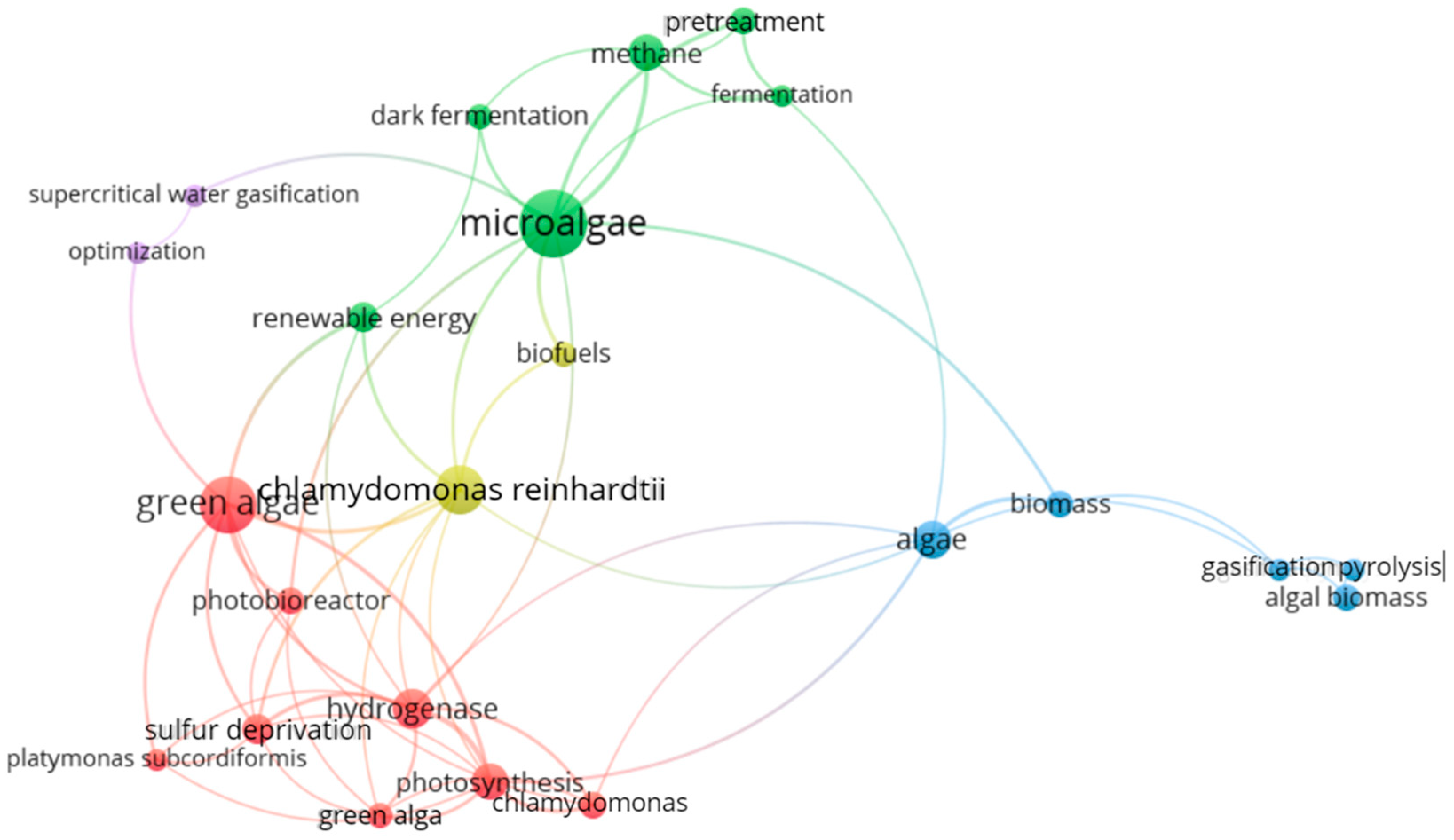{kind=link}
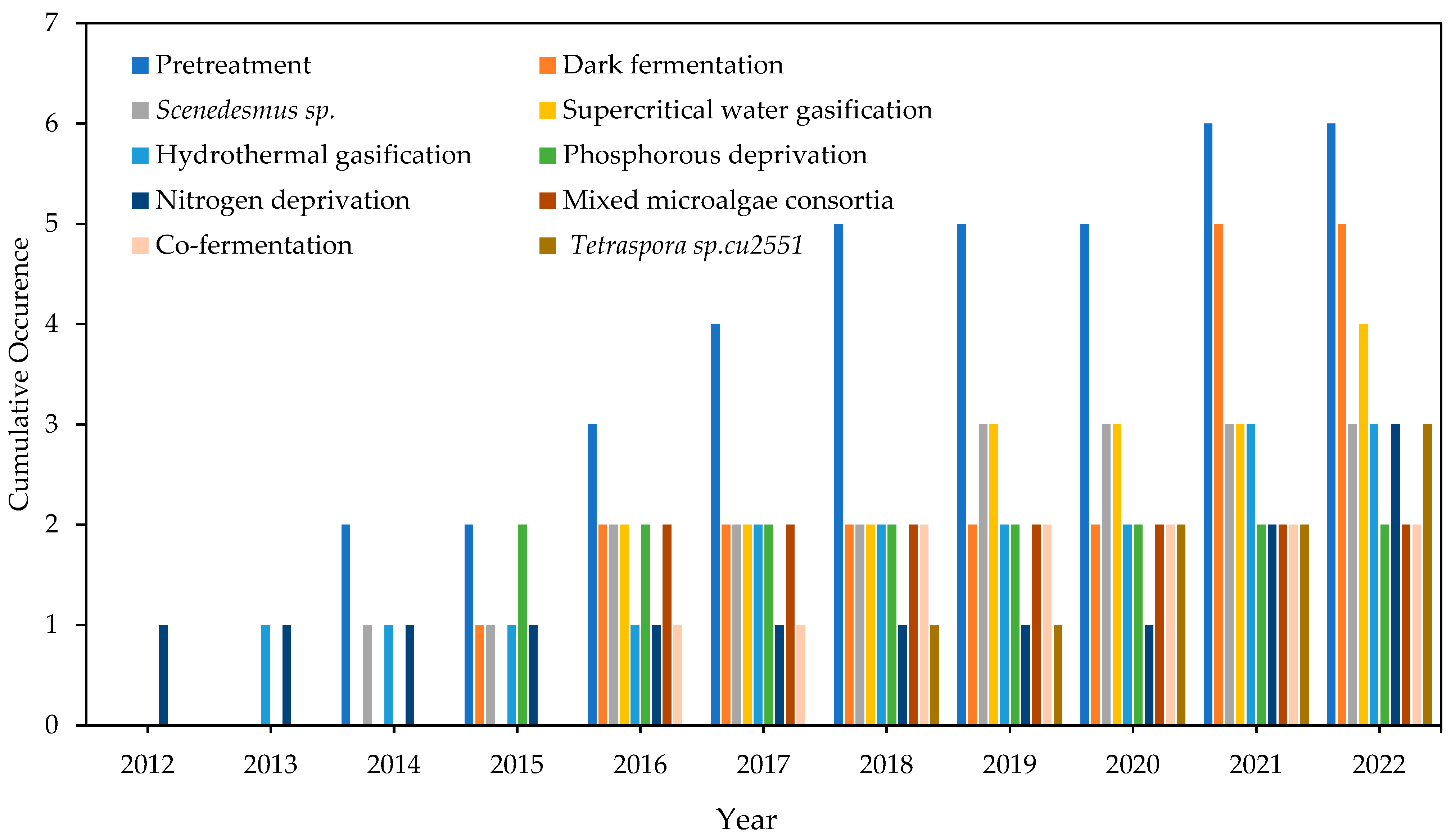{kind=link}
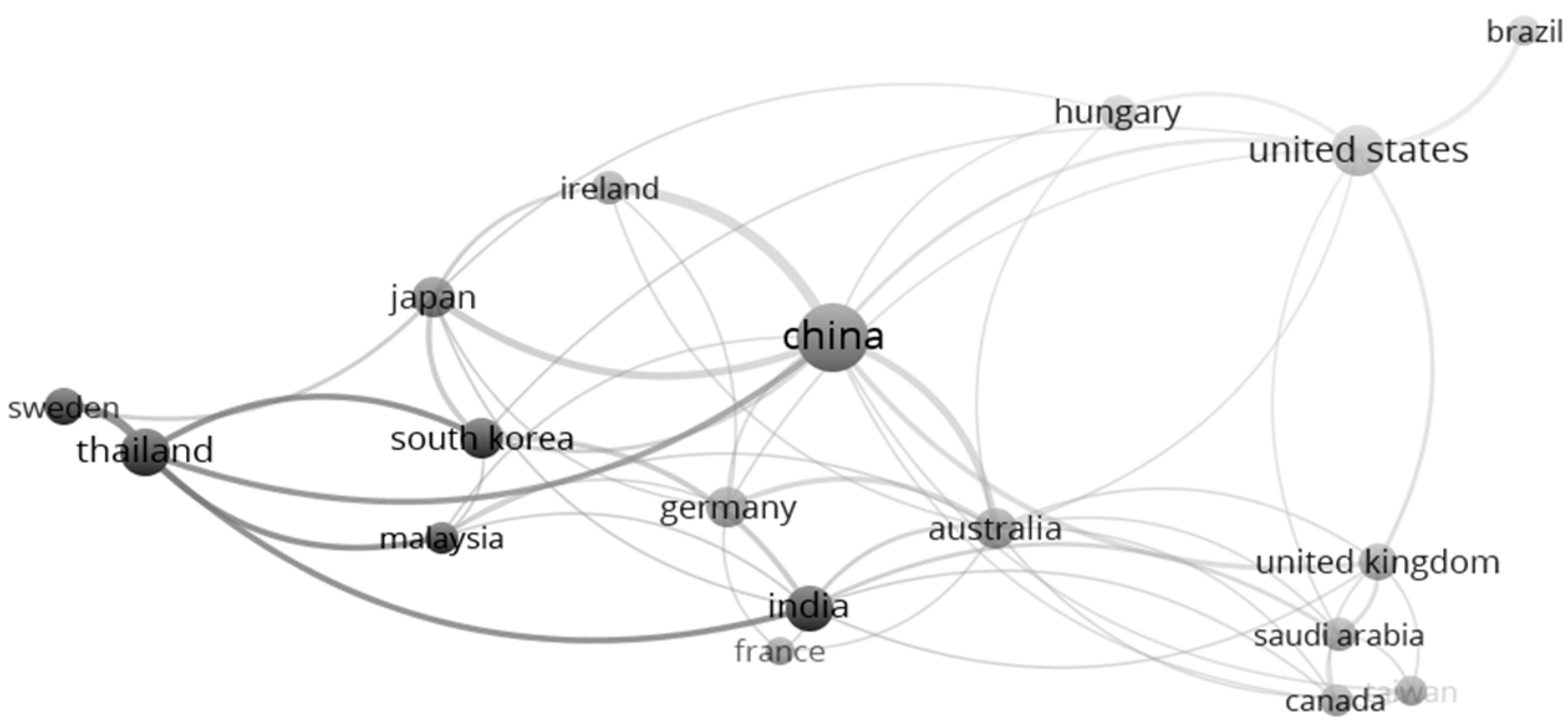{kind=link}
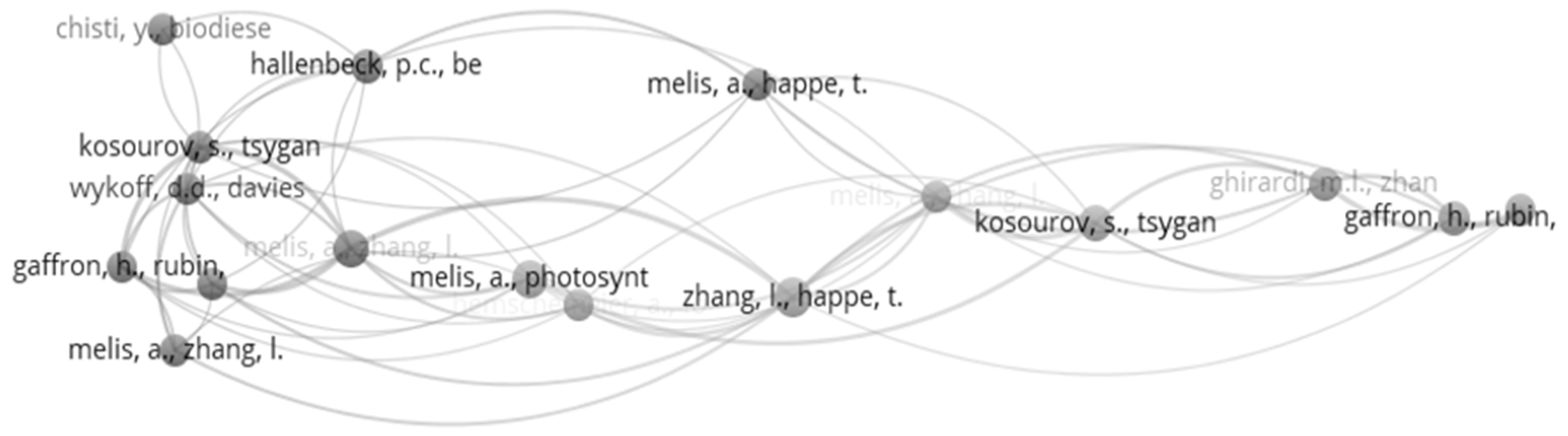{kind=link}
Table 1.
The Total Link Strength of the 23 keywords that have at least four minimum occurrences.
| Rank | Keywords | Occurrences | Total Link Strength |
|---|---|---|---|
| 1 | Microalgae | 34 | 22 |
| 2 | Green algae | 24 | 20 |
| 3 | Chlamydomonas reinhardtii | 18 | 15 |
| 4 | Hydrogenase | 12 | 19 |
| 5 | Algae | 11 | 10 |
| 6 | Methane | 10 | 8 |
| 7 | Photosynthesis | 10 | 18 |
| 8 | Renewable energy | 7 | 7 |
| 9 | Sulfur deprivation | 7 | 11 |
| 10 | Algal biomass | 6 | 2 |
| 11 | Biomass | 6 | 6 |
| 12 | Chlamydomonas | 6 | 8 |
| 13 | Photobioreactor | 6 | 6 |
| 14 | Pretreatment | 6 | 6 |
| 15 | Biofuels | 5 | 5 |
| 16 | Dark fermentation | 5 | 4 |
| 17 | Green alga | 5 | 8 |
| 18 | Fermentation | 4 | 6 |
| 19 | Gasification | 4 | 4 |
| 20 | Optimization | 4 | 3 |
| 21 | Platymonas subcordiformis | 4 | 5 |
| 22 | Pyrolysis | 4 | 2 |
| 23 | Supercritical water gasification | 4 | 3 |
Table 2.
Top Journals ranked by TC, TP, and CPP.
| Ranked by TP * | Ranked by TC * | Ranked by CPP * | ||||||
|---|---|---|---|---|---|---|---|---|
| Rank | Journal Name | TP | Rank | Journal Name | TC | Rank | Journal Name | CPP |
| 1 | Int. J. Hydrog. Energy | 47 | 1 | Int. J. Hydrog. Energy | 1194 | 1 | Plant Physiology | 873 |
| 2 | Bioresource Technology | 17 | 2 | Bioresource Technology | 1016 | 2 | Planta | 341 |
| 3 | Biotechnology for Biofuels | 7 | 3 | Plant Physiology | 873 | 3 | J. Biological Chemistry | 195 |
| 4 | Energy | 5 | 4 | J. Biological Chemistry | 391 | 4 | BBA—Bioenergetics | 165 |
| 5 | Fuel | 4 | 5 | Planta | 341 | 5 | Eukaryotic cell | 111 |
| 6 | J. Applied Phycology | 4 | 6 | Biotechnology for Biofuels | 247 | 6 | Dalton Transactions | 97 |
| 7 | Algal Research | 3 | 7 | Biochemical Eng. J | 175 | 7 | J. Bioscience and Bio. Eng | 94 |
| 8 | Asia-Pacific J. Sci. Technol. | 3 | 8 | BBA—Bioenergetics | 165 | 8 | Biochemical Eng. J | 87 |
| 9 | Biotechnology Letters | 3 | 9 | Fuel | 125 | 9 | Nature Communications | 80 |
| 10 | Biotechnology Progress | 3 | 10 | PLOS One | 124 | 10 | Water Research | 78 |
Note(s): * TP: Total Publications, TC: Total Citations, CPP: Citation per publication; Int. J. Hydrog. Energy: International Journal of Hydrogen Energy, Asia-Pacific J. Sci. Technol: Asia-Pacific Journal of Science Technology, J. Applied Phycology: Journal of Applied Phycology, J. Biological Chemistry: Journal of Biological Chemistry, BBA—Bioenergetics: Biochimica et Biophysica Acta-Bioenergetics, Biochemical Eng. J: Biochemical Engineering Journal, J. Bioscience, and Bio. Eng: Journal of Bioscience and Bioengineering.
Table 3.
Influential countries ranked by TP, TC, and CPP.
| Ranked by TP | Ranked by TC | Ranked by CPP | ||||||
|---|---|---|---|---|---|---|---|---|
| Rank | Country | TP | Rank | Country | TC | Rank | Country | CPP |
| 1 | China | 51 | 1 | USA | 1944 | 1 | Germany | 96 |
| 2 | USA | 26 | 2 | China | 1636 | 2 | USA | 75 |
| 3 | Thailand | 17 | 3 | Germany | 1255 | 3 | France | 71 |
| 4 | India | 16 | 4 | Australia | 897 | 4 | Australia | 64 |
| 5 | Australia | 14 | 5 | Japan | 581 | 5 | Ireland | 50 |
| 6 | Germany | 13 | 6 | United Kingdom | 421 | 6 | Japan | 45 |
| 7 | Japan | 13 | 7 | Ireland | 397 | 7 | United Kingdom | 38 |
| 8 | United Kingdom | 11 | 8 | France | 355 | 8 | South Korea | 33 |
| 9 | South Korea | 10 | 9 | India | 335 | 9 | China | 32 |
| 10 | Hungary | 9 | 10 | South Korea | 326 | 10 | Canada | 31 |
Table 4.
Top authors ranked by NCS.
| Ranked by Normalized Citation Score (NCS) | ||||
|---|---|---|---|---|
| Rank | Author | Affiliation | Country | NCS |
| 1 | Jerry D Murphy | University College Cork | Ireland | 10 |
| 2 | Ao Xia | Chongqing University | China | 9 |
| 3 | Maria Lucia Ghirardi | National Renewable Energy Lab. | USA | 9 |
| 4 | Jie Cheng | Zhejiang University | China | 8 |
| 5 | Lin Zhengyan | Zhejiang University | China | 8 |
| 6 | Mei Zhao | Jiangnan University | China | 7 |
| 7 | Michael Seibert | National Renewable Energy Lab. | USA | 6 |
| 8 | Ben Hankamer | The University of Queensland | Australia | 6 |
| 9 | Gopalakrishnan Kumar | University of Stavanger | Norway | 6 |
| 10 | Thomas Happe | Ruhr-Universitat Bochum | Germany | 6 |
Table 5.
Top articles based on local and global citations.
| Ranked by Global Citations | Ranked by Local Citations | ||||
|---|---|---|---|---|---|
| Rank | Article | Global Citations | Rank | Article | Local Citations |
| 1 | Melis et al. (2000) [51] | 873 | 1 | Melis et al. (2000) [51] | 60 |
| 2 | Zhang et al. (2002) [52] | 341 | 2 | Zhang et al. (2002) [52] | 20 |
| 3 | Kruse et al. (2005) [56] | 286 | 3 | Kruse et al. (2005) [56] | 12 |
| 4 | Antal et al. (2003) [57] | 165 | 4 | Guan et al. (2004) [58] | 11 |
| 5 | Onwudili et al. (2013) [50] | 147 | 5 | Maneeruttanarungroj et al. (2010) [28] | 9 |
| 6 | Guan et al. (2004) [58] | 142 | 6 | Lakatos et al. (2014) [59] | 7 |
| 7 | Xia et al. (2016) [2] | 134 | 7 | Skjanes et al. (2008) [49] | 7 |
| 8 | Duman et al. (2014) [60] | 124 | 8 | Hwang et al. (2014) [61] | 7 |
| 9 | Nguyen et al. (2008) [62] | 111 | 9 | Antal et al. (2003) [57] | 7 |
| 10 | Srirangan et al. (2011) [4] | 110 | 10 | Onwudili et al. (2013) [50] | 6 |
Table 6.
Research themes and recent advancements in the field.
| Research Theme | Recent Advancement |
|---|---|
| Techniques to produce bioH2 using microalgae | Co-fermentation of cyanobacteria and glucose with Fe3O4 nanoparticles [69], co-fermentation of pretreated rice residue and microalgae [70], anaerobic co-digestion of Taihu Algae and food waste [71], bioH2 production without sulfur using fermenter effluents enriched in acetate [72]. Effect of magnetite (Fe3O4) supplement on bioH2 production of Clostridium Butyricum DSM 10702 by anaerobic fermentation [73]. Dosing of nano zero-valent iron (nZVI) in the dark co-fermentation system [74]. Pyrolysis of dry spirulina algae in an atmospheric pressure microwave plasma reactor [75].Integrated supercritical water gasification (SCWG), syngas chemical looping (SCL), and H2ation system [76,77]. Catalytic SCWG with Nickel (Ni)/zeolite catalysts [78], catalytic and non-catalytic gasification with a hybrid functional mixture of waste eggshell-derived Calcium oxide (CaO), Ni catalyst, and Yttrium oxide Y2O3 [79], Catalytic hydrothermal gasification using noble metals Platinum (Pt) and Ruthenium (Ru) [35]. BioH2 production by Chlorella vulgaris var. vulgaris TISTR 8261 using frozen food industrial wastewater [80]. Pressurized entrained flow pyrolysis of Chlorella vulgaris microalgae [81].Co-gasification of algae-plastic waste for bioH2 production by Aspen Plus simulation model [82]. Application of Calcium ions (Ca2+) to enhance algal photolysis bioH2 production [83]. Chemical flocculation of the green algae Chlorella pyrenoidosa to form aggregates for 11 days of continuous photobiological hydrogen production [84]. |
| Optimization of factors affecting bioH2 production from microalgae | Response surface methodology to optimize sulfur concentration, run time, and algal biomass concentration [44,85], response surface methodology with central composite design for optimization of factors such as crude glycerol, microalgal biomass, and inoculum of bioH2 production from co-digestion of crude glycerol and microalgae by anaerobic sludge [86]. Potassium deprivation conditions to enhance bioH2 production [87]. Nitrogen-deprived conditions gave 4–5-fold more bioH2 [88]. Optimization of conditions for supercritical gasification using GAMS software [45]. Optimization of essential process parameters such as reaction time, sulfur concentrations, and the medium pH for bio photolytic cyclic process [89]. Optimization of temperature, catalyst loading reaction time for catalytic gasification by central composite design [11], integrated pyrolysis and air gasification processes of algal waste, Chlorella vulgaris, Rhizoclonium sp., and Spirogyra by simulation model developed using Aspen Plus software to optimize the gasifier temperature, gasifier pressure, and air flowrate [22]. |
| Alternative and potential microalgal species | Newly isolated green algae Chlorella sp. KLSc59: [53], Chlorella sp. KLSc61 [90]. Another newly isolated green alga, Scenedesmus acuminatus from Thailand [54], Parachlorella kessleri isolated from Armenia [91], and Chlorococcum minutum [92]. Chlamydomonas reinhardtii having PSI-H2ase chimera polypeptide for enhanced H2 production [93]. H2 generation from the diatoms by culturing it in various solar panel photobioreactors [94]. |
| Strategies to improve the bioH2 production from microalgae | Anaerobic co-digestion of food waste and high-pressure homogenization pretreated Taihu lake wet algae [3], hydrothermal sulfuric acid pretreatment to improve fermentative bioH2 of Dianchi Lake algal bloom [46,95]. An innovative free ammonia pretreatment technology [47]. Immobilized cells of Tetraspora sp. CU2551 in alginate matrix under aerobic sulfur deprivation and under anaerobic sulfur deprivation to maximize bioH2 production [12,13]. The entrapment of microalgae in silica gels [96]. Microalgae-bacteria consortiums such as Chlamydomonas sp. and Pseudomonas sp. strain D [97], Chlamydomonas and Chlorella genera with a starch-degrading bacterium from the Bacillus genus [98], Chlorella vulgaris MACC360, and Archaea [99]. Removal of flavodiiron proteins in Chlamydomonas reinhardtii showed increased bioH2 [100], regulation of photo bioH2 production in Chlamydomonas reinhardtii by an endogenous microRNA [101], Optogenetic regulation of artificial microRNA in Chlamydomonas reinhardtii [102] and Chlamydomonas defective in thylakoid proton gradient overcame the limitation due to O2 and carbon fixations [103]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Arimbrathodi, S.P.; Javed, M.A.; Hamouda, M.A.; Aly Hassan, A.; Ahmed, M.E. BioH2 Production Using Microalgae: Highlights on Recent Advancements from a Bibliometric Analysis. Water 2023, 15, 185. https://doi.org/10.3390/w15010185
AMA Style
Arimbrathodi SP, Javed MA, Hamouda MA, Aly Hassan A, Ahmed ME. BioH2 Production Using Microalgae: Highlights on Recent Advancements from a Bibliometric Analysis. Water. 2023; 15(1):185. https://doi.org/10.3390/w15010185
Chicago/Turabian StyleArimbrathodi, Shirin P., Muhammad Asad Javed, Mohamed A. Hamouda, Ashraf Aly Hassan, and Mahmoud E. Ahmed. 2023. "BioH2 Production Using Microalgae: Highlights on Recent Advancements from a Bibliometric Analysis" Water 15, no. 1: 185. https://doi.org/10.3390/w15010185
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.