


Comparing Water Use Characteristics of Bromus inermis and Medicago sativa Revegetating Degraded Land in Agro-Pasture Ecotone in North China

Abstract
:1. Introduction
2. Materials and Methods
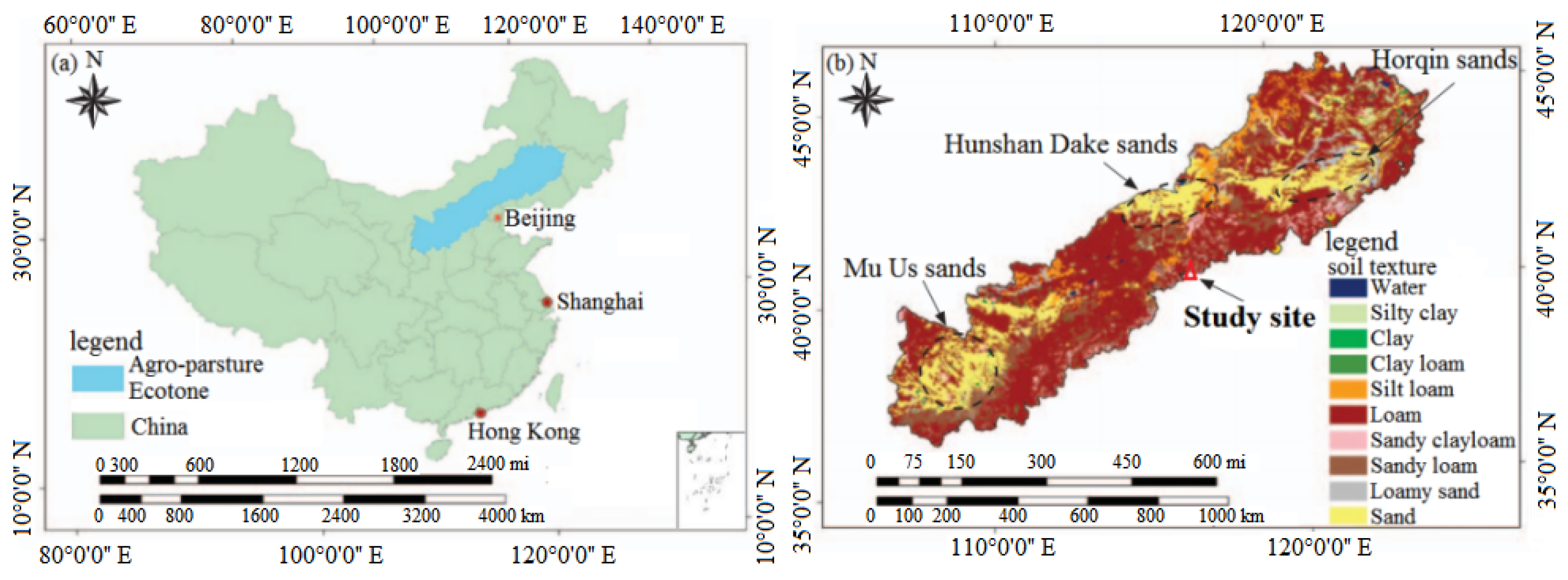
2.1. Site Description
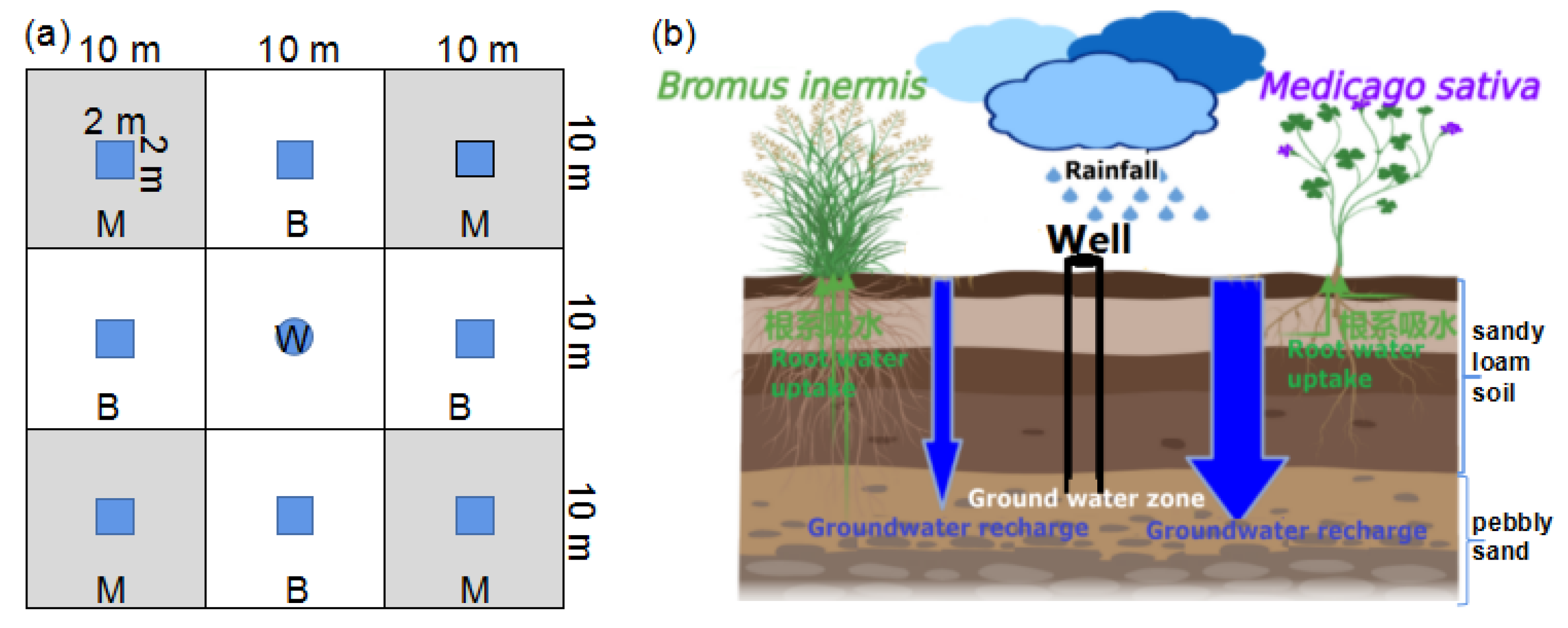
2.2. Sample Collection
2.3. Measurement and Analysis
2.4. Statistical Analysis
3. Results
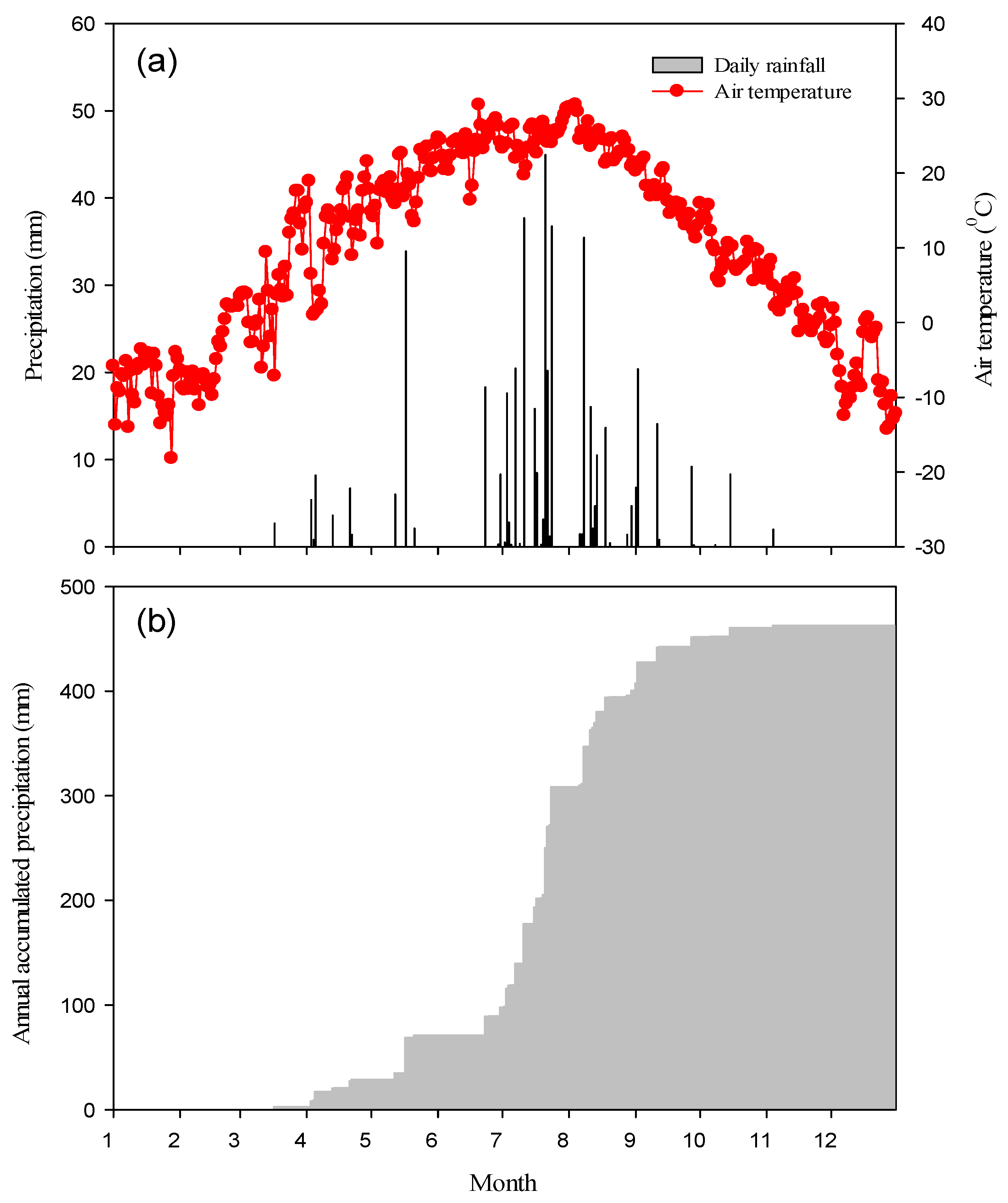
3.1. Meteorological Condition
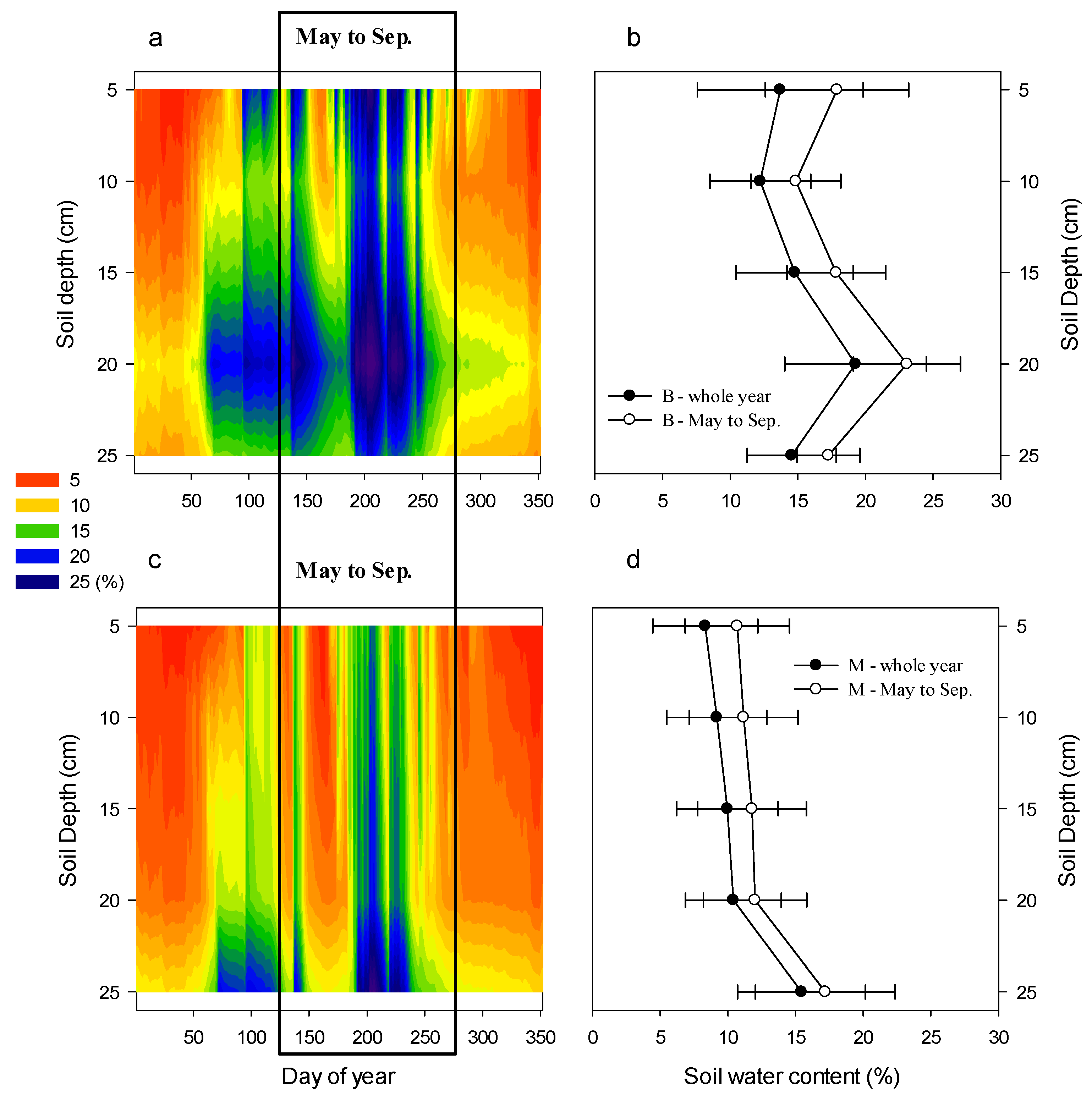
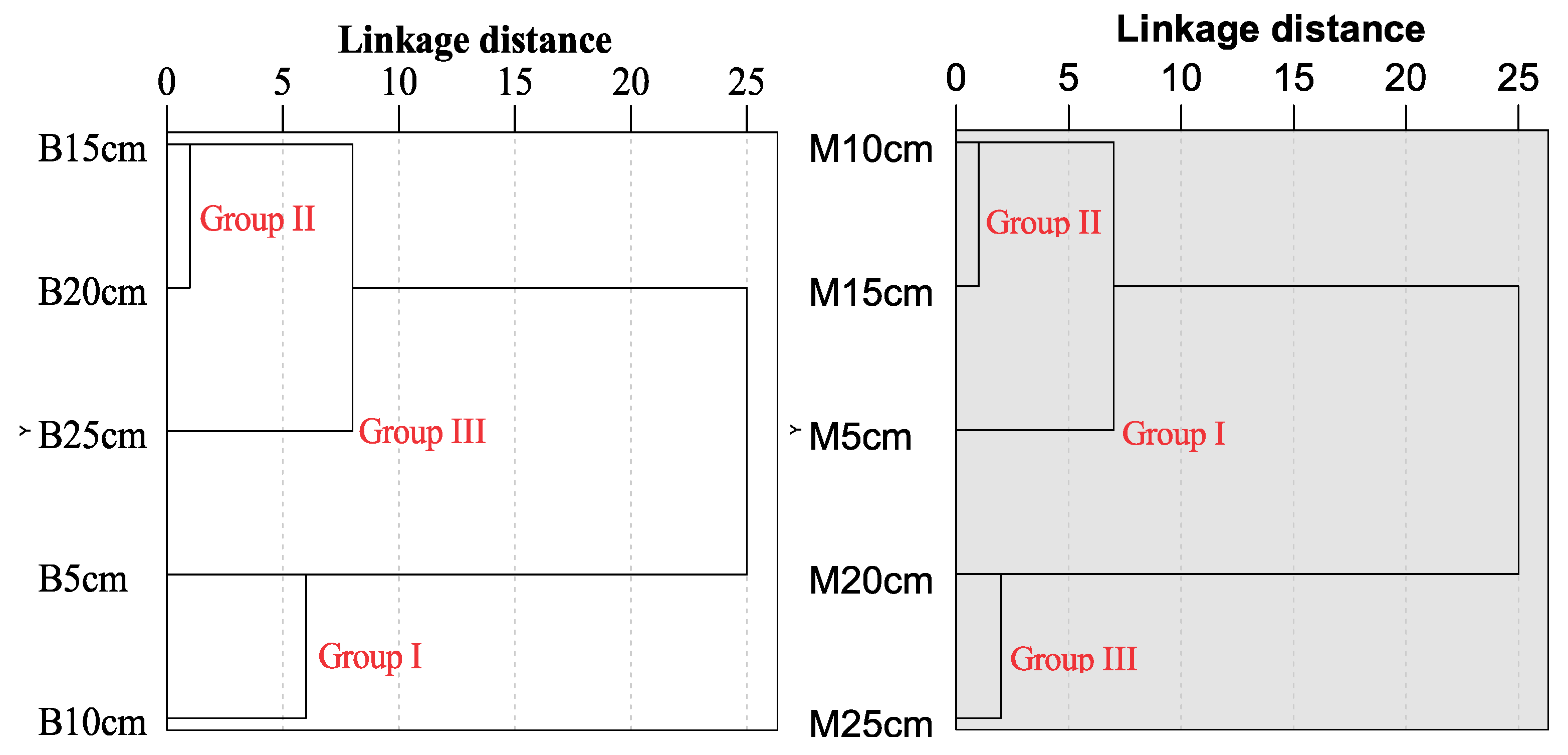
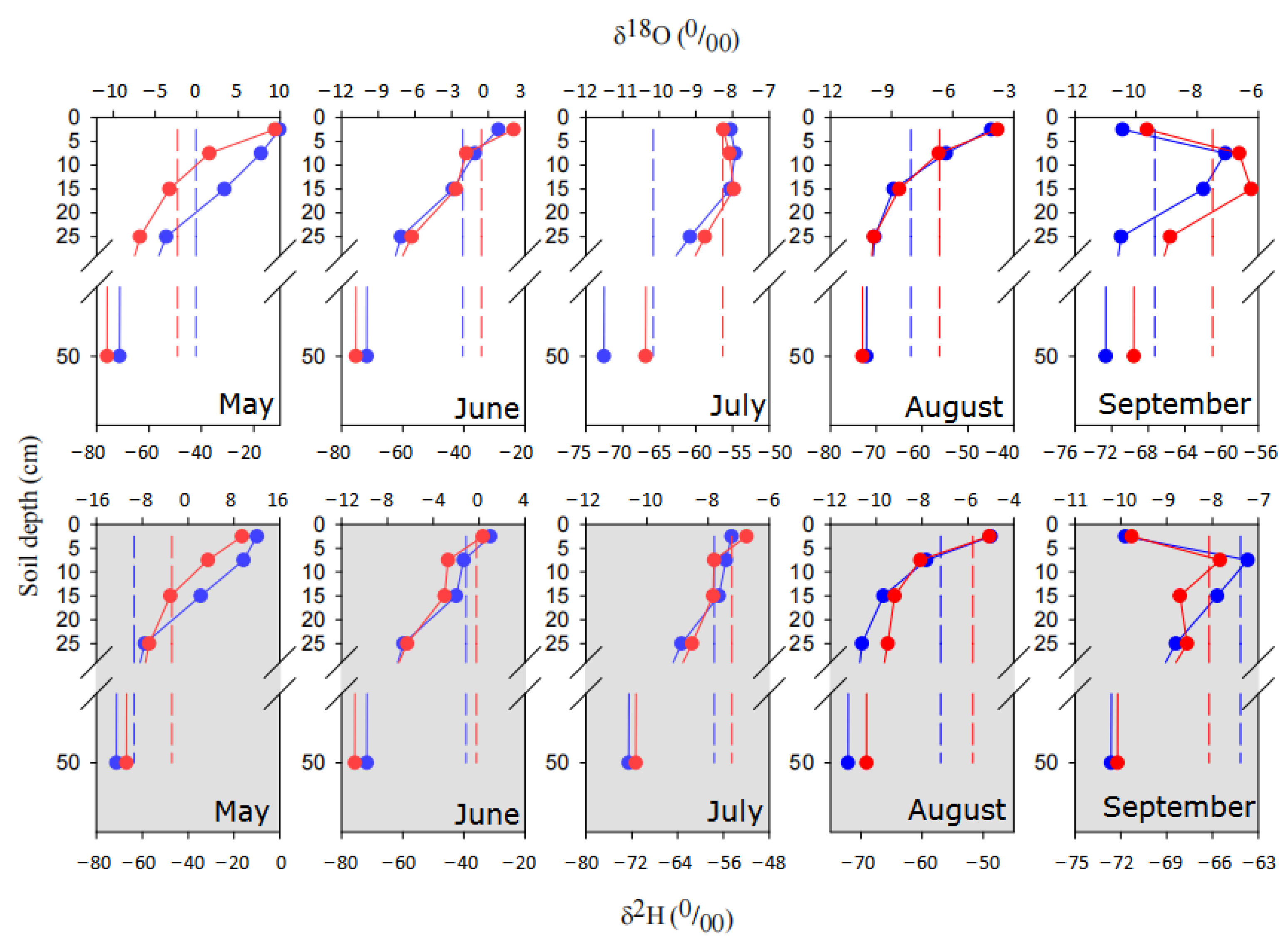
3.2. Soil Water Dynamics and Clustering Analysis
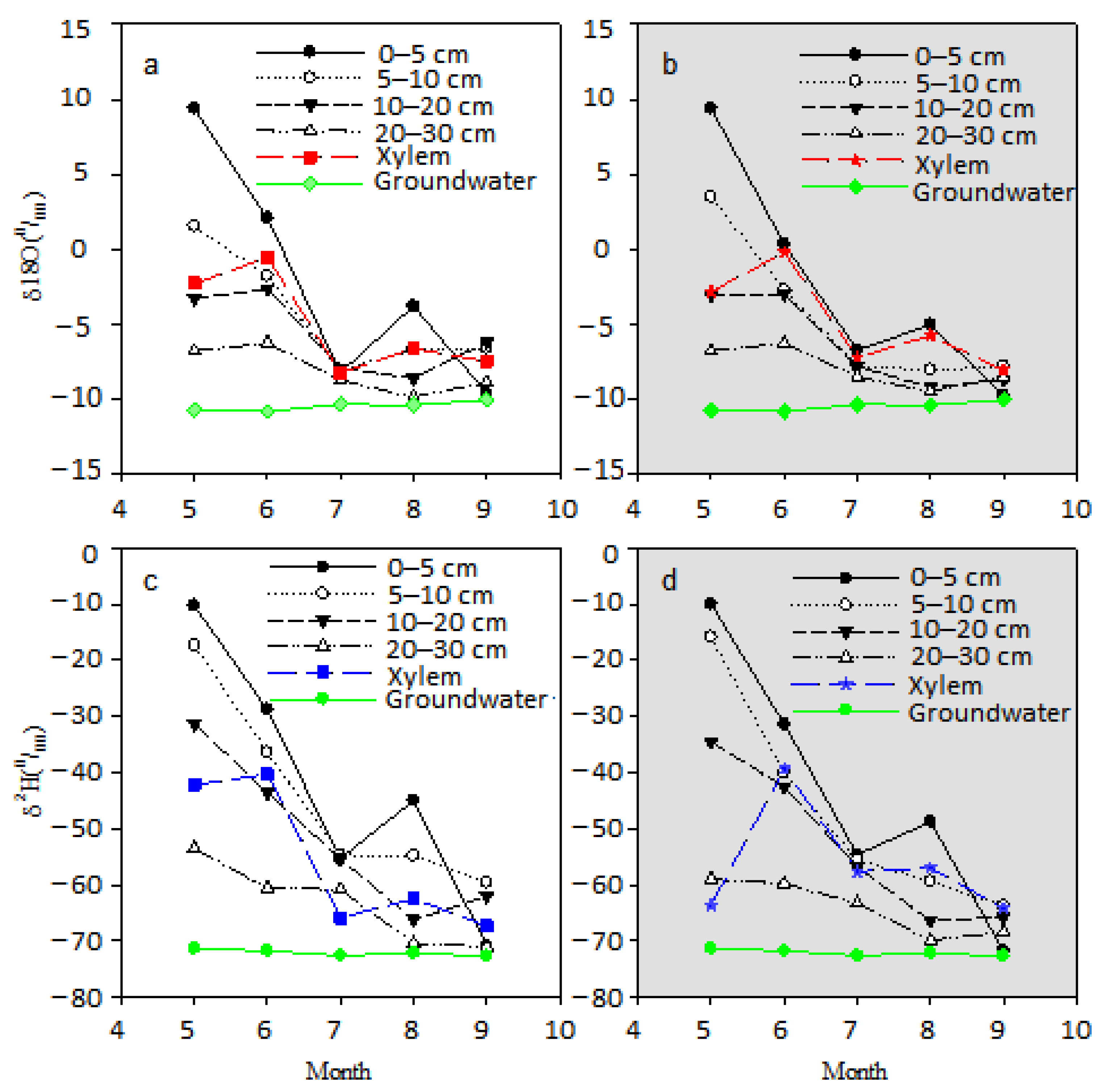
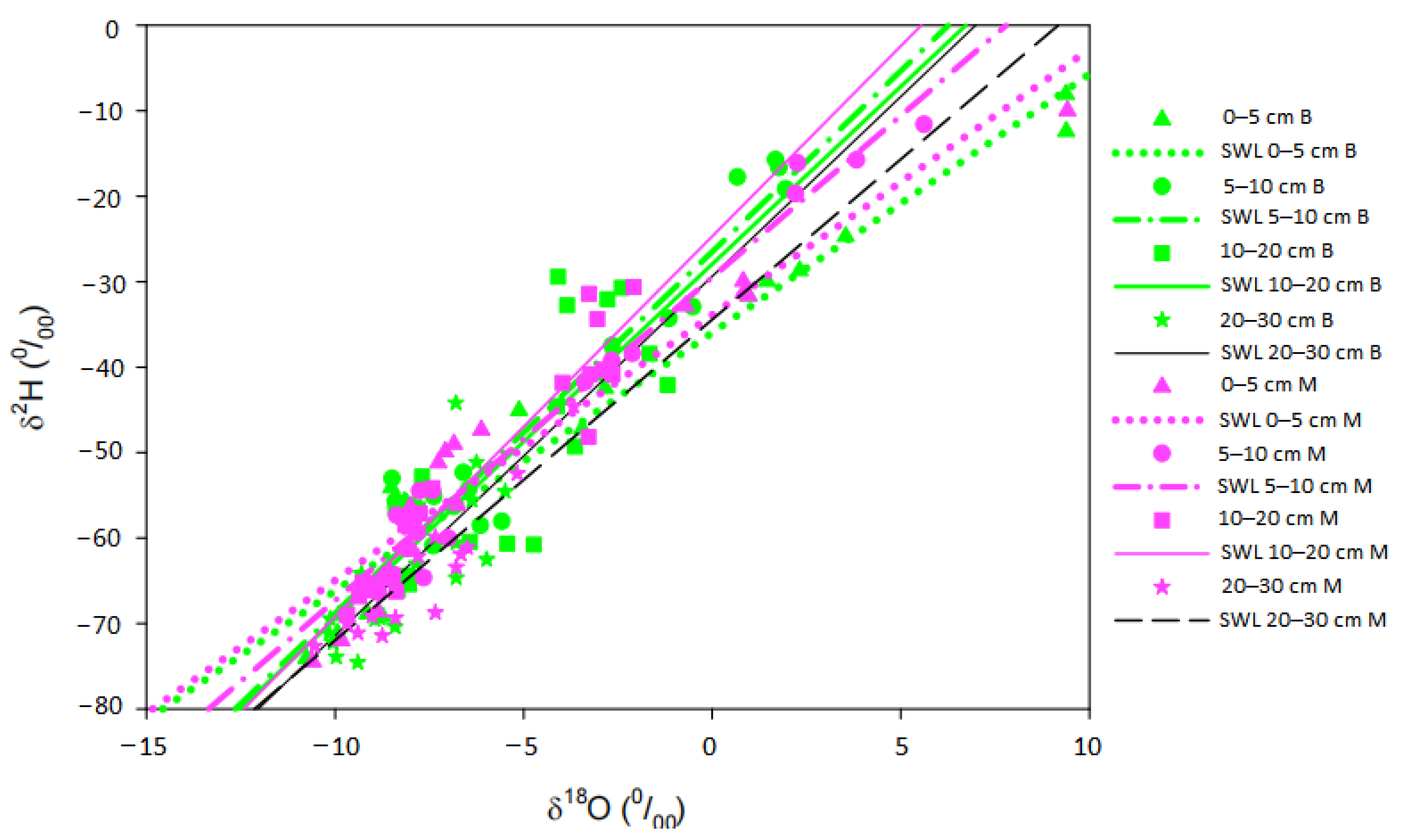
3.3. Isotopic Composition of Precipitation, Groundwater, Xylem Water and Soil Water
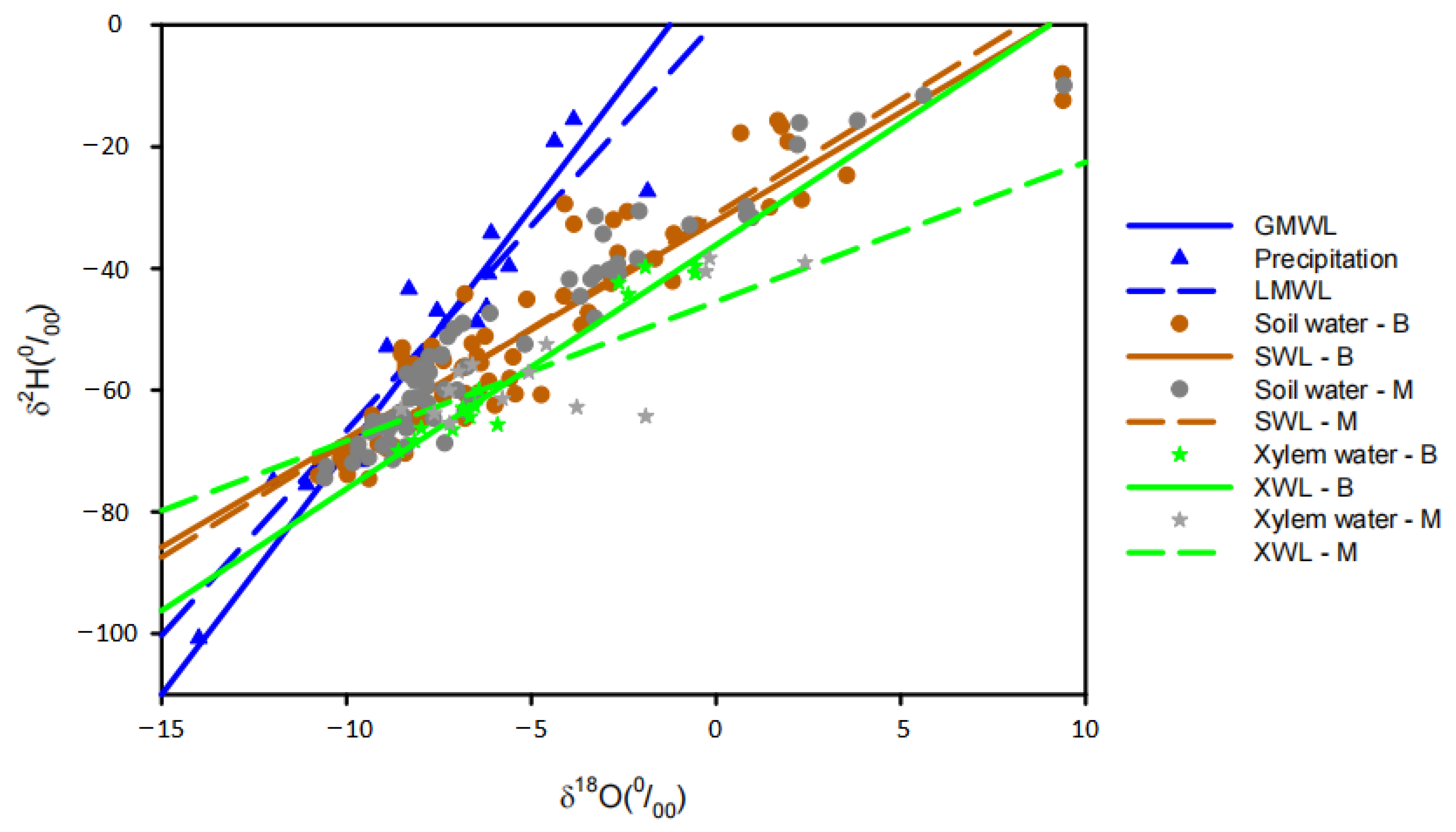
3.4. Intersections of Isotopic Composition between Groundwater, Soil Water and Xylem Water
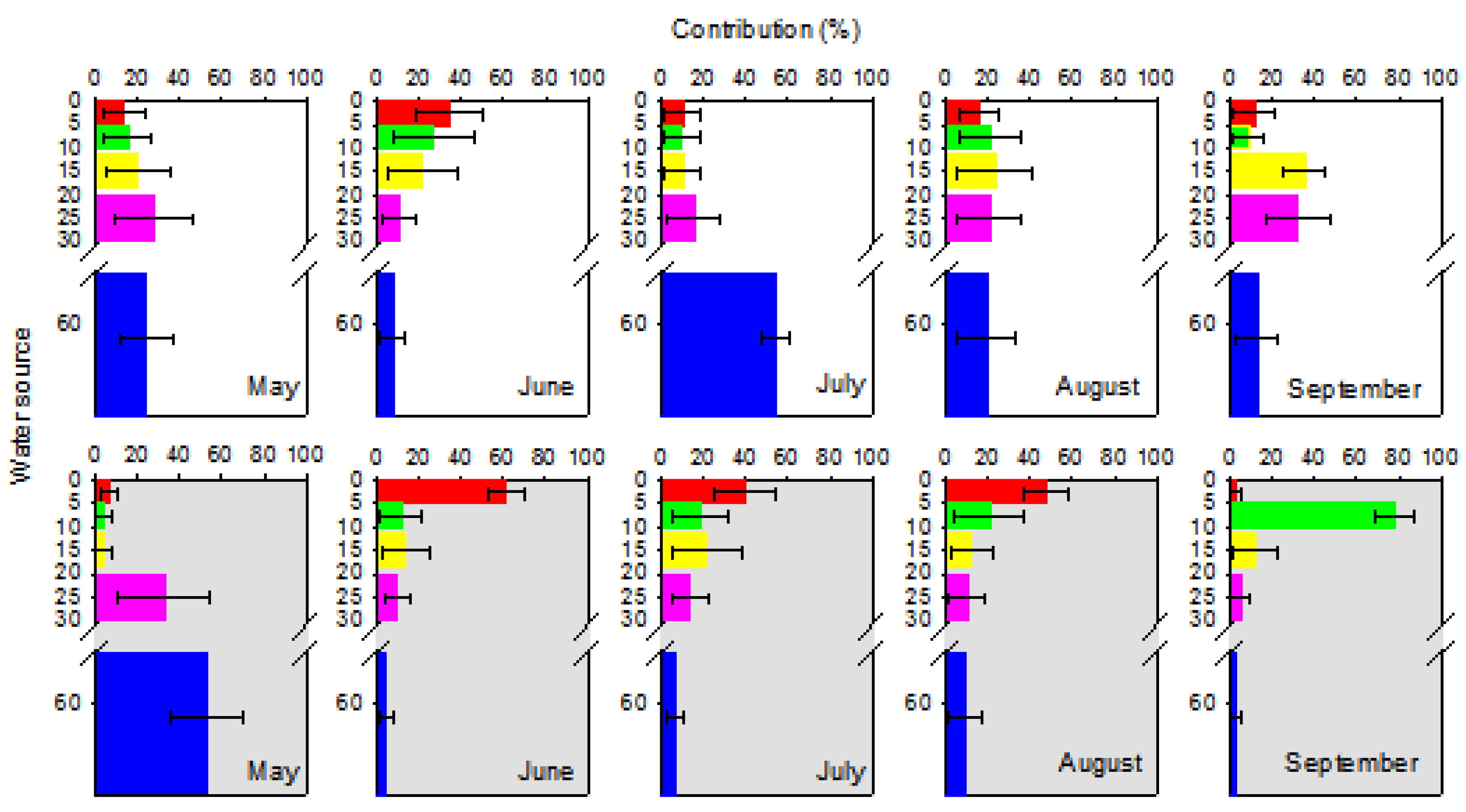
3.5. Contribution of Each Water Source to Plant Water Uptake
4. Discussion
4.1. Water Source Partitioning in the Artificial Grasslands
4.2. Plant Water Use Characteristics in the Different Types of Grassland
4.3. Influences of Land Restoration on Soil Water Dynamics
4.4. Implications
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Guo, Z.; Zobeck, T.M.; Zhang, K.; Li, F. Estimating potential wind erosion of agricultural lands in northern China using the Revised Wind Erosion Equation (RWEQ) and GIS. J. Soil Water Conserv. 2013, 68, 13–21. [Google Scholar] [CrossRef]
- Wang, C.; Gao, Q.; Wang, X.; Yu, M. Decadal trend in agricultural abandonment and woodland expansion in an agro-pastoral transition band in Northern China. PLoS ONE 2015, 10, e0142113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.; Sun, O.J.; Huang, J.; Gao, Y.; Han, X. Land use affects the relationship between species diversity and productivity at the local scale in a semi-arid steppe ecosystem. Funct. Ecol. 2006, 20, 753–762. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wang, K.; Wu, W.L. Dynamics of soil physical and chemical properties and vegetation succession characteristics during grassland desertification under sheep grazing in an agro-pastoral transition zone in Northern China. J. Arid Environ. 2007, 70, 120–136. [Google Scholar] [CrossRef]
- Xu, Z.; Wan, S.; Zhu, G.; Ren, H.; Han, X. The influence of historical land use and water availability on grassland restoration. Restor. Ecol. 2010, 18, 217–225. [Google Scholar] [CrossRef]
- Chen, C.; Huang, D.; Zhang, Y.; Zheng, H.; Wang, K. Invasion of farmland-grassland ecosystems by the exotic sweet clovers, Melilotus officinalis and M. albus. J. Food Agric. Environ. 2013, 11, 1012–1016. [Google Scholar]
- Birch, J.C.; Newton, A.C.; Aquino, C.A.; Cantarello, E.; Echeverría, C.; Kitzberger, T.; Schiappacasse, L.; Garavito, N.T. Cost-effectiveness of dryland forest restoration evaluated by spatial analysis of ecosystem services. Proc. Natl. Acad. Sci. USA 2010, 107, 21925–21930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menz, M.H.M.; Dixon, K.W.; Hobbs, R.J. Hurdles and opportunities for landscape-scale restoration. Science 2013, 339, 526–527. [Google Scholar] [CrossRef] [PubMed]
- Cowie, A.L.; Orr, B.J.; Castillo Sanchez, V.M.; Chasek, P.; Crossman, N.D.; Erlewein, A.; Louwagie, G.; Maron, M.; Metternicht, G.I.; Minelli, S.; et al. Land in balance: The scientific conceptual framework for Land Degradation Neutrality. Environ. Sci. Policy 2018, 79, 25–35. [Google Scholar] [CrossRef]
- Bai, X.; Jia, X.X.; Jia, Y.H.; Shao, M.A.; Hu, W. Modeling long-term soil water dynamics in response to land-use change in a semi-arid area. J. Hydrol. 2020, 585, 124824. [Google Scholar] [CrossRef]
- Vanek, S.J.; Meza, K.; Ccanto, R.; Olivera, E.; Scurrah, M.; Fonte, S.J. Participatory design of improved forage/follow options across soil gradients with farmers of the Central Peruvian Andes. Agr. Ecosyst. Environ. 2020, 300, 106933. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.J.; Wang, L.X.; Lu, N.; Li, J.Y. Water use characteristics of the common tree species in different plantation types in the Loess Plateau of China. Agr. Forest. Meteorol. 2020, 288–289, 108020. [Google Scholar] [CrossRef]
- Fort, F.; Volaire, F.; Guilioni, L.; Barkaoui, K.; Navas, M.L.; Roumet, C. Root traits are related to plant water-use among rangeland Mediterranean species. Funct. Ecol. 2017, 31, 1700–1709. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Liu, Z.; Zhao, X.; Ling, Q.; Huo, G.; Wu, P. Extreme natural drought enhances interspecific facilitation in semiarid agroforestry systems. Agr. Ecosyst. Environ. 2018, 265, 444–453. [Google Scholar] [CrossRef]
- Sanchez, P.A. Improved fallows come of age in the tropics. Agroforest. Syst. 1999, 47, 3–12. [Google Scholar] [CrossRef]
- Mendez, M.O.; Maier, R.M. Phytostabilization of mine tailings in arid and semiarid environments-an emerging remediation technology. Environ. Health Perspect. 2008, 116, 278–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Loaiza, J.; White, S.A.; Root, R.A.; Solis-Dominguez, F.A.; Hammond, C.M.; Chorover, J.; Maier, R.M. Phytostabilization of mine tailings using compost-assisted direct planting: Translating greenhouse results to the field. Sci. Total Environ. 2016, 565, 451–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.B.; Zhao, W.Y.; Yuan, Y.Q.; Morel, J.L.; Chi, H.C.; Feng, W.L.; Wang, S.Z.; Zhang, J.X.; Feng, Z.K.; Tan, H.R.; et al. Mobility of metal(loid)s in Pb/Zn tailings under different revegetation strategies. J. Environ. Manag. 2020, 263, 110323. [Google Scholar] [CrossRef] [PubMed]
- Yin, G.D.; Wang, X.; Zhang, X.; Fu, Y.S.; Hao, F.H.; Hu, Q.H. InVEST model-based estimation of water yield in North China and its sensitivities to climate variables. Water 2020, 12, 1692. [Google Scholar] [CrossRef]
- Shen, C.; Du, H.L.; Chen, Z.; Lu, H.W.; Zhu, F.G.; Chen, H.; Meng, X.Z.; Liu, Q.W.; Liu, P.; Zheng, L.H.; et al. The Chromosome-Level Genome Sequence of the Autotetraploid Alfalfa and Resequencing of Core Germplasms Provide Genomic Resources for Alfalfa Research. Mol. Plant. 2020, 13, 1250–1261. [Google Scholar] [CrossRef]
- Lin, G.H.; Sternberg, L.D.S.L. Utilization of surface water by red mangrove (Rhizophora mangle L.): An isotopic study. B. Mar. Sci. 1994, 54, 94–102. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Meinzer, F.C.; Andrade, J.L.; Goldstein, G.; Holbrook, N.M.; Cavelier, J.; Wright, S.J. Partitioning of soil water among canopy trees in a seasonally dry tropical forest. Oecologia 1999, 121, 293–301. [Google Scholar] [CrossRef]
- Moreno-Gutierrez, C.; Dawson, T.E.; Nicolas, E.; Querejeta, J.I. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New. Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Wen, X.F.; Sun, X.M. Seasonal variations in depth of water uptake for a subtropical coniferous plantation subjected to drought in an East Asian monsoon region. Agric. Forest. Meteorol. 2015, 201, 218–228. [Google Scholar] [CrossRef]
- Dawson, T.E.; Ehleringer, J.R. Streamside trees that do not use stream water. Nature 1991, 350, 335–337. [Google Scholar] [CrossRef]
- Dawson, T.E.; Mambelli, S.; Plamboeck, A.H.; Templer, P.H.; Tu, K.P. Stable isotopes in plant ecology. Annu. Rev. Ecol. Evol. S. 2002, 33, 507–559. [Google Scholar] [CrossRef]
- David, T.S.; Pinto, C.A.; Nadezhdina, N.; Kurz-Besson, C.; Henriques, M.O.; Quilho, T. Root functioning, tree water use and hydraulic redistribution in Quercus suber trees: A modeling approach based on root sap flow. For. Ecol. Manag. 2013, 307, 136–146. [Google Scholar] [CrossRef] [Green Version]
- Evaristo, J.; McDonnell, J.J.; Scholl, M.A.; Bruijnzeel, L.A.; Chun, K.P. Insights into plant water uptake from xylem-water isotope measurements in two tropical catchments with contrasting moisture conditions. Hydrol. Process. 2016, 30, 3210–3227. [Google Scholar] [CrossRef]
- Meissner, M.; Kohler, M.; Schwendenmann, L.; Holscher, D.; Dyckmans, J. Soil water uptake by trees using water stable isotopes (δ2H and δ18O)-a method test regarding soil moisture, texture and carbonate. Plant Soil 2014, 376, 327–335. [Google Scholar] [CrossRef]
- Song, L.N.; Zhu, J.J.; Li, M.C.; Zhang, J.X.; Lv, L.Y. Sources of water used by Pinus sylvestris var. mongolica trees based on stable isotope measurements in a semiarid sandy region of Northeast China. Agric. Water Manag. 2016, 164, 281–290. [Google Scholar] [CrossRef]
- Ma, Y.; Song, X.F. Using stable isotopes to determine seasonal variations in water uptake of summer maize under different fertilization treatments. Sci. Total Environ. 2016, 550, 471–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Song, X.F.; Han, D.M.; Zhang, Y.H.; Liu, X. A study of root water uptake of crops indicated by hydrogen and oxygen stable isotopes: A case in Shanxi Province, China. Agric. Water Manag. 2010, 97, 475–482. [Google Scholar] [CrossRef]
- Wu, Y.J.; Du, T.S.; Li, F.S.; Li, S.E.; Ding, R.S.; Tong, L. Quantification of maize water uptake from different layers and root zones under alternate furrow irrigation using stable oxygen isotope. Agric. Water Manag. 2016, 168, 35–44. [Google Scholar] [CrossRef]
- Zhao, X.; Li, F.D.; Ai, Z.P.; Li, J.; Gu, C.K. Stable isotope evidences for identifying crop water uptake in a typical winter wheat-summer maize rotation field in the North China Plain. Sci. Total Environ. 2018, 618, 121–131. [Google Scholar] [CrossRef]
- Dai, Y.; Zheng, X.J.; Tang, L.S.; Li, Y. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant Soil 2015, 389, 73–87. [Google Scholar] [CrossRef]
- Wu, Y.; Zhou, H.; Zheng, X.J.; Li, Y.; Tang, L.S. Seasonal changes in the water use strategies of three co-occurring desert shrubs. Hydrol. Process. 2014, 28, 6265–6275. [Google Scholar] [CrossRef]
- Bachmann, D.; Gockele, A.; Ravenek, J.M.; Roscher, C.; Strecker, T.; Weigelt, A.; Buchmann, N. No evidence of complementary water use along a plant species richness gradient in temperate experimental grasslands. PLoS ONE 2015, 10, e0116367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirl, R.T.; Schnyder, H.; Ostler, U.; Schaufele, R.; Schleip, I.; Vetter, S.H.; Auerswald, K.; Cabrera, J.C.B.; Wingate, L.; Barbour, M.M.; et al. The 18O ecohydrology of a grassland ecosystem—Predictions and observations. Hydrol. Earth Syst. Sci. 2019, 23, 2581–2600. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.W.; Li, X.Y.; Li, J.; Jiang, Z.Y.; Chen, H.Y.; Ma, Y.J.; Huang, Y.M. Differential soil moisture pulse uptake by coexisting plants in an alpine Achnatherum splendens grassland community. Environ. Earth. Sci. 2016, 75, 914. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.J.; Lu, N.; Zhang, L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Sci. Total. Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Fu, B.J.; Lu, N.; Wang, S.; Zhang, L. Water use characteristics of native and exotic shrub species in the semi-arid Loess Plateau using an isotope technique. Agriv. Ecosyst. Environ. 2019, 276, 55–63. [Google Scholar] [CrossRef]
- Wu, J.Y.; Pang, Z.; Sun, T.J.; Kan, H.M.; Hu, W.; Li, X.N. Soil respiration simulation based on soil temperature and water content in artificial smooth brome grassland. Rangeland J. 2016, 38, 579–589. [Google Scholar] [CrossRef]
- Schultz, N.M.; Griffifis, T.J.; Lee, X.H.; Baker, J.M. Identification and correction of spectral contamination in H2/H1 and O18/O16 measured in leaf, stem, and soil water. Rapid Commun. Mass Spectrom. 2011, 25, 3360–3368. [Google Scholar] [CrossRef]
- Stock, B.C.; Semmens, B.X. MixSIAR GUI User Manual, version 3.1. 2016. Available online: https://github.com/brianstock/MixSIAR (accessed on 20 November 2020).
- Wang, J.; Lu, N.; Fu, B. Inter-comparison of stable isotope mixing models for determining plant water source partitioning. Sci. Total. Environ. 2019, 666, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Craig, H. Isotopic variations in meteoric waters. Science 1961, 133, 1702–1703. [Google Scholar] [CrossRef] [PubMed]
- Ehleringer, J.R.; Dawson, T.E. Water uptake by plants-perspectives from stable isotope composition. Plant Cell Environ. 1992, 15, 1073–1082. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Ehleringer, J.R. Stable isotope composition of stem and leaf water: Applications to the study of plant water use. Funct. Ecol. 1991, 5, 270–277. [Google Scholar] [CrossRef]
- Yang, Y.; Fu, B. Soil water migration in the unsaturated zone of semiarid region in China from isotope evidence. Hydrol. Earth. Syst. Sci. 2017, 21, 1757–1767. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Guo, L.; Huang, Z.; Lopez-Vicente, M.; Wu, G.L. Root morphological characteristics and soil water infiltration capacity in semi-arid artificial grassland soils. Agr. Water Manage. 2020, 235, 106153. [Google Scholar] [CrossRef]
- Wang, G.Y.; Liu, S.; Fang, Y.; Shangguan, Z.P. Adaptive changes in root morphological traits of Gramineae and Leguminosae seedlings in the ecological restoration of the semiarid region of northwest China. Land Degrad. Dev. 2020, 31, 2417–2429. [Google Scholar] [CrossRef]
- Bockstette, S.W.; Pinno, B.D.; Dyck, M.F.; Landhausser, S.M. Root competition, not soil compaction, restricts access to soil resources for aspen on a reclaimed mine soil. Botany 2017, 95, 385–395. [Google Scholar] [CrossRef]
- Li, Z.; Lamb, E.G.; Piper, C.L.; Siciliano, S.D. Plant belowground diversity and species segregation by depth in a semi-arid grassland. Ecoscience 2018, 25, 1–7. [Google Scholar] [CrossRef]
- Bai, W.M.; Zuo, Q.; Huang, Y.F.; Li, B.G. Effect of water supply on root growth and water uptake of alfalfa in Wulanbuhe sandy region. Acta Phytoecol. Sin. 2001, 25, 29–34. (In Chinese) [Google Scholar] [CrossRef]
- Otfinowski, R.; Kenkel, N.C.; Catling, P.M. The biology of Canadian weeds. 134. Bromus inermis Leyss. Can. J. Plant Sci. 2007, 87, 183–198. [Google Scholar] [CrossRef] [Green Version]
- Silvertown, J.; Araya, Y.; Gowing, D.; Cornwell, W. Hydrological niches in terrestrial plant communities: A review. J. Ecol. 2015, 103, 93–108. [Google Scholar] [CrossRef] [Green Version]
- Brooks, J.R.; Barnard, H.R.; Coulombe, R.; McDonnell, J.J. Ecohydrologic separation of water between trees and streams in a Mediterranean climate. Nat. Geosci. 2009, 3, 100–104. [Google Scholar] [CrossRef]
- Goldsmith, G.R.; Munoz-Villers, L.E.; Holwerda, F.; McDonnell, J.J.; Asbjornsen, H.; Dawson, T.E. Stable isotopes reveal linkages among ecohydrological processes in a seasonally dry tropical montane cloud forest. Ecohydrology 2012, 5, 779–790. [Google Scholar] [CrossRef]
- Evaristo, J.; Jasechko, S.; McDonnell, J.J. Global separation of plant transpiration from groundwater and streamflow. Nature 2015, 525, 91–94. [Google Scholar] [CrossRef] [PubMed]
- Eynard, A.; Schumacher, T.E.; Lindstrom, M.J.; Malo, D.D.; Kohl, R.A. Effects of aggregate structure and organic C on wettability of Ustolls. Soil Till. Res. 2006, 88, 205–216. [Google Scholar] [CrossRef]
- Fischer, C.; Tischer, J.; Roscher, C.; Eisenhauer, N.; Ravenek, J.; Gleixner, G.; Attinger, S.; Jensen, B.; de Kroon, H.; Mommer, L.; et al. Plant species diversity affects infiltration capacity in an experimental grassland through changes in soil properties. Plant Soil 2015, 397, 1–16. [Google Scholar] [CrossRef]
- Archer, N.A.L.; Quinton, J.N.; Hess, T.M. Below-ground relationships of soil texture, roots and hydraulic conductivity in two-phase mosaic vegetation in south-east Spain. J. Arid Environ. 2002, 52, 535–553. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Roscher, C.; Jensen, B.; Eisenhauer, N.; Baade, J.; Attinger, S.; Scheu, S.; Weisser, W.W.; Schumacher, J.; Hildebrandt, A. How do earthworms, soil texture and plant composition affect infiltration along an experimental plant diversity gradient in grassland? PLoS ONE 2014, 9, e98987. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.L.; Zhang, Z.N.; Wang, D.; Shi, Z.H.; Zhu, Y.J. Interactions of soil water content heterogeneity and species diversity patterns in semi-arid steppes on the Loess Plateau of China. J. Hydrol. 2014, 519, 1362–1367. [Google Scholar] [CrossRef]
- Gardner, W.R. Some steady state solutions of the unsaturated moisture flow equation with application to evaporation from a water table. Soil Sci. 1958, 85, 228–232. [Google Scholar] [CrossRef]
- Liu, J.H.; Gao, J.X.; Lv, S.H.; Han, Y.W.; Nie, Y.H. Shifting farming-pastoral ecotone in China under climate and land use changes. J. Arid. Environ. 2011, 75, 298–308. [Google Scholar] [CrossRef]
- Qian, W.H.; Lin, X.; Zhu, Y.F.; Xu, Y.; Fu, J.L. Climatic regime shift and decadal anomalous events in China. Clim. Chang. 2007, 84, 167–189. [Google Scholar] [CrossRef]
- Wu, Z.T.; Wu, J.J.; Liu, J.H.; He, B.; Lei, T.J.; Wang, Q.F. Increasing terrestrial vegetation activity of ecological restoration program in the Beijing–Tianjin Sand Source Region of China. Ecol. Eng. 2013, 52, 37–50. [Google Scholar] [CrossRef]
- Wei, B.C.; Xie, Y.W.; Jia, X.; Wang, X.Y.; He, H.J.; Xue, X.Y. Land use/land cover change and it’s impacts on diurnal temperature range over the agricultural pastoral ecotone of Northern China. Land Degrad. Dev. 2018, 29, 3009–3020. [Google Scholar] [CrossRef]
- Mao, D.H.; Wang, Z.M.; Wu, B.F.; Zeng, Y.; Luo, L.; Zhang, B. Land degradation and restoration in the arid and semiarid zones of China: Quantified evidence and implications from satellites. Land Degrad. Dev. 2018, 29, 3841–3851. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, B.; Xu, X.; Tian, J.; He, C. Regional water-energy cycle response to land use/cover change in the agro-pastoral ecotone, Northwest China. J. Hydrol. 2019, 580, 124246. [Google Scholar] [CrossRef]
- Yao, C.; Yang, S.; Qian, W.H.; Lin, Z.M.; Wen, M. Regional summer precipitation events in Asia and their changes in the past decades. J. Geophys. Res. 2008, 113, D17107. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Geruo, A.; Zhang, J.E.; Velicogna, I.; Liang, C.Z.; Li, Z.Y. Ecological restoration impact on total terrestrial water storage. Nat. Sustain. 2020, 4, 56–62. [Google Scholar] [CrossRef]
- Wang, Y.J.; Song, X.F.; Ma, Y.; Zhang, Y.H.; Zheng, F.D.; Yang, L.H.; Bu, H.M. Characterizing the hydrogen and oxygen isotopic compositions of different waters at reclaimed water irrigated district in southeast suburb of Beijing. Geogr. Res. 2017, 36, 361–372. (In Chinese) [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Grassland Types | Month | Soil Volumetric Water Content, SWC (%) | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 5 cm | 10 cm | 15 cm | 20 cm | 25 cm | |||||||||||||||||
| Max | Min | Avg | SD | Max | Min | Avg | SD | Max | Min | Avg | SD | Max | Min | Avg | SD | Max | Min | Avg | SD | ||
| Bromus inermis | May | 25.6 | 12.1 | 17.9 aAA | 3.7 | 19.6 | 13.2 | 15.2 bAA | 1.7 | 23.0 | 16.6 | 18.5 aAA | 1.7 | 28.5 | 21.9 | 24.2 cAA | 1.9 | 19.5 | 16.4 | 17.5 aAA | 1.0 |
| June | 22.7 | 8.8 | 12.8 aBA | 3.6 | 15.4 | 10.7 | 12.1 aBA | 1.3 | 18.2 | 14.0 | 15.2 bBA | 1.3 | 24.2 | 19.0 | 20.8 cBA | 1.7 | 17.9 | 15.2 | 16.4 dBA | 0.8 | |
| July | 28.4 | 16.2 | 23.6 aCA | 3.2 | 22.8 | 11.6 | 18.5 bCA | 2.9 | 25.7 | 14.0 | 21.6 cCA | 3.4 | 30.0 | 18.8 | 26.3 dCA | 4.0 | 23.1 | 15.0 | 19.0 bCA | 2.9 | |
| August | 26.6 | 13.8 | 19.7 aAA | 4.7 | 20.5 | 12.2 | 16.1 bAA | 2.9 | 23.7 | 14.7 | 19.6 aAA | 3.0 | 28.8 | 20.0 | 25.4 cACA | 2.7 | 20.3 | 16.9 | 18.9 aCA | 0.9 | |
| September | 26.0 | 11.0 | 15.4 aDA | 3.8 | 18.7 | 10.2 | 12.3 bBA | 2.2 | 20.3 | 12.1 | 14.3 aBA | 2.2 | 24.6 | 15.7 | 18.6 cDA | 2.5 | 16.7 | 12.6 | 14.6 aDA | 1.4 | |
| May–September | 28.4 | 8.8 | 17.9 aA | 5.3 | 22.8 | 10.2 | 14.9 bA | 3.3 | 25.7 | 12.1 | 17.9 aA | 3.6 | 30.0 | 15.7 | 23.1 cA | 3.9 | 23.1 | 12.6 | 17.3 aA | 2.3 | |
| Medicago sativa | May | 16.0 | 5.8 | 9.4 aAB | 3.0 | 16.2 | 6.8 | 9.7 abAB | 2.8 | 16.8 | 7.6 | 10.3 abAB | 2.6 | 16.8 | 8.7 | 10.9 bAB | 2.3 | 22.0 | 13.1 | 16.1 cAB | 2.7 |
| June | 14.7 | 4.5 | 7.7 aBB | 3.1 | 13.9 | 5.0 | 7.2 aBB | 2.5 | 11.9 | 6.3 | 7.5 aBB | 1.5 | 8.7 | 7.7 | 8.1 aBB | 0.3 | 13.0 | 11.9 | 12.3 bBB | 0.3 | |
| July | 18.9 | 11.1 | 15.3 aCB | 1.9 | 20.1 | 9.7 | 15.8 aCB | 2.4 | 21.2 | 8.1 | 16.4 aCB | 3.1 | 21.2 | 8.5 | 16.3 aCB | 3.6 | 29.1 | 12.7 | 22.2 bCB | 6.1 | |
| August | 16.5 | 7.9 | 12.1 aDB | 3.1 | 17.8 | 9.2 | 13.3 aDB | 3.0 | 18.5 | 10.1 | 14.4 bDB | 2.8 | 18.5 | 10.6 | 14.8 bDB | 2.6 | 26.0 | 15.4 | 21.3 cCB | 3.5 | |
| September | 15.4 | 5.8 | 8.9 aABB | 2.5 | 16.3 | 6.9 | 9.8 abAB | 2.5 | 14.9 | 7.8 | 10.4 bAB | 2.0 | 12.2 | 8.4 | 9.8 abAB | 1.1 | 15.2 | 12.8 | 14.0 cBA | 0.8 | |
| May–September | 18.9 | 4.5 | 10.7 aB | 3.9 | 20.1 | 5.0 | 11.2 abB | 4.0 | 21.2 | 6.3 | 11.8 bB | 4.0 | 21.2 | 7.7 | 12.0 bB | 3.8 | 29.1 | 11.9 | 17.2 cA | 5.2 | |
| Plant Species | Sample Type | Soil Depth (cm) | N | δ18O(‰) | δ2H(‰) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Avg | SD | Max | Min | Avg | SD | ||||
| Bromus inermis | Soil water | 0–5 | 16 | 9.39 | −10.76 | −2.98 | 6.73 | −8.06 | −74.07 | −44.96 | 20.86 |
| 5–10 | 19 | 1.95 | −8.49 | −4.14 | 3.77 | −15.72 | −60.92 | −44.00 | 16.53 | ||
| 10–20 | 18 | −1.18 | −9.73 | −5.47 | 2.65 | −29.41 | −68.80 | −50.65 | 13.56 | ||
| 20–30 | 19 | −5.49 | −10.16 | −8.09 | 1.55 | −44.20 | −74.58 | −63.37 | 8.49 | ||
| Total | 72 | 9.39 | −10.76 | −5.26 | 4.37 | −8.06 | −74.58 | −50.99 | 16.91 | ||
| Xylem water | 15 | −0.55 | −8.60 | −4.86 | 3.03 | −39.55 | −69.99 | −55.60 | 12.42 | ||
| Medicago sativa | Soil water | 0–5 | 14 | 9.42 | −10.59 | −4.29 | 5.67 | −9.96 | −74.48 | −47.20 | 18.39 |
| 5–10 | 17 | 5.62 | −8.86 | −4.01 | 4.87 | −11.60 | −66.39 | −44.64 | 18.68 | ||
| 10–20 | 19 | −2.09 | −9.69 | −6.25 | 2.87 | −30.61 | −68.96 | −52.52 | 13.32 | ||
| 20–30 | 19 | −3.68 | −10.54 | −7.90 | 1.64 | −44.59 | −72.64 | −64.08 | 6.86 | ||
| Total | 69 | 9.42 | −10.59 | −5.75 | 4.14 | −9.96 | −74.48 | −52.68 | 16.27 | ||
| Xylem water | 14 | 2.41 | −8.52 | −4.52 | 3.34 | −38.29 | −65.50 | −55.79 | 9.68 | ||
| Groundwater | 16 | −9.96 | −11.48 | −10.51 | 0.38 | −70.99 | −73.51 | −72.06 | 0.76 | ||
| Precipitation | 18 | −1.86 | −14.00 | −7.52 | 2.95 | −15.49 | −100.76 | −49.98 | 21.11 | ||
| Grassland Type | Soil Depth (cm) | N | Soil Organic Carbon Content (g·kg−1) | Soil Capillary Porosity (%) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Max | Min | Avg | SD | Max | Min | Avg | SD | |||
| Bromus inermis | 0–5 | 4 | 22.11 | 16.67 | 20.18 aA | 2.40 | 36.52 | 34.16 | 35.37 A | 1.02 |
| 5–10 | 4 | 17.94 | 15.01 | 16.26 bA | 1.32 | |||||
| 10–20 | 4 | 10.67 | 7.43 | 8.84 cA | 1.53 | |||||
| 20–30 | 4 | 8.53 | 3.02 | 5.28 dA | 2.61 | |||||
| Total | 4 | 22.11 | 3.02 | 12.64 A | 6.35 | |||||
| Medicago sativa | 0–5 | 4 | 29.45 | 21.38 | 26.19 aB | 3.79 | 41.09 | 34.49 | 37.19 A | 2.98 |
| 5–10 | 4 | 24.58 | 17.30 | 21.83 aB | 3.15 | |||||
| 10–20 | 4 | 12.38 | 7.47 | 10.07 bA | 2.01 | |||||
| 20–30 | 4 | 15.51 | 4.17 | 9.96 bA | 6.02 | |||||
| Total | 4 | 29.45 | 4.17 | 17.01 B | 8.23 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pang, Z.; Xu, H.; Chen, C.; Zhang, G.; Fan, X.; Wu, J.; Kan, H. Comparing Water Use Characteristics of Bromus inermis and Medicago sativa Revegetating Degraded Land in Agro-Pasture Ecotone in North China. Water 2023, 15, 55. https://doi.org/10.3390/w15010055
Pang Z, Xu H, Chen C, Zhang G, Fan X, Wu J, Kan H. Comparing Water Use Characteristics of Bromus inermis and Medicago sativa Revegetating Degraded Land in Agro-Pasture Ecotone in North China. Water. 2023; 15(1):55. https://doi.org/10.3390/w15010055
Chicago/Turabian StylePang, Zhuo, Hengkang Xu, Chao Chen, Guofang Zhang, Xifeng Fan, Juying Wu, and Haiming Kan. 2023. "Comparing Water Use Characteristics of Bromus inermis and Medicago sativa Revegetating Degraded Land in Agro-Pasture Ecotone in North China" Water 15, no. 1: 55. https://doi.org/10.3390/w15010055