
Influence of Salinity on Copper Toxicity in Paracentrotus lividus and Arbacia lixula Embryos
by
 ,
,
Lorenzo Morroni
1,*,
Andrea Gaion
1,2,
Andrea Broccoli
1,
Stefano Ferrari
1,
David Pellegrini
1 and
Davide Sartori
1 1
Istituto Superiore per la Protezione Ambientale (ISPRA), Via del Cedro 38, 57126 Livorno, Italy
2
Higher Education Department, University Centre South Devon, Long Road, Paignton TQ4 7EJ, UK
*
Author to whom correspondence should be addressed.
Water 2023, 15(1), 65; https://doi.org/10.3390/w15010065
Submission received: 17 November 2022
/
Revised: 20 December 2022
/
Accepted: 21 December 2022
/
Published: 25 December 2022
(This article belongs to the Special Issue Ecotoxicological Risk in Aquatic Environments)
Abstract
:The sea urchins Paracentrotus lividus and Arbacia lixula have a key role in benthic dynamics and they are often used to assess the toxicity of environmental contaminants. The aim of this research was to investigate the influence of salinity on copper toxicity on P. lividus and A. lixula embryos, evaluating the impact of future ocean salinity variations and estimating an optimum range for embryos. For this purpose, sea urchin embryos were exposed to different copper concentrations under varying salinity regimes, estimating the toxic effects with the percentage of abnormal embryos and the Integrative Toxicity Index (ITI). Results revealed acceptable levels of anomalies in a salinity range of 33–36‰ for A. lixula and 33–39‰ for P. lividus. The effect concentrations (EC50) of copper reveals increasing toxicity at increasing salinity for both sea urchin species, but between 33–36‰ (A. lixula) and 36–39‰ (P. lividus) the toxicity appear inversely related with salinity. This study illustrates the difficulty in relating a biological response to bioavailability, because the uptake and the sensitivity to copper across salinities is influenced by differences in osmoregulation, enhancing our knowledge about optimal salinity ranges and the potential impact of climate change on sea urchin embryo development.
1. Introduction
The purple sea urchin Paracentrotus lividus (Lamarck, 1816) is a widespread species in the Mediterranean Sea and in the north-eastern Atlantic [1], often used as a model organism in ecological and ecotoxicological studies [2]. Despite this echinoid sharing its habitat of shallow waters in rocky coastal areas with the black sea urchin Abracia lixula (Linnaeus, 1758), the ecological niche of the two species appears not to overlap because of different adaptations to physical stress and feeding preferences on hard substrate. P. lividus has been reported to feed mainly on erect algae, seagrass, and particulate matter, while A. lixula has been described as a grazer of encrusting coralline algae and possibly sessile invertebrates [2,3]. In addition, the gap between the test plates could be seen as a discriminant feature which reflects the different microhabitat distribution of the two species: in A. lixula, this gap has dimensional characteristics which make them particularly well fitted to bear the hydrodynamic physical stress on its apical region (vertical or subvertical rocky substrates), whereas in P. lividus, this gap is narrow, in response to the ecological strategy to live in tight cavities in rocks and on horizontal or gently inclined substrates [4].
In the Mediterranean Sea, these two species live in coastal areas where environmental conditions could vary rapidly and dramatically. Changes in salinity, for example, could occur naturally in shallow waters, as a result of river discharge or evaporation when the air temperature is high [5,6,7,8]. In addition, over the next century, the interaction between large-scale changes in ocean circulation and an altered freshwater cycle due to anthropogenic climate change is going to increase the salinity in the Atlantic Ocean and the Mediterranean Sea, with shifts in physiological and biological capabilities to cope with stress in marine species [9]. Indeed, the projected rise in seawater temperature will affect rainfall patterns, ice melting and evaporation [8,10]. Echinoderms are generally affected by changes in salinity, with important disruption of fertilization success [11,12], survival [13,14] and embryonic/larval development [8,15,16]. Results from previous studies demonstrated that in echinoderms, embryological development and following larval transformations are more sensitive stages to salinity changes than subsequent adulthood. Concerning specifically P. lividus embryological development, the optimal salinity range for Mediterranean populations varies between 33 and 35‰ [17,18,19,20], while for the Atlantic population it adjusts between 28 and 34‰ [12,21,22,23]. To the best of our knowledge, there are only two studies which assess the effects of salinity on A. lixula development, with tolerance ranges of 29–39‰ for the south Atlantic Ocean [24] and 29–35.5‰ for the North-Eastern Atlantic [12]. Nevertheless, the tolerance to salinity appears strongly related to the area of origin of the adults, with a natural variability between different locations [25].
In addition to their ecological importance in rocky-bottom ecosystems, embryos of these two species, and in particular P. lividus, are commonly used in marine toxicity tests due to their sensitivity to a wide range of environmental pollutants [26,27,28,29,30]. Among the environmental pollutants, trace metals are the category mainly affected by chemical speciation in the seawater and show a decrease of free cations activity at increasing salinity due to chloro-complexation [31,32]. Considering copper toxicity, several studies have demonstrated that Cu effects tend to be reduced at increasing salinities in different species of fish and invertebrates [33], although some contrasting results are present [34]. In recent decades, different anthropogenic activities, such as mining and smelting, municipal wastes, agricultural, industrial emissions and aquaculture plants, resulted in an increase in environmental Cu concentration [35].
In the literature, different studies focused on Cu toxicity on embryos of A. lixula [36] and P. lividus [37,38,39,40,41], describing copper-induced malformations and developmental delay. Other papers describe the effects of salinity on A. lixula [12,24] and P. lividus well [12,17,18,19,20,21,22,23]. Reading these studies, it is interesting to note that the influences of salinity and copper on sea urchin embryo development are always treated separately.
This study moves to investigate, for the first time, the interaction between Cu and salinity in P. lividus and A. lixula embryos, starting from salinity ranges and Cu concentrations used by other authors, in order to implement and integrate the results of past studies. In particular, the aim of this study was to analyze the influence of salinity on copper toxicity on P. lividus and A. lixula embryos, to understand the impact of future changes in ocean salinity on sea urchin populations and to establish the optimum range for embryo bioassays. To achieve these goals, sea urchin embryos were exposed to different copper concentrations under varying salinity regimes; this approach showed developmental anomalies in a dose-dependent manner in agreement with the literature data [41]. Morphological data were reported with the standard criteria of normal embryos (%) and weighting the severity of developmental anomalies, using the Integrative Toxicity Index (ITI) which allows for a better categorization and classification of the potential teratogenic effects of pollutants [28].
2. Materials and Methods
2.1. Test Solutions
A stock solution of 45‰ salinity was prepared to reach the predetermined salinity by adding hypersaline brine (HSB), obtained by concentrating natural seawater by evaporation to 0.22 µm filtered seawater (FSW), collected offshore in an area far from evident sources of pollutants, already characterized and used in previous studies [28]. Water solutions at different salinities (27, 30, 33, 36, 39, 42, 45‰) were prepared by adding an appropriate volume of Milli-Q® (Millipore Co., Bedford, MA, USA) water to the stock solution. The desired salinity was confirmed with the detection using a salinity probe (Hanna HI98319).
Copper was used as reference toxicant in the embryo-development test with sea urchins. A solution of copper nitrate (Cu(NO3)2 H2O) 10 mg/L was prepared in bi-distilled water (BDW) by a stock solution of 1000 g/L (Sigma Aldrichl, St. Louis, MO, USA). For sea urchin embryo incubation, further dilutions in filtered seawater (FSW) were prepared to obtain the nominal concentrations of 10, 15, 20, 25 µg/L Cu for A. lixula and 10, 20, 25, 30, 35, 40 µg/L for P. lividus. The exposure concentrations were differentiated between the two sea urchin species on the basis of the literature ranges [28,41,42] and after preliminary experiments, with the aim of validating these values.
2.2. Chemical Analysis of Test Solutions
Fresh standard solutions of Cu were prepared before the analysis and were used in bioassays. Three replicates were carried out for each Cu concentration and were analyzed by a graphite furnace atomic absorption spectrophotometer (AAS-GF, Varian Spectra AA-220 Z), following the procedures reported in EPA 7010, to compare the nominal Cu concentration with the obtained solutions. No matrix modifiers were employed. The determination was carried out using a calibration curve between 10 and 100 µg/L. Standard solutions from Sigma Aldrich were analyzed to guarantee quality control, and the percentage of recovery was always between −10% and +10%.
2.3. Sea Urchin Collection and Bioassays
Adult sea urchins of P. lividus and A. lixula were collected from October 2018 to February 2019 in an intertidal rocky site along the southern coast of Livorno, Italy (43°25.6020 N–10° 23.7800 E) and immediately transported in an insulated box to the laboratory. Bioassays were conducted as described by Morroni et al. (2018) [29]. Briefly, sea urchins were induced to spawn by injecting 1 mL of 0.5 M KCl solution (Carlo Erba, Milan, Italy) into the coelom, through the peribuccal membrane. Sperm was collected dry from at least three males using a Pasteur pipette, pooled, and stored at 4 °C until use. For each tested salinity, oocytes obtained from at least three females were shed into 100 mL beaker previously filled with 0.22 µm filtered seawater (FSW) at requested salinity. As per internal Quality Assurance in our laboratory, eggs were examined under an Olympus GX53 inverted microscope (Tokyo, Japan) before conducting the experiments to determine their maturity, and were then pooled into a 500 mL beaker. The final concentration of 1000 eggs/mL was prepared by counting 100 μL subsamples with an inverted microscope at 4× objectives.
Once sperm motility had been checked, 10 µL of sperm was gently diluted in 500 mL eggs suspension (1000 eggs/mL). A few minutes after fertilization, an aliquot of embryos was observed to verify the success of fertilization (formation of the fertilization envelope > 95%) [41].
After fertilization, for each tested salinity and sea urchin species, 1 mL of embryo suspension (1000 embryos) was exposed to untreated negative controls (0.22 µm FSW) and positive controls with exposure to copper nitrate (reference toxicant) at the following Cu nominal concentrations: 10, 20, 25, 30, 35, 40 µg/L. Working solution were obtained by diluting a stock solution of 1 g/L of Cu(NO3)2*3H2O (Sigma Aldrich, St. Louis, MO, USA). Six replicates were prepared for each concentration using sterile capped polystyrene microplates. Embryos were incubated in a dark room at 18 °C for 72 h; at the end of incubation, samples were fixed with 1% Lugol solution, and observed with transmitted light microscopy. Morphological anomalies were monitored counting 100 embryos per replica under a light inverted microscope (Leica DMI 3000B, Milan, Italy). The acceptability of the results was fixed at a percentage of normal plutei ≥80% in the negative control test [35].
2.4. Toxicity Criteria
The toxicity of Cu at different salinities was estimated by calculating the percentage of abnormal embryos at the pluteus stage (72 h of development), according to the standard criteria and the Integrative Toxicity Index (ITI) [28,43]. The standard criteria calculate the percentage of normal and abnormal embryos without considering different typologies of malformations or the phase in which they appear. Conversely, all the ITI methodologies count the frequency of delayed and/or abnormal embryonic morphologies and quantitatively rank the severity of the effects. Embryos were classified as normal if they satisfied the morphological criteria as follows: (i) suitable schedule in reaching the developmental endpoint (pluteus at 72 h); (ii) correct skeleton development and patterning; (iii) right ectoderm, mesoderm, and endoderm germ layer differentiation; (iv) conform left/right or dorso/ventral axis symmetry. On the other hand, embryos displaying impairment of the axis symmetry, as well as germ layer defects, were marked as abnormal [28]. In this study, ITI 4.0 [43] was applied and validated for the first time for A. lixula embryos also.
In particular, ITI 4.0 classifies embryos on a toxicity scale from 0 (absence of toxicity) to 5 (maximum toxicity). The lowest level (0) assigned to each ITI was associated with a “no effect” or light delay in development, including normal plutei and early plutei. A score of 1 was associated with the pluteus and early pluteus stage displaying malformations, a score of 2 was associated with the stage of the prism, and a score of 3 with the prism displaying malformations. The highest levels of toxicity (4 and 5) were associated with the delayed and/or malformed embryos at the gastrula stage, and with the delayed and/or malformed embryos from fertilized egg to the blastula stage [43].
The ITIs applied in this study are calculated as follows:
where Si is the score associated with each abnormal embryonic morphology and Fi is the frequency observed for that abnormality expressed as a percentage (n = 6).
ITI = ∑ni = 1 (Si × Fi)/100,
2.5. Data Analysis
The median Effective Concentration (EC50) inducing abnormal larvae was considered to evaluate toxic effects of Cu. The EC values with 95% confidence limits were estimated using the MOSAIC web-interface for statistical analyses in ecotoxicology [44]. The calculations within MOSAIC are based on the R package ‘morse’ [45].
Embryo development anomalies related to the Cu-exposure at different salinities was examined using two-way ANOVA (copper concentration, salinity) after checking the normality and homoscedasticity of data, the former using a Shapiro–Wilk test and the latter being tested through a Cochran’s C-test [46]. Significant differences between the percentage of abnormal larvae in FSW (control) and in the reference substance solutions were determined by Dunnett’s test for multiple comparison [26,47]. Two levels of significance were established: p < 0.05 and p < 0.01. Significance was established at 95% (p < 0.05). The statistical tests were performed using GMAV 5.0 software (University of Sydney, Australia) and PAST statistical software [48].
3. Results
Copper concentrations, measured in each test solution, differed by 3–7.8% from the nominal concentrations.
ANOVA results showed significant effects on embryo development related to the factor salinity, copper nitrate concentration and their interaction (Table 1); post hoc pairwise comparison indicated significant differences between salinity at all copper concentrations (p < 0.01).
3.1. Paracentrotus lividus
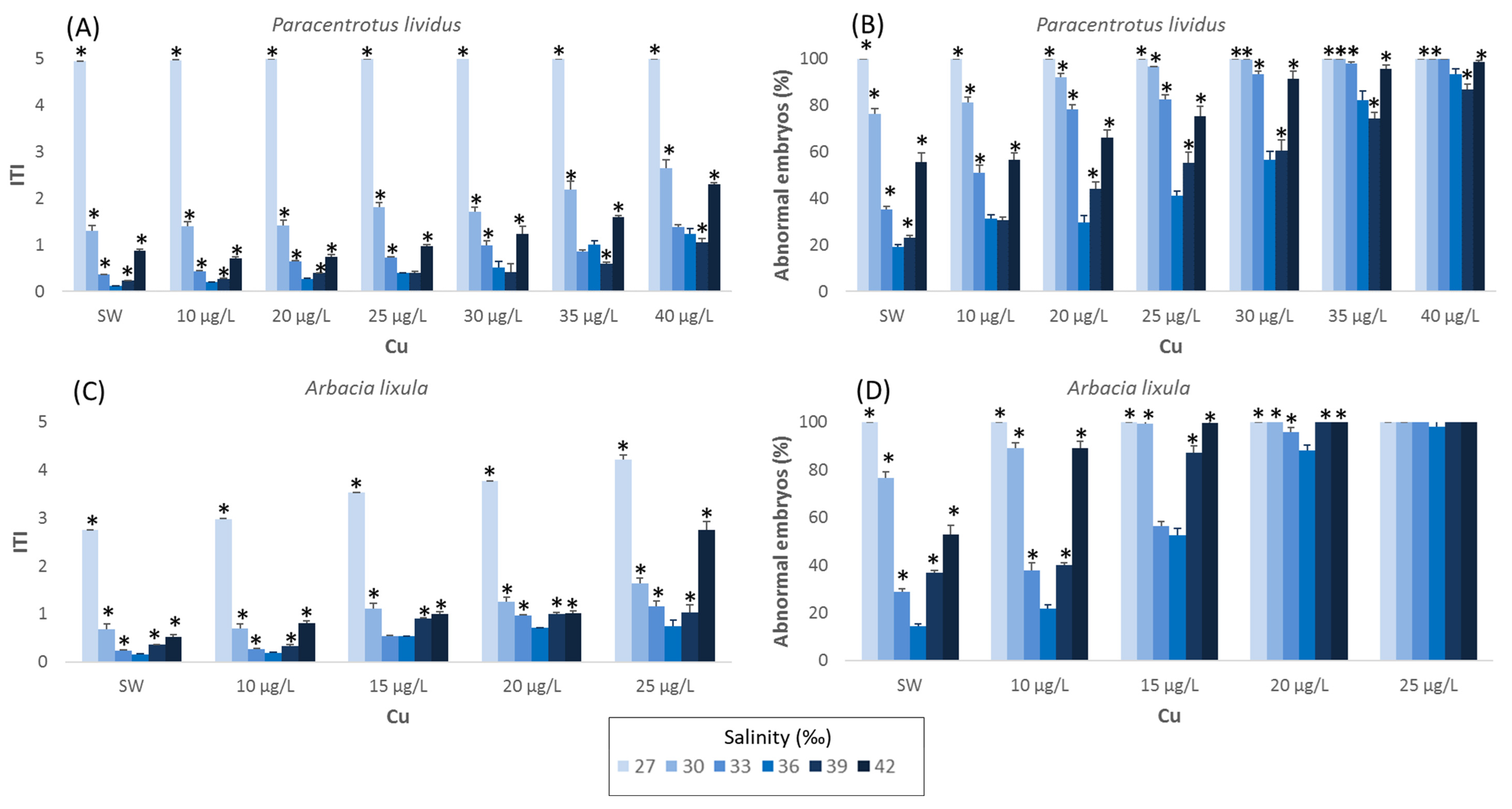
The effects of salinity on larval development of P. lividus are shown in further detail in Figure 1. In the range of salinities tested (27–45‰), an acceptable level of anomalies (abnormal embryos < 30%; ITI < 0.36) was observed at salinity ranging from 33 to 36‰. The lowest percentage of abnormal plutei (14.4%) occurred at a salinity of 36‰, corresponding to ITI values of 0.11. Correct larval development in P. lividus was achieved even at 39‰ (23% of abnormal larvae and 0.96 of ITI), although developmental endpoints differ significantly (p < 0.01) from the percentage obtained at 36‰. At a salinity of 27‰ and higher, of 42‰, larval growth was completely blocked at the early larval stage either in the negative (FSW) and positive (copper solution) control, with ITI values higher than 2 manly due to undeveloped and malformed embryos.
Similar to what was reported for A. lixula, the developmental anomalies of P. lividus in the negative control increased significantly in the range between 27 and 36‰ (p < 0.01). Abnormal embryos were observed, rising to 19% (ITI of 0.91) for a salinity of 36‰. The EC50 value obtained with copper ranged from 10.7 (8.3–12.9) µg/L to 28.8 (28.4–29.3) µg/L for a salinity of 33 and 36‰, respectively. At a salinity of 39‰, the percentage of well-developed plutei remained above 80%, with an EC50 value of 24.7 (23.6–25.7) µg/L for copper nitrate (Table 1).
Aberrations in P. lividus embryos increased significantly (Figure 1) up to 60% at 42‰; tests conducted with the reference toxicant showed a percentage of malformations in larvae exceeding 90% starting from the concentration of 25 µg/L with an EC50 value of 21.7 (19.3–23.8) µg/L.
3.2. Arbacia lixula
The effects of salinity on larval development of P. lividus are shown in further detail in Figure 1. In the range of salinities tested (27–45‰), an acceptable level of anomalies (abnormal embryos < 30%; ITI < 0.36) was observed at salinity ranging from 33 to 39‰. The lowest percentage of abnormal plutei for A. lixula (14.4%) occurred at the salinity of 36‰, corresponding to ITI values of 0.16. At salinity of 27‰ and higher of 42‰, larval growth was completely blocked at early larval stage, either in the negative (FSW) or positive control, with ITI values higher than 2, mainly due to undeveloped and malformed embryos.
Outcomes from A. lixula showed, in FSW, a high percentage (77%) of well-developed plutei even at 30‰, whereas at 33‰, the malformed plutei observed was lower than 30% (ITI of 0.24) Testing the effect of reference toxicant at this salinity, the percentage of malformed plutei was higher than 90%, starting from a copper concentration of 20 µg/L (ITI of 0.96). At 36‰, the percentage of malformed plutei in FSW decreases significantly (p < 0.01) compared to those observed at 30‰ and 33‰, although the sensitivity of sea urchin embryos versus copper remains similar to those obtained at lower salinities (15.5 µg/L, Figure 1).
With a further increase in salinity (39‰), the percentage of malformed plutei in FSW decreased significantly (p < 0.05) with respect to 36‰. At 39‰ the adverse effects on embryo development started from a concentration of 20 µg/L, with 87% of abnormal embryos (ITI of 0.33). The EC50 value for copper at 39‰ drops to 13.6 (13.3–13.9) µg/L (Table 2). Embryonic development of A. lixula was found to be strongly affected at 42‰; the percentage of well-developed plutei in the negative control fell below 50%. Tests carried out with reference toxicant showed a percentage of malformed larvae ranging between 89 and 100% for all tested concentrations of copper.
4. Discussion
Previous results have shown that larvae of many sea urchin species are stenohaline, and their survival and growth are significantly influenced by salinity changes [11,17,24,49,50]. Concerning the literature data, Bressan et al. (1995) [17] found that the optimal salinities range for P. lividus embryonic development was 34–35‰.
Those data were confirmed by Saco-Álvarez et al. (2010) [23], who reported an ideal range of salinities for P. lividus embryo development and early larval growth within 31‰ and 35‰, whereas Carballera et al. (2011) [12], studying the effect of salinities on fertilization and larval development on sea urchin collected in the Bay of Algeciras (Western Mediterranean Sea), recorded an optimal larval development (≥90%) for salinities ranging from 29 to 33‰.
Data were confirmed for the Atlantic specimen of P. lividus by Pétinay et al. (2009) [22], who established, for the execution of the embryo development test, a salinity ranging between 28 and 34‰. Our study confirms these values, as we obtained a correct embryonic development for P. lividus at salinities ranging from 33 to 36‰.
Contrary to what was reported in the literature for A. lixula [24,26], the outcome of our study highlighted a narrower range of salinities tolerance, ranging from 33 to 36‰. The stenohaline attitude of both sea urchins reported in our trials was confirmed by the sharp decrease in the percentage of well-developed embryos passing from 33 to 27‰ (30% and 23% for P. lividus and A. lixula, respectively).
Carballera et al. (2011) [26] observed, in both species, significantly lower (p < 0.05) percentage of normal embryos for salinities < 29‰, although the percentage of normal embryos observed at 25.5‰ was greater (>50%) than those obtained in our trials at 30‰. To explain this discrepancy, it is possible to hypothesize the presence of genetic differences linked to the spatial variability of populations within the Mediterranean basin, as already demonstrated, by the different sensitivity to heavy metals of P. lividus and A. lixula embryos obtained from specimens collected from geographically distant areas [41]. Analyzing the genetic sequences of the P. lividus population within the Mediterranean, Carreras et al. (2019) [51] highlighted differences in allele composition involved in processes related to temperature and salinity, revealing a progressive adaptation to salinity from west to east, and that the populations of western Mediterranean areas are differentiated from those in the eastern Mediterranean [51].
The effects of salinity on the larval development of sea urchin were reported for many species. In Evechinus chloroticus fertilization rate, development rate, gastrulation, and normal development were significantly affected by low salinity (29‰) compared to those obtained at 34, 35 and 37‰ [8]. Sarifudin et al. (2014; 2017) [52,53] found that Diadema setosum and Salmacis sphaeroides successfully grow and develop within the salinity range from 28 to 37‰ and from 25 to 35‰, respectively. A similar optimal range of salinity (28 to 34‰) was observed in Tripneustes gratilla by Shamim Parvez et al. (2018) [54], whereas Greenwood and Bennet (1981) [55], studying the effect of 45 temperature–salinity combinations on the fertilization success of the gametes of the sea urchin Parechinus angulosus, observed that temperature had the greatest effect on sperm, whereas low salinities relevantly affected the ova, but had little apparent effect on sperm. Even in Heliocidaris crassispina, low salinity and high temperature had interactive effects in reducing larval survivorship [16], whereas Metaxas (1998) [56] reported for Echinometra lucunter short-term (24 h) and long-term (over the entire developmental period) effects of salinity on larval survival and rate of development. In the short term, the majority of larvae could survive brief periods of reduced salinity, whereas for extended periods, low salinity had more adverse effects, causing 100% of mortality.
Concerning the sensitivity to copper, in the optimal range of salinities for the larval development (36–39‰), tests with P. lividus showed good repeatability, highlighting an EC50 value ranging from 24.7 (23.6–25.7) to 28.8 (28.4–29.3) µg/L. Data were consistent with the values reported in the literature for this specimen [25,36,37,57,58,59,60,61,62,63], whereas for A. lixula, an EC50 value of 15.5 (15.1–15.8) µg/L was observed at 36‰. The greater sensitivity of A. lixula reported in a previous study [41] was confirmed by our trials, although the EC50 values obtained this time for copper nitrate, both for P. lividus and A. lixula, were lower. To our knowledge, only one study has focused on the sensitivity of A. lixula to copper, obtaining an EC50 value ranging between 46.0 and 53.0 µg/L [51]. This discrepancy could refer both to the different chemical standard employed in the experiment, CuSO4*5H2O instead Cu(NO3)2*3H2O, or to genetic differences between the Atlantic and Mediterranean population of A. lixula, although the order of magnitude of these data was in good agreement to what was observed in our previous study [41].
It is interesting to notice that for P. lividus, the percentage of malformed plutei observed in the presence of the reference toxicant at a salinity of 39‰ was lower than those found at a salinity of 36‰. This trend was also observed in A. lixula, albeit with less intensity, at salinities of 36 and 33‰. The toxicity of some heavy metals depends on chemical speciation and is modified by the trace metal’s ability to complex with other ions and compounds [64,65,66]. It is well known that, for example, the toxicity of Cu2+ is moderated by organic and inorganic chelators, such as humic substances, naturally present in many aquatic systems [67,68,69,70]; these compounds have carboxylic and phenolic groups which can complex trace metals and lower their toxicity [71,72].
In general, the toxicity of trace metals to invertebrates has been found to be inversely related to salinity [33,73,74,75,76,77,78,79], probably related to the increased bioavailability of metal ions (toxic form) at low salinity [72].
In the blue crab Callinectes sapidus, the toxicity of copper is higher at a salinity of 2‰ (96 h-LC50 = 5.3 (3.50–8.05) µM Cu) than at 30‰ (96 h-LC50 = 53.0 (27.39–102.52) µM Cu) [76]. Similarly, the estuarine crabs Cancer irroratus and Carcinus maenus exposed to increasing copper concentrations showed a much more pronounced reduction in osmoregulation function at lower salinity [80]. Studies conducted on Crassostrea gigas, Crassostrea Virginica and Mytilus edulis showed an increase in copper toxicity at lower salinity which could not be accounted for by osmotic stress alone [73,81,82]. Additionally, Riba et al. (2004) [75] noted that the toxicity of the heavy metals measured in Ruditapes philippinarum exposed to sediments rich in Zn, Cd, Pb, and Cu increased at low values of pH (6.5) and salinity (10‰). Moreover, Moreira et al. (2018) [79] reported an inverse correlation between the increase of arsenic toxicity and the salinity reduction for Crassostrea angulata and Crassostrea gigas. Other studies conducted on the American lobster (Homarus americanus) exposed to 560 µg/L at 13 °C [83], the striped bass (Morone saxatilis) [84], the polychaete, Nereis diversicolor [85,86], the clam, Rangia cuneata [87], the pompano, Trachinotus carolinus [88] and the larval topsmelt, Atherinops affinis [89] converge to the same statement: the highest sensitivity occurs at the lowest salinity. Furthermore, not all marine species exhibit increased metal toxicity associated with decreased salinity. Deruytter et al. (2015) [90] reported that in marine mussel larvae (Mytilus galloprovincialis), an increase in Cu toxicity (modeled with dissolved Cu or Cu2+ activity) was observed with increasing salinity, suggesting a salinity-induced alteration in the physiology of the mussel larvae. Similar results have been reported for the marine gastropod Hydrobia ventrosa, which exhibited a greater reduction in growth in the presence of cadmium at a salinity of 33‰ than at a salinity of 13‰ (Forbes, 1991) [91].
The results of this study illustrate the difficulty in relating a biological response (e.g., growth or mortality) to bioavailability, because the uptake and the relative sensitivity to trace metal across salinities could be influenced by species-specific differences in osmoregulation [92,93]. To the best of our knowledge, this is the first publication assessing the influence of different salinities on copper toxicity for P. lividus and A. lixula. The correlation observed in literature studies between lower salinities and the higher toxicity of heavy metal seems to confirm the goodness of our results. Lastly, our study introduces new knowledge about the possible impact climate change has on sea urchins, and consequently on the whole trophic net.
Author Contributions
Conceptualization, L.M. and D.S.; methodology, L.M.; validation, L.M. and D.S.; formal analysis, L.M.; investigation, L.M., A.B. and S.F.; data curation, L.M.; writing—original draft preparation, L.M. and D.S; writing—review and editing, A.G., D.S., A.B. and D.P.; supervision, D.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are available from the corresponding author upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Boudoureque, C.F.; Verlaque, M. Paracentrotus lividus. In Developments in Aquaculture and Fisheries Science; Elsevier: Amsterdam, The Netherlands, 2020; pp. 447–485. [Google Scholar]
- Pagano, G.; Thomas, P.J.; Guida, M.; Palumbo, A.; Romano, G.; Trifuoggi, M.; Oral, R.; Trifuoggi, M. Sea urchin bioassays in toxicity testing: II. Sediment evaluation. Expert Opin. Environ. Biol. 2017, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Tuya, F.; Cisneros-Aguirre, J.; Ortega-Borges, L.; Haroun, R.J. Bathymetric segregation of sea urchins on reefs of the Canarian Archipelago: Role of flow-induced forces. Estuar. Coast. Shelf Sci. 2007, 73, 481–488. [Google Scholar] [CrossRef]
- Régis, M.B. Particularités microstructurales du squelette de Paracentrotus lividus et Arbacia lixula: Rapports avec l’écologie et l’éthologie de ces échinoïdes. Mar. Biol. 1979, 54, 373–382. [Google Scholar] [CrossRef]
- Lawrence, J.M. The effect of temperature-salinity combinations on the functional well- being of adult Lytechinus variegatus (Lamarck) (Echinodermata, Echinoidea). J. Exp. Mar. Biol. Ecol. 1975, 18, 271–275. [Google Scholar] [CrossRef]
- Trowbridge, C.D. Life at the edge: Population dynamics and salinity tolerance of a high intertidal, pool-dwelling ascoglossan opisthobranch on New Zealand rocky shores. J. Exp. Mar. Biol. Ecol. 1994, 182, 65–84. [Google Scholar] [CrossRef]
- Russell, M.P. Echinoderm responses to variation in salinity. Adv. Mar. Biol. 2013, 66, 171–212. [Google Scholar] [PubMed]
- Delorme, N.J.; Sewell, M.A. Temperature and salinity: Two climate change stressors affecting early development of the New Zealand sea urchin Evechinus chloroticus. Mar. Biol. 2014, 161, 1999–2009. [Google Scholar] [CrossRef]
- Bindoff, N.L.; Cheung, J.G.; Kairo, J.; Arístegui, V.A.; Guinder, R.; Hallberg, N.; Hilmi, N.; Jiao, M.S.; Karim, L.; Levin, S.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities. In IPCC Special Report on the Ocean and Cryosphere in a Changing Climate; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2019; pp. 447–587. [Google Scholar]
- IPCC (2014) Climate Change. Impact, Adaptation and vulnerability. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, 1132. Available online: https://www.ipcc.ch/report/ar6/wg2/ (accessed on 16 November 2022).
- Allen, J.D.; Pechenik, J.A. Understanding the Effects of Low Salinity on Fertilization Success and Early Development in the Sand Dollar Echinarachnius parma. Biol. Bull. 2010, 218, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Carballeira, C.; Martín-Díaza, L.; DelVallsa, T.A. Influence of salinity on fertilization and larval development toxicity tests with two species of sea urchin. Mar. Environ. Res. 2011, 72, 196–203. [Google Scholar] [CrossRef]
- Sewell, M.A.; Young, C.M. Temperature limits to fertilization and early development in the tropical sea urchin Echinometra lucunter. J. Exp. Mar. Biol. Ecol. 1999, 236, 291–305. [Google Scholar] [CrossRef]
- Li, L.; Li, Q.; Sun, X.; Kong, L. Effects of temperature and salinity on larval growth, survival, and development of the sea cucumber Apostichopus japonicus. N. Am. J. Aquacult. 2011, 73, 296–303. [Google Scholar] [CrossRef]
- Pia, T.S.; Johnson, T.; George, S.B. Salinity-induced morphological changes in Pisaster ochraceus (Echinodermata: Asteroidea) larvae. J. Plankton. Res. 2012, 34, 590–601. [Google Scholar] [CrossRef] [Green Version]
- Mak, K.K.Y.; Chan, K.Y.K. Interactive effects of temperature and salinity on early life stages of the sea urchin Heliocidaris crassispina. Mar. Bio. 2018, 165, 57. [Google Scholar] [CrossRef]
- Bressan, M.; Marin, M.; Brunetti, R. Influence of temperature and salinity on embryonic development of Paracentrotus lividus (Lmk, 1816). Hydrobiologia 1995, 304, 175–184. [Google Scholar] [CrossRef]
- His, E.; Seaman, M.N.L.; Beiras, R. A simplification the bivalve embryogenesis and larval development bioassay method for water quality assessment. Wat Res. 1997, 31, 351–355. [Google Scholar] [CrossRef]
- Volpi Ghirardini, A.V.; Arizzi Novelli, A.; Likar, B.; Pojana, G.; Ghetti, P.F.; Marcomini, A. Sperm cell toxicity test using sea Urchin Paracentrotus lividus Lamarck (Echinodermata: Echinoidea): Sensitivity and discriminatory ability toward anionic and nonionic surfactants. Environ. Toxicol. Chem. 2001, 20, 644–651. [Google Scholar] [CrossRef]
- Marín, M.G.; Da Ros, L.; Moschino, V.; Campesan, G. Sediment elutriate toxicity testing with embryos of sea urchin (Paracentrotus lividus). Aquat. Ecosyst. Health Manag. 2001, 4, 215–221. [Google Scholar] [CrossRef]
- Fernández, N. Evaluación Biológica de la Contaminación Marina Costera Mediante Bioensayos con Embriones del Erizo de mar Paracentrotus lividus; Universidad de Vigo: Vigo, Spain, 2002. [Google Scholar]
- Pétinay, S.; Chataigner, C.; Basuyaux, O. Standardisation du développement larvaire de l’oursin, Paracentrotus lividus, pour l’évaluation de la qualité d’une eau de mer. Comptes Rendus Biol. 2009, 332, 1104–1114. [Google Scholar] [CrossRef]
- Saco-Álvarez, L.; Durán, I.; Ignacio Lorenzo, J.; Beiras, R. Methodologicalbasis for the optimization of amarine sea-urchinembryo test (SET) for the ecological assessment of coastal water quality. Ecotox. Environ. Safe 2010, 73, 491–499. [Google Scholar] [CrossRef]
- Máximo, M.V.; Mottola, L.S.M.; Resgalla, C., Jr. Sensibilidade do ouriço Arbacia lixula (Echinodermata: Echinoidea) em testes de toxicidade. J. Braz. Soc. Ecotoxicol. 2008, 3, 47–52. [Google Scholar] [CrossRef]
- His, E.; Heyvang, I.; Geffard, O.; de Montaudouin, X. A comparison between oyster (Crassostrea gigas) and sea urchin (Paracentrotus lividus) larval bioassays for toxicological studies. Water Res. 1999, 33, 1706–1718. [Google Scholar] [CrossRef]
- Carballeira, C.; De Orte, M.R.; Viana, I.G.; Delvalls, T.A.; Carballeira, A. Assessing the toxicity of chemical compounds associated with land-based marine fish farms: The sea urchin embryo bioassay with Paracentrotus lividus and Arbacia lixula. Arch. Environ. Contam. Toxicol. 2012, 63, 249–261. [Google Scholar] [CrossRef] [PubMed]
- Maisano, M.; Cappello, T.; Catanese, E.; Vitale, V.; Natalotto, A.; Giannetto, A.; Barreca, D.; Brunelli, E.; Mauceri, A.; Fasulo, S. Developmental abnormalities and neurotoxicological effects of CuO NPs on the black sea urchin Arbacia lixula by embryotoxicity assay. Mar. Environ. Res. 2015, 111, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Morroni, L.; Pinsino, A.; Pellegrini, D.; Regoli, F.; Matranga, V. Development of a new integrative toxicity index based on an improvement of the sea urchin embryo toxicity test. Ecotoxicol. Environ. Saf. 2016, 123, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Morroni, L.; Pinsino, A.; Pellegrini, D.; Regoli, F. Reversibility of metal induced malformations in sea urchin embryos. Ecotoxicol. Environ. Saf. 2018, 148, 923–929. [Google Scholar] [CrossRef]
- Gambardella, C.; Marcellini, F.; Falugi, C.; Varrella, S.; Corinaldesi, C. Early-stage anomalies in the sea urchin (Paracentrotus lividus) as bioindicators of multiple stressors in the marine environment: Overview and future perspectives. Environ. Pollut. 2021, 287, 117608. [Google Scholar] [CrossRef]
- Du Laing, G.; De Vos, R.; Vandecasteele, B.; Lesage, E.; Tack, F.M.; Verloo, M.G. Effect of salinity on heavy metal mobility and availability in intertidal sediments of the Scheldt estuary. Estuar. Coast. Shelf Sci. 2008, 77, 589–602. [Google Scholar] [CrossRef]
- Lewis, C.; Ellis, R.; Vernon, E.; Elliot, K.; Newbatt, S.; Wilson, R.W. Ocean acidification increases copper toxicity differentially in two key marine invertebrates with distinct acid-base responses. Sci. Rep. 2016, 6, 21554. [Google Scholar] [CrossRef] [Green Version]
- Grosell, M.; Blanchard, J.; Brix, K.V.; Gerdes, R. Physiology is pivotal for interactions between salinity and acute copper toxicity to fish and invertebrates. Aquatic. Toxicol. 2007, 84, 162–172. [Google Scholar] [CrossRef]
- Nielson, C.; Hird, C.; Lewis, C. Ocean acidification buffers the physiological responses of the king ragworm Alitta virens to the common pollutant copper. Aquat. Toxixol. 2019, 212, 120–127. [Google Scholar] [CrossRef]
- Morroni, L.; Sartori, D.; Costantini, M.; Genovesi, L.; Magliocco, T.; Ruocco, N.; Buttino, I. First molecular evidence of the toxicogenetic effects of copper on sea urchin Paracentrotus lividus embryo development. Water Res. 2019, 160, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Sartori, D.; Pellegrini, D.; Gaion, A. Analysis of variability in Embryological response of two sea urchin species to spatial and temporal features-can these factors influence responses in standardized Ecotoxicological assays? Expert Opin. Environ. Biol. 2016, S1, 2. [Google Scholar] [CrossRef] [Green Version]
- Fernández, N.; Beiras, R. Combined toxicity of dissolved mercury with copper, lead and cadmium on embryogenesis and early larval growth of the Paracentrotus lividus sea-urchin. Ecotoxicology 2001, 10, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Arizzi Novelli, A.; Losso, C.; Ghetti, P.F.; Ghirardini, A.V. Toxicity of heavy metals using sperm cell and embryo toxicity bioassays with Paracentrotus lividus (Echinodermata: Echinoidea): Comparisons with exposure concentrations in the lagoon of Venice, Italy. Environ. Toxicol. Chem. Int. J. 2003, 22, 1295–1301. [Google Scholar] [CrossRef]
- Manzo, S.; Buono, S.; Cresimini, C. Predictability of copper, Irgarol, and diuron combined effects on Sea Urchin Paracentrotus lividus. Arch. Environ. Contam. Toxicol. 2008, 54, 57–68. [Google Scholar] [CrossRef]
- Manzo, S.; Buono, S.; Cresimini, C. Cadmium, lead and their mixtures with copper: Paracentrotus lividus embryotoxicity assessment, prediction, and offspring quality evaluation. Ecotoxicology 2010, 19, 1209–1223. [Google Scholar] [CrossRef]
- Gharred, T.; Ezzine, I.K.; Naija, A.; Bouali, R.R.; Jebali, J. Assessment of toxic interactions between deltamethrin and copper on the fertility and developmental events in the Mediterranean sea urchin, Paracentrotus lividus. Environ. Monit. Assess. 2015, 187, 193. [Google Scholar] [CrossRef]
- Giannetto, A.; Cappello, T.; Oliva, S.; Parrino, V.; De Marco, G.; Fasulo, S.; Mauceri, A.; Maisano, M. Copper oxide nanoparticles induce the transcriptional modulation of oxidative stress-related genes in Arbacia lixula embryos. Aquatic. Toxicol. 2018, 201, 187–197. [Google Scholar] [CrossRef]
- Bonaventura, R.; Zito, F.; Morroni, L.; Pellegrini, D.; Regoli, F.; Pinsino, A. Development and validation of new analytical methods using sea urchin embryo bioassay to evaluate dredged marine sediments. J. Environ. Manag. 2021, 281, 111862. [Google Scholar] [CrossRef]
- Charles, C.; Veber, P.; Delignette-Muller, M.L. MOSAIC: A web-interface for statistical analyses in ecotoxicology. Environ. Sci. Pollut. Res. 2018, 25, 11295–11302. [Google Scholar] [CrossRef]
- Delignette-Muller, M.L.; Ruiz, P.; Charles, S.; Duchemin, W.; Lopes, C.; Kon-Kam-King, G.; Veber, P. Morse: Modelling Tools for Reproduction and Survival Data in Ecotoxicology. R Package Version 2.2.0. 2016. Available online: https://cran.r-project.org/web/packages/morse/index.html (accessed on 16 November 2022).
- Underwood, A.J. Experiments in Ecology. Their Logical Design and Interpretation Using Analysis of Variance; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Murado, M.A.; Prieto, M.A. NOEC and LOEC as merely concessive expedients: Two unambiguous alternatives and some criteria to maximize the efficiency of dose–response experimental designs. Sci. Total Environ. 2013, 461, 576–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammer, O.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Morroni, L.; Rakaj, A.; Grosso, L.; Fianchini, A.; Pellegrini, D.; Regoli, F. Sea cucumber Holothuria polii (Delle Chiaje, 1823) as new model for embryo bioassays in ecotoxicological studies. Chemosphere 2020, 240, 124819. [Google Scholar] [CrossRef] [PubMed]
- Rakaj, A.; Morroni, L.; Grosso, L.; Fianchini, A.; Pensa, D.; Pellegrini, D.; Regoli, F. Towards sea cucumbers as a new model in embryo-larval bioassays: Holothuria tubulosa as test species for the assessment of marine pollution. Sci. Total Environ. 2021, 787, 147593. [Google Scholar] [CrossRef]
- Carreras, C.; García-Cisneros, A.; Wangensteen, O.S.; Ordóñez, V.; Palacín, C.; Pascual, M.; Turon, X. East is East and West is West: Population genomics and hierarchical analyses reveal genetic structure and adaptation footprints in the keystone species Paracentrotus lividus (Echinoidea). Divers. Distrib. 2020, 26, 382–398. [Google Scholar] [CrossRef] [Green Version]
- Sarifudin, M.; Aminur Rahman, M.; Yu, S.H.; Yusoff, F.M.; Arshad, A. Effects of Salinity on Embryonic and Early Larval Development of a Tropical Sea Urchin, Salmacis sphaeroides. Iran. J. Fish. Sci. 2014, 13, 456–468. [Google Scholar]
- Sarifudin, M.; Aminur Rahman, M.; Yusoff, F.M.; Arshad, A.; Tans, S.G. Influence of salinity variations on the embryonic and early larval development of long-spined black sea urchin (Diadema setosum). J. Anim. Plant Sci. 2017, 27, 316–324. [Google Scholar]
- Shamim Parvez, M.D.; Aminur Rahman, M.; Yusoff, F.M.; Arshad, A.; Lee, S.-G. Salinity effects on the development of embryos and larvae of a high-valued sea urchin, Tripneustes gratilla (Linnaeus, 1758). J. Environ. Biol. 2018, 39, 785–794. [Google Scholar] [CrossRef]
- Greenwood, P.J.; Bennett, T. Some effects of temperature salinity combinations on the early development of the sea urchin Parechinus angulosus (leske). Fertilization. J. Exp. Mar. Biol. 1981, 51, 119–131. [Google Scholar] [CrossRef]
- Metaxas, A. The effect of salinity on larval survival and development in the sea urchin Echinometra lucunter. Invertebr. Reprod. Dev. 1998, 34, 323–330. [Google Scholar] [CrossRef]
- Dinnel, P.A.; Link, J.M.; Stober, Q.J.; Letourneau, M.W.; Roberts, W.E. Comparative sensitivity of sea urchin sperm bioassays to metals and pesticides. Arch. Environ. Contam. Toxicol. 1989, 18, 748–755. [Google Scholar] [CrossRef] [PubMed]
- Carr, S.R.; Chapman, D.C.; Long, E.R.; Windom, H.L.; Thursby, G.; Sloane, G.M.; Wolfe, D.A. Sediment quality assessment studies of Tampa Bay, Florida. Environ. Toxicol. Chem. 1996, 15, 1218–1231. [Google Scholar] [CrossRef]
- Warnau, M.; Temara, A.; Jangoux, M.; Dubois, P.; Iaccarino, M.; De Biase, A.; Pagano, G. Spermiotoxicity and embryotoxicity of heavy metals in the echinoid Paracentrotus lividus. Environ. Toxicol. Chem. 1996, 15, 1931–1936. [Google Scholar] [CrossRef]
- Philips, B.M.; Anderson, B.S.; Hunt, J.W. Spatial and temporal variation in results of purple urchin (Strongylocentrotus purpuratus) toxicity tests with zinc. Environ. Toxicol. Chem. 1998, 17, 453–459. [Google Scholar] [CrossRef]
- Radenac, G.; Fichet, D.; Miramand, P. Bioaccumulation and toxicity of four dissolved metals in Paracentrotus lividus sea-urchin embryo. Mar. Environ. Res. 2001, 51, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Gaion, A.; Scuderi, A.; Pellegrini, D.; Sartori, D. Arsenic exposure affects embryo development of sea urchin, Paracentrotus lividus (Lamarck, 1816). Bull. Environ. Contam. Toxicol. 2013, 91, 565–570. [Google Scholar] [CrossRef] [PubMed]
- Sartori, D.; Scuderi, A.; Sansone, G.; Gaion, A. Echinoculture: The rearing of Paracentrotus lividus in a recirculating aquaculture system. Experiments of artificial diets for the maintenance of sexual maturation. Aquacult. Int. 2015, 23, 111–125. [Google Scholar] [CrossRef]
- Florence, T.M. Trace element speciation and aquatic toxicology. Trends Anal. Chem. 1983, 2, 162–166. [Google Scholar] [CrossRef]
- Blust, R.; Fontaine, A.; Decleir, W. Effect of hydrogen ions and inorganic complexing on the uptake of copper by the brine shrimp Artemia franciscana. Mar. Ecol. Prog. Ser. 1991, 76, 273–282. [Google Scholar] [CrossRef]
- Vercauteren, K.; Blust, R. Bioavailability of dissolved zinc to the common mussel Mytilus edulis in complexing environments. Mar. Ecol. Prog. Ser. 1996, 137, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Lewis, A.G.; Whitfield, P.; Ramnarine, A. Some particulate and soluble agents affecting the relationship between metal toxicity and organism survival in the calanoid copepod Euchaeta japonica. Mar. Biol. 1972, 17, 215–221. [Google Scholar] [CrossRef]
- Lewis, A.G.; Whitfield, P.; Ramnarine, A. The reduction of copper toxicity in a marine copepod by sediment extract. Limnol. Oceanogr. 1973, 18, 324–326. [Google Scholar] [CrossRef]
- Davey, E.W.; Morgan, M.J.; Erickson, S.J. A biological measurement of the copper complexation capacity of sea water. Limnol. Oceanogr. 1973, 18, 993–997. [Google Scholar] [CrossRef]
- Morris, O.P.; Russel, G. Effect ofchelation on the toxicity of copper. Mar. Pollut. Bull. 1973, 4, 159–160. [Google Scholar] [CrossRef]
- Turner, D.R.; Whitfield, M.; Dickson, A.G. The equilibrium speciation of dissolved components in freshwater and seawater at 25 °C and 1 atm pressure. Geochim. Cosmochim. Acta 1981, 45, 855–881. [Google Scholar] [CrossRef]
- Newman, M.C.; Jagoe, C.H. Ligands and the bioavailability of metals in aquatic environments. In Bioavailability. Physical, Chemical and Biological Interactions; Hamelink, S.L., Landrum, P.F., Bergman, H.L., Benson, W.H., Eds.; CRC Press: New York, NY, USA, 1994; pp. 39–61. [Google Scholar]
- Coglianese, M.P. The effects of salinity on copper and silver toxicity to embryos of the Pacific oyster. Arch. Environ. Contam. Toxicol. 1983, 11, 297. [Google Scholar] [CrossRef]
- Hall, L.W.; Anderson, R.D. The Influence of Salinity on the Toxicity of Various Classes of Chemicals to Aquatic Biota. Crit. Rev. Toxicol. 1995, 2, 281–346. [Google Scholar] [CrossRef]
- Riba, I.; DelValls, T.A.; Forja, J.M.; Gómez-Parra, A. The influence of ph and salinity on the toxicity of heavy metals in sediment to the estuarine clam Ruditapes philippinarum. Environ. Toxicol. Chem. 2004, 23, 1100–1107. [Google Scholar] [CrossRef] [Green Version]
- Martins, C.D.M.G.; Barcarolli, I.F.; de Menezes, E.J.; Giacomin, M.M.; Wood, C.M.; Bianchini, A. Acute toxicity, accumulation and tissue distribution of copper in the blue crab Callinectes sapidus acclimated to different salinities: In vivo and in vitro studies. Aquat. Toxicol. 2011, 101, 88–99. [Google Scholar] [CrossRef]
- Park, J.; Kim, S.; Yoo, J.; Lee, J.S.; Park, J.W.; Jung, J. Effect of salinity on acute copper and zinc toxicity to Tigriopus japonicus: The difference between metal ions and nanoparticles. Mar. Pollut. Bull. 2014, 85, 526–531. [Google Scholar] [CrossRef]
- Piazza, V.; Gambardella, C.; Canepa, S.; Costa, E.; Faimali, M.; Garaventa, F. Temperature and salinity effects on cadmium toxicity on lethal and sublethal responses of Amphibalanus amphitrite nauplii. Ecotox. Environ. Saf. 2016, 123, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Moreira, A.; Figueira, E.; Libralato, G.; Soares, A.M.V.M.; Guida, M.; Freitas, R. Comparative sensitivity of Crassostrea angulata and Crassostrea gigas embryo-larval development to As under varying salinity and temperature. Mar. Environ. Res. 2018, 140, 135–144. [Google Scholar] [CrossRef] [PubMed]
- Thurberg, F.P.; Dawson, M.A.; Collier, R.S. Effects of copper and cadmium on osmoregulation and oxygen consumption in two species of estuarine crabs. Mar. Biol. 1973, 23, 171–175. [Google Scholar] [CrossRef]
- MacInnes, J.R.; Calabrese, A. Combined effects of salinity, temperature and copper on embryos and early larvae of the American oyster, Crassostrea virginica. Arch. Environ. Contam. Toxicol. 1979, 8, 553. [Google Scholar] [CrossRef]
- Webber, R.E.; Swaan, A.; Bang, A. Interactive effects of ambient copper and anoxia, temperature and salinity stress on survival and hemolymph and muscle tissue osmotic effectors in Mytilus edulis. J. Exp. Mar. Biol. Ecol. 1992, 159, 135. [Google Scholar] [CrossRef]
- Mcleese, D.W. Toxicity of copper at 2 temperatures and 3 salinities to American lobster (Homarus americanus). J. Fish. Res. Bd. Can. 1974, 31, 1949–1952. [Google Scholar] [CrossRef]
- Reardon, I.S.; Harrell, R.M. Acute toxicity of formalin and coppersulfate to striped bass fingerlings held in varying salinities. Aquaculture 1990, 87, 255–270. [Google Scholar] [CrossRef]
- Fernandez, T.V.; Jones, N.V. Studies on the toxicity of zinc and copper applied singly and jointly to Neries diversicolor at different salinities and temperatures. Trop. Ecol. 1990, 31, 47–55. [Google Scholar]
- Jones, L.H.; Jones, N.V.; Radlett, A.J. Some effects of salinity on toxicity of copper to polychaete Nereis diversicolor. Estuar. Coast. Mar. Sci. 1976, 4, 107–111. [Google Scholar] [CrossRef]
- Olson, K.R.; Harrel, R.C. Effect of salinity on acute toxicity of mercury, copper, and chromium for Rangia cuneata (Pelecypoda, Mactridae). Contrib. Mar. Sci. 1973, 17, 9–13. [Google Scholar]
- Birdsong, C.L.; Avault, J.W. Toxicity of certain chemicals to juvenile pompano. Prog. Fish Cult. 1971, 33, 76. [Google Scholar] [CrossRef]
- Anderson, B.S.; Hunt, J.W.; Piekarski, W.J.; Phillips, B.M.; Englund, M.A.; Tjeerdema, R.S.; Goetzl, J.D. Influence of salinity on copper and azide toxicity to larval topsmelt Atherinops affinis (Ayres). Arch. Environ. Contam. Physiol. 1995, 29, 366–372. [Google Scholar] [CrossRef] [PubMed]
- Deruytter, D.; Vandegehuchte, M.B.; Garrevoet, J.; De Laender, F.; Vergucht, E.; Delbeke, K.; Blust, R.; De Schamphelaere, K.A.; Vincze, L.; Janssen, C.R. Salinity and dissolved organic carbon both affect copper toxicity in mussel larvae: Copper speciation or competition cannot explain everything. Environ. Toxicol. Chem. 2015, 34, 1330–1336. [Google Scholar] [CrossRef] [PubMed]
- Forbes, V.E. Response of Hydrobia ventrosa (Montagu) to environmental stress: Effects of salinity fluctuations and cadmium exposure on growth. Funct. Ecol. 1991, 5, 642–648. [Google Scholar] [CrossRef]
- Knezovich, J.P. Chemical and biological factors affecting bioavailability of contaminants in seawater. In Bioavailability: Physical, Chemical and Biological Interactions; Hamelink, J.L., Landrum, P.F., Bergman, H.L., Benson, W.H., Eds.; Lewis Publishers: London, UK, 1994; pp. 23–30. [Google Scholar]
- Blanchard, J.; Grosell, M. Effects of salinity on copper accumulation in the common killifish (Fundulus heteroclitus). Environ. Toxicol. Chem. 2005, 24, 1403–1413. [Google Scholar] [CrossRef]
Figure 1.
Effects of salinity on larval development at different Cu concentrations (SW indicates sea water without copper addition). Data are expressed as mean percentage of abnormal embryos (±standard error) ((A) Paracentrotus lividus, (B) Arbacia lixula) and as ITI ((C) Paracentrotus lividus, (D) Arbacia lixula). Note: The asterisk (*) indicates significant differences (p < 0.05) between the standard salinity of 36‰ and other salinities.
Figure 1.
Effects of salinity on larval development at different Cu concentrations (SW indicates sea water without copper addition). Data are expressed as mean percentage of abnormal embryos (±standard error) ((A) Paracentrotus lividus, (B) Arbacia lixula) and as ITI ((C) Paracentrotus lividus, (D) Arbacia lixula). Note: The asterisk (*) indicates significant differences (p < 0.05) between the standard salinity of 36‰ and other salinities.


{kind=link}
Table 1.
Analysis of variance (two-way ANOVA) of embryo development in response to different salinities and copper concentrations. df: degrees of freedom, MS: mean squares, p: significance (p < 0.01).
Table 1.
Analysis of variance (two-way ANOVA) of embryo development in response to different salinities and copper concentrations. df: degrees of freedom, MS: mean squares, p: significance (p < 0.01).
| Paracentrotus lividus | Arbacia lixula | |||||||
|---|---|---|---|---|---|---|---|---|
| Source of Variation | df | MS | p | df | MS | p | ||
| Salinity | 6 | 34957 | 0.001 | 6 | 17131 | 0.001 | ||
| [Cu] | 4 | 21670 | 0.001 | 4 | 31848 | 0.001 | ||
| Salinity × Cu | 20 | 1734.3 | 0.001 | 20 | 3303.4 | 0.001 | ||
| Res | 333 | 5.7033 | 333 | 67.257 | ||||
| Total | 363 | 363 | ||||||
Table 2.
Median effective concentration (EC50) of sea urchin embryos exposed to different salinities. Values are reported with respective 95% confidential limits; n.c. = not calculated.
Table 2.
Median effective concentration (EC50) of sea urchin embryos exposed to different salinities. Values are reported with respective 95% confidential limits; n.c. = not calculated.
| Salinity (‰) | EC50 (µg/L) | |
|---|---|---|
| Paracentrotus lividus | Arbacia lixula | |
| 27 | n.c. | n.c. |
| 30 | 10.7 (8.3–12.9) | 9.82 (9.31–10.3) |
| 33 | 17.6 (16.3–18.9) | 15.8 (15.6–16.1) |
| 36 | 28.8 (28.4–29.3) | 15.5 (15.1–15.8) |
| 39 | 24.7 (23.6–25.7) | 13.6 (13.3–13.9) |
| 42 | 21.7 (19.3–23.8) | 9.0 (8.6–9.3) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Morroni, L.; Gaion, A.; Broccoli, A.; Ferrari, S.; Pellegrini, D.; Sartori, D. Influence of Salinity on Copper Toxicity in Paracentrotus lividus and Arbacia lixula Embryos. Water 2023, 15, 65. https://doi.org/10.3390/w15010065
AMA Style
Morroni L, Gaion A, Broccoli A, Ferrari S, Pellegrini D, Sartori D. Influence of Salinity on Copper Toxicity in Paracentrotus lividus and Arbacia lixula Embryos. Water. 2023; 15(1):65. https://doi.org/10.3390/w15010065
Chicago/Turabian StyleMorroni, Lorenzo, Andrea Gaion, Andrea Broccoli, Stefano Ferrari, David Pellegrini, and Davide Sartori. 2023. "Influence of Salinity on Copper Toxicity in Paracentrotus lividus and Arbacia lixula Embryos" Water 15, no. 1: 65. https://doi.org/10.3390/w15010065
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.