
Interactive Effects of Nutrients and Salinity on Phytoplankton in Subtropical Plateau Lakes of Contrasting Water Depths
 and
and 
Abstract
:1. Introduction
2. Materials and Methods
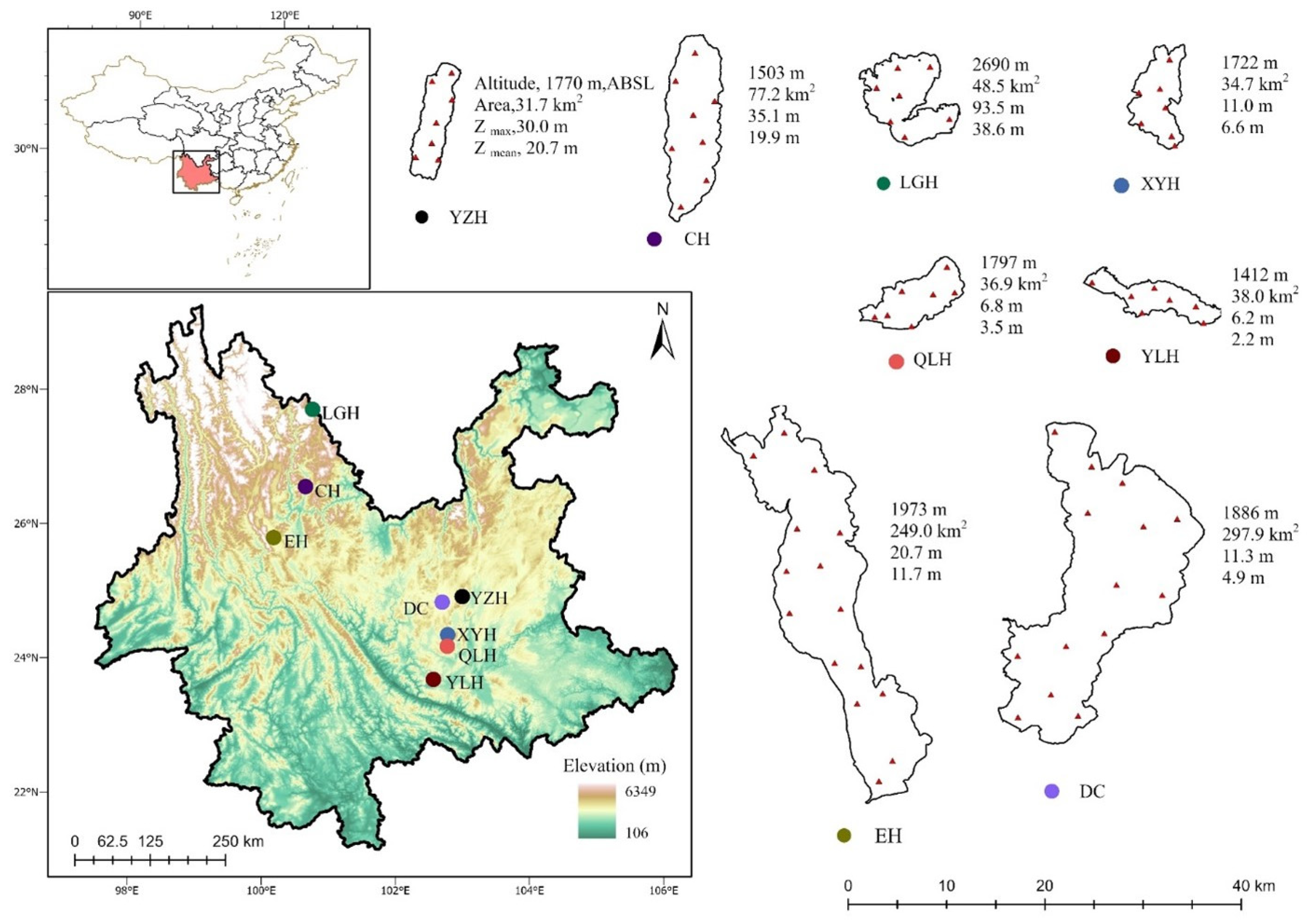
2.1. Field Sampling and Measurements
2.2. Data Processing and Analyses
3. Results
3.1. Environmental Conditions and Assemblage Structure of Phytoplankton
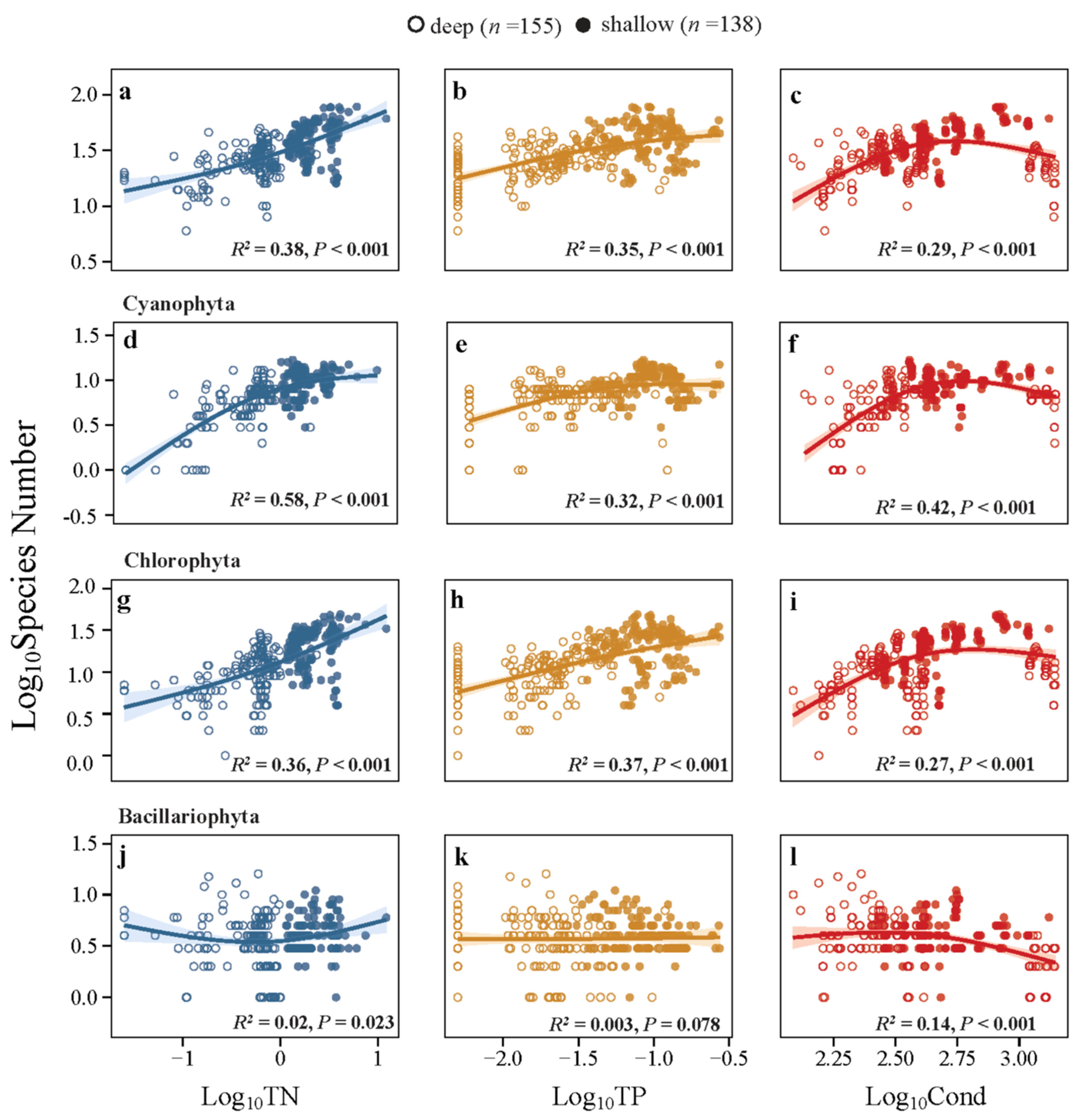
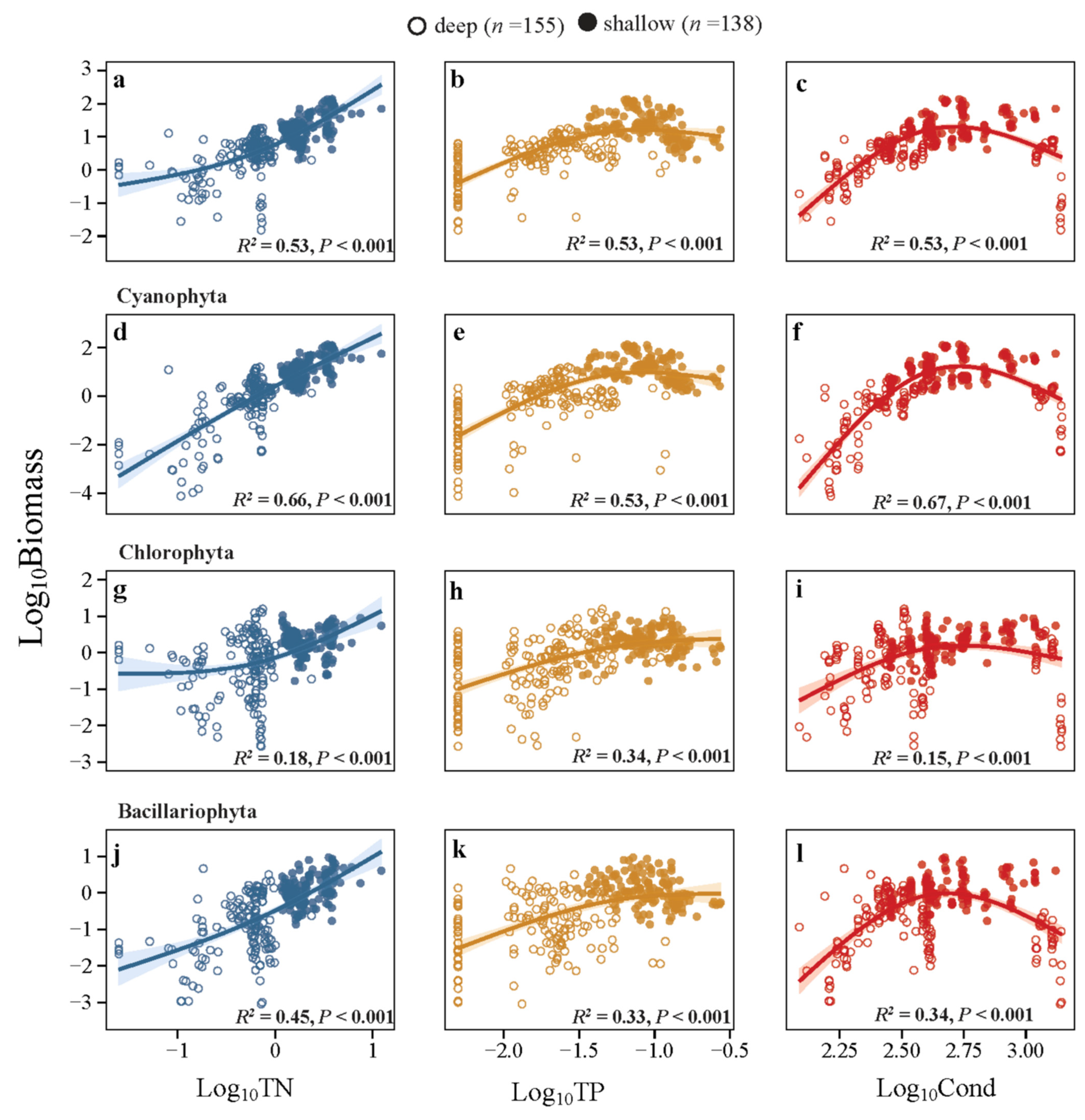
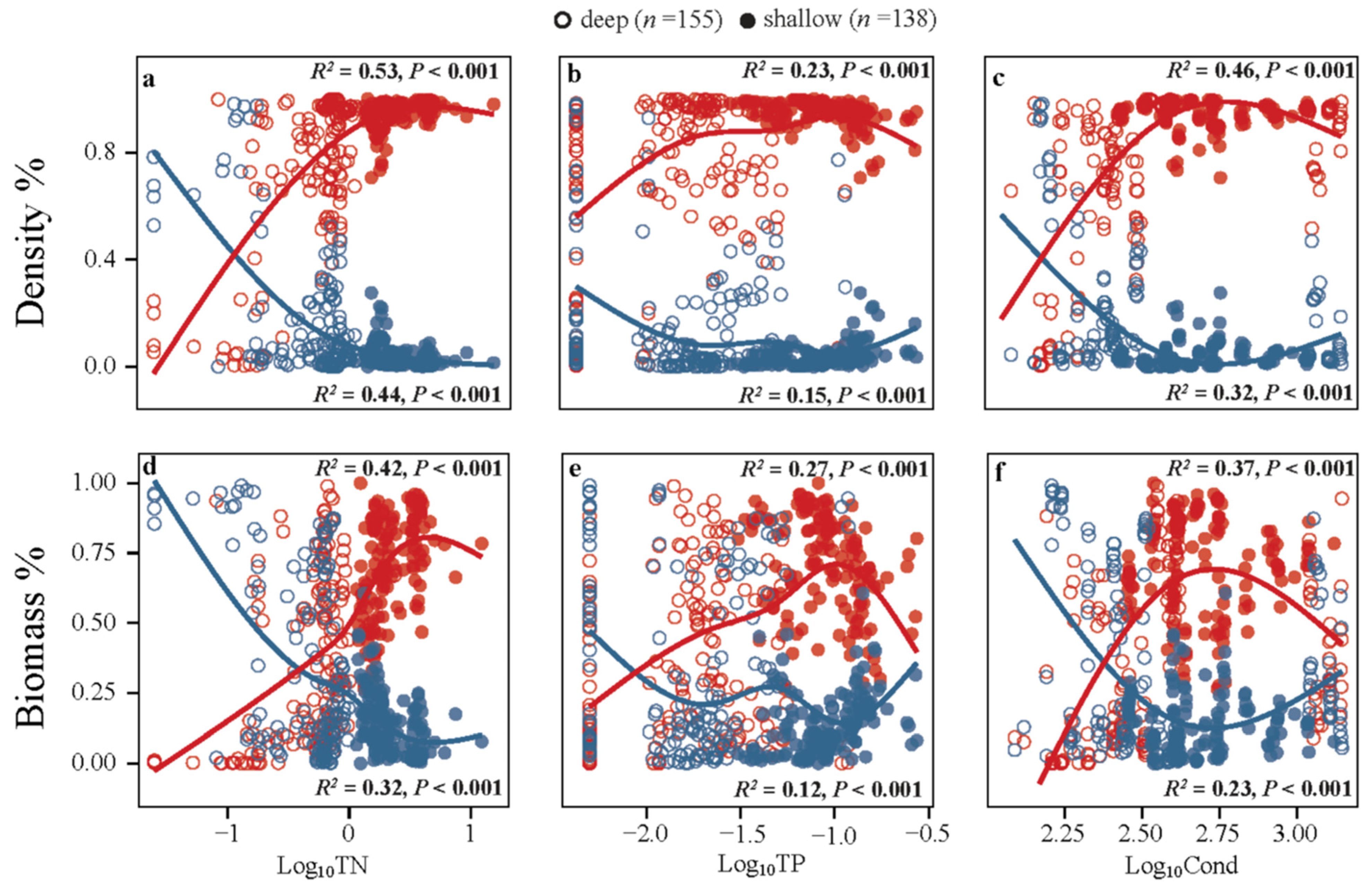
3.2. Relationships of Phytoplankton with the Environments
4. Discussion
4.1. Changes in Species Number and Abundance of Phytoplankton
4.2. Changes in the Dominance of the First Two Main Phytoplankton Taxa
5. Conclusions
- Species number and abundance of phytoplankton increased with increasing TN and TP concentrations; however, the increasing trends declined at high TP concentrations in shallow lakes. Shortage of nitrogen might be the limiting factor as suggested by a low TN/TP ratio in the high-TP lakes.
- Phytoplankton species richness and abundance did not decline in shallow lakes but did so in deep lakes at similar Cond levels. This could perhaps be explained by the buffering effects of eutrophication since the deep lakes were in oligotrophic or mesotrophic states, while the shallow lakes were eutrophic.
- The two dominant phytoplankton taxa by numbers, cyanophyta (Cyan) and chlorophyta (Chlo), showed clearly different trends along the nutrient and Cond gradients, with Chlo being dominant at the lower end and Cyan being dominant at the higher ends of the environmental gradients. In summer, similar shifts in taxa dominance occurred based on biomass data but not for density, where Cyan dominated along the entire nutrient and Cond gradients.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, V.H.; Schindler, D.W. Eutrophication Science: Where Do We Go from Here? Trends Ecol. Evol. 2009, 4, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Le Moal, M.; Gascuel-Odoux, C.; Ménesguen, A.; Souchon, Y.; Étrillard, C.; Levain, A.; Moatar, F.; Pannard, A.; Souchu, P.; Lefebvre, A.; et al. Eutrophication: A New Wine in an Old Bottle? Sci. Total Environ. 2019, 651, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeppesen, E.; Pierson, D.; Jennings, E. Effect of Extreme Climate Events on Lake Ecosystems. Water 2021, 13, 282. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Climate Change: A Catalyst for Global Expansion of Harmful Cyanobacterial Blooms. Environ. Microbiol. Rep. 2009, 1, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Rosén, G. Phytoplankton Indicators and Their Relations to Certain Chemical and Physical Factors. Limnologica 1981, 13, 263–290. [Google Scholar]
- Saturday, A.; Lyimo, T.J.; Machiwa, J.; Pamba, S. Spatial and Temporal Variations of Phytoplankton Composition and Biomass in Lake Bunyonyi, South-Western Uganda. Environ. Monit. Assess. 2022, 194, 288. [Google Scholar] [CrossRef]
- Ochieng, B.; Mbao, E.O.; Zhang, Z.; Shi, L.; Liu, Q. Phytoplankton Community Structure of Tang-Pu Reservoir: Status and Ecological Assessment in Relation to Physicochemical Variability. Environ. Monit. Assess. 2022, 194, 382. [Google Scholar] [CrossRef]
- Álvarez-Góngora, C.; Herrera-Silveira, J.A. Variations of Phytoplankton Community Structure Related to Water Quality Trends in a Tropical Karstic Coastal Zone. Mar. Pollut. Bull. 2006, 52, 48–60. [Google Scholar] [CrossRef]
- Lepistö, L.; Holopainen, A.-L.; Vuoristo, H. Type-Specific and Indicator Taxa of Phytoplankton as a Quality Criterion for Assessing the Ecological Status of Finnish Boreal Lakes. Limnologica 2004, 34, 236–248. [Google Scholar] [CrossRef] [Green Version]
- Feng, C.; Jia, J.Y.; Wang, C.; Han, M.Q.; Dong, C.C.; Huo, B.; Li, D.P.; Liu, X.J. Phytoplankton and Bacterial Community Structure in Two Chinese Lakes of Different Trophic Status. Microorganisms 2019, 7, 621. [Google Scholar] [CrossRef] [Green Version]
- Schulhof, M.A.; Van de Waal, D.B.; Declerck, S.A.J.; Shurin, J.B. Phytoplankton Functional Composition Determines Limitation by Nutrients and Grazers across a Lake Productivity Gradient. Ecosphere 2022, 13, e4008. [Google Scholar] [CrossRef]
- Domingues, R.B.; Barbosa, A.; Galvão, H. Nutrients, Light and Phytoplankton Succession in a Temperate Estuary (the Guadiana, South-Western Iberia). Estuar. Coast. Shelf Sci. 2005, 64, 249–260. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling Harmful Cyanobacterial Blooms in a World Experiencing Anthropogenic and Climatic-Induced Change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.D.; Xu, X.W.; Qi, M.; Sun, J.J.; Zhang, Y.Y.; Zhang, W.; Wang, M.Z.; Wang, X.J.; Zhang, Y. Lake Warming Intensifies the Seasonal Pattern of Internal Nutrient Cycling in the Eutrophic Lake and Potential Impacts on Algal Blooms. Water Res. 2021, 188, 116570. [Google Scholar] [CrossRef]
- Hu, Y.; Jiang, X.Y.; Shao, K.Q.; Tang, X.G.; Qin, B.Q.; Gao, G. Convergency and Stability Responses of Bacterial Communities to Salinization in Arid and Semiarid Areas: Implications for Global Climate Change in Lake Ecosystems. Front. Microbiol. 2022, 12, 741645. [Google Scholar] [CrossRef]
- Han, L.; Li, Y.M.; Zou, Y.F.; Gao, X.B.; Gu, Y.G.; Wang, L. Relationship between Lake Salinity and the Climatic Gradient in Northeastern China and Its Implications for Studying Climate Change. Sci. Total Environ. 2022, 805, 150403. [Google Scholar] [CrossRef]
- Hintz, W.D.; Relyea, R.A. A Review of the Species, Community, and Ecosystem Impacts of Road Salt Salinisation in Fresh Waters. Freshw. Biol. 2019, 64, 1081–1097. [Google Scholar] [CrossRef] [Green Version]
- Hintz, W.D.; Arnott, S.E.; Symons, C.C.; Greco, D.A.; McClymont, A.; Brentrup, J.A.; Cañedo-Argüelles, M.; Derry, A.M.; Downing, A.L.; Gray, D.K.; et al. Current Water Quality Guidelines across North America and Europe Do Not Protect Lakes from Salinization. Proc. Natl. Acad. Sci. USA 2022, 119, e2115033119. [Google Scholar] [CrossRef]
- Lin, Q.Q.; Xu, L.; Hou, J.Z.; Liu, Z.W.; Jeppesen, E.; Han, B.-P. Responses of Trophic Structure and Zooplankton Community to Salinity and Temperature in Tibetan Lakes: Implication for the Effect of Climate Warming. Water Res. 2017, 124, 618–629. [Google Scholar] [CrossRef]
- Jeppesen, E.; Sondergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B.; et al. Lake Responses to Reduced Nutrient Loading—An Analysis of Contemporary Long-Term Data from 35 Case Studies. Freshw. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Osburn, F.S.; Wagner, N.D.; Taylor, R.B.; Chambliss, C.K.; Brooks, B.W.; Scott, J.T. The Effects of Salinity and N: P on N-rich Toxins by Both an N-fixing and Non-N-fixing Cyanobacteria. Limnol. Oceanogr. Lett. 2022, lol2.10234. [Google Scholar] [CrossRef]
- Chakraborty, P.; Acharyya, T.; Raghunadh Babu, P.V.; Bandyopadhyay, D. Impact of Salinity and PH on Phytoplankton Communities in a Tropical Freshwater System: An Investigation with Pigment Analysis by HPLC. J. Environ. Monit. 2011, 13, 614. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Geng, M.D.; Yu, J.L.; Du, Y.X.; Xu, M.; Zhang, W.Z.; Wang, J.J.; Su, H.J.; Wang, R.; Chen, F.Z. Eutrophication Decrease Compositional Dissimilarity in Freshwater Plankton Communities. Sci. Total Environ. 2022, 821, 153434. [Google Scholar] [CrossRef] [PubMed]
- Lind, L.; Schuler, M.S.; Hintz, W.D.; Stoler, A.B.; Jones, D.K.; Mattes, B.M.; Relyea, R.A. Salty Fertile Lakes: How Salinization and Eutrophication Alter the Structure of Freshwater Communities. Ecosphere 2018, 9, e02383. [Google Scholar] [CrossRef]
- Qin, B.Q.; Zhou, J.; Elser, J.J.; Gardner, W.S.; Deng, J.M.; Brookes, J.D. Water Depth Underpins the Relative Roles and Fates of Nitrogen and Phosphorus in Lakes. Environ. Sci. Technol. 2020, 54, 3191–3198. [Google Scholar] [CrossRef]
- Xie, P. Biological Mechanisms Driving the Seasonal Changes in the Internal Loading of Phosphorus in Shallow Lakes. Sci. China Ser. D 2006, 49, 14–27. [Google Scholar] [CrossRef] [Green Version]
- Szklarek, S.; Górecka, A.; Wojtal-Frankiewicz, A. The Effects of Road Salt on Freshwater Ecosystems and Solutions for Mitigating Chloride Pollution—A Review. Sci. Total Environ. 2022, 805, 150289. [Google Scholar] [CrossRef]
- Zadereev, E.; Drobotov, A.; Anishchenko, O.; Kolmakova, A.; Lopatina, T.; Oskina, N.; Tolomeev, A. The Structuring Effects of Salinity and Nutrient Status on Zooplankton Communities and Trophic Structure in Siberian Lakes. Water 2022, 14, 1468. [Google Scholar] [CrossRef]
- Christensen, I.; Pedersen, L.K.; Søndergaard, M.; Lauridsen, T.L.; Tserenpil, S.; Richardson, K.; Amorim, C.A.; Pacheco, J.P.; Jeppesen, E. Impact of Zooplankton Grazing on Phytoplankton in North Temperate Coastal Lakes: Changes along Gradients in Salinity and Nutrients. Hydrobiologia 2022, 1–18. [Google Scholar] [CrossRef]
- Janssen, A.B.G. Characterizing 19 Thousand Chinese Lakes, Ponds and Reservoirs by Morphometric, Climate and Sediment Characteristics. Water Res. 2021, 202, 117427. [Google Scholar] [CrossRef]
- Liu, W.Z.; Li, S.Y.; Bu, H.M.; Zhang, Q.F.; Liu, G.H. Eutrophication in the Yunnan Plateau Lakes: The Influence of Lake Morphology, Watershed Land Use, and Socioeconomic Factors. Environ. Sci. Pollut. Res. 2012, 19, 858–870. [Google Scholar] [CrossRef]
- Yan, D.N.; Xu, H.; Lan, J.H.; Yang, M.; Wang, F.S.; Hou, W.G.; Zhou, K.G.; An, Z.S. Warming Favors Subtropical Lake Cyanobacterial Biomass Increasing. Sci. Total Environ. 2020, 726, 138606. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.K.; Liu, X.B.; Peng, W.Q.; Dong, F.; Chen, Q.C.; Sun, Y.L.; Wang, R.N. Hydroclimatic Influence on the Salinity and Water Volume of a Plateau Lake in Southwest China. Sci. Total Environ. 2019, 659, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Yu, Y.H. Comprehensive Drought Monitoring in Yunnan Province, China Using Multisource Remote Sensing Data. J. Mt. Sci. 2021, 18, 1537–1549. [Google Scholar] [CrossRef]
- Water and Wastewater Monitoring and Analysis Methods, 4th ed.; Editorial Board of Water and Wastewater Monitoring and Analysis, Methods of the Ministry of Environmental Protection of the People’s Republic of China (Ed.) China Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Zhang, Z.; Huang, X. Freshwater Plankton Research Methods; Science Press: Beijing, China, 1991. [Google Scholar]
- Hu, H.; Wei, Y. Freshwater Algae in China—System, Classification and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Weng, J.; Xu, H. Atlas of Common Freshwater Planktonic Algae in China; Shanghai Science and Technology Press: Shanghai, China, 2010. [Google Scholar]
- Zhang, Z.; Huang, X. Chinese Freshwater Algae Journal; Science Press: Beijing, China, 2007. [Google Scholar]
- Jin, X.; Tu, Q. Specification for Eutrophication Survey of Lakes; China Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Jing, H.W.; Hua, L.; Guo, J. Analysis on urban lakes’ eutrophication status in Beijing. J. Lake Sci. 2008, 20, 357–363. [Google Scholar] [CrossRef] [Green Version]
- Moffett, E.R.; Baker, H.K.; Bonadonna, C.C.; Shurin, J.B.; Symons, C.C. Cascading Effects of Freshwater Salinization on Plankton Communities in the Sierra Nevada. Limnol. Oceanogr. Lett. 2020, lol2.10177. [Google Scholar] [CrossRef]
- Boros, E.; Vörös, L. A Magyarországi Szikes Tavak Sótartalma És Ionösszetétele. Acta Biol. Debrecina Suppl. Oecologica Hung. 2010, 22, 37–51. [Google Scholar]
- Jeppesen, E.; Jensen, J.P.; Søndergaard, M.; Lauridsen, T.; Landkildehus, F. Trophic Structure, Species Richness and Biodiversity in Danish Lakes: Changes along a Phosphorus Gradient: A Detailed Study of Danish Lakes along a Phosphorus Gradient. Freshw. Biol. 2000, 45, 201–218. [Google Scholar] [CrossRef]
- Reinl, K.L.; Brookes, J.D.; Carey, C.C.; Harris, T.D.; Ibelings, B.W.; Morales-Williams, A.M.; De Senerpont Domis, L.N.; Atkins, K.S.; Isles, P.D.F.; Mesman, J.P.; et al. Cyanobacterial Blooms in Oligotrophic Lakes: Shifting the High-nutrient Paradigm. Freshw. Biol. 2021, 66, 1846–1859. [Google Scholar] [CrossRef]
- Abell, J.M.; Özkundakci, D.; Hamilton, D.P.; Reeves, P. Restoring Shallow Lakes Impaired by Eutrophication: Approaches, Outcomes, and Challenges. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1199–1246. [Google Scholar] [CrossRef]
- Quirós, R. The Nitrogen to Phosphorus Ratio for Lakes: A Cause or a Consequence of Aquatic Biology? In El Agua en Iberoamerica: De la Limnología a la Gestion en Sudamerica, 2nd ed.; Fernandez Cirelli, A., Chalar Marquisa, G., Eds.; CYTED XVII; Centro de Estudios Transdiciplinarios del Agua, Facultad de Veterinaria, Universidad de Buenos Aires: Buenos Aires, Argentina, 2002; pp. 11–26. [Google Scholar]
- Klausmeier, C.A.; Litchman, E.; Daufresne, T.; Levin, S.A. Optimal Nitrogen-to-Phosphorus Stoichiometry of Phytoplankton. Nature 2004, 429, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Filstrup, C.; Heathcote, A.; Kendall, D.; Downing, J. Phytoplankton Taxonomic Compositional Shifts across Nutrient and Light Gradients in Temperate Lakes. Inland Waters 2016, 6, 234–249. [Google Scholar] [CrossRef]
- Bergström, A.-K. The Use of TN:TP and DIN:TP Ratios as Indicators for Phytoplankton Nutrient Limitation in Oligotrophic Lakes Affected by N Deposition. Aquat. Sci. 2010, 72, 277–281. [Google Scholar] [CrossRef]
- Zhou, J.; Leavitt, P.R.; Zhang, Y.; Qin, B. Anthropogenic Eutrophication of Shallow Lakes: Is It Occasional? Water Res. 2022, 221, 118728. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.J.; Wang, H.Z.; Liang, X.M.; Wu, S.K. Total Phosphorus Thresholds for Regime Shifts Are Nearly Equal in Subtropical and Temperate Shallow Lakes with Moderate Depths and Areas. Freshw. Biol. 2014, 59, 1659–1671. [Google Scholar] [CrossRef]
- Dolman, A.M.; Mischke, U.; Wiedner, C. Lake-Type-Specific Seasonal Patterns of Nutrient Limitation in German Lakes, with Target Nitrogen and Phosphorus Concentrations for Good Ecological Status. Freshw. Biol. 2016, 61, 444–456. [Google Scholar] [CrossRef]
- Mo, Y.; Peng, F.; Gao, X.; Xiao, P.; Logares, R.; Jeppesen, E.; Ren, K.; Xue, Y.; Yang, J. Low Shifts in Salinity Determined Assembly Processes and Network Stability of Microeukaryotic Plankton Communities in a Subtropical Urban Reservoir. Microbiome 2021, 9, 128. [Google Scholar] [CrossRef]
- Hintz, W.D.; Mattes, B.M.; Schuler, M.S.; Jones, D.K.; Stoler, A.B.; Lind, L.; Relyea, R.A. Salinization Triggers a Trophic Cascade in Experimental Freshwater Communities with Varying Food-Chain Length. Ecol. Appl. 2017, 27, 833–844. [Google Scholar] [CrossRef]
- Coldsnow, K.D.; Relyea, R.A. The Combined Effects of Macrophytes and Three Road Salts on Aquatic Communities in Outdoor Mesocosms. Environ. Pollut. 2021, 287, 117652. [Google Scholar] [CrossRef]
- Greco, D.A.; Arnott, S.E.; Fournier, I.B.; Schamp, B.S. Effects of chloride and nutrients on freshwater plankton communities. Limnol. Oceanogr. Lett. 2021, lol2.10202. [Google Scholar] [CrossRef]
- Ersoy, Z.; Abril, M.; Cañedo-Argüelles, M.; Espinosa, C.; Vendrell-Puigmitja, L.; Proia, L. Experimental Assessment of Salinization Effects on Freshwater Zooplankton Communities and Their Trophic Interactions under Eutrophic Conditions. Environ. Pollut. 2022, 313, 120127. [Google Scholar] [CrossRef]
- Li, Z.; Gao, Y.; Wang, S.; Lu, Y.; Sun, K.; Jia, J.; Wang, Y. Phytoplankton Community Response to Nutrients along Lake Salinity and Altitude Gradients on the Qinghai-Tibet Plateau. Ecol. Indic. 2021, 128, 107848. [Google Scholar] [CrossRef]
- McClymont, A.; Arnott, S.E.; Rusak, J.A. Interactive Effects of Increasing Chloride Concentration and Warming on Freshwater Plankton Communities. Limnol. Oceanogr. Lett. 2022, lol2.10278. [Google Scholar] [CrossRef]
- Ballot, A.; Kotut, K.; Novelo, E.; Krienitz, L. Changes of Phytoplankton Communities in Lakes Naivasha and Oloidien, Examples of Degradation and Salinization of Lakes in the Kenyan Rift Valley. Hydrobiologia 2009, 632, 359–363. [Google Scholar] [CrossRef]
- Zhang, H.H.; Zong, R.R.; He, H.Y.; Liu, K.W.; Yan, M.M.; Miao, Y.T.; Ma, B.; Huang, X. Biogeographic Distribution Patterns of Algal Community in Different Urban Lakes in China: Insights into the Dynamics and Co-Existence. J. Environ. Sci. 2021, 100, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Rakocevic, J. Spatial and Temporal Distribution of Phytoplankton in Lake Skadar. Arch. Biol. Sci. 2012, 64, 585–595. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LGH | CH | YZH | EH | XYH | DC | YLH | QLH | |
|---|---|---|---|---|---|---|---|---|
| Longitude (E°) | 100.79 | 100.66 | 103.01 | 100.02 | 102.78 | 102.71 | 102.59 | 102.79 |
| Latitude (N°) | 27.72 | 26.54 | 24.91 | 25.78 | 24.34 | 24.83 | 23.67 | 24.17 |
| ZSD (m) | 8.21 ± 0.70 | 2.29 ± 0.37 | 1.91 ± 0.09 | 1.26 ± 0.05 | 0.46 ± 0.03 | 0.38 ± 0.03 | 0.19 ± 0.02 | 0.33 ± 0.02 |
| Turb (NTU) | 2.41 ± 1.03 | 4.29 ± 0.46 | 2.74 ± 0.39 | 5.82 ± 0.38 | 24.11 ± 1.57 | 37.77 ± 2.20 | 32.39 ± 2.16 | 24.70 ± 1.55 |
| WT (°C) | 15.91 ± 0.70 | 21.14 ± 0.69 | 19.68 ± 0.58 | 19.00 ± 0.60 | 19.77 ± 0.88 | 18.95 ± 0.60 | 21.71 ± 0.98 | 20.16 ± 0.85 |
| DO (mg/L) | 6.69 ± 0.33 | 7.31 ± 0.18 | 7.63 ± 0.20 | 7.81 ± 0.16 | 8.19 ± 0.31 | 9.99 ± 1.43 | 7.35 ± 0.33 | 9.10 ± 0.32 |
| Cond (μS/cm) | 181.8 ± 4.2 | 1236 ± 17.9 | 389.1 ± 3.8 | 277.6 ± 4.4 | 519.0 ± 11.6 | 360.9 ± 6.6 | 500.4 ± 11.2 | 880.7 ± 30.7 |
| pH | 7.83 ± 0.10 | 8.72 ± 0.05 | 8.22 ± 0.08 | 8.17 ± 0.05 | 8.31 ± 0.08 | 8.56 ± 0.06 | 8.10 ± 0.08 | 8.38 ± 0.06 |
| TN (mg/L) | 0.19 ± 0.03 | 0.77 ± 0.02 | 0.59 ± 0.02 | 0.64 ± 0.04 | 1.69 ± 0.03 | 1.69 ± 0.05 | 3.24 ± 0.12 | 3.99 ± 0.37 |
| TP (mg/L) | 0.01 ± 0.004 | 0.03 ± 0.003 | 0.02 ± 0.001 | 0.02 ± 0.003 | 0.16 ± 0.013 | 0.10 ± 0.005 | 0.06 ± 0.004 | 0.11 ± 0.004 |
| N/P | 25.8 ± 2.4 | 48.7 ± 7.6 | 31.6 ± 1.8 | 40.2 ± 4.7 | 13.2 ± 1.7 | 19.0 ± 0.7 | 59.3 ± 3.7 | 37.4 ± 2.9 |
| Chl a (μg/L) | 1.89 ± 0.50 | 5.41 ± 0.50 | 13.71 ± 0.57 | 10.20 ± 0.72 | 45.45 ± 3.47 | 52.21 ± 4.24 | 72.60 ± 6.64 | 76.24 ± 9.12 |
| TLI | 15.98 ± 1.61 | 33.74 ± 1.31 | 35.12 ± 0.38 | 35.71 ± 0.61 | 51.36 ± 0.46 | 52.42 ± 0.69 | 57.13 ± 0.93 | 57.33 ± 0.57 |
| Trophic status | oligotrophic | mesotrophic | mesotrophic | mesotrophic | eutrophic | eutrophic | eutrophic | eutrophic |
| Multiple Stepwise Regression Analysis (n = 293) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Non-standardized coefficient | Standardization coefficient | t | p | VIF | R2 | adjusted R2 | F | D-W | ||
| B | SE | Beta | ||||||||
| ZM | −0.196 | 0.023 | −0.451 | 8.544 | <0.001 | 1.54 | 0.482 | 0.477 | 88.838 (p < 0.001) | 0.689 |
| TP | 0.117 | 0.023 | 0.283 | 5.138 | <0.001 | 1.677 | ||||
| Cond | 0.083 | 0.035 | 0.106 | 2.358 | 0.019 | 1.124 | ||||
| dependent variable: Species Number | ||||||||||
| TN | 0.591 | 0.112 | 0.363 | 5.263 | <0.001 | 3.143 | 0.566 | 0.562 | 124.511 (p < 0.001) | 0.674 |
| TP | 0.42 | 0.09 | 0.273 | 4.65 | <0.001 | 2.27 | ||||
| ZM | −0.326 | 0.093 | −0.202 | 3.528 | <0.001 | 2.155 | ||||
| dependent variable: Biomass | ||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Jiang, X.; Li, Y.-L.; Yang, L.-J.; Li, Y.-H.; Liu, Y.; Zhou, L.; Wang, P.-Z.; Zhao, X.; Wang, H.-J.; et al. Interactive Effects of Nutrients and Salinity on Phytoplankton in Subtropical Plateau Lakes of Contrasting Water Depths. Water 2023, 15, 69. https://doi.org/10.3390/w15010069
Wang Y, Jiang X, Li Y-L, Yang L-J, Li Y-H, Liu Y, Zhou L, Wang P-Z, Zhao X, Wang H-J, et al. Interactive Effects of Nutrients and Salinity on Phytoplankton in Subtropical Plateau Lakes of Contrasting Water Depths. Water. 2023; 15(1):69. https://doi.org/10.3390/w15010069
Chicago/Turabian StyleWang, Ying, Xia Jiang, Yan-Ling Li, Li-Juan Yang, Ye-Hao Li, Ying Liu, Long Zhou, Pu-Ze Wang, Xu Zhao, Hai-Jun Wang, and et al. 2023. "Interactive Effects of Nutrients and Salinity on Phytoplankton in Subtropical Plateau Lakes of Contrasting Water Depths" Water 15, no. 1: 69. https://doi.org/10.3390/w15010069