
First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea


1
Institute for Marine and Coastal Research, University of Dubrovnik, Kneza Damjana Jude 12, 20000 Dubrovnik, Croatia
2
Institute of Oceanography and Fisheries, Šetalište Ivana Meštrovića 63, 21000 Split, Croatia
3
Department of Applied Ecology, University of Dubrovnik, Ćira Carića 4, 20000 Dubrovnik, Croatia
*
Author to whom correspondence should be addressed.
Water 2023, 15(10), 1821; https://doi.org/10.3390/w15101821
Submission received: 16 April 2023
/
Revised: 3 May 2023
/
Accepted: 8 May 2023
/
Published: 10 May 2023
(This article belongs to the Special Issue The Study of Plankton in the Mediterranean Sea)
Abstract
:The tintinnid ciliate Rhizodomus tagatzi has spread rapidly in recent decades in neritic areas of the Mediterranean and adjacent seas, where it is classified as a non-indigenous species. The present study documents the first finding of the species in the Adriatic Sea, in the transitional waters of the Neretva River estuary. Zooplankton material was collected with 5 L Niskin bottles in three layers (1, 5 and 7 m) at two stations, at the mouth of the Neretva River and 16 km upstream, in the period from May 2022 to January 2023. In addition to the morphological characteristics of R. tagatzi, we discuss the state and variability of the populations based on the abundance of the species and the influence of abiotic environmental factors (temperature and salinity) and possible pathways of entry into the Adriatic Sea. The maximum abundance of R. tagatzi in the Neretva River estuary of ~2 × 103 cells L−1 is among the highest in the Mediterranean Sea, and the species has a significant contribution to the tintinnid community (>80%), indicating successful establishment of the population in the estuary and intense influence on ecosystem functioning until the strong river flow completely displaces the wedge from the seabed. Statistical methods confirm the differences between stations in the total abundance of tintinnids and differences in their vertical distribution at the upstream station.
1. Introduction
Tintinnid ciliates are a common, occasionally abundant and important component of planktonic food webs. They represent a monophyletic branch of the subclass Choreotrichia consisting exclusively of loricate forms, but also suggest paraphyly of certain genera [1,2]. The use of molecular methods in taxonomic research allows for more reliable species identification, but is still rare in this group of organisms [1,3]. Tintinnid loricae exhibit significant intraspecific polymorphism, but their morphology, i.e., differences in size, overall shape, ornamentation, fine surface structures and the presence or absence of agglutinated particles, remains the most important feature for their taxonomy.
According to the available literature, Rhizodomus tagatzi is distributed only in the Northern Hemisphere in tropical to temperate latitudes [4]. It was probably first described by Busch in 1925 as a form of Tintinnopsis karajensis in the coastal waters of Indonesia [5]. It was later recorded under the name Rhizodomus tagatzi in the Pacific coastal waters of Russia [6] and Japanese coastal waters [7], from where it spread to the Gulf of Mexico, the Caribbean Sea [8,9] and the Arabian Gulf [10] under the name Tintinnopsis corniger. In the 1980s, the species was found in the coastal waters of the western Mediterranean, where it was described as the new species Tintinnopsis nudicauda [11] and T. corniger [12]. Subsequently, specimens were determined in other Mediterranean regions: T. corniger in the central Mediterranean [13], in the eastern Mediterranean [14,15] and in the Marmara Sea [16], as T. nudicauda in the eastern Mediterranean [17] and as Rhizodomus tagatzi in the Black Sea [18]. According to the European Alien Species Information Network (EASIN) [19,20] tintinnid Rhizodomus tagatzi Strelkow and Wirketis 1950 is classified as an alien species introduced into the eastern Mediterranean Sea in 2007 [15].
This paper reports the first record of Rhizodomus tagatzi in the Adriatic Sea from plankton samples collected during the monitoring program in the Neretva River estuary in the southern part of the Adriatic Sea. The paper focuses simultaneously on the morphology of Rhizodomus tagatzi, the status of its population in the estuary and its relationships with thermohaline features of the environment, taking into account the temporal variability of all parameters. Considering that EASIN classifies the species as alien [20], the origin of this species in the Adriatic Sea is also discussed.
2. Materials and Methods
2.1. Study Area
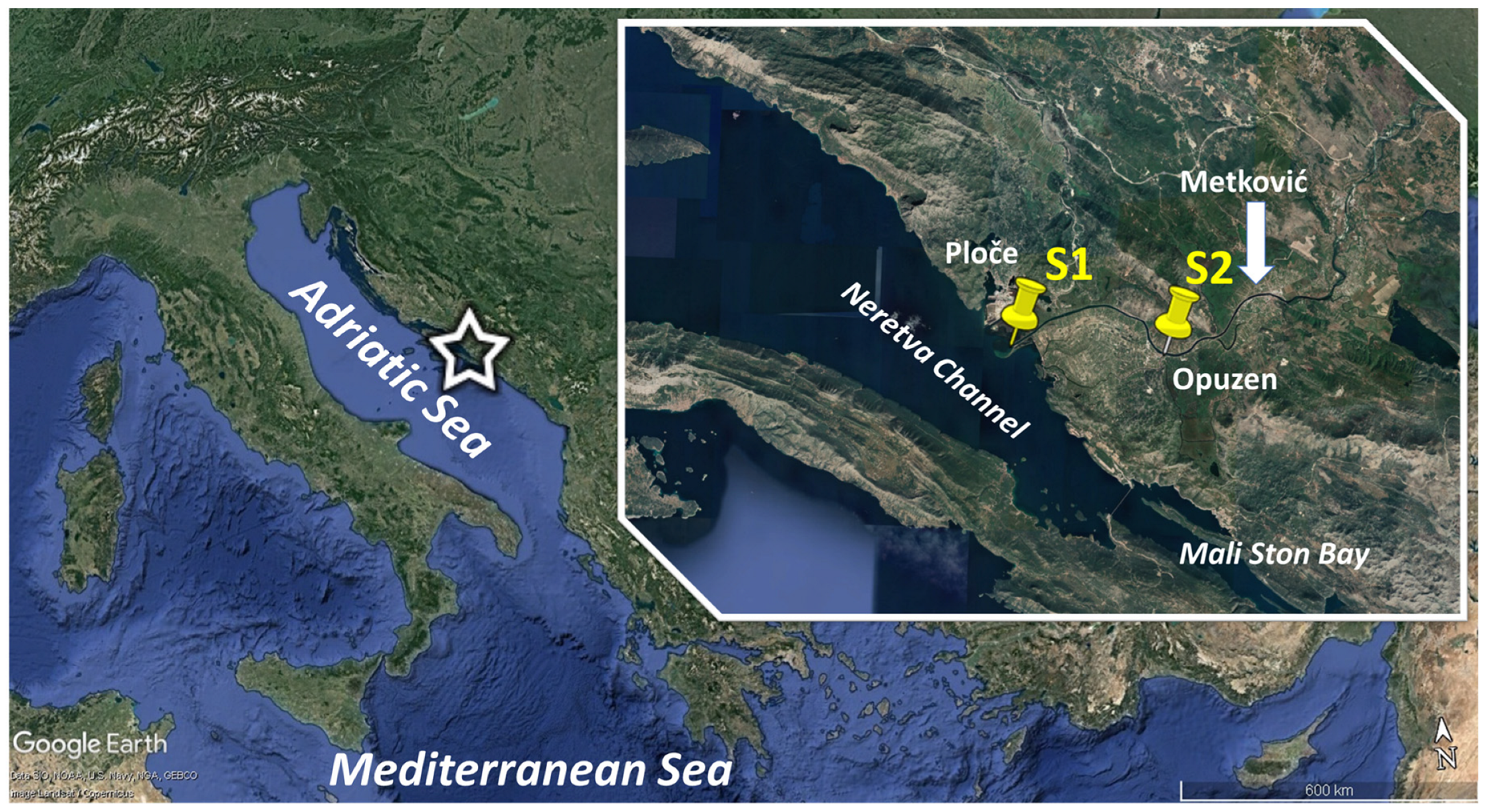
The Neretva is the largest river on the eastern coast of the Adriatic Sea, surrounded by karst hills to the north, east and south and the Adriatic Sea to the west. The river flows through Bosnia and Herzegovina (193 km) and Croatia (22 km). In the lower alluvial valley in Croatia, the Neretva divides into several river courses and forms a delta with an area of about 280 km2. The delta has been reduced by extensive land reclamation projects, and the marshes, lagoons and lakes that once characterised this plain have been largely reduced and transformed into one of the most productive agricultural areas in Croatia. In addition, the estuary and adjacent areas have been intensively exploited for various economic purposes, including transportation, fishing and more recently, tourism and recreation. The lower Neretva region consists of five ornithological, ichthyological and landscape protected areas with a total area of 1624 ha.
The Neretva River estuary is a salt-coast type, where the inflow of river water is much greater than that of sea water, and vertical mixing is limited to a thin transitional boundary between the freshwater at the surface and the salt water below [21]. The influence of the inertial effect of the sea is particularly strong when the inflows of the Neretva are low, and is observed up to above Metković, 23 km from the mouth (Figure 1). According to the author, the water regime of the Neretva River is characterised by high winter water levels (November and December and sometimes until April) and low summer water levels (June to October). The average annual flow of the Neretva in Metković is 355 m3 s−1, with a minimum of 32 m3 s−1 and a maximum of over 2400 m3 s−1 [22]. The dynamics of seawater intrusion is influenced by the tidal fluctuations of the Adriatic Sea and the upstream inflows, which depend mainly on the operation of hydroelectric power plants. The salt wedge reaches Metković at discharges of 180 m3 s−1, while no seawater can enter the riverbed at discharges higher than 500 m3 s−1 [23]. Seawater intrusion can threaten ecosystem function and affect the quality of freshwater for agriculture or water supply. Therefore, sea level rise has been identified as one of the major threats to such low-lying areas [24].
2.2. Field Sampling and Laboratory Processing
Samples were collected monthly or sometimes more frequently from May 2022 to January 2023 by a small fishing boat at two stations in the Neretva River estuary (Figure 1). Station S1 is located at the mouth of the river near the port of Ploče and station S2 is 16 km upstream from it in the main stream. The depth of both stations is 8.0 m. Zooplankton were sampled at three depths of 1 m, 5 m, and near the bottom (~7 m) using 5 L Niskin bottles. Samples were preserved in a 2.5% formaldehyde–seawater solution previously buffered with CaCO3. Preparation of samples for microscopic analysis has been described in detail in [25]. Since the cells were not visible, we counted all loricae. Counting and taxonomic identification were performed using an inverted microscope (Olympus IMT-2) at 100× and 400× magnification. The entire bottom of the sedimentation chamber was examined, and abundance was expressed as the number of cells per litre (cells L−1). To determine the morphometric characteristics of Rhizodomus tagatzi, 56 specimens were measured microscopically at 200× and 400× magnification using a calibrated ocular micrometre.
Vertical temperature and conductivity profiles were measured with a mobile probe SEBA Hidrometrie KLL-Q-2 with an accuracy of >±0.01 °C and ±0.02, respectively. Thermohaline properties were determined in parallel with the collection of zooplankton material at the same oceanographic levels.
2.3. Data Analysis
The Mann–Whitney and Kruskal–Wallis nonparametric statistical tests were used to detect differences among stations and sampling depths in total tintinnid abundance (Addinsoft 2023, XLSTAT statistical and data analysis solution. New York, NY, USA. https://www.xlstat.com). Similarity patterns between stations and sampling depths were described using non-metric multidimensional scaling (MDS) and hierarchical clustering (HC) methods [26]. The distance between the extracted factor station/depth was calculated using Bray–Curtis similarity. Clustering was performed using the group average method, with the “similarity profile” permutation test (SIMPROF) applied to test the significance of the internal structure within the constructed cluster. An MDS plot was created to illustrate the similarity, with the overlapping clusters coming from a SIMPROF cluster dendrogram. These statistical analyses were performed using PRIMER 7 software (version 7.1.21 of PRIMER-E Ltd., Plymouth, UK) [26].
3. Results
Taxonomic classification of species Rhizodomus tagatzi Strelkow and Wirketis, 1950 according to World Ciliophora Database [27]:
Class Oligotrichea
Subclass Oligotrichia
Order Choreotrichida
Suborder Tintinnina
Family Tintinnida incertae sedis
3.1. Morphological Features of Rhizodomus tagatzi
The lorica of R. tagatzi is 149.1–217.6 μm long overall in specimens preserved in formaldehyde and consists of a cylindrical bowl that gradually narrows toward the aboral end and terminates in a branched aboral horn (Table 1 and Figure 2A). The walls of the cylindrical part of the bowl appear rigid; they are flat and covered with coarse material that makes them opaque, and at the aboral end, the bowl tapers to a horn-like projection. Due to the agglutination of the lorica, it is not possible to detect the presence of spiral turns. The mouth opening is complete, rough and 28.7–36.1 μm wide. The aboral end is 34.3–56.5 μm long and consists of a large, rather flat, pointed horn surrounded by one to five smaller, single-pointed appendages. The length of these lateral buds is quite small compared to the main horn (Figure 2B,C). Only sometimes (in less than 5% of the measurements) were two main horns observed. In contrast to the rest of the lorica, the aboral end has a hyaline structure with a fine reticulation.
3.2. Distribution Pattern of Rhizodomus tagatzi and Environmental Settings in the Neretva River Estuary
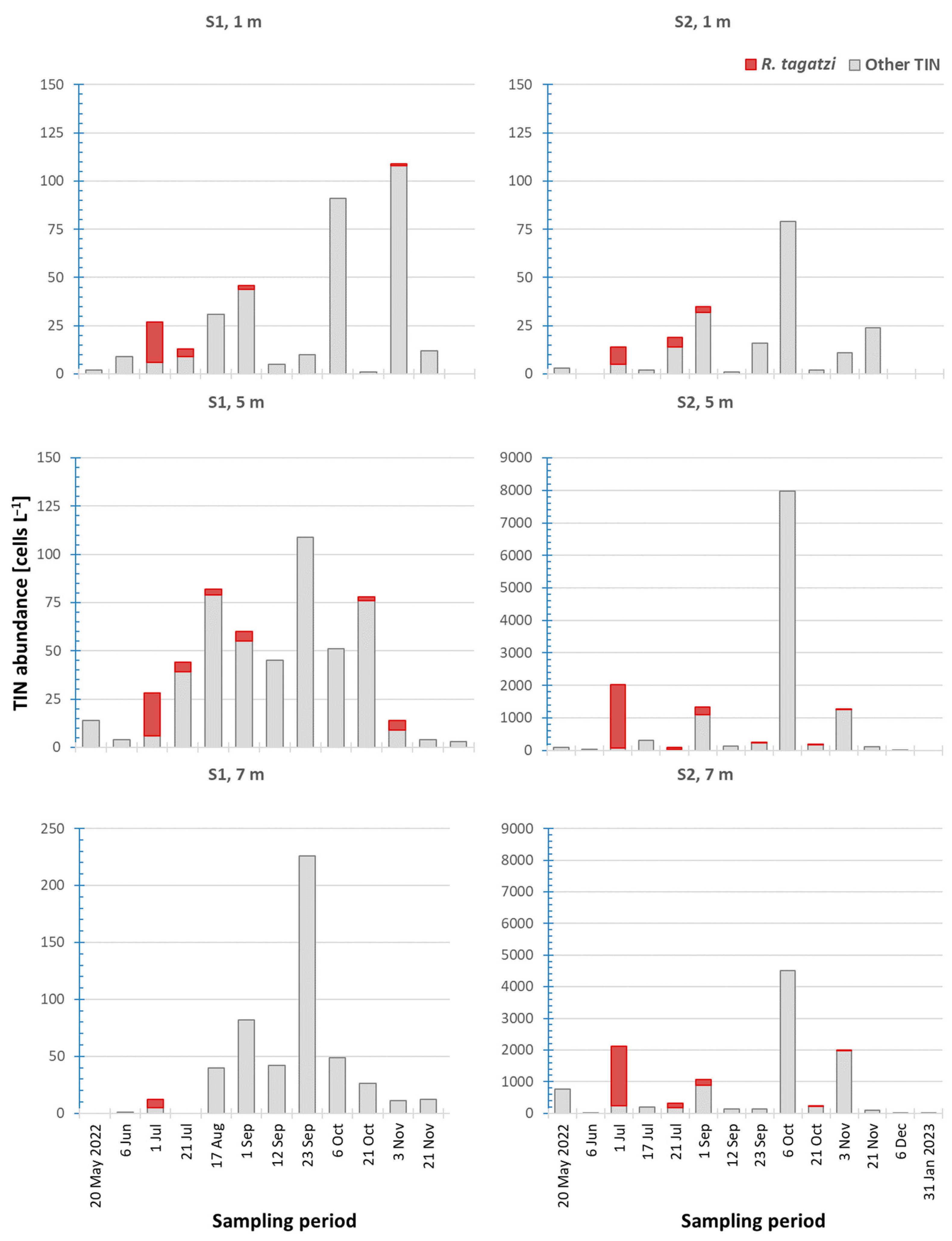
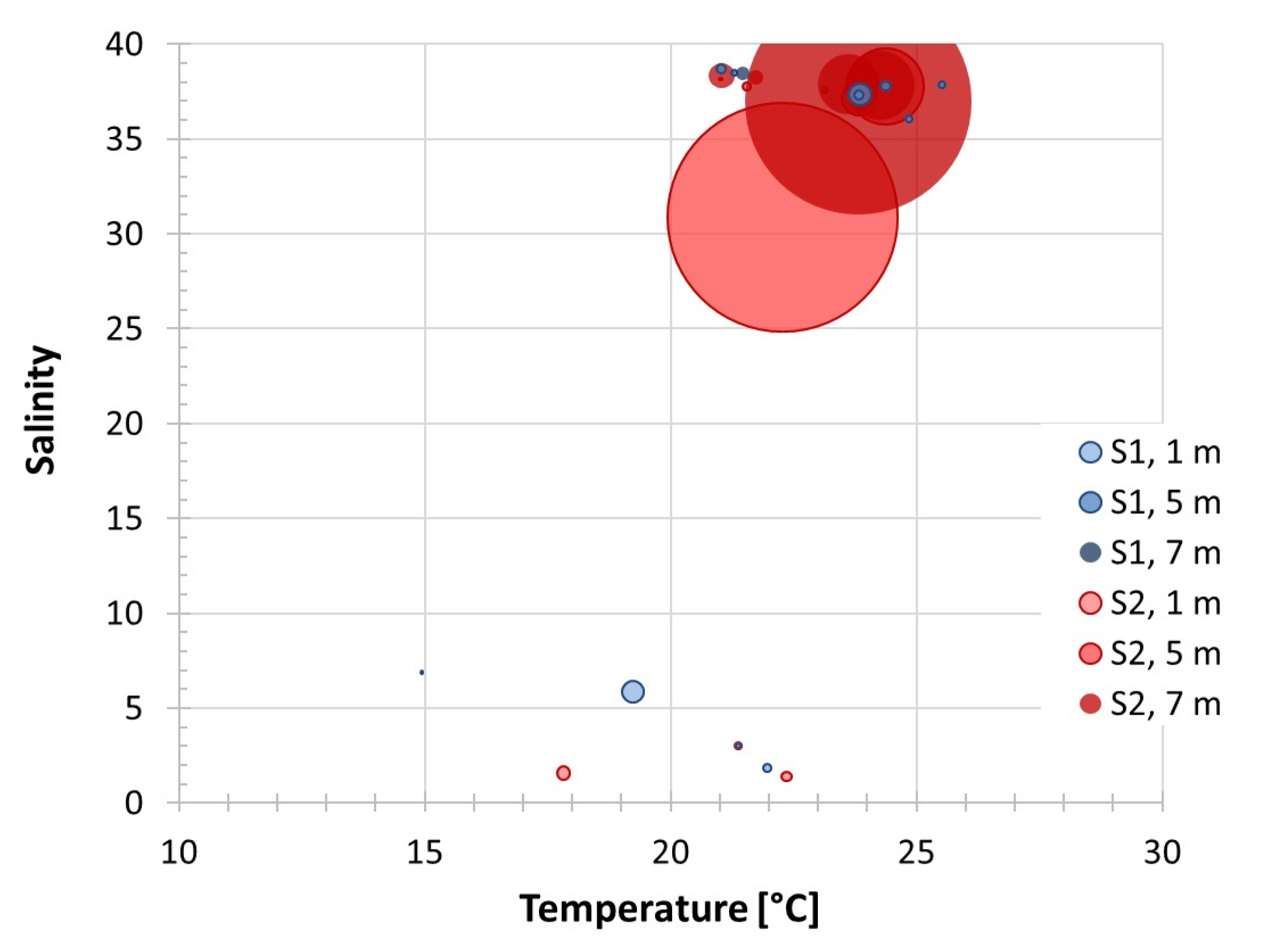
At upstream station S2, the species occurs from early July to early November, with most specimens occurring in the 5 and 7 m layers in a temperature and salinity range of 21.01–24.37 °C and 30.88–38.36, respectively (Figure 3, Figure 4 and Figure 5). The highest abundance, 1958 cells L−1, was recorded on 1 July, at 5 m depth at 22.28 °C and 30.88. In the same period, very low abundances of the species were recorded at the mouth of the Neretva River, station S1 (less than 5 cells L−1). Except for on 1 July, they were not found at a depth of 7 m, and the highest abundance was only 22 cells L−1 (5 m), at a temperature of 23.85 °C and salinity of 37.35. On this basis, we estimate the limits of hydrographic parameters for Rhizodomus tagatzi in the Neretva estuary to be a temperature of 21.01–25.51 °C and salinity of 30.88–38.71. However, a few specimens were also found at lower temperature (min. 14.94 °C) and salinity values (1.39–6.90), but due to the way the plankton samples were collected and processed, we cannot determine with certainty whether the specimens were alive.
During the study period, Rhizodomus tagatzi contributed an average percentage of 8.68 ± 20.96% on S1 and 11.19 ± 24.91% on S2 to the total tintinnid abundance. This species dominated the community at both stations in early July (Figure S1). At the upstream station (S2), an average of 83.32 ± 16.96% and a maximum of 96.83% were recorded at 5 m depth, and a very high value of 55.68% was also recorded at 5 m depth on 21 July. In contrast, at the station in the Neretva estuary (S1), these values were lower, with an average of 71.56 ± 11.46% and a maximum of 78.58% at 5 m depth on 1 July. A relatively high community percentage of 35.71% was measured at 5 m depth in early November. Statistical analysis revealed significant differences between stations in total abundance of tintinnids (U = 1029.5, p = 0.045), while differences in vertical distribution were significant only at station S1 in the surface layer compared to deeper layers (K = 15.864, p = 0.001). A list of tintinnid taxa detected in the Neretva River estuary during this study can be found in Table S1.
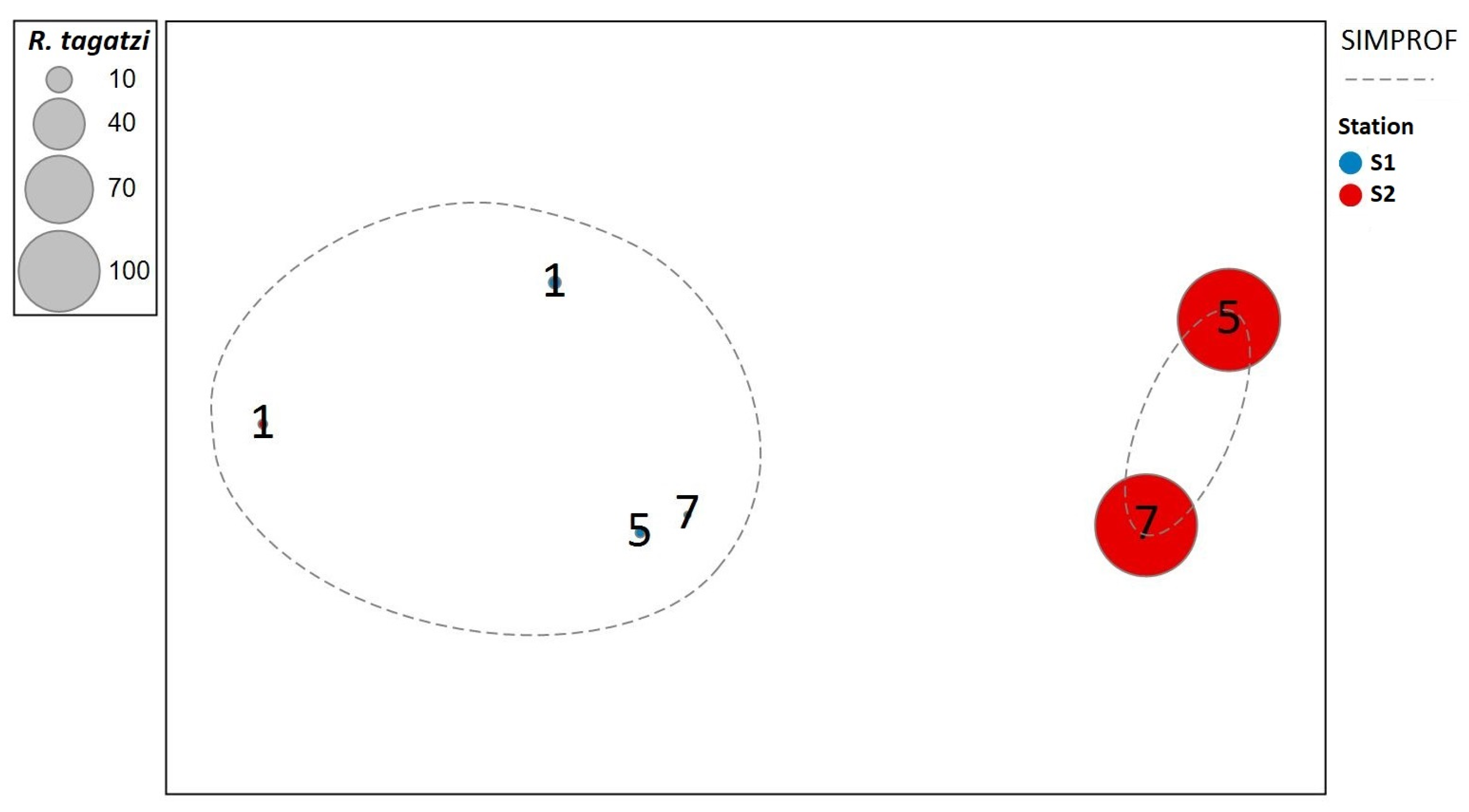
To investigate the degree of similarity of the tintinnid community in the Neretva River estuary, the total abundance of these organisms was subjected to hierarchical clustering and MDS ordination (Figure 6). According to the SIMPROF test, two clusters were statistically significant (indicated by dashed lines) at a significance level of p = 0.004: (I) the cluster comprising the sampling layers of 5 and 7 m depth at the upstream station S2, where the highest values of total abundance of tintinnids as well as R. tagatzi were detected, and (II) the cluster comprising all other samples. Although the surface layer at both stations (as two branches) is separated from the deeper layers at the confluence station (S1), the differences are not statistically significant.
4. Discussion
The tintinnid Rhizodomus tagatzi is common in ecosystems with high trophic levels and stable water column, such as the Thau Lagoon [12] and the lagoon of Urbino in the northern Mediterranean [11], the Damietta Harbour in Egypt [14], the coast of Hurghada in the Red Sea [28], Lake Faro in Sicily (central Mediterranean) [4], northern Lebanon in the eastern Mediterranean [29], Izmir Bay in the Aegean Sea [15], the Gulf of Gemlik in the Sea of Marmara [16], and Sevastopol Bay in the Black Sea [18,30]. For a more detailed overview of the geographic distribution of R. tagatzi outside the Mediterranean Sea, the reader can refer to the work of Dolan and Pierce [31] and Saccà and Giufrè [4].
In general, data on the status of plankton communities in the Neretva River estuary are still sparse and mostly limited to the lowest part of the estuary [32,33,34,35]. According to Article 2 of the Water Framework Directive (WFD 2000/60/EC), this area is classified as “transitional waters”, the main characteristics of which is the occurrence of pronounced vertical stratification of the water column in terms of salinity, nutrient concentration, oxygen content, and plankton community composition. The occurrence of stratification is caused by a relatively low amplitude of ocean changes, the energy of which is insufficient to cause significant vertical mixing of the water column [36]. The sea penetrates upstream along the Neretva riverbed into a clearly separated bottom layer of saline water upstream to behind Metković [23]. The thickness of the upper freshwater layer is variable and depends primarily on the flow of the river, the characteristics of the riverbed, and synoptic conditions.
In addition, water quality is classified as mesotrophic due to extensive anthropogenic activities in the lower estuary and its surroundings [34]. Long-term phytoplankton surveys in the area of the adjacent Mali Ston Bay have confirmed that the nanoplankton component of the phytoplankton is equally represented as the microphytoplankton in all seasons [37]. Size fractions of nano- and picoplankton are considered a suitable food source for tintinnids [38,39,40,41]. Under such environmental conditions, tintinnids can develop high population densities. Some species can rapidly adapt to a new habitat, reproduce, and reach much higher densities than in their original habitats. This phenomenon has already been observed in the oceanic species Xystonella lohmanni and Eutintinnus tubulosus in the Krka River estuary (middle Adriatic Sea) [42] and in Amphorides laackmanni in the northern part of the Adriatic Sea [43].
The high abundances (~2 × 103 cells L−1) of R. tagatzi that we found at the 16 km upstream station (S2) of the Neretva estuary in layers more than 5 m deep were due to the entry of a seawater wedge into the riverbed itself. Especially in summer, when the species was most abundant, the salt wedge penetrates up to 25 km from the mouth due to the lower flow of the river [23]. The significantly higher population densities at the upstream station (S2) compared to the estuary (S1) indicate favourable environmental conditions. Indeed, the sharp interface between layers, which in the estuary is a halocline and often a thermocline [23], is also associated with the nutricline, which creates suitable conditions for the development of phytoplankton and microbial communities, as well as microzooplankton as their primary consumers [41,44,45,46]. Depending on physical conditions such as wind-driven turbulent mixing, plankton organisms can also be transported out of the pycnocline into frontal areas and even cause a surface bloom [47]. In our study, the lowest abundance of tintinnids was found in the surface layer, indicating the lack of intense vertical mixing of the layers.
From what is known so far, the species has been observed in land-sea transition zones (estuaries, lagoons, coastal lakes) and in the neritic marine environment offshore of estuaries. It appears to tolerate a relatively wide range of salinity (16–37) and prefers relatively warmer water conditions (18–29 °C) [4]. Therefore, it was detected in samples only during the summer–autumn period of the year, with a maximum in summer, which is consistent with our results. Table 1 provides a summary overview of the abundance of the species and the temperature and salinity data in which it was detected. The preference of tintinnids for high temperatures appears to be common in many nearshore and estuarine marine waters, which may be related to better growth at higher temperatures and chlorophyll a concentrations [14]. The abundance of R. tagatzi in the Neretva River estuary is among the highest recorded in the Mediterranean and surrounding seas (Table 1). Higher values were found only in Uranouchi Inlet in Japan in summer 1997 (5 cells mL−1) [48] and in Lake Faro in July 2002 (4.5 × 103 cells L−1) [13]. The formation of dense populations and their dominance in the tintinnid community may have a strong impact on ecosystem functioning. Future research must therefore address the composition and quantity of the entire plankton community.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Abundance and environmental data for Rhizodomus tagatzi, Tintinnopsis corniger or Tintinnopsis nudicauda in the Mediterranean Sea and adjacent seas. (The bold values for temperature and salinity indicate the environmental conditions under which the highest abundance was recorded).
Table 1.
Abundance and environmental data for Rhizodomus tagatzi, Tintinnopsis corniger or Tintinnopsis nudicauda in the Mediterranean Sea and adjacent seas. (The bold values for temperature and salinity indicate the environmental conditions under which the highest abundance was recorded).
| Study Area | Tem./Sal. | Abundance/Biomass | Reference |
|---|---|---|---|
| Thau Lagoon, northern Mediterranean Sea | Warmer waters with high salinity 11.8–23.2 °C/31.5–37.9 | June 1994 25,215 µg m−3 | [12] 2 |
| Lake Faro, NE corner of Sicily, Italy (central Mediterranean Sea) | 12.5–29.3 °C/34.1–37.0 19.0 °C and 36.4 | July–September 2022 4.52 × 103 cells L−1 (July 2002) | [4,13] 1,2 |
| Damietta Harbour, Egypt (Eastern Mediterranean Sea) | 27–33 °C | July 2003 Max. total TIN abundance 73.5 × 103 cells m−3 (no data for T. corniger) | [14] 2 |
| Northern Lebanese coastal waters (Eastern Mediterranean Sea) | No data | July–October September 2006 and 2014 (7 cells L−1) August 2010 | [17] 3 |
| Novorossiysk Harbour and Gelendzhik Bay, North-eastern Black Sea | 26–28 °C | 5 × 104 cells m−3 (August-September 2015) | [49] 1 |
| Sevastopol Bay, Black Sea | Summer: 18.2–28.0 °C (23.6 ± 1.7 °C)/15.5–18.0 (17.6 ± 0.2) Autumn: 5.0–20.9 °C (14.1 ± 3.0 °C)/17.2–17.88 (17.63 ± 0.11) | summer–autumn (summer 2010: 45% of total TIN abundance) Mean 1: 12,819 cells m−3 1.01 × 105 cells m−3 (summer) | [18] 1 |
| Marmara Sea, Gulf of Gamlik, Turkish coastal waters | 18–28 °C/16–18 18 °C/16 19 °C and 18 | August–October 2010 850 cells L−1 (October) | [16] 2 |
| Al-Max Bay, Alexandria, Egypt | ~22 °C/~24 | Autumn 2014 16,902 ± 6453 cells m−3 | [50] 3 |
| Neretva River estuary, Adriatic Sea | 21.01–25.51 °C/30.88–38.71 22.28 °C and 30.88 | July–November 2022 1958 cells L−1 (1 July 2022) | This paper 1 |
1, Rhizodomus tagatzi; 2, Tintinnopsis corniger; 3, Tintinnopsis nudicauda.
A relatively small number of publications address the measurement of lorica (Table 2), and a small number of these studies are supplemented with descriptions of lorica and/or environmental parameters. The agglutinated lorica in the upper, more or less cylindrical part, with a hyaline caudal extension, resembled those of specimens of R. tagatzi from the Black Sea [49], T. nudicauda found in the Urbino Lagoon in Corsica [11] and in Lebanese coastal waters [17], and T. corniger found on the Red Sea coast near Hurghada [28] and in Izmir Bay in the coastal waters of Turkey [15]. On the other hand, differences in the appearance of the main horn and lateral branches, as well as in the agglutination of the lorica, were observed most frequently. In contrast to the specimens from the Neretva River estuary, the aboral horns are much more robust in R. tagatzi from Peter Great Bay in Russia [6], Jiaozhou Bay in China [51], and Sevastopol Bay [18], and in T. corniger from the Gulf of Gemlik in Turkish coastal waters [16]. A considerably longer caudal horn (more than one-third of the total lorica length) was found in T. corniger from the Strait of Hormoz and waters of the United Arab Emirates [10], while a completely hyaline lorica was found in R. tagatzi from Lake Faro in Sicily [52].
The intense shipping traffic that characterises the Adriatic Sea favours the expanding speed of marine organisms. Ballast water has been a vector for tintinnid transport for many years, and it may no longer be possible to determine the original distribution of many tintinnid species [53]. Ballast water has been used in maritime transport for more than a hundred years, so it is reasonable to assume that some tintinnid species, like other planktonic organisms, have successfully established populations in new locations. Although it cannot be said with certainty that every occurrence of a new species is related to ballast water, it is very likely that ballast water, which is regularly discharged in the port of Ploče during the loading of ships, served as a transmission vector for this neritic tintinnid species. The non-indigenous copepod species Pseudodiaptomus marinus [54] and a very high proportion of non-indigenous macrozoobenthic species have already been found in this port [55]. Interestingly, P. marinus was also found upstream from the port of Ploče (S2 station in our study), confirming seawater intrusion into the upper reaches of the river [56]. According to the available data, a total of 1.035 × 106 m3 of ballast water was discharged into the Port of Ploče since 2013, with the largest amount in 2021 [57]. More than 90% of the ballast discharged into this port originated from the Mediterranean Sea (70% from the Adriatic Sea). The presence of R. tagatzi in ballast water tanks [15,49,53] suggests that transport by ballast water is the likely mechanism for the spread of this species in the Adriatic. The formation of cysts is an additional advantage for the spread of this genus [58,59]. In addition to transmission through ballast water, transfer through aquaculture has also been mentioned as one of the introduction or dispersal routes for this tintinnid species [4]. According to the authors, transfer of juvenile mussels from natural spawning areas to nurseries (mussel farms) is a common practice. However, since mussel farming in Mali Ston Bay (Figure 1) has been carried out in a traditional way with autochthonous juveniles since the 16th century [60], we can consider this method of introduction of R. tagatzi into the Adriatic Sea as unlikely.
One of the problems in the identification of non-indigenous species (NIS) species is the insufficient number of studies. R. tagatzi is a relatively large tintinnid with a very peculiar morphology, so it is difficult to assume that it was not noticed in previous studies. Extensive surveys of zooplankton at the terminals used for cargo loading in the twelve Adriatic ports were conducted in 2013–2016 as part of the BALMAS project (www.balmas.eu), but the finding of this species was not reported. However, the zooplankton survey focused on mesozooplankton [61], while microzooplankton was surveyed only in four Croatian ports (Pula, Rijeka, Šibenik and Split) [62]. As a semi-enclosed sea with limited water exchange with other seas, the Adriatic Sea is highly susceptible to any form of pollution, introduction of non-indigenous species, and thus to any hazards arising from the discharge of ballast water. It is unclear what impact introduced tintinnids may have on the local food web.
There is a possibility that many introduced species will not have a significant impact at the point of introduction. However, even if an introduced organism does not cause an obvious ecological problem, it may displace native species and eventually lead to a decline in biodiversity. These introductions are expected to increase in the future. Therefore, it is of interest to monitor endangered areas to assess ecological status and identify introductions and their impact on the native community. Thus, this study could be a good basis for further research and contribute to a better understanding of the functioning of pelagic ecosystems, especially in ecologically sensitive ecosystems.
5. Conclusions
The paper documents the first occurrence of the alien tintinnid ciliate Rhizodomus tagatzi in the Adriatic Sea, in the habitat of the transitional waters of the Neretva River estuary.
The results confirm that the species occurs in the warm season (20.01–25.51 °C) and tolerates a wide range of salinity (30.88–38.71). The maximum abundance of ~2 × 103 L−1 cells is among the highest in the Mediterranean and surrounding seas and is related to the intrusion of a seawater wedge into the riverbed itself. The formation of dense populations and their dominance in the tintinnid community may have a strong impact on ecosystem functioning.
According to our study, it is very likely that ballast water is the main transmission vector for this neritic tintinnid species in the Adriatic Sea. Although this species is not currently an obvious ecological problem in the estuary, it may displace native species and reduce biodiversity. Therefore, this study could be a good basis for further research and contribute to a better understanding of the functioning of pelagic ecosystems, especially in ecologically sensitive environments.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15101821/s1, Figure S1: Relative abundance of Rhizodomus tagatzi in tintinnid community at two stations and three sampling depths in the Neretva River estuary from May 2022 to January 2023; Table S1: List of the tintinnid taxa identified at two stations in the Neretva River estuary in the period from May 2022 to January 2023 (+, present; blank, absent).
Author Contributions
Conceptualization, N.B. and D.L.; methodology, J.N. and N.B.; formal analysis, N.B.; investigation, J.N., D.L. and I.V.; resources, J.N. and D.L.; data curation, J.N.; writing—original draft preparation, N.B. and J.N.; writing—review and editing, N.B., I.V. and D.L.; visualization, N.B. and I.V.; supervision, D.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The data supporting the findings of this research are available upon reasonable request from the first author.
Acknowledgments
We thank the fishermen and residents of the Neretva estuary, especially Mile Marević for his active support during the fieldwork. We are very grateful to the reviewers who contributed to the improvement of this manuscript with their comments, valuable advice, and useful suggestions. We also thank our colleague Jasna Arapov from the Institute of Oceanography and Fisheries (Split, Croatia) who helped us with the preparation and photographing of R. tagatzi with the scanning electron microscope.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Agatha, S.; Strüder-Kypke, M.C. Phylogeny of the order Choreotrichida (Ciliophora, Spirotricha, Oligotrichea) as inferred from morphology, ultrastructure, ontogenesis, and SSr-RNA gene sequences. Eur. J. Protistol. 2007, 43, 37–63. [Google Scholar] [CrossRef] [PubMed]
- Strüder-Kypke, M.; Lynn, D.H. Morphological versus molecular data-phylogeny of tintinnid ciliates (Ciliophora, Choreotrichia) inferred from small subunit rRNA gene sequences. Denisia 2008, 23, 417–424. [Google Scholar]
- Bachy, C.; Gómez, F.; López-García, I.; Dolan, J.R.; Moreira, D. Molecular phylogeny of tintinnid ciliates (Tintinnida, Ciliophora). Protist 2012, 163, 873–887. [Google Scholar] [CrossRef] [PubMed]
- Saccà, A.; Giuffrè, G. Biogeography and ecology of Rhizodomus tagatzi, a presumptive invasive tintinnid ciliate. J. Plankton Res. 2013, 35, 894–906. [Google Scholar] [CrossRef]
- Dolan, J.R. Planktonic protists: Little bugs pose big problems for biodiversity assessment. J. Plankton Res. 2016, 38, 1044–1051. [Google Scholar] [CrossRef]
- Strelkow, A.A.; Wirketis, M.A. New planktonic infusorian (suborder Tintinnoinea) from Peter the Great Bay. Rep. USSR Acad. Sci. 1950, 74, 389–391. (In Russian) [Google Scholar]
- Hada, Y. New species of the Tintinnida found from Inland Sea. Bull. Suzugamine Women Coll. Nat. Sci. 1964, 11, 1–4. [Google Scholar]
- Balech, E. Tintinnoinea del Mediterraneo; Trabajo Numero 28; Instituto Espanol de Oceanographia: Madrid, Spain, 1968; p. 84. [Google Scholar]
- Aladro-Lubel, M.A. Distribucion de los Tintinidos (Protozoa, Ciliata) de la parte oester suroeste y sur del Golfo de Mexico. Rev. De Las Soc. Mex. De Hist. Nat. 1974, 35, 45–75. [Google Scholar]
- Sharaf, G.M. On some tintinnids (Protozoa: Ciliata) from the Strait of Hormoz and the United Arab Emirates waters. Qatar Univ. Sci. J. 1995, 15, 389–398. [Google Scholar]
- Paulmier, G. Tintinnides (Ciliophora, Oligotrichida, Tintinnina) de l’Atlantique Boréal, de l’Océan Indien et de Quelques Mers Adjacentes: Méditerranée, Mer Caraibe, Mer Rouge. Inventaire et distribution. Observations Basées sur les Loricas; Rapport IFRAMER DRV/RH-ECOHAL: L’Houmeau, France, 1997; p. 127. Available online: https://archimer.ifremer.fr/doc/00424/53609/ (accessed on 22 March 2023).
- Lam-Hoai, T.; Rougoer, C.; Lasserre, G. Tintinnids and rotifers in a northern Mediterranean coastal lagoon. Structural diversity and function through biomass estimation. Mar. Ecol. Prog. Ser. 1997, 152, 13–25. [Google Scholar] [CrossRef]
- Saccà, A.; Guglielmo, L.; Bruni, V. Vertical and temporal microbial community patterns in a meromictic coastal lake influenced by the Straits of Massina upwelling system. Hydrobiologia 2008, 600, 89–104. [Google Scholar] [CrossRef]
- Dorgham, M.M.; Abdel-Aziz, E.N.; El-Ghobashy, E.A.; El-Tohamy, S.W. Preliminary study on protozoan community in Damietta Harbor, Egypt. Glob. Vet. 2009, 3, 495–502. [Google Scholar]
- Yurga, L. Non-native two new Tintinnida (Oligotrichida) taxa in Izmir Bay: Leprotintinnus nordquisti (Brandt 1906) ve Tintinnopsis corniger (Hada, 1964). Ege J. Fish. Aqua. Sci. 2012, 29, 69–72. [Google Scholar] [CrossRef]
- Durmuş, T.; Balci, M.; Balkis, N. Species of genus Tintinnopsis Stein, 1867 in Turkish coastal waters and new record of Tintinnopsis corniger Hada, 1964. Pakisten J. Zool. 2011, 44, 383–388. [Google Scholar]
- Abbou-Abi Saab, M. Tintinnids of the Lebanese Coastal Waters (Eastern Mediterranean); CNRS-Lebanon/UNEP/MAP/RAS/SPA Lebanon: Batroun, Lebanon, 2008; p. 192. [Google Scholar]
- Gavrilova, N.A.; Dovgal, I.V. Tintinnid Ciliates (Ciliophora, Tintinnida) of the Black Sea and the Sea of Azov; A.O. Kovalevsky Institute of Biology of the Southern Seas of Ras: Sevastopol, Crimea, 2019; p. 176. (In Russian) [Google Scholar]
- Katsanevakis, S.; Bogucarskis, K.; Gatto, F.; Vandekerkhove, J.; Deriu, I.; Cardoso, A.C. Building the European Alien Species Information Network (EASIN): A novel approach for the exploration of distributed alien species data. BioInvasions Rec. 2012, 1, 235–245. [Google Scholar] [CrossRef]
- European Commission-Joint Research Centre-European Alien Species Information Network (EASIN). Available online: https://easin.jrc.ec.europa.eu/ (accessed on 10 February 2023).
- Matić, F. Air-Sea Interaction in Split, Brač and Neretva Channel. Master’s Thesis, Faculty of Science, University of Zagreb, Zagreb, Croatia, 2005; p. 75. (In Croatian). [Google Scholar]
- Samokovlija Dragičević, J. Quality of water and improvement of the delta and drainage area of the Neretva River. Građevinar 2023, 55, 759–765. (In Croatian) [Google Scholar]
- Ljubenkov, I.; Vranješ, M. Numerical model of stratified flow-case study of the Neretva river bed salination (2004). Građevinar 2012, 64, 101–113. [Google Scholar]
- Krvavica, N.; Ružić, I. Assessment of sea-level rise impacts on salt-wedge intrusion in idealized and Neretva River Estuary. Estuar. Coast. Shelf Sci. 2020, 234, 106638. [Google Scholar] [CrossRef]
- Bojanić, N.; Šolić, M.; Krstulović, N.; Šestanović, S.; Marasović, I.; Ninčević, Ž. Temporal variability and abundance and biomass of ciliates and copepods in the eutrophicated part of Kaštela Bay (Middle Adriatic Sea). Helgol. Mar. Res. 2005, 59, 107–120. [Google Scholar] [CrossRef]
- Clark, K.R.; Gorley, R.N. PRIMER v7: User Manual/Tutorial, 1st ed.; PRIMER-e Ltd.: Plymouth, UK, 2015; pp. 1–300. [Google Scholar]
- Xu, K.; Agatha, S.; Dolan, J. World Ciliophora Database. Rhizodomus tagatzi Strelkow & Wirketis, 1950. Accessed through: World Register of Marine Species 2023. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=689307 (accessed on 10 February 2023).
- Abou Zaid, M.M.; Hellal, A.M. Tiintinnids (Protozoa: Ciliata) from the coast of Hurghada Red Sea, Egypt. Egypt. J. Aquat. Res. 2012, 38, 249–268. [Google Scholar] [CrossRef]
- Abboud-Abi Saab, M.; Ouba, A. Seasonal and interannual changes in abundance and community structure of tintinnids in the Levantine Sea in relation to hydrographic conditions from 2002 to 2014. Reg. Stud. Mar. Sci. 2021, 47, 101990. [Google Scholar] [CrossRef]
- Gavrilova, N.A.; Dovgal, I.V. Tintinnid ciliates (Spirotrichea, Choreotrichia, Tintinnida) of the Black Sea: Recent invasions. Protistology 2016, 10, 91–96. [Google Scholar] [CrossRef]
- Dolan, J.R.; Pierce, R.W. Diversity and distribution of tintinnids. In The biology and ecology of tintinnid ciliates Models for marine plankton, 1st ed.; Dolan, J.R., Montagnes, D.J.S., Agatha, S., Coats, D.W., Stoecker, D.K., Eds.; Wiley-Blackwell A. John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 214–243. [Google Scholar]
- Vidjak, O.; Bojanić, N.; Kušpilić, G.; Gladan Ninčević, Ž.; Tičina, V. Zooplankton community and hidrographycal properties of the Neretva Channel (eastern Adriatic Sea). Helgol. Mar. Res. 2007, 61, 267–282. [Google Scholar] [CrossRef]
- Jasprica, N.; Hafner, D.; Batistić, M.; Kapetanović, T. Phytoplankton in three freshwater lakes in the Neretva River delta /Eastern Adriatic, NE Mediterranean). Nova Hedwig. 2005, 81, 37–54. [Google Scholar] [CrossRef]
- Jasprica, N.; Carić, M.; Kršinić, F.; Kapetanović, T.; Batistić, M.; Njire, J. Planktonic diatoms and their environment in the lower Neretva River estuary (Eastern Adriatic Sea, NE Mediterranean). Nova Hedwig. 2012, 141, 405–430. [Google Scholar]
- Bužančić, M.; Ninčević Glada, Ž.; Marasović, I.; Kušpilić, G.; Grbec, B. Eutrophication influence on phytoplankton community composition in three bays on the eastern Adriatic coast. Oceanologia 2016, 58, 302–316. [Google Scholar] [CrossRef]
- Lončar, G.; Krvavica, N.; Gotovac, H.; Oskoruš, D.; Kulić, T. Numerical analysis of the impact of the dam on the prevention of saltwater intrusion into the Neretva riverbed. Hrvat. Vode 2020, 28, 113–124. (In Croatian) [Google Scholar]
- Skejić, S.; Vilibić, I.; Matijević, S.; Jozić, S.; Ninčević Gladan, Ž.; Morović, M.; Marasović, I.; Prelesnik, H. Long-term regulating mehanisms of phytoplankton biomass in a tradicional shellfish aquaculture area. Fresenius Environ. Bull. 2015, 24, 3001–3013. [Google Scholar]
- Bojanić, N.; Šolić, M.; Krstulović, N.; Šestanović, S.; Ninčević Gladan, Ž.; Marasović, I.; Brautović, I. The role of ciliates within the microbial food web in the eutrophicated part of Kaštela Bay (middle Adriatic Sea). Sci. Mar. 2006, 70, 431–442. [Google Scholar] [CrossRef]
- Bojanić, N.; Vidjak, O.; Šolić, M.; Krstulović, N.; Brautović, I.; Matijević, S.; Kušpilić, G.; Šestanović, S.; Ninčević Gladan, Ž.; Marasović, I. Community structure and seasonal dynamics of tintinnid ciliates in Kaštela Bay (middle Adriatic Sea). J. Plankton Res. 2012, 34, 510–530. [Google Scholar] [CrossRef]
- Gavrilova, N.; Dolan, J.R. A note on species list and ecosystem shift: Black Sea Tintinnids, Ciliates of the microzooplankton. Acta Protozool. 2007, 46, 279–288. [Google Scholar]
- Šolić, M.; Krstulović, N.; Šantić, D.; Šestanović, S.; Ordulj, M.; Bojanić, N.; Kušpilić, G. Structure of microbial communities in phosphorus-limited estuaries along the eastern Adriatic coast. J. Mar. Biol. Assoc. United Kingd. 2015, 95, 1565–1578. [Google Scholar] [CrossRef]
- Kršinić, F. Tintinnids (Tintinnida, Choreotrichia, Ciliata) in the Adriatic Sea, Mediterranean Part II. Ecology; Institute of Oceanography and Fisheries: Split, Croatia, 2010; p. 116. [Google Scholar]
- Kršinić, F.; Precali, R. On the occurrence of oceanic tintinnines with a particular consideration of the species Amphorides laackmanni (Jörgensen, 1924), (Ciliophora, Oligotrichida, Tintinnina) in the northern Adriatic Sea. Mar. Ecol. 1997, 18, 67–81. [Google Scholar] [CrossRef]
- Lunven, M.; Guillaud, J.F.; Youénou, A.; Crassous, M.P.; Berric, R.; Le Gall, E.; Kérouel, R.; Labry, C.; Aminot, A. Nutrient and phytoplankton distribution in the Loire River plume (Bay of Biscay, France) resolved by a new Fine Scale Sampler. Estuar. Coast. Shelf Sci. 2005, 65, 94–108. [Google Scholar] [CrossRef]
- Cermeño, P.; Dutkiewicz, S.; Harris, R.P.; Follows, M.; Schofield, O.; Falkowski, G. The role of nutricline depth in regulation the ocean carbon cycle. Proc. Natl. Acad. Sci. USA 2009, 105, 20344–20349. [Google Scholar] [CrossRef]
- Richardson, K.; Bendtsen, J. Vertical distribution of phytoplankton and primary production in relation to nutricline depth in the open ocean. Mar. Ecol. Prog. Ser. 2019, 620, 34–46. [Google Scholar] [CrossRef]
- Tyler, M.A. Dye tracing of a subsurface chlorophyll maximum of a red-tide dinoflagellate to surface frontal regions. Mar. Biol. 1984, 78, 285–300. [Google Scholar] [CrossRef]
- Akizawa, K.; Iwasaki, N.; Ueda, H. List of microplankton in Uranouchi Inlet, Kochi, Japan during the summer of 1997. Bull. Mar. Sci. Fish. Kochi Univ. 1998, 18, 57–63. [Google Scholar]
- Selifonova, Z.P.; Makarevich, P.R. Invasive alien species of tintinnid ciliates from the northeaster Black Sea, Russian and Abkhazian coast. Protistology 2018, 12, 185–190. [Google Scholar] [CrossRef]
- El-Damhougy, K.A.; El-Sayed, A.A.; Aboul Ezz, S.M.; Abu Husein, M.S. Tintinnida (Ciliophora) as bio-indicator for certain pollutants at al-max area, Alexandria, Egypt. Int. J. Fish. Aquat. Stud. 2017, 5, 297–305. [Google Scholar]
- Feng, M.; Wang, C.; Zhang, W.; Zhang, G.; Xu, H.; Zhao, Y.; Xiao, T.; Wang, C.; Wang, W.; Bi, Y.; et al. Dataset of long-term variation in species occurrence and abundance of tintinnid assemblages in Jiaozhou Bay, China. Data Brief 2018, 19, 1856–1864. [Google Scholar] [CrossRef] [PubMed]
- Saccà, A.; Strüder-Kypke, M.C.; Lynn, D.H. Redescription of Rhizodomus tagatzi (Ciliophora: Spirotrichea: Tintinnida), based on morphology and small subunit ribosomal RNA gene sequence. J. Eukaryot. Microbiol. 2012, 59, 218–231. [Google Scholar] [CrossRef] [PubMed]
- Pierce, W.R.; Carlton, J.T.; Carlton, D.A.; Geller, J.B. Ballast water as a vector for tintinnid transport. Mar. Ecol. Prog. Ser. 1997, 149, 295–297. [Google Scholar] [CrossRef]
- Uttieri, M.; Aguzzi, L.; Aiese Cigliano, R.; Amato, A.; Bojanić, N.; Brunetta, M.; Camatti, E.; Carotenuto, Y.; Damjanović, T.; Delpy, F.; et al. WGEUROBUS–Working Group “Towards a EURopean OBservarory of the non-indigenous calanoid copepod Pseudodiaptomus marinUS”. Biol. Invasions 2020, 22, 885–906. [Google Scholar] [CrossRef]
- Spagnolo, A.; Auriemma, R.; Bacci, T.; Balković, I.; Bertasi, F.; Bolognini, L.; Cabrini, M.; Cilenti, L.; Cuicchi, C.; Cvitković, I.; et al. Non-indigenous macrozoobentic species on hard substrata of selected harbours in the Adriatic Sea. Mar. Pollut. Bull. 2019, 147, 150–158. [Google Scholar] [CrossRef]
- Lučić, D.; Onofri, I.; Garić, R.; Violić, I.; Vranješ, M.; Gangai Zovko, B.; Jurinović, J.; Njire, J.; Hure, M. Ingeression og the hydromedusa Neotima lucullana (delle chiaje, 1822) into the ecosystem of the Neretva River estuary (south-eastern Adriatic, Croatia). Acta Adriat. 2022, 63, 165–174. [Google Scholar] [CrossRef]
- Hasanspahić, N.; Pećarević, M.; Hrdalo, N.; Čampara, L. Analysos of ballast waer discharged in port–A case study of the port of Ploče (Croatia). J. Mar. Sci. Eng. 2022, 10, 1700. [Google Scholar] [CrossRef]
- Kamiyama, T. Comparative biology of tintinnid cysts. In The Biology and Ecology of Tintinnid Ciliates Models for Marine Plankton, 1st ed.; Dolan, J.R., Montagnes, D.J.S., Agatha, S., Coats, D.W., Stoecker, D.K., Eds.; Wiley-Blackwell A. John Wiley & Sons, Ltd.: Chichester, UK, 2013; pp. 171–185. [Google Scholar]
- Ganser, M.H.; Bartel, H.; Weiβenbavher, B.; Andosch, A.; Lütz-Meindl, U.; Radacher, P.; Agatha, S. A light and electron microscopical study on resting cyst on the tintinnid Schmidingerella (Alveolata. Ciliophora) including a phylogeny-aware comparison. Eur. J. Protistol. 2022, 86, 125922. [Google Scholar] [CrossRef]
- Basoli, J. Fishing in the Adriatic. Special edition; Znanje: Zagreb, Croatia, 1984; p. 392. (In Croatian) [Google Scholar]
- Vidjak, O.; Bojanić, N.; de Olazabal, A.; Benzi, M.; Brautović, I.; Camatti, E.; Hure, M.; Lipej, L.; Lučić, D.; Pansera, M.; et al. Zooplankton in Adriatic port environments: Indigenous communities and non-indigenous species. Mar. Pollut. Bull. 2019, 147, 133–149. [Google Scholar] [CrossRef]
- Ninčević Gladan, Ž.; Vidjak, O.; Žuljević, A.; Šolić, M.; Cvitković, I.; Matić Skoko, S. Port Baseline Survey; (Final Report) BALMAS project. WP5. Activity 5.1.; Institute of Oceanography and Fisheries: Split, Croatia, 2016; pp. 1–31. [Google Scholar]
Figure 1.
Study area with the sampling stations in the Neretva River estuary (Southern Adriatic Sea) (S1, 43°1′10.0″ N 17°26′36.0″ E and S2, 43°01′4.65″ N 17°33′57.7″ E).
Figure 1.
Study area with the sampling stations in the Neretva River estuary (Southern Adriatic Sea) (S1, 43°1′10.0″ N 17°26′36.0″ E and S2, 43°01′4.65″ N 17°33′57.7″ E).

Figure 2.
Photomicrographs of Rhizodomus tagatzi from the Adriatic Sea: (A) the overall lorica morphology of a specimen fixed in formaldehyde solution (scale = 50 µm), (B) aboral horn with (C) lateral buds imaged with a scanning electron microscope (SEM). [Photos by J. Njire (A) and J. Arapov and N. Bojanić (B,C)].
Figure 2.
Photomicrographs of Rhizodomus tagatzi from the Adriatic Sea: (A) the overall lorica morphology of a specimen fixed in formaldehyde solution (scale = 50 µm), (B) aboral horn with (C) lateral buds imaged with a scanning electron microscope (SEM). [Photos by J. Njire (A) and J. Arapov and N. Bojanić (B,C)].

Figure 3.
Temporal and vertical variability of Rhizodomus tagatzi and other tintinnids at two stations in the Neretva River estuary from May 2022 to January 2023.
Figure 3.
Temporal and vertical variability of Rhizodomus tagatzi and other tintinnids at two stations in the Neretva River estuary from May 2022 to January 2023.

Figure 4.
Scatterplot of Rhizodomus tagatzi abundances as a function of thermohaline water properties at two stations (S1 and S2) and three layers (1, 5 and 7 m depth) in the Neretva River estuary.
Figure 4.
Scatterplot of Rhizodomus tagatzi abundances as a function of thermohaline water properties at two stations (S1 and S2) and three layers (1, 5 and 7 m depth) in the Neretva River estuary.

Figure 5.
Vertical temperature and salinity profiles at two stations (S1 and S2) in the Neretva River estuary from May 2022 to January 2023.
Figure 5.
Vertical temperature and salinity profiles at two stations (S1 and S2) in the Neretva River estuary from May 2022 to January 2023.

Figure 6.
Non-metric multidimensional scaling (MDS) ordination of two stations (S1 and S2) and three sampling depths (1, 5 and 7 m) based on Bray–Curtis similarities from untransformed abundance data of Rhizodomus tagatzi and other tintinnids. Statistically significant separations are indicated by the dashed line at a similarity level of 6.97, Π = 9.63, and p = 0.0039. Superimposed bubbles indicate relative abundance of R. tagatzi at sampling stations and depths. (Stress = 0.01).
Figure 6.
Non-metric multidimensional scaling (MDS) ordination of two stations (S1 and S2) and three sampling depths (1, 5 and 7 m) based on Bray–Curtis similarities from untransformed abundance data of Rhizodomus tagatzi and other tintinnids. Statistically significant separations are indicated by the dashed line at a similarity level of 6.97, Π = 9.63, and p = 0.0039. Superimposed bubbles indicate relative abundance of R. tagatzi at sampling stations and depths. (Stress = 0.01).

Table 2.
Morphological characteristics and localities for the species Rhizodomus tagatzi, Tintinnopsis corniger or Tintinnopsis nudicauda. (The number in parentheses indicates the mean with standard deviation).
Table 2.
Morphological characteristics and localities for the species Rhizodomus tagatzi, Tintinnopsis corniger or Tintinnopsis nudicauda. (The number in parentheses indicates the mean with standard deviation).
| Study Area | Total Lorica Length [µm] | OLD [µm] | Horn Length [µm] | Reference |
|---|---|---|---|---|
| Peter the Great Bay, Russia (subpolar sector of the Japan Sea) | 168.3–244.8 (213.65 ± 1.44) | 30.6–35.7 (30.6) | 40.8–91.8 (70.25 ± 0.93) | [6] 1 |
| Strait of Hormoz and the United Arab Emirates waters | 166–200 | 29–31 | >1/3 total length | [10] 2 |
| Urbino Lagoon (Corsica, Western Mediterranean Sea | 170–190 | 30–35 | No data | [11] 3 |
| Lebanese coastal waters (Eastern Mediterranean Sea) | 130–190 | 27–28 | 44–46 | [17] 3 |
| Marmara Sea, Gulf of Gamlik, Turkish coastal waters | 165–170 (167 ± 1.9) | 30–33 (31 ± 1.1) | 40–45 (43 ± 2) | [16] 2 |
| Lake Faro, NE corner of Sicily (central Mediterranean Sea) | 133–250 (178) | 29–35 (32) external | 35–60 (50) | [52] 1 |
| Izmir Bay, Turkish coastal water | 185 | 33 | Up to 40 | [15] 2 |
| Sevastopol Bay, Black Sea | 133–205 | 30–40 | 44–46 | [18] 1 |
| Adriatic Sea, Neretva River estuary | 149.1–217.6 (175.3 ± 19.7) | 28.7–36.1 (32.1 ± 1.6) | 34.3–56.5 (44.7 ± 6.0) | This paper 1 |
1, Rhizodomus tagatzi; 2, Tintinnopsis corniger; 3, Tintinnopsis nudicauda.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Njire, J.; Bojanić, N.; Lučić, D.; Violić, I. First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water 2023, 15, 1821. https://doi.org/10.3390/w15101821
AMA Style
Njire J, Bojanić N, Lučić D, Violić I. First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea. Water. 2023; 15(10):1821. https://doi.org/10.3390/w15101821
Chicago/Turabian StyleNjire, Jakica, Natalia Bojanić, Davor Lučić, and Ivana Violić. 2023. "First Record of the Alien Tintinnid Ciliate Rhizodomus tagatzi Strelkow and Wirketis 1950 in the Adriatic Sea" Water 15, no. 10: 1821. https://doi.org/10.3390/w15101821
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.