
Synergistic Removal of Ciprofloxacin and Sulfamethoxazole by Lemna minor and Salvinia molesta in Mixed Culture: Implications for Phytoremediation of Antibiotic-Contaminated Water
 ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Bioassays
2.2. Physiological Evaluations
2.3. Combined Toxicity Evaluation
2.4. Relative Yield and Competitive Balance Index
2.5. Chemical Analyses
2.6. Statistical Analysis
3. Results
3.1. Antibiotic Concentrations in Growth Media at T0 and Antibiotic Degradation
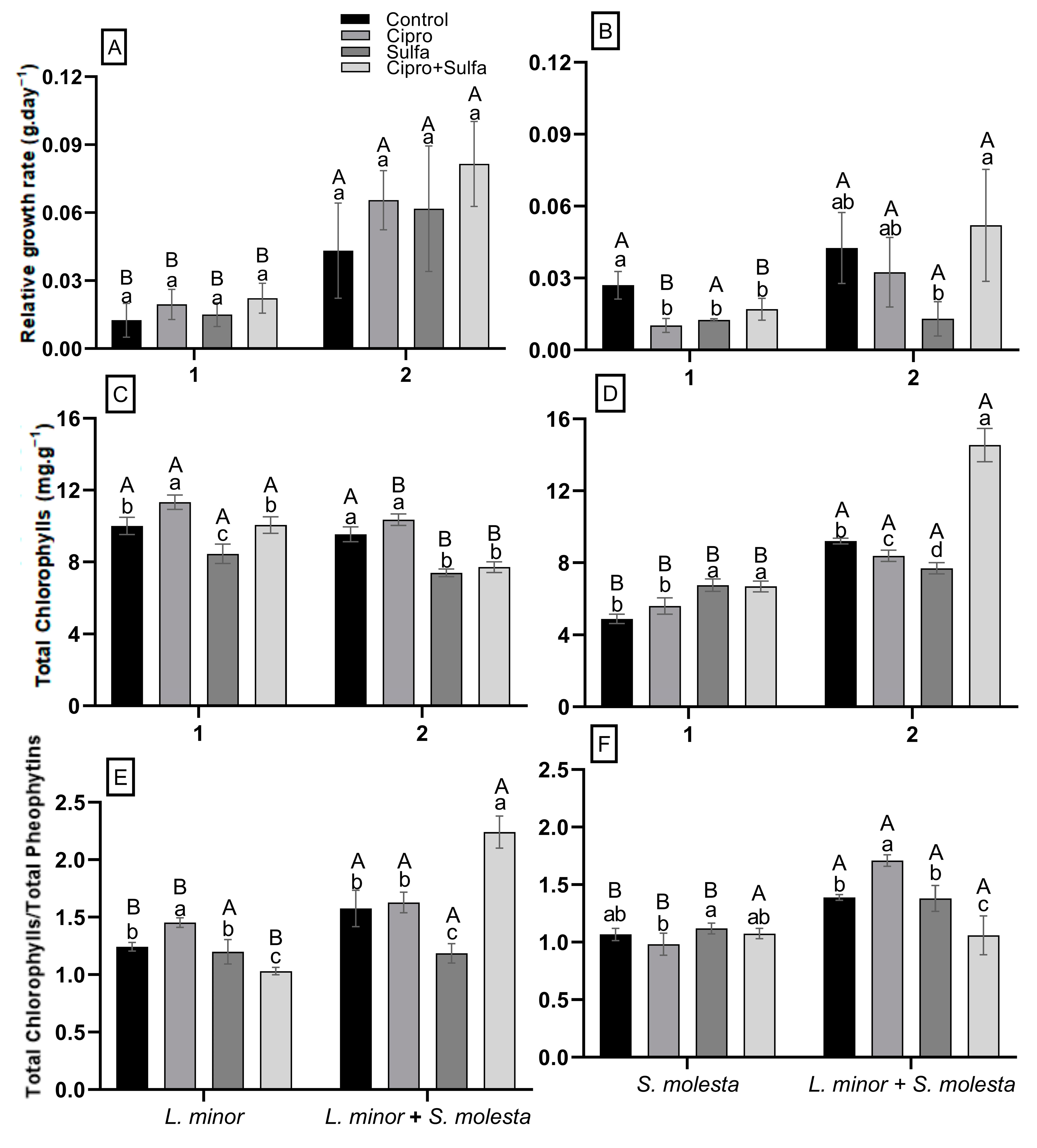
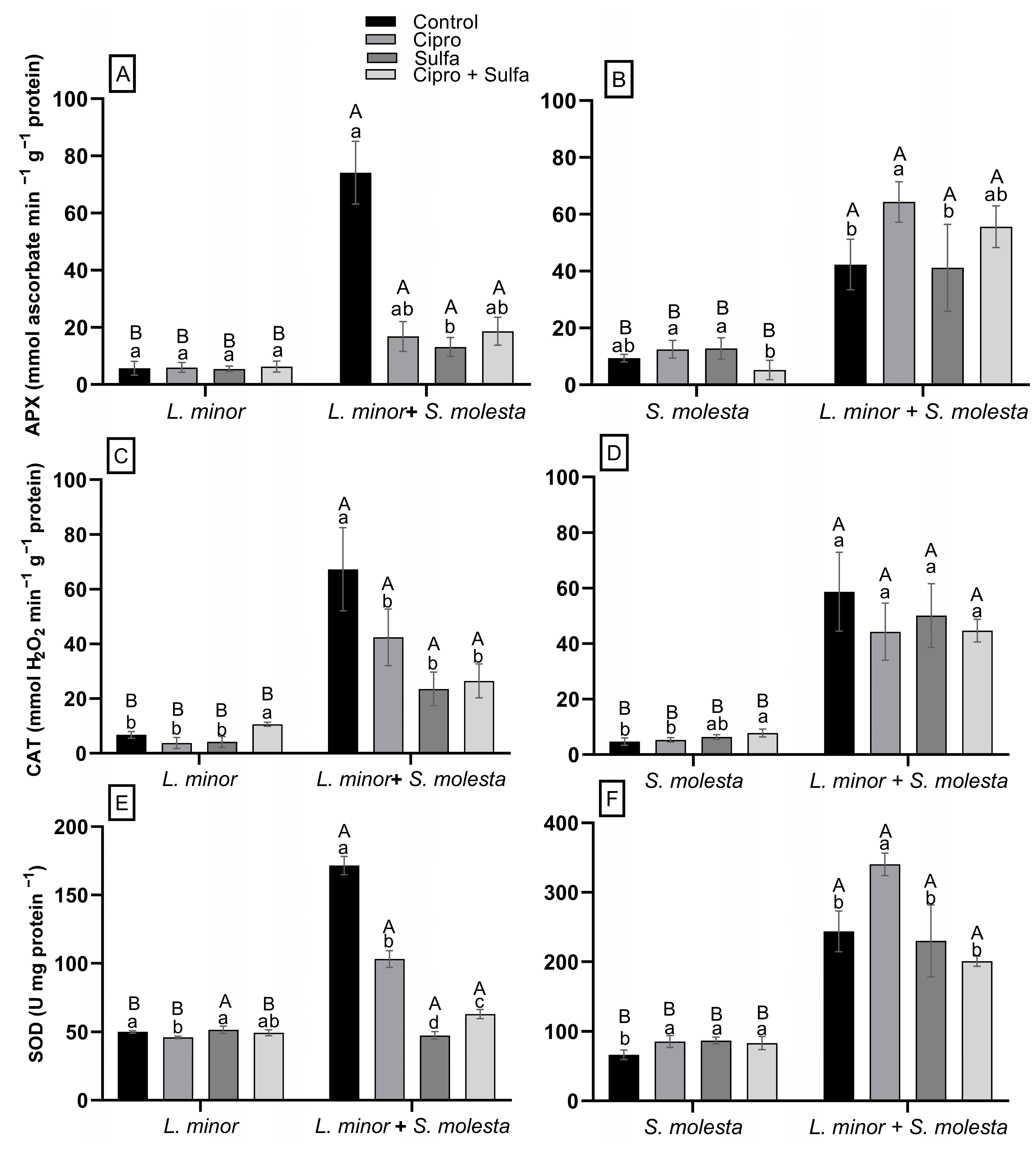
3.2. Physiological Responses of L. minor When Exposed Alone or Together with S. molesta to Antibiotics
3.3. Physiological Responses of S. molesta When Exposed Alone or Together with L. minor to Antibiotics
3.4. Combined Toxicity Evaluation
3.5. Relative Yield and Competitive Balance Index
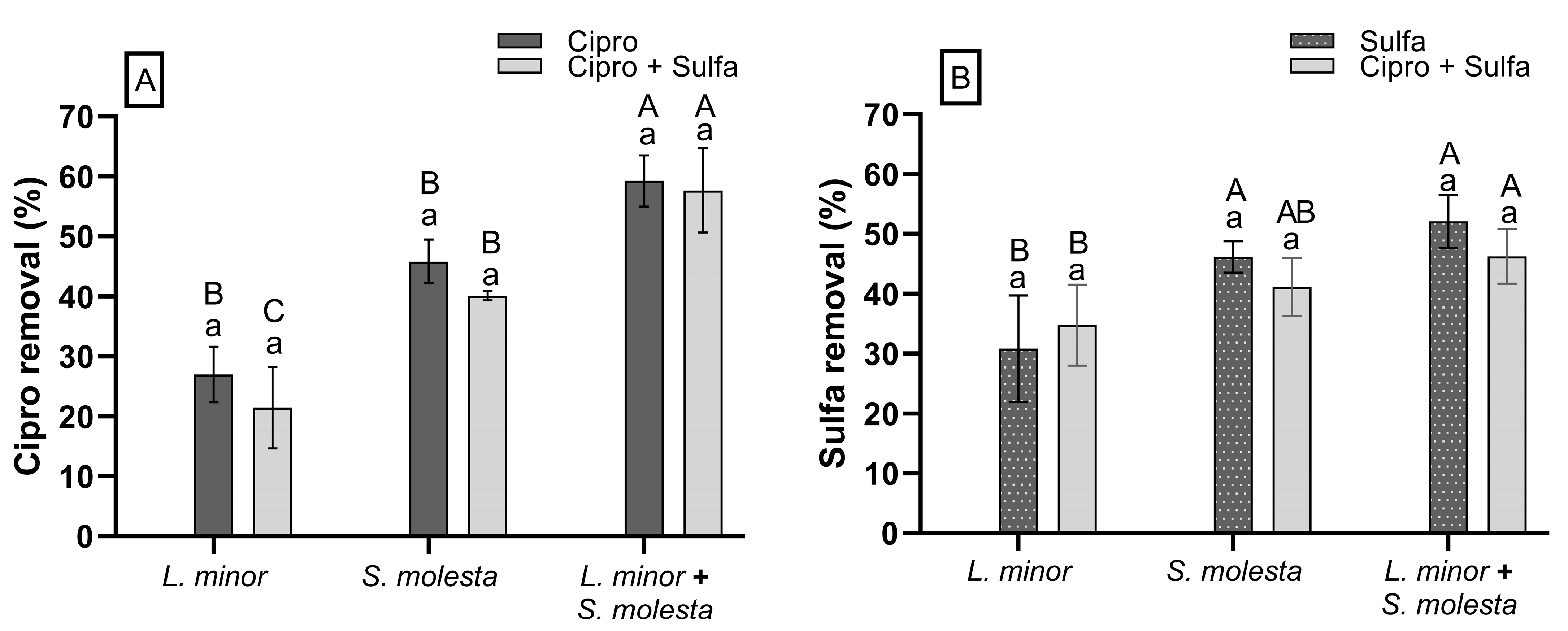
3.6. Antibiotic Removal
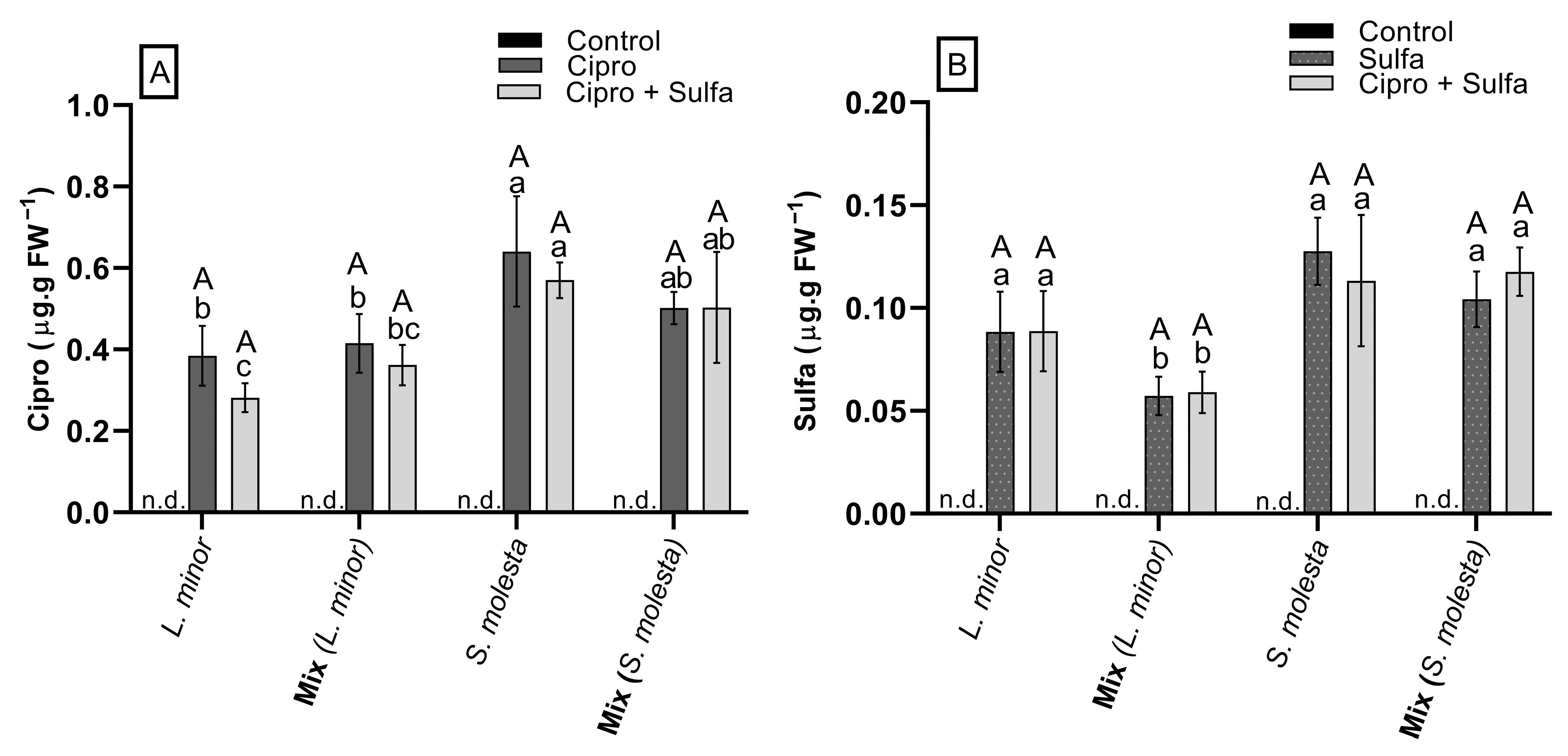
3.7. Antibiotic Concentration in Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shi, H.; Ni, J.; Zheng, T.; Wang, X.; Wu, C.; Wang, Q. Remediation of wastewater contaminated by antibiotics. A review. Environ. Chem. Lett. 2020, 18, 345–360. [Google Scholar] [CrossRef]
- Kotyza, J.; Soudek, P.; Kafka, Z.; Vaněk, T. Phytoremediation of Pharmaceuticals—Preliminary Study. Int. J. Phytoremediation 2010, 12, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Uniyal, P.L.; Prasanna, R.; Ahluwalia, A.S. Phytoremediation Potential of Aquatic Macrophyte, Azolla. AMBIO 2012, 41, 122–137. [Google Scholar] [CrossRef] [PubMed]
- Kochi, L.Y.; Freitas, P.L.; Maranho, L.T.; Juneau, P.; Gomes, M.P. Aquatic Macrophytes in Constructed Wetlands: A Fight against Water Pollution. Sustainability 2020, 12, 9202. [Google Scholar] [CrossRef]
- Maldonado, I.; Terrazas, E.G.M.; Vilca, F.Z. Application of duckweed (Lemna sp.) and water fern (Azolla sp.) in the removal of pharmaceutical residues in water: State of art focus on antibiotics. Sci. Total Environ. 2022, 838, 156565. [Google Scholar] [CrossRef]
- Bianchi, E.; Biancalani, A.; Berardi, C.; Antal, A.; Fibbi, D.; Coppi, A.; Lastrucci, L.; Bussotti, N.; Colzi, I.; Renai, L.; et al. Improving the Efficiency of Wastewater Treatment Plants: Bio-Removal of Heavy-Metals and Pharmaceuticals by Azolla filiculoides and Lemna minuta. Sci. Total Environ. 2020, 746, 141219. [Google Scholar] [CrossRef]
- Topal, M.; Öbek, E.; Şenel, G.U.; Topal, E.I.A. Removal of tetracycline antibiotic by Lemna gibba L. from aqueous solutions. Water Environ. J. 2020, 34, 37–44. [Google Scholar] [CrossRef]
- Gomes, M.P.; Brito, J.C.M.; Rocha, D.C.; Navarro-Silva, M.A.; Juneau, P. Individual and combined effects of amoxicillin, enrofloxacin, and oxytetracycline on Lemna minor physiology. Ecotoxicol. Environ. Saf. 2020, 203, 111025. [Google Scholar] [CrossRef]
- Rocha, C.S.; Kochi, L.Y.; Ribeiro, G.B.; Rocha, D.C.; Carneiro, D.N.M.; Gomes, M.P. Evaluating Aquatic Macrophytes for Removing Erythromycin from Contaminated Water: Floating or Submerged? Int. J. Phytoremediation 2022, 24, 995–1003. [Google Scholar] [CrossRef]
- Iatrou, E.I.; Stasinakis, A.S.; Aloupi, M. Cultivating Duckweed Lemna Minor in Urine and Treated Domestic Wastewater for Simultaneous Biomass Production and Removal of Nutrients and Antimicrobials. Ecol. Eng. 2015, 84, 632–639. [Google Scholar] [CrossRef]
- Kitamura, R.S.A.; Brito, J.C.M.; de Assis, H.C.S.; Gomes, M.P. Physiological responses and phytoremediation capacity of floating and submerged aquatic macrophytes exposed to ciprofloxacin. Environ. Sci. Pollut. Res. 2022, 30, 622–639. [Google Scholar] [CrossRef]
- Mendes, E.J.; Malage, L.; Rocha, D.C.; Kitamura, R.S.A.; Gomes, S.M.A.; Navarro-Silva, M.A.; Gomes, M.P. Isolated and Combined Effects of Glyphosate and Its By-Product Aminomethylphosphonic Acid on the Physiology and Water Remediation Capacity of Salvinia molesta. J. Hazard. Mater. 2021, 417, 125694. [Google Scholar] [CrossRef]
- Gomes, M.P.; Le Manac’h, S.G.; Maccario, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Differential Effects of Glyphosate and Aminomethylphosphonic Acid (AMPA) on Photosynthesis and Chlorophyll Metabolism in Willow Plants. Pestic. Biochem. Physiol. 2016, 130, 65–70. [Google Scholar] [CrossRef]
- Gulçin, İ.; Taslimi, P. Sulfonamide Inhibitors: A Patent Review 2013–Present. Expert Opin. Ther. Pat. 2018, 28, 541–549. [Google Scholar] [CrossRef]
- Picó, Y.; Andreu, V. Fluoroquinolones in soil—Risks and challenges. Anal. Bioanal. Chem. 2007, 387, 1287–1299. [Google Scholar] [CrossRef]
- Agunbiade, F.O.; Moodley, B. Pharmaceuticals as emerging organic contaminants in Umgeni River water system, KwaZulu-Natal, South Africa. Environ. Monit. Assess. 2014, 186, 7273–7291. [Google Scholar] [CrossRef] [PubMed]
- Duong, H.A.; Pham, N.H.; Nguyen, H.T.; Hoang, T.T.; Pham, H.V.; Pham, V.C.; Berg, M.; Giger, W.; Alder, A.C. Occurrence, Fate and Antibiotic Resistance of Fluoroquinolone Antibacterials in Hospital Wastewaters in Hanoi, Vietnam. Chemosphere 2008, 72, 968–973. [Google Scholar] [CrossRef]
- K’Oreje, K.; Vergeynst, L.; Ombaka, D.; De Wispelaere, P.; Okoth, M.; Van Langenhove, H.; Demeestere, K. Occurrence patterns of pharmaceutical residues in wastewater, surface water and groundwater of Nairobi and Kisumu city, Kenya. Chemosphere 2016, 149, 238–244. [Google Scholar] [CrossRef] [PubMed]
- Su, H.-C.; Ying, G.-G.; Tao, R.; Zhang, R.-Q.; Zhao, J.-L.; Liu, Y.-S. Class 1 and 2 Integrons, Sul Resistance Genes and Antibiotic Resistance in Escherichia coli Isolated from Dongjiang River, South China. Environ. Pollut. 2012, 169, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Kitamura, R.S.A.; Marques, R.Z.; Barbato, M.L.; Zámocký, M. The Role of H2O2-Scavenging Enzymes (Ascorbate Peroxidase and Catalase) in the Tolerance of Lemna minor to Antibiotics: Implications for Phytoremediation. Antioxidants 2022, 11, 151. [Google Scholar] [CrossRef]
- Gorham, P.R.; McLachlan, J.; Hammer, U.T.; Kim, W.K. Isolation and Culture of Toxic Strains of Anabaena Flos-Aquae (Lyngb.) de Bréb. SIL Proc. 1922–2010 1964, 15, 796–804. [Google Scholar] [CrossRef]
- Jacinavicius, F.R.; Gama, W.A.; Azevedo, M.T.d.P.; Sant’anna, C.L. Manual Para Cultivo de Cianobactérias. Inst. de Botânica 2013, 32, 1–27. [Google Scholar]
- Zagatto, P.A.; Aragão, M.A. Implantação de Métodos Para Avaliação de Algas Tóxicas; Universidade de São Paulo: São Paulo, Brazil, 1992. [Google Scholar]
- Oecd Test No. 221: Lemna Sp. Growth Inhibition Test. In OECD Guidelines for the Testing of Chemicals, Section 2; OECD: Paris, France, 2006; pp. 1–26. [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of Total Carotenoids and Chlorophylls a and b of Leaf Extracts in Different Solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Vernon, L.P. Spectrophotometric Determination of Chlorophylls and Pheophytins in Plant Extracts. Anal. Chem. 1960, 32, 1144–1150. [Google Scholar] [CrossRef]
- Velikova, V.; Yordanov, I.; Edreva, A. Oxidative stress and some antioxidant systems in acid rain-treated bean plants: Protective role of exogenous polyamines. Plant Sci. 2000, 151, 59–66. [Google Scholar] [CrossRef]
- Hodges, D.M.; DeLong, J.M.; Forney, C.F.; Prange, R.K. Improving the Thiobarbituric Acid-Reactive-Substances Assay for Estimating Lipid Peroxidation in Plant Tissues Containing Anthocyanin and Other Interfering Compounds. Planta 1999, 207, 604–611. [Google Scholar] [CrossRef]
- Beyer, W.F., Jr.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Nakano, Y.; Asada, K. Hydrogen Peroxide is Scavenged by Ascorbate-specific Peroxidase in Spinach Chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Aebi, H. Catalase In Vitro. ScienceDirect 1984, 105, 121–126. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Iswarya, V.; Sharma, V.; Chandrasekaran, N.; Mukherjee, A. Impact of Tetracycline on the Toxic Effects of Titanium Dioxide (TiO2) Nanoparticles towards the Freshwater Algal Species, Scenedesmus obliquus. Aquat. Toxicol. 2017, 193, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Chesworth, J.; Donkin, M.; Brown, M. The interactive effects of the antifouling herbicides Irgarol 1051 and Diuron on the seagrass Zostera marina (L.). Aquat. Toxicol. 2004, 66, 293–305. [Google Scholar] [CrossRef]
- Bottollier-Curtet, M.; Planty-Tabacchi, A.-M.; Tabacchi, E. Competition between young exotic invasive and native dominant plant species: Implications for invasions within riparian areas. J. Veg. Sci. 2013, 24, 1033–1042. [Google Scholar] [CrossRef]
- Wilson, J.B. The Effect of Initial Advantage on the Course of Plant Competition. Oikos 1988, 51, 19–24. [Google Scholar] [CrossRef]
- Palmada, J.; March, R.; Torroella, E.; Espigol, C.; Baleri, T. Determination of Enrofloxacin and Its Active Metabolite (Ciprofloxacin) at the Residue Level in Broiler Muscle Using HPLC with Fluorescence Detector. Proc. Euroresidue IV. Residues Vet. Drugs Food 2000, 23, 3078–3083. [Google Scholar]
- Migliore, L.; Cozzolino, S.; Fiori, M. Phytotoxicity to and Uptake of Enrofloxacin in Crop Plants. Chemosphere 2003, 52, 1233–1244. [Google Scholar] [CrossRef]
- Böger, B.; Vilhena, R.d.O.; Fachi, M.M.; Concentino, V.; Junkert, A.M.; Santos Josiane, M.M.F.; Domingos, E.L.; Zamora, P.G.-P.; Pontarolo, R. HPLC-MS/MS Method for Quantification of Pharmaceuticals in Subtropical Rivers and Water Treatment Plants in Brazil. J. Environ. Sci. Public Health 2020, 4, 390–408. [Google Scholar] [CrossRef]
- Gomes, M.P.; Gonçalves, C.A.; de Brito, J.C.M.; Souza, A.M.; Cruz, F.V.D.S.; Bicalho, E.M.; Figueredo, C.C.; Garcia, Q.S. Ciprofloxacin induces oxidative stress in duckweed (Lemna minor L.): Implications for energy metabolism and antibiotic-uptake ability. J. Hazard. Mater. 2017, 328, 140–149. [Google Scholar] [CrossRef]
- Gomes, M.P.; de Brito, J.C.M.; Carneiro, M.M.L.C.; da Cunha, M.R.R.; Garcia, Q.S.; Figueredo, C.C. Responses of the nitrogen-fixing aquatic fern Azolla to water contaminated with ciprofloxacin: Impacts on biofertilization. Environ. Pollut. 2018, 232, 293–299. [Google Scholar] [CrossRef]
- Mao, H.; Yang, H.; Xu, Z.; Yang, Y.; Zhang, X.; Huang, F.; Wei, L.; Li, Z. Microplastics and Co-Pollutant with Ciprofloxacin Affect Interactions between Free-Floating Macrophytes. Environ. Pollut. 2023, 316, 120546. [Google Scholar] [CrossRef]
- Lv, Y.; Xu, J.; Xu, K.; Liu, X.; Guo, X.; Lu, S.; Xi, B. Accumulation Characteristics and Biological Response of Ginger to Sulfamethoxazole and Ofloxacin. Environ. Pollut. 2020, 262, 114203. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liu, X.; Lv, Y.; Guo, X.; Lu, S. Response of Cyperus Involucratus to Sulfamethoxazole and Ofloxacin-Contaminated Environments: Growth Physiology, Transportation, and Microbial Community. Ecotoxicol. Environ. Saf. 2020, 206, 111332. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Tavares, D.S.; Richardi, V.S.; Marques, R.Z.; Wistuba, N.; de Brito, J.C.M.; Soffiatti, P.; Sant’anna-Santos, B.F.; da Silva, M.A.N.; Juneau, P. Enrofloxacin and Roundup® interactive effects on the aquatic macrophyte Elodea canadensis physiology. Environ. Pollut. 2019, 249, 453–462. [Google Scholar] [CrossRef]
- Gomes, M.P.; Rocha, D.C.; de Brito, J.C.M.; Tavares, D.S.; Marques, R.Z.; Soffiatti, P.; Sant’anna-Santos, B.F. Emerging contaminants in water used for maize irrigation: Economic and food safety losses associated with ciprofloxacin and glyphosate. Ecotoxicol. Environ. Saf. 2020, 196, 110549. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, R.S.A.; Marques, R.Z.; Kubis, G.C.; Kochi, L.Y.; Barbato, M.L.; Maranho, L.T.; Juneau, P.; Gomes, M.P. The phytoremediation capacity of Lemna minor prevents deleterious effects of anti-HIV drugs to nontarget organisms. Environ. Pollut. 2023, 329, 121672. [Google Scholar] [CrossRef]
- Gao, Z.-Y.; Wang, S.-C.; Zhang, Y.-X.; Liu, F.-F. Single and combined toxicity of polystyrene nanoplastics and copper on Platymonas helgolandica var. tsingtaoensis: Perspectives from growth inhibition, chlorophyll content and oxidative stress. Sci. Total Environ. 2022, 829, 154571. [Google Scholar] [CrossRef]
- Gomes, M.P.; Kubis, G.C.; Kitamura, R.S.A.; Figueredo, C.C.; Nogueira, K.D.S.; Vieira, F.; Navarro-Silva, M.A.; Juneau, P. Do anti-HIV drugs pose a threat to photosynthetic microorganisms? Chemosphere 2022, 307, 135796. [Google Scholar] [CrossRef]
- Rocha, D.C.; Rocha, C.D.S.; Tavares, D.S.; Calado, S.L.D.M.; Gomes, M.P. Veterinary antibiotics and plant physiology: An overview. Sci. Total Environ. 2021, 767, 144902. [Google Scholar] [CrossRef]
- Barbosa, K.B.F.; Costa, N.M.B.; Alfenas, R.C.G.; Paula, S.O.; Minim, V.P.R.; Bressan, J. Estresse oxidativo: Conceito, implicações e fatores modulatórios. Rev. Nutr. 2010, 23, 629–643. [Google Scholar] [CrossRef]
- Nurnaeimah, N.; Mat, N.; Mohd, K.S.; Badaluddin, N.A.; Yusoff, N.; Sajili, M.H.; Mahmud, K.; Adnan, A.F.M.; Khandaker, M.M. The Effects of Hydrogen Peroxide on Plant Growth, Mineral Accumulation, as Well as Biological and Chemical Properties of Ficus deltoidea. Agronomy 2020, 10, 599. [Google Scholar] [CrossRef]
- Khandaker, M.M.; Boyce, A.N.; Osman, N. The Influence of Hydrogen Peroxide on the Growth, Development and Quality of Wax Apple (Syzygium samarangense, [Blume] Merrill & L.M. Perry Var. Jambu Madu) Fruits. Plant Physiol. Biochem. 2012, 53, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Chételat, A.; Reymond, P.; Farmer, E.E. Selective and Powerful Stress Gene Expression in Arabidopsis in Response to Malondialdehyde. Plant J. 2004, 37, 877–888. [Google Scholar] [CrossRef]
- Yalcinkaya, T.; Uzilday, B.; Ozgur, R.; Turkan, I. The Roles of Reactive Carbonyl Species in Induction of Antioxidant Defence and ROS Signalling in Extreme Halophytic Model Eutrema parvenu and Glycophytic Model Arabidopsis thaliana. Environ. Exp. Bot. 2019, 160, 81–91. [Google Scholar] [CrossRef]
- Vollenweider, S.; Weber, H.; Stolz, S.; Chételat, A.; Farmer, E.E. Fatty acid ketodienes and fatty acid ketotrienes: Michael addition acceptors that accumulate in wounded and diseased Arabidopsis leaves. Plant J. 2008, 24, 467–476. [Google Scholar] [CrossRef]
- Liang, X.; Qian, R.; Wang, D.; Liu, L.; Sun, C.; Lin, X. Lipid-Derived Aldehydes: New Key Mediators of Plant Growth and Stress Responses. Biology 2022, 11, 1590. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Garcia, Q.S.; Barreto, L.C.; Pimenta, L.P.S.; Matheus, M.T.; Figueredo, C.C. Allelopathy: An overview from micro- to macroscopic organisms, from cells to environments, and the perspectives in a climate-changing world. Biologia 2017, 72, 113–129. [Google Scholar] [CrossRef]
- Gostyńska, J.; Pankiewicz, R.; Romanowska-Duda, Z.; Messyasz, B. Overview of Allelopathic Potential of Lemna minor L. Obtained from a Shallow Eutrophic Lake. Molecules 2022, 27, 3428. [Google Scholar] [CrossRef]
- Bich, T.T.N.; Kato-Noguchi, H. Allelopathic Potential of Two Aquatic Plants, Duckweed (Lemna minor L.) and Water Lettuce (Pistia stratiotes L.), on Terrestrial Plant Species. Aquat. Bot. 2012, 103, 30–36. [Google Scholar] [CrossRef]
- Hussain, N.; Abbasi, T.; Abbasi, S. Vermiremediation of an invasive and pernicious weed salvinia (Salvinia molesta). Ecol. Eng. 2016, 91, 432–440. [Google Scholar] [CrossRef]
- Sandanandan, L.; Chandran, C.; Geethu, C.B. Studies on the Biochemistry and Allelopathy of Salvinia molesta D.S. Mitchell. Trends Biosci. 2018, 11, 1569–1571. [Google Scholar]
- Dodgen, L.K.; Ueda, A.; Wu, X.; Parker, D.R.; Gan, J. Effect of Transpiration on Plant Accumulation and Translocation of PPCP/EDCs. Environ. Pollut. 2015, 198, 144–153. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Monoculture | Co-Culture | ||||||
|---|---|---|---|---|---|---|---|---|
| Cipro (µg L−1) | Sulfa (µg L−1) | Cipro (µg L−1) | Sulfa (µg L−1) | |||||
| T0 | T7 | T0 | T7 | T0 | T7 | T0 | T7 | |
| Control | n.d. | n.d | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| Cipro | 1.480 ± 0.066 a | 1.087 ± 0.061 b | n.d. | n.d. | 1.550 ± 0.086 a | 1.137 ± 0.128 b | n.d. | n.d. |
| Sulfa | n.d. | n.d. | 0.285 ± 0.050 a | 0.142 ± 0.029 b | n.d. | n.d. | 0.373 ± 0.064 a | 0.191 ± 0.020 b |
| Cipro + Sulfa | 1.473 ± 0.061 a | 1.088 ± 0.094 b | 0.283 ± 0.051 a | 0.130 ± 0.020 b | 1.498 ± 0.050 a | 1.103 ± 0.095 b | 0.383 ± 0.033 a | 0.117 ± 0.019 b |
| Index | Culture | Species | Observed Inhibition (%) | Expected Inhibition (%) | RI | Interaction |
|---|---|---|---|---|---|---|
| Growth rate | Monoculture | L. minor | 78.00 ± 52.40 | −87.20 | −0.89 ± 0.60 | antagonistic |
| S. molesta | 60.46 ± 1.34 | 82.42 | −0.44 ± 0.20 | antagonistic | ||
| Co-culture | L. minor | 88.43 ± 43.31 | −116.22 | −0.76 ± 0.37 | antagonistic | |
| S. molesta | 47.28 ± 15.48 | 76.88 | 0.28 ± 0.70 | additive | ||
| Chlorophylls | Monoculture | L. minor | −15.57 ± 5.39 | 4.43 | −1.21 ± 0.19 | antagonistic |
| S. molesta | 36.56 ± 6.08 | −58.01 | −0.63 ± 0.10 | antagonistic | ||
| Co-culture | L. minor | −19.11 ± 3.14 | 15.76 | 0.10 ± 1.04 | additive | |
| S. molesta | 58.05 ± 10.05 | 23.86 | 2.43 ± 0.42 | synergistic | ||
| H2O2 | Monoculture | L. minor | 14.61 ± 15.48 | 6.93 | 2.10 ± 1.06 | synergistic |
| S. molesta | −36.57 ± 5.81 | 37.72 | −0.96 ± 0.15 | antagonistic | ||
| Co-culture | L. minor | 57.24 ± 42.74 | −51.85 | −1.10 ± 0.82 | antagonistic | |
| S. molesta | −8.68 ± 1.40 | 15.68 | −0.55 ± 0.89 | antagonistic | ||
| MDA | Monoculture | L. minor | 2.20 ± 13.23 | 10.78 | 0.20 ± 1.22 | antagonistic |
| S. molesta | −34.04 ± 6.10 | 29.44 | −1.15 ± 0.20 | antagonistic | ||
| Co-culture | L. minor | −1.28 ± 1.18 | 0.52 | −2.45 ± 2.26 | antagonistic | |
| S. molesta | 32.12 ± 6.56 | −1.32 | −24.27 ± 4.94 | antagonistic | ||
| APX | Monoculture | L. minor | 10.00 ± 3.28 | −0.18 | −54.57 ± 179.07 | antagonistic |
| S. molesta | −44.29 ± 39.95 | −82.19 | 0.53 ± 0.44 | antagonistic | ||
| Co-culture | L. minor | −74.83 ± 6.57 | 95.96 | −0.77 ± 0.06 | antagonistic | |
| S. molesta | 31.40 ± 17.25 | −47.90 | −0.62 ± 0.36 | antagonistic | ||
| CAT | Monoculture | L. minor | 57.39 ± 10.97 | 65.42 | 0.87 ± 0.16 | antagonistic |
| S. molesta | 64.57 ± 30.37 | −51.30 | −1.25 ± 0.59 | antagonistic | ||
| Co-culture | L. minor | 60.64 ± 8.25 | 77.95 | −0.77 ± 0.11 | antagonistic | |
| S. molesta | −23.85 ± 6.88 | 35.55 | −0.67 ± 0.19 | antagonistic | ||
| SOD | Monoculture | L. minor | −1.32 ± 6.88 | 5.22 | −0.25 ± 0.82 | antagonistic |
| S. molesta | 25.42 ± 14.24 | −68.55 | −0.37 ± 0.20 | antagonistic | ||
| Co-culture | L. minor | −63.23 ± 1.96 | 83.36 | −0.75 ± 0.02 | antagonistic | |
| S. molesta | −17.89 ± 2.94 | −31.82 | 0.55 ± 0.09 | antagonistic |
| Treatments | RY L. minor | RY S. molesta | CBI L. minor |
|---|---|---|---|
| Control | 3.58 a | 1.59 b | 0.81 b |
| Cipro | 3.47 a | 3.20 a | 0.08 d |
| Sulfa | 4.13 a | 1.00 b | 1.41 a |
| Cipro + Sulfa | 3.68 a | 3.05 a | 0.18 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kochi, L.Y.; Kitamura, R.S.A.; Rocha, C.S.; Brito, J.C.M.; Juneau, P.; Gomes, M.P. Synergistic Removal of Ciprofloxacin and Sulfamethoxazole by Lemna minor and Salvinia molesta in Mixed Culture: Implications for Phytoremediation of Antibiotic-Contaminated Water. Water 2023, 15, 1899. https://doi.org/10.3390/w15101899
Kochi LY, Kitamura RSA, Rocha CS, Brito JCM, Juneau P, Gomes MP. Synergistic Removal of Ciprofloxacin and Sulfamethoxazole by Lemna minor and Salvinia molesta in Mixed Culture: Implications for Phytoremediation of Antibiotic-Contaminated Water. Water. 2023; 15(10):1899. https://doi.org/10.3390/w15101899
Chicago/Turabian StyleKochi, Leticia Yoshie, Rafael Shinji Akiyama Kitamura, Camila Silva Rocha, Julio Cesar Moreira Brito, Philippe Juneau, and Marcelo Pedrosa Gomes. 2023. "Synergistic Removal of Ciprofloxacin and Sulfamethoxazole by Lemna minor and Salvinia molesta in Mixed Culture: Implications for Phytoremediation of Antibiotic-Contaminated Water" Water 15, no. 10: 1899. https://doi.org/10.3390/w15101899