
Seasonal Changes in Upper Thermal Tolerances of Freshwater Thai Fishes
by
 and
and
Sampan Tongnunui
1,*,
Treerat Sooksawat
2,3,*,
Charoonroj Chotwiwatthanakun
4,
Weerayuth Supiwong
5,
Amnuay Wattanakornsiri
6 and
and
F. W. H. Beamish
7,8 1
Conservation Biology, Mahidol University, Kanchanaburi Campus, Lum Sum, Sai York, Kanchanaburi 71150, Thailand
2
Department of Microbiology, Faculty of Science, Khon Kaen University, Khon Kaen 40002, Thailand
3
Visiting Professor Program, Conservation Biology, Mahidol University, Kanchanaburi Campus, Lum Sum, Sai York, Kanchanaburi 71150, Thailand
4
Center of Excellence for Shrimp Molecular Biology and Biotechnology (Centex Shrimp), Faculty of Science, Mahidol University, Nakhonsawan Campus, Bangkok 10400, Thailand
5
Faculty of Interdisciplinary Studies, Khon Kaen University, Nong Khai Campus, Mueang, Nong Khai 43000, Thailand
6
Department of Agriculture and Environment, Faculty of Science and Technology, Surindra Rajabhat University, Mueang, Surin 32000, Thailand
7
Environmental Science Program, Faculty of Science, Burapha University, Bangsean, Chonburi 20131, Thailand
8
Department of Zoology, University of Guelph, Guelph, ON N1G2W1, Canada
*
Authors to whom correspondence should be addressed.
Water 2023, 15(2), 350; https://doi.org/10.3390/w15020350
Submission received: 9 December 2022
/
Revised: 6 January 2023
/
Accepted: 8 January 2023
/
Published: 14 January 2023
(This article belongs to the Special Issue Impacts of Human Activities and Climate Change on Freshwater Fish, Volume II)
Abstract
:Seasonal change inferred to climate change inevitably influences Critical thermal maximum (CTmax) of riverine fishes. In this study, we investigated CTmax as thermal tolerance for four common riverine fishes, i.e., Danio regina, Channa gachua, Rasbora caudimaculata and Mystacoleucus chilopterus, in the Kwae Noi river system in western Thailand. The acute thermal tolerance was lower in the wet season (mean river temperature ∼25 °C) and higher in the dry season (mean river temperature ∼23 °C) with medians of wet season-CTmax for those four fishes of 35.3 ± 0.4, 36.2 ± 0.5, 37.3 ± 0.5 and 37.5 ± 0.6 °C, respectively, and high values of dry season-CTmax of 37.4 ± 0.5, 38.3 ± 0.5, 38.7 ± 0.7 and 39.1 ± 0.5 °C, respectively. The variations of CTmax for all of the four species in this study, throughout the wet and dry seasons, attribute to their seasonal plasticity in response to the dynamics of thermal stress. Under climate variability and climate change with increasing the higher temperatures of air and river, and altering the habitat, R. caudimaculata and M. chilopterus had higher capacities to tolerate the acute heat stress across wet and dry seasons.
1. Introduction
Among the other effects of climate change, the increase in atmospheric greenhouse gases from anthropogenic activities in the world (especially carbon dioxide) has led to a rising average global temperature and seasonal variation. It is expected that the peak and average of diurnal water temperature will experience gradual increase in accordance with higher air temperature. Interestingly, the over-release of greenhouse gases influences both global atmospheric circulation and ocean circulation linked to redistribution change of the daylength [1,2]. The influence of the climate change has not been focused with regards to the daylength change compared with the AT and water temperature (WT) increasing. However, both WT and photoperiod are the important environmental cues impacting fishes’ physiological tolerance, which is considered as an adaptive mechanism in response to the seasonal climate change.
Seasonal changes on the earth occur annually and affect a multitude of responses in fish, especially in growth and reproduction [3,4]. WT and photoperiod are environmental factors associated with seasonal changes [5,6]. The annual seasons in the places located in the temperate (mid-latitude) regions are distinctly different in the four-season year which can be marked by the weather conditions as spring, summer, autumn and winter. In tropical regions (equator), i.e., Southeast Asian countries including Thailand, winter is less associated with dramatic changes in temperature and lower rainfall and is referred to as dry season; summer provides higher rainfall and is referred to as the wet season.
Fish are poikilothermal species and are very sensitive to changes in surrounding temperature. Tropical riverine fishes living in a relatively small thermal fluctuation environment and close to upper thermal limits are vulnerable to increases in small rivers providing high biodiversity of fish, including rare, endemic and stenothermal species. Rising temperatures in the small rivers can affect all levels of their biological and ecological organization (individual, population, community and ecosystem).
Lethal temperatures of high latitude ectothermic fishes [7] tend to be higher in summer and lower in winter, in accordance with higher and lower ambient temperatures during these respective seasons [8,9]. Freshwater fishes in low latitude regions experience a smaller seasonal variation in ambient temperatures [10] and the scant information suggests the difference between maximum ambient and lethal temperature is less than those in high latitude regions [11]. Furthermore, the thermal tolerance capacity of the ectothermic animals including fish is likely to depend on the photoperiod experiences relating to the seasonal cycle [12,13,14,15]. The seasonal change in photoperiods have been suggested as being an important environmental cue driving specific metabolisms underlaying the behavior, physiology and phenology of fish [16] but, unfortunately, the photoperiod has been less mentioned in previous studies [15].
Freshwater fishes confronted by persistent high ambient temperatures may respond by moving to a more favorable habitat, except where physical, chemical or behavior barriers prohibit such a change. Adaptations have been accepted for understanding fish distribution in relation to temperature and referring to the development of physiological, behavioral or genetic characteristics that enable an organism to cope with environmental change [17,18]. Thermal adaptations are less well understood for tropical riverine fishes, particularly species from Southeast Asian countries, including Thailand. From searching the information available for a small number of species and interpreting a variety of methods, fishes from relatively stable environments, especially in the tropics, are sensitive to small changes in environmental temperature [7].
WT may impose lethal and sub-lethal effects on riverine fish, particularly impacting metabolism, appetite, growth, maturity and host of other inter-related activities [19,20,21]. Lethal temperatures impose their effects at low and high temperatures and are generally measured by either of two procedures. Critical thermal maximum or minimum (CTmax or CTmin) is measured by heating or cooling water at a constant rate that continues to a defined end point, such as a loss in equilibrium or respiratory movement, that suggests an imminent death. When the rate of temperature increases or decreases it usually accommodates, or at least should accommodate, an alignment of deep body temperature with ambient [22]. The second common procedure, incipient upper or lower lethal temperatures, exposes fish to an abrupt change in a static temperature, upward or downward that continues until death occurs [8]. Both CTmax or CTmin, and incipient upper or lower lethal temperatures, are also called upper or lower thermal tolerance [23].
In this study, we measured CTmax as thermal tolerance for four common Thai fishes, acclimated to 25.0 °C, a common average temperature in low-order Thai rivers [24], at the beginning and the end of the wet and dry seasons. Devario regina and Rasbora caudimaculata, are endemic species in Thailand, but a high abundance are found in head rivers in western Thailand [25]. Mystacoleucus chilopterus and Channa gachua are found in high abundance in head rivers and widely distributed throughout Thailand. These four species were representatives for studying the seasonal changes in heat tolerance of some of the population of Thai riverine freshwater fishes to cope with the future climate variability and change. Moreover, we compared the CTmax values for seasonal changes in tolerance of fishes from high latitude regions, and the CTmax values among those fishes with current seasonal maximum ambient temperatures in small rivers in Western Thailand.
2. Materials and Methods
2.1. Study Area
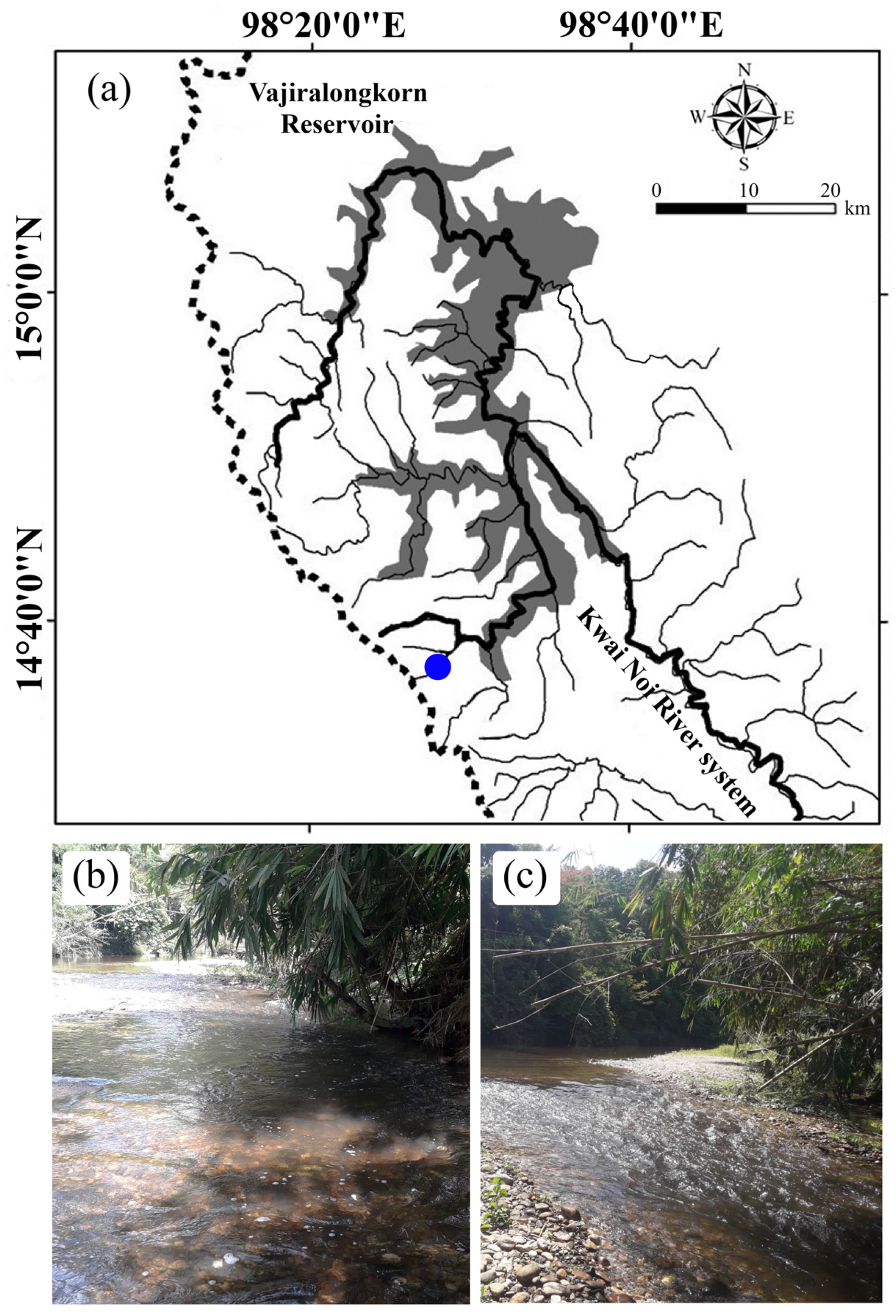
To study the CTmax of freshwater fish under the seasonal change, the Pakkok River (latitude 14°36′22″ N, longitude 98°28′14″ E), a tributary of Kwae Noi River system, in Kanchanburi province, Western Thailand (Figure 1), was considered as the study area. The sampling location, Pakkok river, the distance of which is 27 km from the border of Myanmar. This river is covered by Thongphaphoom national park, a highly pristine region contributing to high biodiversity of fish with a complexity of trophic levels [26,27] and has rare agricultural areas and few local villages with sparse populations [28]. The meteorological data during the period of 2015 to 2022 were sourced from Thai Meteorological Department (TMD) for evaluation of average AT, median of annual AT, and annual time series of average AT over Thailand. The median of annual AT was computed by the monthly AT. The maximum AT and minimum AT during the study period were collected from the meteorological station at Thong Pha Phum located near the Pakkok River. The photoperiod in Kanchanaburi province in each season was evaluated from the primary data provided by Thai Astronomical Society. The ambient river temperatures or water temperatures (WT) during the study period of wet season (May and August) and dry season (November and February) of 2017 and 2022 were monitored. Physiochemical measurements for Pakkok River in wet and dry season are shown in Table 1.
2.2. Fish Samples
Four fishes, D. regina, R. caudimaculata, M. chilopterus and C. gachua, were collected by the kick sampling technique from Pakkok River on eight occasions between 2017 and 2022. Fish were collected during daylight hours and the CTmax was measured at intervals of four months during mid-May (early wet season), mid-August (late wet season), mid-November (early dry season) and mid-February (late dry season). The sample size of the tested animal of each fish species was calculated using G*Power program [29] based on the power of test at 0.9, significant level at 0.05, and effect size = 0.6812803, which determined from our preliminary result of CTmax. The number of each fish required for two replicates of CTmax testing on each month was 20 individuals at actual power of 0.9015975. The average sizes (g) of D. regina, R. caudimaculata, M. chilopterus and C. gachua chosen for the experiment were 2.30 ± 0.12 (in range of 2.00–2.60), 1.69 ± 0.16 (in range of 1.50–2.00), 4.35 ± 0.14 (in range of 4.2–4.6), and 2.98 ± 0.09 (in range of 2.80–3.20), respectively. The experiments during 2017 followed the ethical principle and guideline for the use and care of animal in science, which were approved by the Ethics Committee at Faculty of Science, Burapha University. For the experiment during 2022, the animal care and use license was U1-07864–2561, approved by Mahidol University, Thailand.
2.3. Fish Rearing Condition
In preparation for each season’s measurement, fish were captured in Pakkok River and transported within 2–3 h to our laboratory in insulated tanks incorporated with the heating element (300 W heater; coded 3018, LifeTech; digital thermoregurator, model XT121C, Dixell, Italy; and J type thermocouple) to control the temperature (25.0 ± 0.1 °C). Thereafter, the fish were housed in the animal facility consisting of three units of the recirculating systems (RS) at greenhouse laboratory in Mahidol University, Kanchanaburi Campus. The unit of RS system comprised six 15-L fish tanks, solid separation tank, microbead filter, biofilter, heating tank, water pump and oxygen and aeration system. For each species, 10 individuals of similar weight were assigned to each of two holding RS tanks and acclimated in recirculating freshwater of RS system under the acclimation temperature at 25.0 °C and natural photoperiod (~12:12 and ~11:13 for the wet and dry seasons, respectively) for two days prior to CTmax experiment. The water temperature during the acclimation was allowed to fluctuate in a narrow range of 24.9–25.1 °C. The holding RS tanks were filled with lightly chlorinated tap water. The water flow rate in RS system was 600 L·h−1. Water was replaced daily. The water qualities were measured twice daily at approximately 10 h and 20 h with calibrated probes including DO (±0.01 mg O2·L−1; Hanna, models HI9147), pH (±0.1 units; Hanna, models HI98127) and total ammonia (±0.01mg NH3·L−1; Merck-NOVA, Model 60). The DO, total ammonia, and pH in each RS system were controlled at >75% of air saturation, and <0.001 mg NH3·L−1 and 7.3–7.5, respectively.
2.4. CTmax Measurement
CTmax was taken as the calculated highest temperatures fish tolerated for 1 h following increases of 1.0 °C·h−1 above the acclimation temperature (thermal tolerance). The experimental end point was the cessation of respiratory movements as applied earlier by Tongnunui and Beamish [17]. Fish were deprived of food for 24 h following the final feeding on the second day in the holding RS tank. Thereafter, fish were transferred to an experimental RS tank (15 L) at 25 °C and food was withheld for an additional 12 h. An earlier study found the total of 36 h of food deprivation was sufficient for complete gastric evacuation, minimizing metabolic stress [24].
CTmax measurements began by increasing the temperature 1.0 °C above 25.0 °C, which was held for 1 h. Stepwise hourly increases of 1.0 °C were continued until individual fish sequentially exhibited a visible absence of opercular movements when death was assumed. It is noted that this endpoint may not always represent fish death, but a qualitative assessment in the present study suggested it did for the majority of fish and it is less subjective than behavioral irregularities sometimes applied in these studies [5]. All fish were removed when opercula movements ceased. Afterward, they were blotted with tissues to remove excessive water and weight (0.01 g). CTmax was calculated as the highest temperature at which an individual survived for a full hour plus the time fraction of the unit temperature during which opercular movements ceased. CTmax was calculated for each species and season as the mean highest temperature that fish (n = 20) tolerated for 1 h following abrupt increments of 1 °C·h−1 above the 25 °C acclimation temperature. Chemical measurements were made in each RS tank at the beginning and end of each experiment to ensure that the water quality is maintained during the CTmax experiment, i.e., 7.5–8.5 mg O2·L−1; pH 7.1–7.8; and 0.00–0.02 mg NH3·L−1.
2.5. Statistical Analysis
The relationship between AT and WT in the study area was tested using two methods: (1) linear correlation using Pearson’s method and (2) simple linear regression (GraphPad, Prism version 9.111, San Diego, CA, USA). Paired replications of CTmax measurements for each species and month (seasons) were tested for differences by t-test; p < 0.05. CTmax values among pooled replicates for each of the four species were tested for significant differences by analysis of variance. Post hoc comparisons, Tukey HSD (Honestly Significant Difference) were applied to distinguish differences among the four months (seasons) for each of four species; p < 0.05 was accepted for significant value. Minitab statistical analysis program, version 17.0 was used to test mean significance value.
3. Results
3.1. Environmental Data
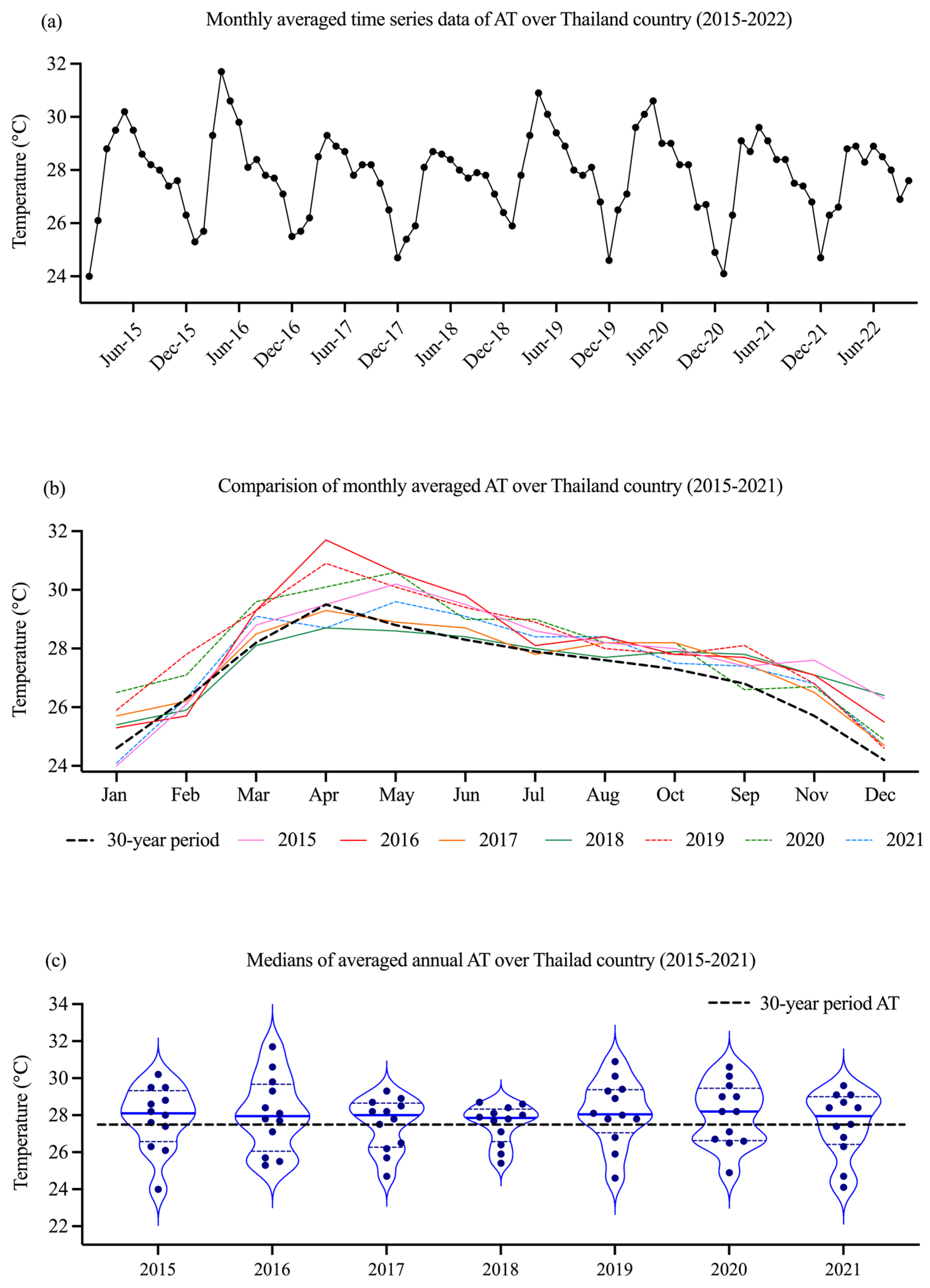
Rivers are among the most sensitive ecosystems to the effects of seasonal change and climate change scenario. Monthly averaged AT during 2015 and 2022 indicated that the month high and the month low AT were April (28.7–31.7 °C) and December (24.2–26.4 °C), respectively (Figure 2a,b). Seasonally, the periods showing differences in AT were wet season (May–October) and mid dry season (December–January). However, there was no considerable difference between summer season (March–April) and early wet season (May–June). As the Figure 2b, the monthly averaged AT from 2015 to 2022 showed a trend to be higher than that of monthly averaged AT during previous 30-year period (Figure 2b). The medians of annual AT over Thailand (Figure 2c) have been increasing between 27.9 °C and 28.2 °C during 2015 to 2021, when compared with the previous 30-year period AT (27.5 °C).
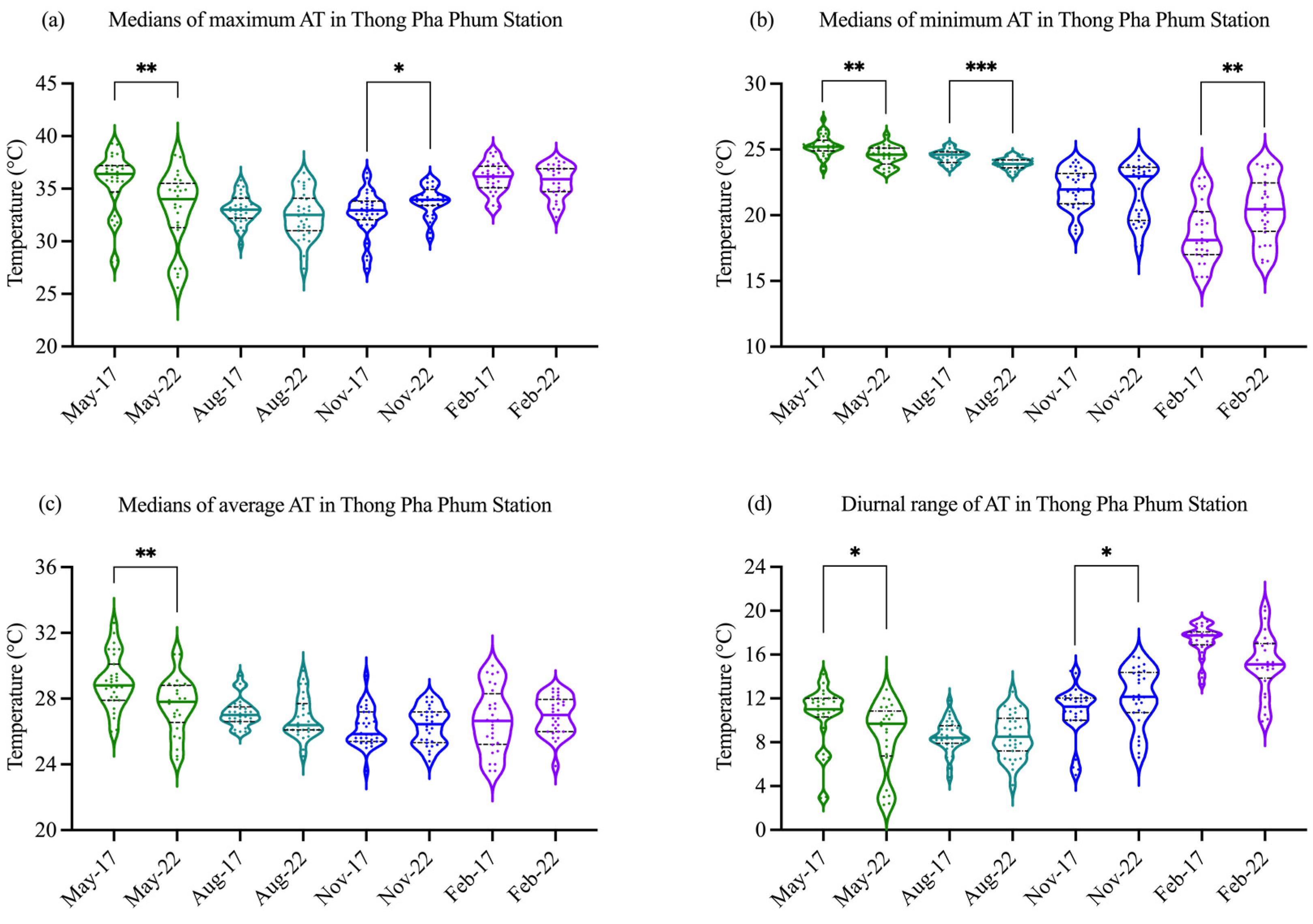
In Thong Pha Phum, located near the Pakkok River, the maximum and minimum temperatures were demonstrated in Figure 3a,b. During the study period for CTmax testing on 2017 and 2022, the maximum AT observed in the dry season (November and February) varied between 27.4 °C and 38.5 °C, and the maximum AT observed in the wet season (May and August) varied between 25.6 °C and 39.3 °C. As the Figure 3b, the medians of minimum AT observed in November and February of the dry season (18.1–22.9 °C) lower than that of May and August of the wet season (23.9–25.2 °C). The medians of average AT observed in May, a month of the early wet season, were higher than those observed in August, November and February (Figure 3c). The difference between the daily maximum and daily minimum AT (diurnal range of daily AT) during a specified time interval of the dry and wet seasons varied by about 5.0–20.4 °C and 2.3–14.2 °C, respectively (Figure 3d). Ambient hours of daylight at the river site varied seasonally with the shortest in November (~11.6 h) and February (~11.8 h) and longest in August (~12.7 h) and May (~12.8 h), respectively.
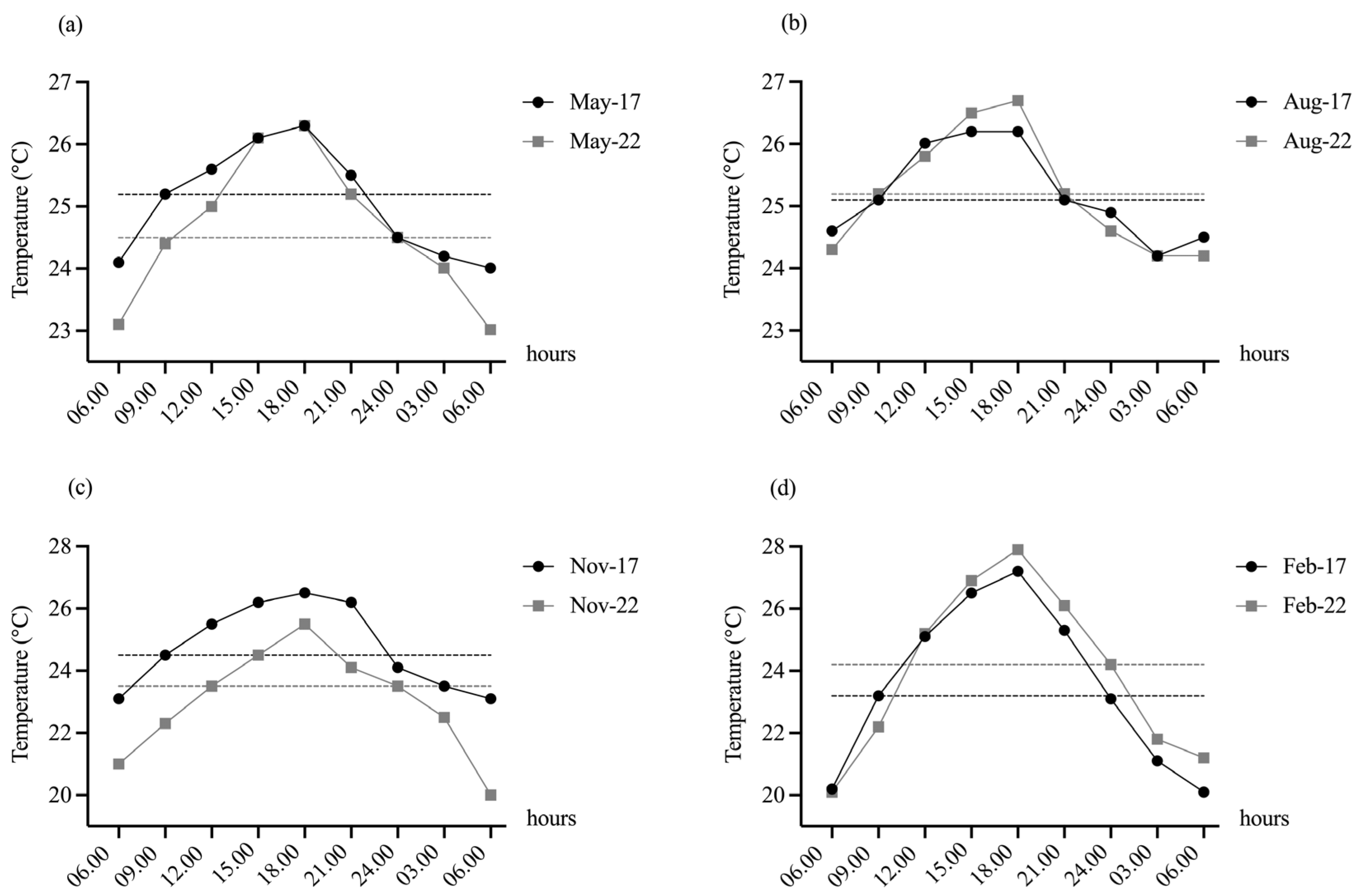
During the study periods, the WT of the Pakkok River at our field site represented the fluctuation in the stream that expose to the wild native fishes shown in Figure 4. The daily WT of the Pakkok River on November of 2017 and 2022 of dry season varied between 20.0 °C and 26.5 °C (Figure 4a). While the daily WT of the Pakkok River on February of 2017 and 2022 fluctuated 20.0 °C and 27.0 °C (Figure 4b). The fluctuation of ambient WT on the dry season varied by 3.4–7.8 °C (Figure 4c,d), which was higher than that observed on the wet season of the study period showing the daily fluctuation of ambient WT varied by 2.0–3.3 °C (Figure 4a,b). The lowest and highest average river temperatures of the wet season observed in the Pakkok River were about 23.0 °C and 26.7 °C, respectively. Average daylight ambient river temperature varied by about 5–6 °C with average daily maximum temperature varying seasonally from approximately 20 to 27 °C.
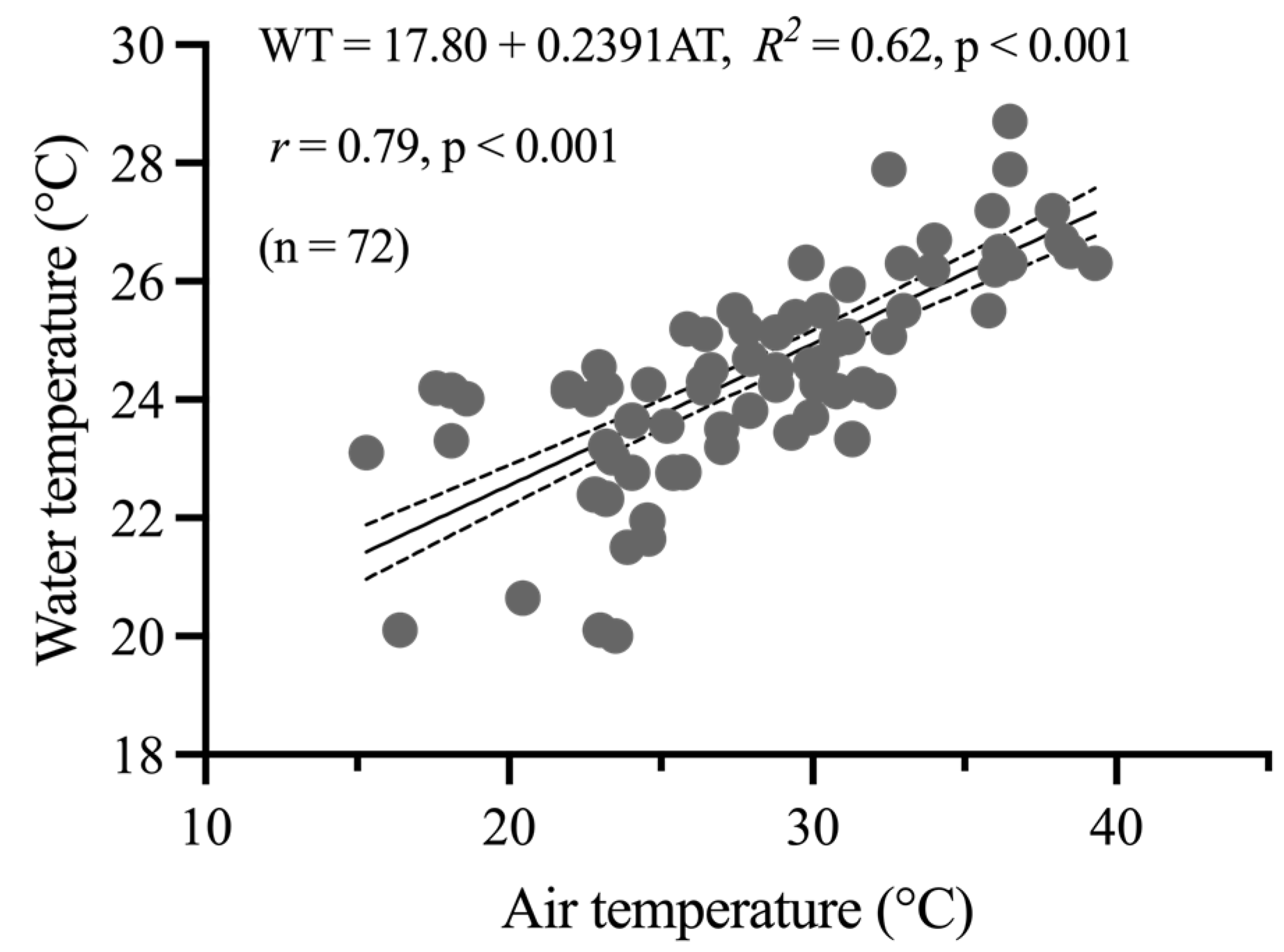
The daily surface WT of Pakkok River on each month of wet and dry season in 2017 and 2022 showed a lower median temperature compared to the daily AT at Thong Pha Phum meteorological Station. As shown in Figure 5, a linear correlation analysis of WT and AT in Pakkok River was performed. The regression indicated correlation between the AT and surface WT with a slope of 0.2391 and the general equation of WT = 17.80 + 0.2391AT.
3.2. CTmax of the Fish in Wet Season and Dry Season
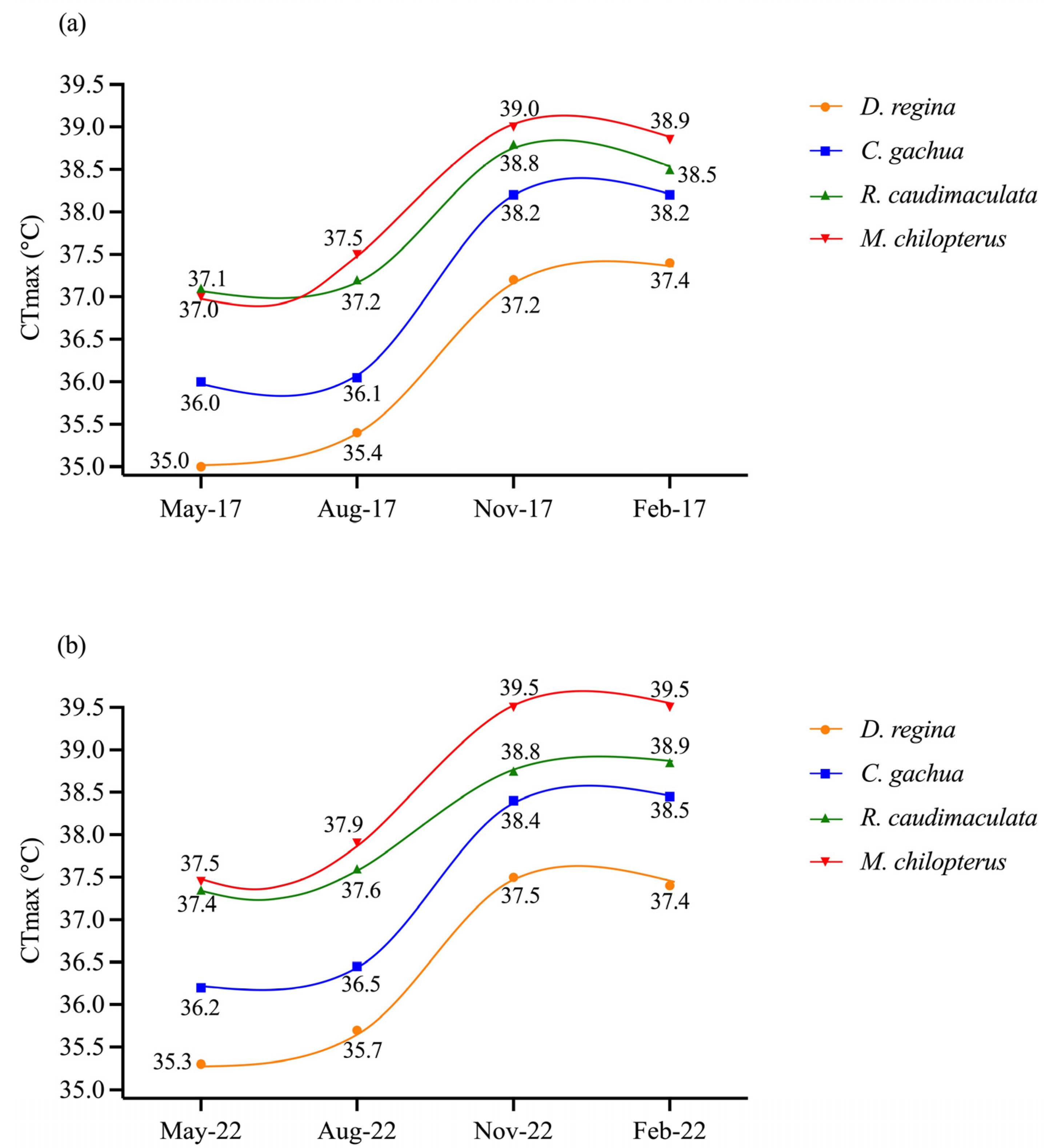
CTmax values of fish in paired replicate tanks did not differ significantly during each month (p ≥ 0.05) and were combined for each of the four species. Body weight of individuals in paired replicate tanks also did not differ significantly for each of the four species (p ≥ 0.05), and were also combined. CTmax was lowest in May with median ± SD for D. regina, C. gachua, R. caudimaculata, and M. chilopterus of 35.0 ± 0.3, 36.0 ± 0.2, 37.1 ± 0.5, and 37.0 ± 0.5 °C, respectively, in 2017, and 35.3 ± 0.3, 36.2 ± 0.5, 37.4 ± 0.4, and 37.5 ± 0.7 °C, respectively, in 2022. Highest CTmax values of each species were 37.2 ± 0.4 to 37.5 ± 0.7 for D. regina, 38.2 ± 0.4 to 38.5 ± 0.6 °C for C. gachua, 38.5 ± 0.7 to 38.9 ± 0.6 °C for R. caudimaculata, and 39.0 ± 0.5 to 39.5 ± 0.7 °C for M. chilopterus, which were observed in dry season during November and February of 2017 and 2022. For each species, CTmax values followed a similar sigmoidal pattern of change between May and February. Between May and August CTmax increased only slightly then abruptly to November and, again, only slightly to February (Figure 6a,b).
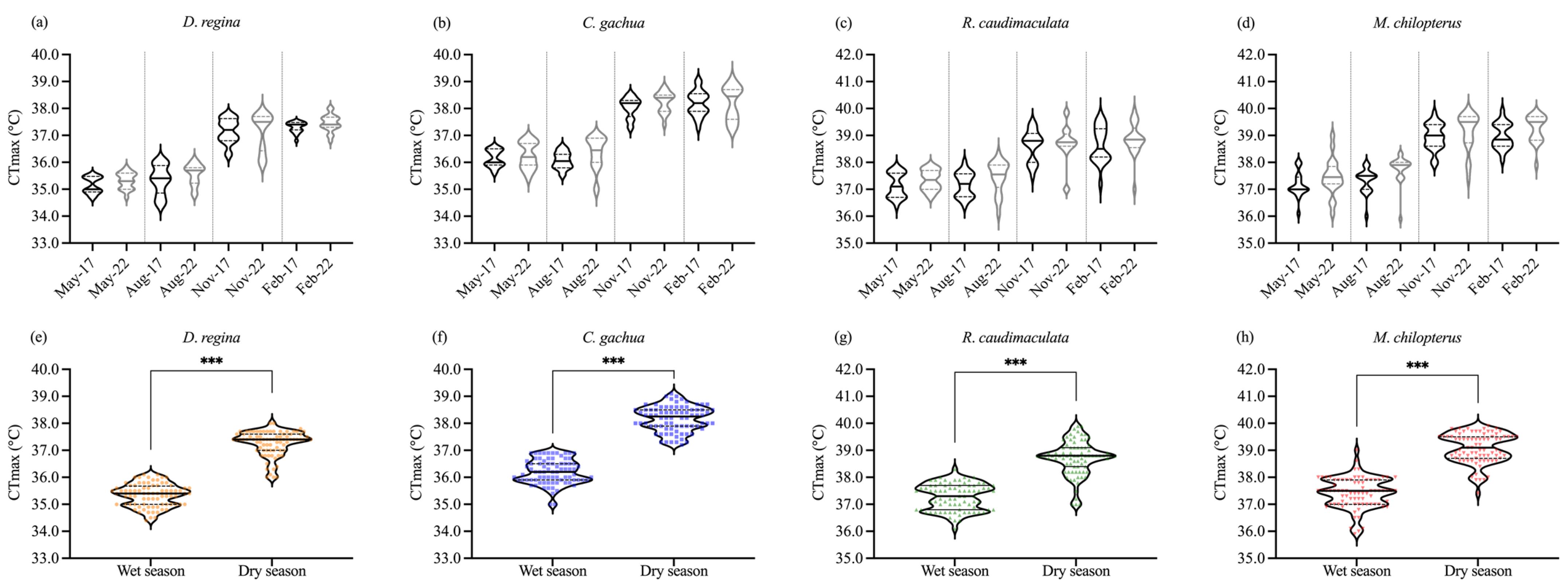
In each fish species, there were no significant difference (p ≥ 0.05) between CTmax value observed from the first study in year 2017 and the second study in year 2022 when considered at the same month of wet and dry season (Figure 7a–d). The increase between May and August was not significant for any species (p ≥ 0.05) as was that between November and February (p ≥ 0.05). However, CTmax values differed (p < 0.05) significantly between months of wet season and months of dry season (Figure 7e–h). Temperature differences of the CTmax between wet and dry season in 2017 and 2022 for D. regina, C. gachua, M. chilopterus, and R. caudimaculata were 2.2 to 2.4, 2.2 to 2.3, 1.5 to 1.8, and 1.9 to 2.0 °C, respectively. CTmax values were inversely related with length of daylight in Kanchanaburi being highest in February and November when daylength was comparatively short, 11.8 and 11.6 h, and lowest in May and August when it was long at 12.8 and 12.7 h.
4. Discussion
The knowledge and data of the thermal tolerance in fishes are very important for predicting the responses to global warming at the population level. The pattern of the relationship between lethal temperature and season was identical for all four species with high and low thermal tolerance values occurring in the dry and wet seasons and inversely with daylengths [30,31,32]. This is in contrast to the pattern in northern or high latitudes which seasonal changes in lethal or stressful temperatures appear to relate directly with photoperiod [12,33]. Our findings demonstrated that the fishes experienced to shorter photoperiod in dry season (~11:13 h) had a higher CTmax than those of exposed to longer photoperiod in wet season (~12:12 h). CTmax values between the wet and dry seasons differed by approximately 2–3 °C for all the fish species in this study. In contrast with other studies, CTmax of various teleost fish in high latitude or temperate regions was higher in the summer season (June to August) when the fishes experience longer daylengths. Brett [8], who measured the thermal tolerance of 13 species of temperate fishes captured from the wild river at Algonquin Park in Ontario, Canada, found that it was highest in early to late spring (March to May) or early summer (June) in according with increasing daylengths. In Tyler’s study [34] the 12 h LC50 for Chrosomus eos and Chromsomus neogaeus was higher in late spring and summer than winter by about 2 °C at each acclimation temperature. Higher lethal temperatures were found also for bluegill sunfish (Lepomis macrochirus), killifish (Fundulus heteroclitus) and blacknose dace (Rhinichthys atratulus) experienced on the longer photoperiod [12,14,16]. This association with photoperiod was in contrast to the pattern found for Thai fishes in the present study. The inconsistency in the pattern of CTmax relating with the photoperiod between tropical fishes in our finding and other temperate fishes in previous findings could be involved with fish species, thermal history, reproductive status, dietary, nutrition, habitat, as well as environmental factors, especially the ambient river temperature [14,16,20,35,36].
Since the early studies by Brett [8] and Tyler [34] a number of thermal tolerance studies have been undertaken with freshwater temperate species and most, if not all, have found a direct association between lethal temperature and photoperiod. Most temperate fishes reproduce only once annually and during the spring and summer suggesting an adaptation to warmer ambient temperatures, higher metabolism, higher water levels and a greater abundance of habitats and food. Thermal tolerance of the four low latitude species in this study was lowest during the wet season when water levels are high and habitat and food abundant, but photoperiod relatively low [30,31]. Importantly, earlier studies have related rainfall with the initiation of reproduction among low latitude fishes [37]. Seasonal variation in photoperiod at low latitudes is much less than that at high latitudes. Furthermore, reproduction strategies among fishes from low latitude regions are more diverse than among temperate species and may exhibit a distinct seasonal pattern of reproduction, an extended period of reproduction or even reproduce more or less continually [38,39,40]. A number of hypotheses have been suggested in relation to the adaptive advantages of different reproductive strategies that may override the importance of photoperiod, especially when seasonal differences are not large as in low latitude regions [38,41]. Spawning at the beginning of the wet season in Thailand coincides with peak food availability in floodplain regions [42], while continuous spawning could represent an opportunistic strategy to take advantage of favorable conditions in a highly competitive environment, while seasonal reproduction may correlate with patterns of egg release [43].
Lethal temperature of the fishes in this study varied inversely with daylength in contrast to the direct association in most seasonal measurements for temperate fishes [44]. Life strategies in fish, to a large degree, represent a balance between the availability and allocation of energy for somatic and gonadal growth [6,37]. Among river fishes, especially those found in fast flowing headwater rivers where currents can be rapid, the energy expended in swimming can be quite significant [45]. Tropical river fishes in Thailand and many temperate river fishes, including the species-rich cyprinids, commonly reproduce during the late spring or early summer [46], which is coincident with high river discharge and currents that tend to be high relative to other seasons. As noted, mature river fishes during their reproductive mode are metabolically stressed under the load of nurturing gametes, particularly oocytes [47]. Furthermore, as reproductive products increase in size, gonads will occupy an increasingly large volume of the body cavity decreasing potential fish appetite and capacity of the digestive tract to contain and process food. Collectively these changes will reduce the energy budget for other activities, including resistance to heat stress. This study supports the suggestion that Thai fish have adopted a reproductive strategy that balances the application of environmental resources that collectively favor reproductive success.
In the present days, the diurnal temperature extremes in the peak of wet and dry seasons at the Pakkok River were ~27 °C and the diurnal temperature fluctuation slightly altered over three years after 2017. In the present study, the dry season-CTmax values were higher than the wet season-CTmax values. CTmax values between the wet and dry seasons differed by approximately 2–3 °C for all fish species. The seasonal variations in thermal tolerance of four fish species captured from Pakkok River and other fishes in previous experiments basically depended on the ambient river temperature and fluctuation. As the previous studies, they found higher CTmax of temperate fishes in high latitudes when the fishes experienced a higher average WT in spring and summer seasons [8,20,34,48]. The ambient WT is directly affected by the ambient AT. The increase in the average of annual AT over Thailand varied between 0.5 and 0.95 °C during a 30-year period until 2022, which is higher than the increased average annual world AT (0.69 °C) [49]. In the period of 2017 to 2022, the highest of mean maximum AT in Thong Pha Phum were 41.4–43.3 °C in March to May, which raised by about 1 to 4 °C from the same interval months of 2017 (Data collected from TDM). In Thailand the average annual AT will likely rise by 0.62 °C per decade [50]. Globally, the average AT may project warming by about 4.5 °C from 1990 to 2100. In Thailand, the average increasing AT of <3.5 °C to >4 °C is projected in next century [51].
The warming AT has implications for freshwater ecosystems caused by the increasing WT. In general, due to the correlation between AT and WT, the current climate change may drive the global average temperature of freshwater system to rise by 0.163 °C per decade [52]. The alteration in WT is also influenced by the change in other environmental factors, which has the potential to rising temperature of the stream. For example, the WT depends on shading level by woody riparian plants, buffer length, width, and canopy cover along the river [53]. The size of the stream and catchment size influence the daily WT fluctuations and rates of heating and cooling [54]. Under a high anthropogenic heat pollutions and high emissions, the riparian landscapes have been altered, thereby giving more impact on the WT in the future. The daily maximum WT especially a small headwater stream in unshaded will increase in the range of 3 °C, while the daily maximum WT in 60% canopy cover will decrease by about 3.5 °C [53,55]. In the period of 2017 to 2022, the highest of mean maximum AT in Thong Pha Phum were 41.4–43.3 °C in March to May, which rose by about 1 to 4 °C from the same interval months of 2017 (Data collected from TDM). Collectively, the extreme temperature in Pakkok River may potentially occur in future summer to early wet season due to global climate change altering the WT, forest degradation, as well as riparian landscape. We employed a regression model to assess how increasing AT influenced the WT in Pakkok River, by the end of this century, the temperature of Pakkok River may increase by 1 °C when the warming AT of >4 °C.
Therefore, the four fish species from Pakkok River in this study may tolerate to the warming temperature of the river in the next century because the increasing WT of ~1 °C above mean WT during 2017 and 2022 is lower than their CTmax across the wet and dry seasons. The wet season-CTmax values for the four species were about 1.5–2.4 °C smaller than those in dry season. This might be explained by the acclimatization of fish to a wider range of daily temperatures in dry season, which can adjust the physiology of response to the environmental changes [56]. The variations in thermal maxima for D. regina, C. gachua, R. caudimaculata, and M. chilopterus, throughout the wet and dry seasons, attribute to their seasonal plasticity in response to the dynamics of thermal stress. The ongoing climate warming is driving extreme weather events, longer warm days and heatwaves in Thailand; the warming habitat has greatly influenced the low plasticity populations in freshwater ecosystem, as well as other ecosystems. According to the results obtained from the experiment, it is found that the R. caudimaculata, and M. chilopterus may have a higher capacity to tolerate acute heat stress caused by extreme thermal events. However, both these species appear to have relatively high thermal maxima and are expected to have a greater risk for the long-term exposure to the warming temperature, indicating the limited ability to acclimatize to warmer conditions because of the higher costs of metabolic and energetic they incur [48,57,58]. The tolerance of the heat stress in fish species relates to a complexity of genetic architecture underlying the thermal tolerance trait [36]. The thermal-tolerant animals appearing a higher baseline expression of thermal response genes; however, they respond to the thermal stress with smaller changes in the expression of those genes [58], which might reveal the association with levels of plasticity [59].
The small shifts in sigmoidal-CTmax pattern in 2022 (second populations), above the sigmoidal-CTmax pattern in 2017 (first population), were observed for all fish species of this study. Interestingly, these may reflect an attempt of adaptive upper thermal limits in the second populations to respond to broader fluctuation of environmental temperature under climate variability and climate change compared with the first populations. Considering the extension in thermal tolerant magnitude of fish, this trait is costly, especially in wet season, in which the energetic costs to exert the expression of thermal response genes (i.e., cluster of heat shock protein genes) might limit other important specific-seasonal traits; for example, the reproductive performance and fecundity [20]. In the worst possible case, we might expect a compromise in reproductive success of fish that would result in decreased population sizes and ongoing increased risk of local extinction in future climate warming. However, our study did not determine the thermal response gene expressions; therefore, these are the most important issues for our future study to predict the vulnerability and phenotypic thermal plasticity in those species acclimatizing to warmer habitat due to climate warming and the detrimental effect on the population sizes and risk to biodiversity.
Author Contributions
Conceptualization, T.S., C.C., W.S., A.W., F.W.H.B. and S.T.; methodology, T.S., F.W.H.B. and S.T.; validation, F.W.H.B. and S.T.; formal analysis, T.S. and S.T.; investigation, T.S. and S.T.; resources, T.S., C.C., W.S., F.W.H.B. and S.T.; writing—original draft preparation, T.S. and S.T.; writing—review and editing, T.S., C.C., W.S., A.W., F.W.H.B. and S.T; visualization, T.S.; supervision, F.W.H.B. and S.T.; project administration, C.C., W.S. and S.T.; funding acquisition, T.S., C.C., W.S., F.W.H.B. and S.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded in part by the National Research Council of Thailand, NRCT (Grant No. N25A650460), Thailand Research Fund through the Royal Golden Jubilee Ph.D. Program (Grant No. PHD0330/2552), and the U.S. National Science Foundation through a Rules of Life award (grant No. NSF 1839915) for the project of Fish of Maeklong Basin, Thailand.
Data Availability Statement
http://www.aws-observation.tmd.go.th/web/climate/climate_report_annual.asp (accessed on 1 December 2022).
Acknowledgments
The fish collecting permit was authorized by the Department of Fisheries, Ministry of Agriculture and Cooperatives, Thailand. We thank the National Research Council of Thailand for the international collaborative research activity for the project of Fish of Maeklong Basin, Thailand permission (NRCT#11880). We appreciate the facilities support by Department of Conservation Biology and Central Instruments in Kanchanaburi Campus, Mahidol University authorized by Thatchavee Leelawat, the Vice President for Kanchanaburi Campus. We are also grateful to the faculty of Science, Burapha University, Chonburi, Thailand for providing the facilities.
Conflicts of Interest
The authors declare no conflict of interest.
References
- de Viron, O.; Dehant, V.; Goosse, H.; Crucifix, M. Effect of global warming on the length-of-day. Geophys. Res. Lett. 2002, 29, 1146. [Google Scholar] [CrossRef]
- Landerer, F.W.; Jungclaus, J.H.; Marotzke, J. Ocean bottom pressure changes lead to a decreasing length-of-day in a warming climate. Geophys. Res. Lett. 2007, 34, L06307. [Google Scholar] [CrossRef] [Green Version]
- Scott, S.G.; Pankhurst, N.W. Interannual variation in the reproductive cycle of the New Zealand snapper Pagrus auratus (Bloch & Schneider) (Sparidae). J. Fish Biol. 1992, 41, 685–696. [Google Scholar]
- Shimizu, A. Effect of photoperiod and temperature on gonadal activity and plasma steroid levels in a reared strain of the mummichog (Fundulus heteroclitus) during different phases of its annual reproductive cycle. Gen. Comp. Endocrinol. 2003, 131, 310–324. [Google Scholar] [CrossRef]
- Moyle, P.B.; Cech, J.J. Fishes: An Introduction to Ichthyology, 4th ed.; Prentice Hall: Uppersaddle River, NJ, USA, 2000; p. 621. [Google Scholar]
- Pankhurst, N.W.; Munday, P.L. Effects of climate change on fish reproduction and early life history stages. Mar. Freshw. Res. 2011, 62, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Beitinger, T.L.; Bennett, W.A.; McCauley, R.W. Temperature tolerance of North American freshwater fishes exposed to dynamic changes in temperature. Environ. Biol. Fishes 2000, 58, 237–275. [Google Scholar] [CrossRef]
- Brett, J.R. Some lethal Temperature Relations of Algonquin Park Fishes; University of Toronto studied biological series, No.52. Publications of the Ontario fisheries research laboratory, No.63; University of Toronto Press: Toronto, ON, Canada, 1944; pp. 1–49. [Google Scholar]
- Golovanov, V.K.; Ruchin, A.B. Critical thermal maximum of Amur Sleeper Perccottus glenii in different season of the year. J. Ichthyol. 2011, 51, 788–793. [Google Scholar] [CrossRef]
- Alexandre, C.M.; Ferreira, M.T.; Almeida, P.R. Life history of a cyprinid species in non-regulated and regulated rivers from permanent and temporary Mediterranean basins. Ecohydrology 2015, 8, 1137–1153. [Google Scholar] [CrossRef]
- Cereja, R. Critical thermal maxima in aquatic ectotherms. Ecol. Indic. 2020, 119, 106856. [Google Scholar] [CrossRef]
- Terpin, K.M.; Spotila, J.R.; Koons, R.R. Effect of photoperiod on the temperature tolerance of the blacknose dace. Rhinichthys atratulus. Comp. Biochem. Physiol. 1976, 53A, 241–244. [Google Scholar] [CrossRef]
- Guderley, H.; Leroy, P.H.; Gagné, A. Thermal acclimation, growth, and burst swimming of threespine stickleback: Enzymatic correlates and influence of photoperiod. Physiol. Biochem. Zool. 2001, 74, 66–74. [Google Scholar] [CrossRef]
- Healy, T.M.; Schulte, P.M. Factors affecting plasticity in whole-organism thermal tolerance in common killifish (Fundulus heteroclitus). J. Comp. Physiol.B 2012, 182, 49–62. [Google Scholar] [CrossRef]
- Huey, R.B.; Buckley, L.B. Designing a seasonal acclimation study presents challenges and opportunities. Integr. Org. Biol. 2022, 4, obac016. [Google Scholar] [CrossRef] [PubMed]
- Lamptey, D.I.; Sparks, R.W.; De Oca, R.M.; Skolik, R.; Menze, M.A.; Martinez, E. Seasonal changes in mitochondrial bioenergetics and physiological performance of the bluegill sunfish, Lepomis macrochirus, from a shallow, Midwest river. J. Therm. Biol. 2022, 104, 103186. [Google Scholar] [CrossRef]
- Kitano, H. Systems biology: A brief overview. Sciences 2002, 295, 1662–1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wood, C.M.; McDonald, D.G. Global Warming: Implications for Freshwater and Marine Fish; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Payne, N.L.; Smith, J.A.; van der Meulen, D.E.; Taylor, M.D.; Watanabe, Y.Y.; Takahashi, A.; Marzullo, T.A.; Gray, C.A.; Cadiou, G.; Suthers, I.M. Temperature dependence of fish performance in the wild: Links with species biogeography and physiological thermal tolerance. Funct. Ecol. 2016, 30, 903–912. [Google Scholar] [CrossRef] [Green Version]
- Turko, A.J.; Nolan, C.B.; Balshine, S.; Scott, G.R.; Pitcher, T.E. Thermal tolerance depends on season, age and body condition in imperilled redside dace Clinostomus elongatus. Conserv. Physiol. 2020, 8, coaa062. [Google Scholar] [CrossRef]
- Li, D.; Dorber, M.; Barbarossa, V.; Verones, F. Global characterization factors for quantifying the impacts of increasing water temperature on freshwater fish. Ecol. Indic. 2022, 142, 109201. [Google Scholar] [CrossRef]
- Becker, C.D.; Genoway, R.G. Evaluation of the critical thermal maximum for determining thermal tolerance of freshwater fish. Environ. Biol. Fishes 1979, 4, 245–256. [Google Scholar] [CrossRef]
- Calosi, P.; Bilton, D.T.; Spicer, J.I. Thermal tolerance, acclimatory capacity and vulnerability to global climate change. Biol. Lett. 2008, 4, 99–102. [Google Scholar] [CrossRef] [Green Version]
- Tongnunui, S.; Beamish, F.W.H. Critical thermal maximum, temperature acclimation and climate effects on Thai freshwater fishes. EA. 2017, 10, 109–117. [Google Scholar]
- Beamish, F.W.H.; Sa-ardrit, P.; Tongnunui, S. Habitat characteristics of the Cyprinidae in small rivers in Central Thailand. Environ. Biol. Fishes 2006, 76, 237–253. [Google Scholar] [CrossRef]
- Hayden, B.; Tongnunui, S.; Beamish, F.W.H.; Nithirojpakdee, P.; Cunjak, R.A. Variation in stable-isotope ratios between fin and muscle tissues can alter assessment of resource use in tropical river fishes. J. Fish Biol. 2017, 91, 574–586. [Google Scholar] [CrossRef]
- Hayden, B.; Tongnunui, S.; Beamish, F.W.H.; Nithirojpakdee, P.; Soto, D.X.; Cunjak, R.A. Functional and trophic diversity of tropical headwater stream communities inferred from carbon, nitrogen and hydrogen stable isotope ratios. Food Webs 2021, 26, e00181. [Google Scholar] [CrossRef]
- Tongnunui, S.; Sooksawat, T.; Kohkaew, R.; Teampanpong, J.; Wattanakornsiri, A. Accumulation of microplastics in the freshwater shrimp, Macrobrachium lanchesteri, from Khwae Noi Watershed in Western Thailand. EnvironmentAsia 2022, 15, 25–37. [Google Scholar]
- Erdfelder, E.; Faul, F.; Buchner, A. GPOWER: A general power analysis program. BRMIC 1996, 28, 1–11. [Google Scholar] [CrossRef]
- Hoar, W.S. Seasonal variation in the resistance of goldfish to temperature. Trans. R. Soc. Can. 1955, 49, 25–34. [Google Scholar]
- Hoar, W.S. Photoperiodism and thermal resistance of goldfish. Nature 1956, 178, 364–365. [Google Scholar] [CrossRef]
- Hoar, W.S.; Robertson, G.B. Temperature resistance of goldfish maintained under controlled photoperiods. Can. J. Zool. 1959, 37, 419–428. [Google Scholar] [CrossRef]
- Sharma, N.K.; Akhtar, M.S.; Pandey, N.; Singh, R.; Singh, A.K. Seasonal variation in thermal tolerance, oxygen consumption, antioxidative enzymes and non-specific immune indices of Indian hill trout, Barilius bendelisis (Hamilton, 1870) from central Himalaya, India. J. Therm. Biol. 2015, 52, 166–176. [Google Scholar] [CrossRef]
- Hines, C.W.; Fang, Y.; Chan, V.K.; Stiller, K.T.; Brauner, C.J.; Richards, J.G. The effect of salinity and photoperiod on thermal tolerance of Atlantic and coho salmon reared from smolt to adult in recirculating aquaculture systems. Comp. Biochem. Physiol., Mol. Amp; Integr. Physiol. 2019, 230, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Tyler, A.V. Some lethal temperature relations of two minnows of the genus Chrosomus. Can. J. Zool. 1966, 44, 349–364. [Google Scholar] [CrossRef]
- Morgan, R.; Sundin, J.; Finnøen, M.H.; Dresler, G.; Vendrell, M.M.; Dey, A.; Sarkar, K.; Jutfelt, F. Are model organisms representative for climate change research? Testing thermal tolerance in wild and laboratory zebrafish populations. Conserv. Physiol. 2019, 7, coz036. [Google Scholar] [CrossRef]
- Haworth, M.R.; Bestgen, K.R. Flow and water temperature affect reproduction and recruitment of a Great Plains cyprinid. Can. J. Fish. Aquat. Sci. 2017, 74, 853–863. [Google Scholar] [CrossRef]
- Lowe-McConnell, R.H. Ecological Studies in Tropical Fish Communities; Cambridge University Press: Cambridge, UK, 1987; p. 382. [Google Scholar]
- Martin-Smith, K.; Laird, L. Reproductive patterns in some Cypriniformes from Borneo. In Proceedings of the 5th Indo Pacific Fish Conference, Paris, Society of French Ichthyology, Noumea, French, 3–8 November 1997. [Google Scholar]
- Muchilisin, Z.; Musman, M.; Azizah, M.S. Spawning season of Rasbora tawarensis (Pisves: Cyprinidae) in Lake Laut Tawar, Aceh Province, Indonesia. Reprod. Biol. Endocrinol. 2010, 8, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winemiller, K.O. Patterns of life history among South American fishes in seasonal environments. Oecologia 1989, 81, 225–241. [Google Scholar] [CrossRef]
- Thapanand-Chaidee, T. Fecundity relationship, maturity size and spawning season of shark catfish Helicophagus waandersii Bleeker, 1858 in the Mun River, Thailand. J. Fish. Environ. 2008, 32, 17–29. [Google Scholar]
- Valbo-Jørgensen, J.; Poulsen, A.F. Using local knowledge as a research tool in the study of river fish biology: Experiences from the Mekong. Environ. Dev. Sustain. 2000, 2, 253–376. [Google Scholar] [CrossRef]
- Petitgas, P.; Rijnsdorp, A.D.; Dickey-Collas, M.; Engelhard, G.H.; Peck, M.A.; Pinnegar, J.K.; Drinkwater, K.; Huret, M.; Nash, R.D. Impacts of climate change on the complex life cycles of fish. Fish. Oceanogr. 2013, 22, 121–139. [Google Scholar] [CrossRef]
- Beamish, F.W.H. Migration and spawning energetics of the anadromous sea lamprey, Petromyzon marinus. Environ. Biol. Fish 1979, 4, 3–7. [Google Scholar]
- Boulton, A.J.; Boyero, L.; Covich, A.P.; Dobson, M.; Lake, S.; Pearson, R. Are tropical streams ecologically different from temperate streams? In Tropical Stream Ecology; Dudgeon, D., Ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 257–284. [Google Scholar]
- Hontela, A.; Stacey, N. Cyprinidae. In Reproductive Seasonality in Teleosts; Munro, A.D., Scott, A.P., Lam, T.J., Eds.; CRC Press: Boca Raton, FL, USA, 2019; pp. 53–78. [Google Scholar]
- Shultz, A.D.; Zuckerman, Z.C.; Suski, C.D. Thermal tolerance of nearshore fishes across seasons: Implications for coastal fish communities in a changing climate. Mar. Biol. 2016, 163, 1–10. [Google Scholar] [CrossRef]
- Suwanpakdee, S.; Sriyasak, P.; Pimolrat, P. Risk of climate variability on tilapia cage culture in songkhram river in northeastern Thailand. AJSTR 2021, 24, 76–83. [Google Scholar] [CrossRef]
- Phanprasit, W.; Rittaprom, K.; Dokkem, S.; Meeyai, A.C.; Boonyayothin, V.; Jaakkola, J.J.; Näyhä, S. Climate warming and occupational heat and hot environment standards in Thailand. Saf. Health Work 2021, 12, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Kiguchi, M.; Takata, K.; Hanasaki, N.; Archevarahuprok, B.; Champathong, A.; Ikoma, E.; Jaikaeo, C.; Kaewrueng, S.; Kanae, S.; Kazama, S.; et al. A review of climate-change impact and adaptation studies for the water sector in Thailand. Environ. Res. Lett. 2021, 16, 023004. [Google Scholar] [CrossRef]
- Liu, S.; Xie, Z.; Liu, B.; Wang, Y.; Gao, J.; Zeng, Y.; Xie, J.; Xie, Z.; Jia, B.; Qin, P.; et al. Global river water warming due to climate change and anthropogenic heat emission. Glob. Planet. Chang. 2020, 193, 103289. [Google Scholar] [CrossRef]
- Kail, J.; Palt, M.; Lorenz, A.; Hering, D. Woody buffer effects on water temperature: The role of spatial configuration and daily temperature fluctuations. Hydrol. Process. 2021, 35, e14008. [Google Scholar] [CrossRef]
- Coats, W.A.; Jackson, C.R. Riparian canopy openings on mountain streams: Landscape controls upon temperature increases within openings and cooling downstream. Hydrol. Process. 2020, 34, 1966–1980. [Google Scholar] [CrossRef]
- Loicq, P.; Moatar, F.; Jullian, Y.; Dugdale, S.J.; Hannah, D.M. Improving representation of riparian vegetation shading in a regional stream temperature model using LiDAR data. Sci. Total Environ. 2018, 624, 480–490. [Google Scholar] [CrossRef]
- Hopkin, R.S.; Qari, S.; Bowler, K.; Hyde, D.; Cuculescu, M. Seasonal thermal tolerance in marine Crustacea. J. Exp. Mar. Biol. Ecol. 2006, 331, 74–81. [Google Scholar] [CrossRef]
- Magozzi, S.; Calosi, P. Integrating metabolic performance, thermal tolerance, and plasticity enables for more accurate predictions on species vulnerability to acute and chronic effects of global warming. Glob. Chang. Biol. 2015, 21, 181–194. [Google Scholar] [CrossRef]
- Barley, J.M.; Cheng, B.S.; Sasaki, M.; Gignoux-Wolfsohn, S.; Hays, C.G.; Putnam, A.B.; Sheth, S.; Villeneuve, A.R.; Kelly, M. Limited plasticity in thermally tolerant ectotherm populations: Evidence for a trade-off. Proc. R. Soc. B Biol. Sci. B 2021, 288, 20210765. [Google Scholar] [CrossRef] [PubMed]
- Rivera, H.E.; Aichelman, H.E.; Fifer, J.E.; Kriefall, N.G.; Wuitchik, D.M.; Wuitchik, S.J.; Davies, S.W. A framework for understanding gene expression plasticity and its influence on stress tolerance. Mol. Ecol. 2021, 30, 1381–1397. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Map of the study area (a), indicating the sampling location (blue dot) in Pakkok River, Kwai Noi River system, Mae Khlong basin, Thong Pha Phum, Kanchanaburi province, Thailand. The habitat of the Pakkok River observed from the same vantage point during the wet season of 2017 (b) and the dry season of 2017 (c).
Figure 1.
Map of the study area (a), indicating the sampling location (blue dot) in Pakkok River, Kwai Noi River system, Mae Khlong basin, Thong Pha Phum, Kanchanaburi province, Thailand. The habitat of the Pakkok River observed from the same vantage point during the wet season of 2017 (b) and the dry season of 2017 (c).

Figure 2.
The monthly averaged time series of AT (a), the comparison of monthly averaged AT (b), and the violin plot of averaged annual AT (c) over Thailand during 2015–2022 and 30-year period. For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. AT; air temperature.
Figure 2.
The monthly averaged time series of AT (a), the comparison of monthly averaged AT (b), and the violin plot of averaged annual AT (c) over Thailand during 2015–2022 and 30-year period. For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. AT; air temperature.

Figure 3.
The medians of maximum (a), minimum (b), average (c), and diurnal range (d) of AT in Thong Pha Phum station during the study periods of dry season (November and February) and wet season (May and August) in 2017 and 2022. For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. *, **, and *** indicate significance at p < 0.05, p < 0.01, and p < 0.001, respectively, as determined by independent samples t-test.
Figure 3.
The medians of maximum (a), minimum (b), average (c), and diurnal range (d) of AT in Thong Pha Phum station during the study periods of dry season (November and February) and wet season (May and August) in 2017 and 2022. For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. *, **, and *** indicate significance at p < 0.05, p < 0.01, and p < 0.001, respectively, as determined by independent samples t-test.

Figure 4.
The diurnal variations of WT at Pakkok River during the study periods of wet season; May (a), August (b) and dry season; November (c), February (d) in 2017 and 2022. The average WT in Pakkok River was monitored on the 15th and 28th of each observed month (May, August, November, and February) during 2017 and 2022. The data presented at the predetermined time intervals of 3.00, 6.00, 9.00, 12.00, 15.00, 18.00, 21.00, and 24.00 h were the average values of the 15th (n = 3) and 28th (n = 3) of the month. Dash lines in black and light grey represent the daily median WT on each month of year 2017 and 2022, respectively.
Figure 4.
The diurnal variations of WT at Pakkok River during the study periods of wet season; May (a), August (b) and dry season; November (c), February (d) in 2017 and 2022. The average WT in Pakkok River was monitored on the 15th and 28th of each observed month (May, August, November, and February) during 2017 and 2022. The data presented at the predetermined time intervals of 3.00, 6.00, 9.00, 12.00, 15.00, 18.00, 21.00, and 24.00 h were the average values of the 15th (n = 3) and 28th (n = 3) of the month. Dash lines in black and light grey represent the daily median WT on each month of year 2017 and 2022, respectively.

Figure 5.
Relationship between daily air temperature (AT) and surface layer water temperature (WT) at Pakkok River during wet and dry season of 2022. The AT and WT water temperature in Pakkok River was monitored at 15th and 28th of each observed month (May, August, November, and February) during 2022 (n = 72).
Figure 5.
Relationship between daily air temperature (AT) and surface layer water temperature (WT) at Pakkok River during wet and dry season of 2022. The AT and WT water temperature in Pakkok River was monitored at 15th and 28th of each observed month (May, August, November, and February) during 2022 (n = 72).

Figure 6.
Seasonal changes of thermal tolerance or critical thermal maximum (CTmax) for four species acclimated to 25 °C and measured in mid-May (early wet season), mid-August (late wet season), mid-November (early dry season) and mid-February (late dry season) during 2017 (a) and 2020 (b). Values are median for 20 fish.
Figure 6.
Seasonal changes of thermal tolerance or critical thermal maximum (CTmax) for four species acclimated to 25 °C and measured in mid-May (early wet season), mid-August (late wet season), mid-November (early dry season) and mid-February (late dry season) during 2017 (a) and 2020 (b). Values are median for 20 fish.

Figure 7.
Critical thermal maximum (CTmax) values of four species (a–d) captured from the wild in May, August, November, and February are compared between 2017 (black) and 2022 (light grey). For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. Seasonal median CTmax for wet- and dry-captured four fish species (e–h). For each box plot, the dots show the outlier data, the bold horizontal line in the center of the box represents the median, the top and bottom of the box represent the first and third quartiles, and the whiskers indicate the highest and lowest values within 1.5× the interquartile range. *** indicate significance at p < 0.001, respectively, as determined by independent samples t-test.
Figure 7.
Critical thermal maximum (CTmax) values of four species (a–d) captured from the wild in May, August, November, and February are compared between 2017 (black) and 2022 (light grey). For each violin plot, the bold horizontal line in the center of the box represents the median, and the dash lines at the bottom and top of the box represent the first and third quartiles, respectively. Seasonal median CTmax for wet- and dry-captured four fish species (e–h). For each box plot, the dots show the outlier data, the bold horizontal line in the center of the box represents the median, the top and bottom of the box represent the first and third quartiles, and the whiskers indicate the highest and lowest values within 1.5× the interquartile range. *** indicate significance at p < 0.001, respectively, as determined by independent samples t-test.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Physiochemical parameters of Pakkok River, Kwai Noi River system, Mae Khlong basin, Kanchanaburi province, Thailand during the wet and dry seasons of 2017 and 2022.
Table 1.
Physiochemical parameters of Pakkok River, Kwai Noi River system, Mae Khlong basin, Kanchanaburi province, Thailand during the wet and dry seasons of 2017 and 2022.
| Physicochemical Parameters | Wet Season (May and August) | Dry Season (November and February) | ||
|---|---|---|---|---|
| Year 2017 | Year 2022 | Year 2017 | Year 2022 | |
| Dissolved oxygen (DO, mgO2/L) | 8.1–8.5 | 8.7–9.1 | 8.7–9.1 | 8.6–9.1 |
| 8.3 ± 0.1 | 8.9 ± 0.1 | 8.8 ± 0.1 | 8.8 ± 2.0 | |
| Alkalinity (mgCaCO3/L) | 34–36 | 34–38 | 34–36 | 32–36 |
| 34.7 ± 0.8 | 35.7–1.5 | 35.0 ± 0.7 | 34.1 ± 1.6 | |
| Silica (mgSiO2/L) | 34–35 | 34–37 | 35–38 | 35–38 |
| 37.7 ± 0.4 | 35.5 ± 1.2 | 36.3 ± 1.3 | 36.5 ± 1.2 | |
| Ammonia (mgNH3N/L) | 0.01–0.02 | 0.01–0.02 | 0.01–0.02 | 0.01–0.2 |
| 0.012 ± 0.00 | 0.01 ± 0.00 | 0.01 ± 0.00 | 0.04 ± 0.1 | |
| Total iron (mgFe/L) | 0.01–0.02 | 0.01–0.2 | 0.01–0.02 | 0.01–0.02 |
| 0.014 ± 0.00 | 0.02 ± 0.05 | 0.01 ± 0.00 | 0.01 ± 0.00 | |
| pH | 7.4–7.8 | 6.5–7.8 | 7.4–8.1 | 7.4–8.2 |
| 7.5 ± 0.1 | 7.4 ± 0.4 | 7.7 ± 0.3 | 7.8 ± 0.3 | |
| Canopy (%) | 45% | 40% | 30% | 30% |
| 45.0 ± 0.0 | 40.0 ± 0.0 | 30.0 ± 0.0 | 30.0 ± 0.0 | |
Note(s): The range (upper row for each parameter) and average (lower row for each parameter) value of the physicochemical parameters in Pakkok River was monitored on the 15th and 28th of each observed month (May, August, November, and February) during 2017 and 2022.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Tongnunui, S.; Sooksawat, T.; Chotwiwatthanakun, C.; Supiwong, W.; Wattanakornsiri, A.; Beamish, F.W.H. Seasonal Changes in Upper Thermal Tolerances of Freshwater Thai Fishes. Water 2023, 15, 350. https://doi.org/10.3390/w15020350
AMA Style
Tongnunui S, Sooksawat T, Chotwiwatthanakun C, Supiwong W, Wattanakornsiri A, Beamish FWH. Seasonal Changes in Upper Thermal Tolerances of Freshwater Thai Fishes. Water. 2023; 15(2):350. https://doi.org/10.3390/w15020350
Chicago/Turabian StyleTongnunui, Sampan, Treerat Sooksawat, Charoonroj Chotwiwatthanakun, Weerayuth Supiwong, Amnuay Wattanakornsiri, and F. W. H. Beamish. 2023. "Seasonal Changes in Upper Thermal Tolerances of Freshwater Thai Fishes" Water 15, no. 2: 350. https://doi.org/10.3390/w15020350
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.