

Floodplain Forest Restoration as a Nature-Based Solution to Create Climate-Resilient Communities in European Lowland Estuaries
1
NIOZ Royal Netherlands Institute for Sea Research, Department of Estuarine and Delta Systems, Utrecht University, P.O. Box 140, 4400 AC Yerseke, The Netherlands
2
Science Department, University College Roosevelt, P.O. Box 94, 4330 AB Middelburg, The Netherlands
3
Geographical Institute, University of Bonn, 53115 Bonn, Germany
*
Author to whom correspondence should be addressed.
Water 2023, 15(3), 440; https://doi.org/10.3390/w15030440
Submission received: 22 December 2022
/
Revised: 13 January 2023
/
Accepted: 17 January 2023
/
Published: 22 January 2023
(This article belongs to the Special Issue Natural Flood Management as an Adaptive Pathway to Climate-Resilient Communities)
Abstract
:Anthropogenic impact has largely altered the estuarine environment. Today, Nature-based solutions (NbS) are required to respond to challenges such as flooding that affect both the human population and estuarine ecosystems. Estuarine ecosystems such as saltmarshes and floodplain forests provide valuable services, and wetland restoration for flood control is suggested. Recently, flow velocities attenuating the effects of juvenile floodplain willows and wave attenuation through White willow trees under extreme conditions have been found. Services in flood control seemed promising. However, large woods do not bend and may cause turbulence. On the contrary, complex and flexible woody vegetation bends in response to currents and reduces velocities, thereby inducing lower turbulence, but knowledge of the diversity and structure of tidal willow floodplain forests (TWFF) is lacking. Thus, first, an observational study on the occurrence of TWFF in European estuaries along the North Sea was carried out. Based on findings in aerial images, only small TWFF fragments were confirmed in the field for the Elbe, Ems, Dutch delta, and the Scheldt estuary. Based on these findings and the analyses of management plans, the restoration of TWFF was planned, implemented, and studied, including a long-term study at the Elbe estuary upstream stretch. The results of the restoration efforts are provided, including the long-term study (2012–2022) on a restored TWFF in the Elbe estuary, which could serve as a pilot for TWFF restoration and adapting estuarine communities to climate change via NbS.
1. Introduction
Resilience to disturbances that result from different colonisation behaviors of diverse biota leads to landscape self-organisation across different biogeomorphic systems [1]. The floodplain forest ecosystem function of woody vegetation drives (i) the construction of resilient floodplains and (ii) the function of stabilised floodplain forests to act as ‘diversity reservoirs’ which can support regeneration after destructive floods [2]. The plant species’ life history traits thereby influence landscape morphodynamics [3]. Hydrodynamic studies demonstrated that plant species traits interact with environmental conditions resulting in scale-dependent feedback, which explains why the effects of vegetation, e.g., on salt marsh landscape formation, are species-specific [4], e. g., salt marsh plant species flexibility and height determined different wave dissipation potential [5]. This is of importance for estuaries as hotspots for anthropogenic impacts where changes have dramatically accelerated over the past 150 to 300 years with the destruction of more than half of the worldwide tidal wetland habitats and depleted species diversity [6].
Lowland floodplain forests in the temperate climate of the northern hemisphere are composed of members of the Salicaceae plant family, poplars of the genus Populus, and willows of the genus Salix [7] but only Salix species have settled up to the estuarine mouths at the North Sea [8]. Floodplain forests in Eurasia are characterised by White willow (Salix alba L.) as a keystone species [9]. However, more than 99% of European Salix floodplain forests are lost [10]. Conserving and restoring floodplain forests may promote climate change mitigation and adaptation through nature-based solutions (NbS) in flood defense [11]. Coastal flooding is one of the key risks under a medium to low adaptation to climate change, and NbS similar to floodplain and watershed restoration together with a social policy is suggested to mediate some of the limitations of grey infrastructure as a strategy of climate resilience in cities and settlements [12]. Such NbS links natural flood management (NFM) and sustainable urban drainage systems (SuDS) as sustainable catchment-wide flood management combining rural and urban approaches [13]. Wetland vegetation decreases surges and attenuates waves by simultaneously maintaining shallow water and absorbing flood waters which leads to the saving of about 5000 human lives/per year globally [14]. Thus, wetland restoration is strongly suggested for ecosystem-based coastal defense [15]. Estuarine biota is adapted to the tidal systems’ inherent variability in hydrodynamics and salinity and confers resilience to the effects of environmental changes, including climate change [16]. Mangroves serve for flood coastal control in the tropics [17], whereas the floodplain Salix species to protect banks from soil erosion and for streamside restoration by establishing riparian buffers has long since become popular [18]. Floodplain trees increase the surface roughness, which slows down flood flows and, in parallel, enhances flood storage which can lead to overbank flow in rivers, thus, floodplain forests should be placed strategically to alleviate flooding [19]. A multi-criteria evaluation, including topography, physical conditions, land use, ownership, and conservation status to define appropriate sites, is required for forest initialisation and management in floodplains [20]. Therefore, the question of estuarine site selection and Salix species selection for the tidal floodplain forest ecosystem function, structure, and services in estuarine restoration and flood protection in the temperate climate is of high importance and has to be answered.
Plant traits of the floodplain Salix specimen have been valuable for civilisations for millennia: flexible branches were used as woven baskets, wickerwork for house construction, and fences for shelter, which makes them one of the first articles manufactured by human floodplain communities worldwide [9]. More recently, Salix was found to be able to absorb both organic and inorganic pollutants making it an efficient tool for bioremediation [21]. Today, the value of Salix is well documented, and numerous varieties have been collected in National Willow Collections for diverse utilisation purposes [22].
In floodplain forests, diverse Salix species are able to colonise bare sediments rapidly with abundant seeds or vegetative propagules (branch fragments). The established floodplain Salix specimen has exceptional mechanical properties, such as a high bending capacity and breaking resistance enabling resilience to river currents and waves [7]. A high bending capacity was recently confirmed for juvenile tidal floodplain Salix that was grown from vegetative propagules in a mesocosm experiment using tidal flooding and salinity treatment corresponding to estuarine conditions [23]. Floodplain forest S. alba trees reduced waves even at heights up to 2.5 m under extreme conditions in a real-scale study in the large Delta flume, The Netherlands (NL). The wave-attenuation capacity under extreme conditions suggests that floodplain willow forests can play a significant role in flood risk reduction services [24]. This confirms earlier suggestions for integrating S. alba into flood protection foreshore designs along the dike line [25]; this would also prove significant for estuarine floodplain forest restoration due to flow velocities attenuating the effects of juvenile Salix plants (Figure 1a) that were tested alive in a small-scale flume study [26]. In this flume study, flow velocities were reduced in both the leafy status and the leafless stage. The increased canopy permeability but maintained shoot flexibility could have led to a similar flow reduction in dormancy (leafless stage) during the storm surge season in winter. Moreover, flow velocity was reduced most directly behind the Salix specimen in this flume study. In the field, Salix alba specimens were planted on banks along the dike line in a river-dominated tidal freshwater system for flood protection purposes (Figure 1b): modelling results on this plantation scheme indicated wave height reduction by 60–80%, suggesting the beneficial application of S. alba floodplain forests on the macro scale, e.g., for the reduction in waves overtopping the dikes [25]. However, the findings of both van Wesenbeeck et al. [24] and Borsje et al. [25] are based on studies of planted S. alba stumps with equal structure.
The biological diversity of Salix is characteristic in riverine floodplains (e.g., [7]) and in the tidal freshwater wetlands of Elbe and the Scheldt estuaries above the mean high-water line [27]. Salix alba and S. viminalis even occur up to the mesohaline estuarine stretch [28], but both Salix species settle along estuaries in the temperate climate high in the intertidal where they can tolerate semidiurnal flooding at least up to 60 cm [23]. Salix alba floodplain forest (Salicion albae) is a protected habitat type according to the European Habitats Directive (HD) [29]. Contrary to NFM techniques which mainly focus from the source to the upstream catchment, designs implemented in the estuarine and lower catchment context are more driven by the need to protect habitats according to the HD with supplemental benefits for flood defence [13]. In Europe, many estuaries are designated as Natura 2000 sites as the legally binding system of protected areas in EU member states. Within the Natura 2000 network, both habitat types (HD annex I) and species (HD annex II) are listed, for which the conservation and restoration of good conservation status are required. This includes habitat type 91E0*—Salix alba floodplain forest [30]. In the European Forest Gap Analyses [10], the Netherlands, Belgium and Germany were found to have a potential forest cover of more than 95% but ranked among the countries with the highest forest loss (NL > 90%; Belgium (B) > 80%; GER > 70%). Generally, alluvial forests suffered from the largest proportional loss (>90%), including willow floodplain forests in particular (>99.5%).
Lowland estuaries in The Netherlands, Belgium, and Germany were selected to investigate the occurrence, diversity, structure, and restoration of tidal willow floodplain forests (hereinafter named TWFF) with regard to ecosystem services, particularly in flood defence. Second, a long-term study lasting ten years from 2012 to 2022 on TWFF restoration, maintenance, and monitoring in the estuarine environment was carried out. Restoration sites were investigated, and the Salix vegetation was analysed regarding the diversity and structure to answer the questions: (i) does TWFF occur in European lowland estuaries, (ii) what is an appropriate site and design for TWFF restoration, and (iii) do diverse floodplain Salix species differ in structure and function at the estuarine restoration sites?
2. Materials and Methods
2.1. Study Area
The European lowland estuaries of Ems, Elbe, Dutch Delta, and Scheldt were selected as study areas (Figure 2). These estuaries are drastically changed due to dredging for navigation: Elbe (Hamburg port), Ems (cruise shipyard Papenburg), Scheldt (port of Antwerp). This leads to increasing macrotidal conditions (2–6 m tidal amplitude), whereas most of the Dutch Delta shows microtidal conditions due to storm surge barriers between the estuary and the North Sea except for Nieuwe Waterweg (Rotterdam port) [27].
2.2. Determination of Estuarine Sites and Design for Tidal Willow Forest Restoration
Sites for the determination of estuarine sites and the design of tidal willow forest restoration were defined in two steps. First, to receive an overview, a qualitative study on the occurrence of the TWFF in European coastal plain estuaries along the North Sea coast was carried out. Satellite aerial images (Sentinal-2; 2018) of the Elbe estuary (GER), the Ems (GER/NL), the Dutch Delta estuaries (NL), and the Scheldt estuary (NL/B) (Figure 2) were analysed regarding the occurrence of TWFF and Salix species in estuarine foreshores. Based on these findings, an observational study was conducted. The TWFF were studied in the field, and the recorded results of TWFF sites were stated with GPS-position data (eTrex Vista HCx; Garmin International, Inc.; Olathe, KS, USA) and in four overview maps which show the locations of TWFF (Figures 3–6). These maps were created with the freeware program QGIS v.3.22.5 based on Sentinel-2 Data (Sentinel-2 Data © European Union, containing Copernicus Sentinel-2 Data (2018), processed by the Federal Agency for Cartography and Geodesy, GER).
Second, a study of the literature was conducted in order to analyse the policy, governance, and strategies for TWFF restoration and to review estuarine management plans for the Elbe and the Ems estuary, the Dutch Delta estuaries, and the Scheldt estuary.
2.3. Salix Species Selection for Tidal Willow Floodplain Forest Restoration
Tidal willow floodplain forests that occur in tidal wetlands up to the North Sea are composed of Salix alba, S. x rubens, S. fragilis, S. triandra, S. viminalis, and additionally, S. purpurea at sandy locations [8] (Table 1). In tidal freshwater forests along the Scheldt, Dutch delta estuaries, and the Elbe and Ems estuary, S. alba and S. fragilis are characteristic Salix species accompanied by shrubby S. alba, S. caprea, S. dasyclados, S. purpurea, S. triandra, and S. viminalis [27]. However, S. dasyclados (triple hybrid S. caprea × S. cinerea × S. viminalis) has been predominantly planted for willow coppices along the Elbe estuary; S. rubens is an S. alba × S. fragilis hybrid is and thus collectively referred as S. fragilis agg., and S. caprea is not characteristic of floodplain forests [31]. Thus, five willow species (Salix alba, S. fragilis, S. purpurea, S. triandra, and S. viminalis) were selected for the here reported long-term study. The species selection corresponds to a primary investigation on Salix in tidal freshwater wetlands where autochthonous plant material from the tidal freshwater wetlands of the Elbe estuary was collected, cultivated, and used for TWFF restoration measures (see Markus-Michalczyk and Michalczyk [30]).
2.4. Long-Term Structure Analysis of Tidal Willow Floodplain Forest Restoration
Based on the results from the determination of estuarine sites and designs for tidal willow forest restoration, the Elbe estuary was selected as a study site for the long-term analysis of tidal floodplain forest restoration. The Salix specimens that were investigated in the long-term study originated from rods that were collected from the selected individuals of S. alba, S. fragilis, S. purpurea, S. triandra, and S. viminalis in the tidal freshwater wetlands of the Elbe estuary and cultivated in a professional tree nursery. One-year-old rooted 50–80 cm long Salix cuttings were planted in April 2014 at an estuarine upstream site (53°24′25.00′′ N 10°14′46.10′′ E). The juvenile Salix (50 S. alba, 100 S. fragilis, 100 S. purpurea, 110 S. triandra, and 110 S. viminalis specimen) were placed at a 2 m distance among them and planted in groups at the site (see Markus-Michalczyk and Michalczyk [30] for details on the plantation and early establishment).
The early establishment and structure of the different Salix species at the estuarine restoration site were first recorded in September 2015 and in August 2016. The height was measured using a telescope benchmark. Diverse disturbances such as storm surges, wind, and water currents might have affected the Salix specimen close to the marsh edge in addition to periodical tidal flooding, which causes roots to die off and induce inclined up to prostrate trunks. Thus, trunk position and trunk shape were recorded to refer to the complexity of the estuarine environment. Trunk parameters were recorded in categories according to [32]: trunk position (1: erect; 2: inclined; 3: prostrate); trunk shape (1: straight; 2: arching; 3: crooked).
In spring 2017, a substitution plantation was carried out at places where Salix’s early establishment in the plantation failed. It was planned to substitute S. alba (50 individuals) and S. triandra (50 individuals). In autumn 2017, an initial study on the early establishment of the newly planted specimens was conducted. Based on these findings, restoration maintenance measures were applied, in particular, to the exclusion of competition by herbaceous plants and winds. In spring 2019, a second study on the substitution plantation of the Salix specimens was carried out.
In spring 2021, a final study on the initial Salix plantation and the substitution plantation was conducted to record the structure of the TWFF at the restoration site in the tidal freshwater wetlands of the Elbe estuary. The specimen height was measured with a Suunto PM-5/1520 height measurement device (Suunto, Finnland). The diameter was measured at breast height. The trunk shape and trunk position were recorded according to the procedure described for the measurement in 2016 and 2017. Each Salix specimen was given a measured GPS position (Garmin eTrex; Vista HCx), and the distance to the marsh edge was measured via aerial images. ANOVAs were applied to test for the effects of diverse species (Salix alba, S. fragilis, S. purpurea, S. triandra, and S. viminalis) on the height and diameter at breast height.
3. Results
3.1. Estuarine Sites and Design for Tidal Willow Floodplain Forest Restoration
In the qualitative study on the occurrence of TWFF in European lowland estuaries along the North Sea based on aerial images, only several TWFF fragments were found and verified in the field. The location—from the estuarine upstream to the downstream stretch—was determined with GPS-position data (Garmin etrx; Vista HCx) along the Elbe (GER), the Ems (GER/NL), the Dutch Delta estuaries (NL), and the Scheldt estuary (NL/B) which were displayed in four separate satellite maps for each studied estuary (Figure 3, Figure 4, Figure 5 and Figure 6).
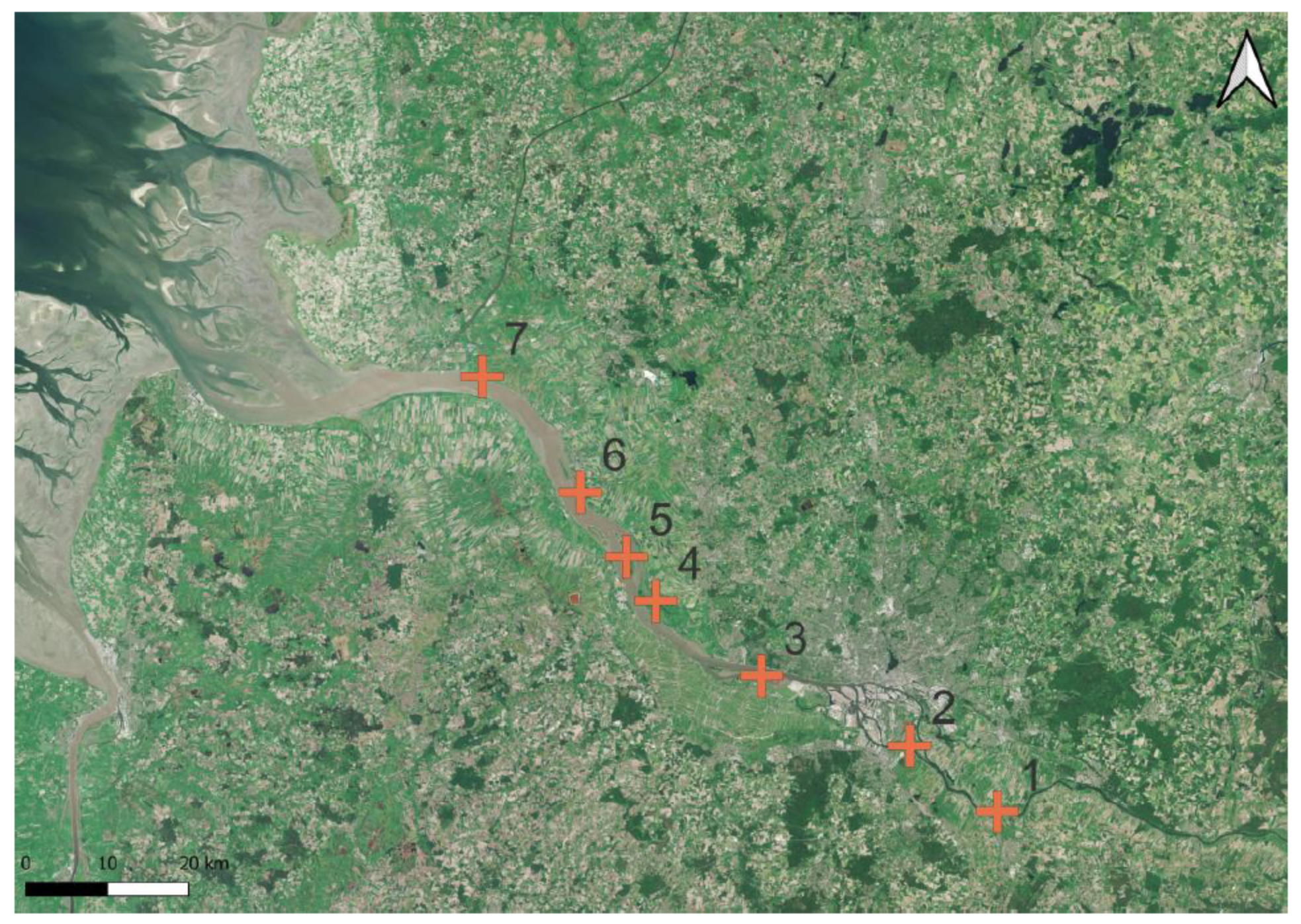
Figure 3.
Locations of TWFF in the Elbe estuary; GER (red plus; numbers 1–7); from the upstream weir (1) to the downstream estuarine mouth (7). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).
Figure 3.
Locations of TWFF in the Elbe estuary; GER (red plus; numbers 1–7); from the upstream weir (1) to the downstream estuarine mouth (7). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).

In the Elbe estuary, from the estuarine border at a weir 140 km distant from the river mouth, TWFF were found in fragments at many locations and at some locations as extended TWFF (Figure 3). Field observations showed that Salix belts along the tidal freshwater stretch occurred particularly in the southern foreshore belonging to the federal state of Lower Saxony. Extended tidal floodplain forests consisting of Salix occurred in the Nature protection areas of Zollenspieker, Heuckenlock, and on the Elbe estuary islands Neßsand in the state of Hamburg. Along the oligohaline stretch, extended TWFF was particularly found on the Elbe estuary islands of Pagensand and Rhinplate as well as in the Nature protection areas of Haseldorfer Binnenelbe mit Elbvorland and Bielenberger Wäldchen (Figure 7a). Small TWFF fragments occurred at some locations at the oligohaline stretch, but the most downstream located potential of TWFF was found in the foreshore of Sankt Margarethen and at the mesohaline stretch of the Elbe estuary.
In the Ems estuary, from the estuarine border at the weir to the river mouth, and similarly to the Elbe estuary, TWFF were found in fragments at many locations and at some locations as extended TWFF (Figure 4). In the field study, numerous small TWFF fragments were particularly found along the tidal freshwater stretch (Figure 7b). An outstanding finding downstream of the Dollart in the mesohaline stretch close to the open sea was an extended TWFF established directly in front of the dike line, which was eventually exposed to seawater during storm surges.
Figure 4.
Location of TWFF in the Ems estuary; GER/NL (red plus; numbers 1–6); from the upstream weir (1) to downstream estuarine mouth (6). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).
Figure 4.
Location of TWFF in the Ems estuary; GER/NL (red plus; numbers 1–6); from the upstream weir (1) to downstream estuarine mouth (6). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).

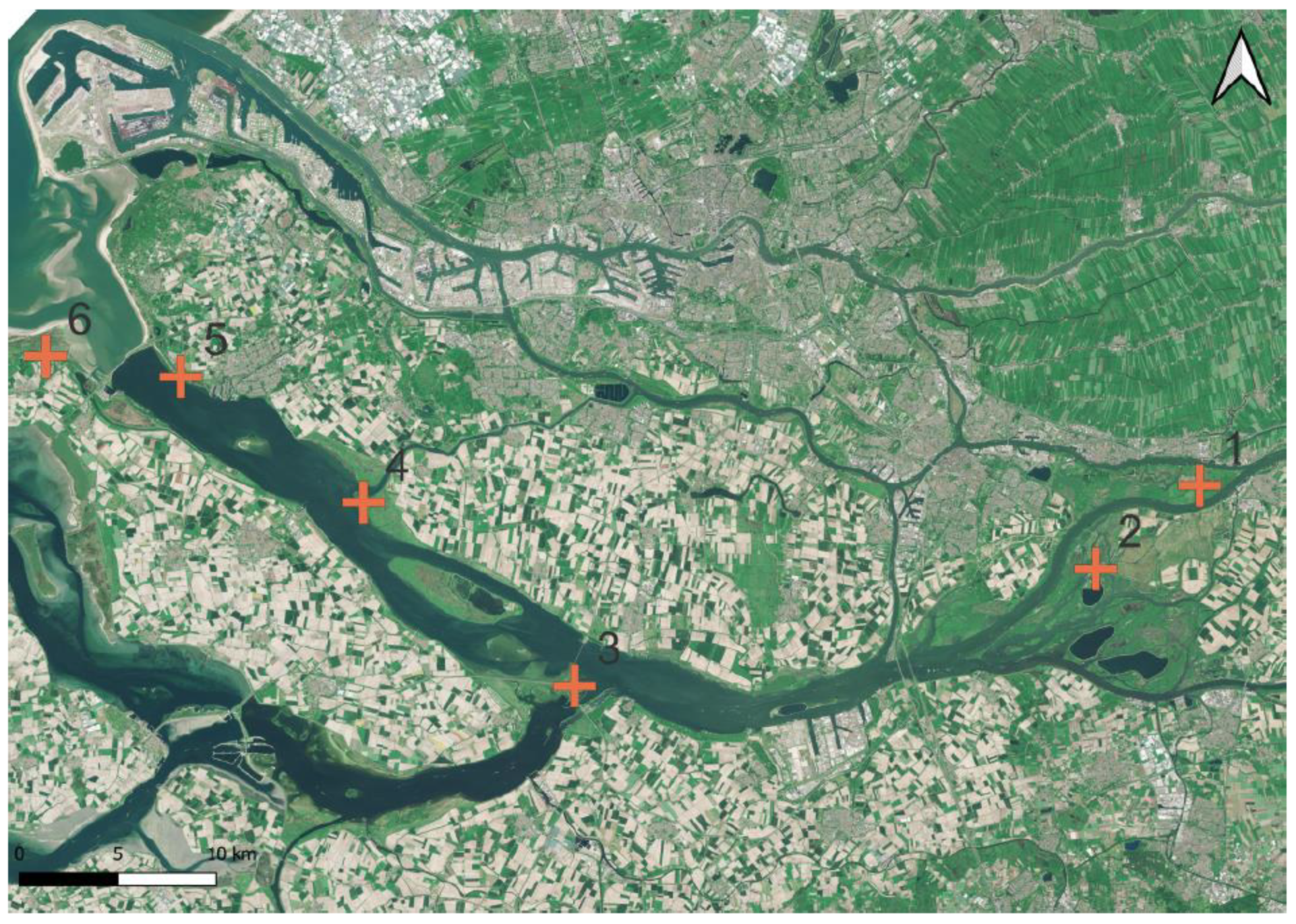
In the Dutch Delta estuaries, TWFF in the aerial images occurred at many locations in small up to medium-sized fragments (Figure 5). In the field, these small TWFF fragments were verified. In the tidal freshwater area of De Biesbosch, the most extended TWFF in the Dutch Delts system was established.
Figure 5.
Locations of TWFF in the Dutch Delta estuaries; NL (red plus; numbers 1-6); from the upstream direction (1) to the downstream estuarine mouth (6). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).
Figure 5.
Locations of TWFF in the Dutch Delta estuaries; NL (red plus; numbers 1-6); from the upstream direction (1) to the downstream estuarine mouth (6). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).

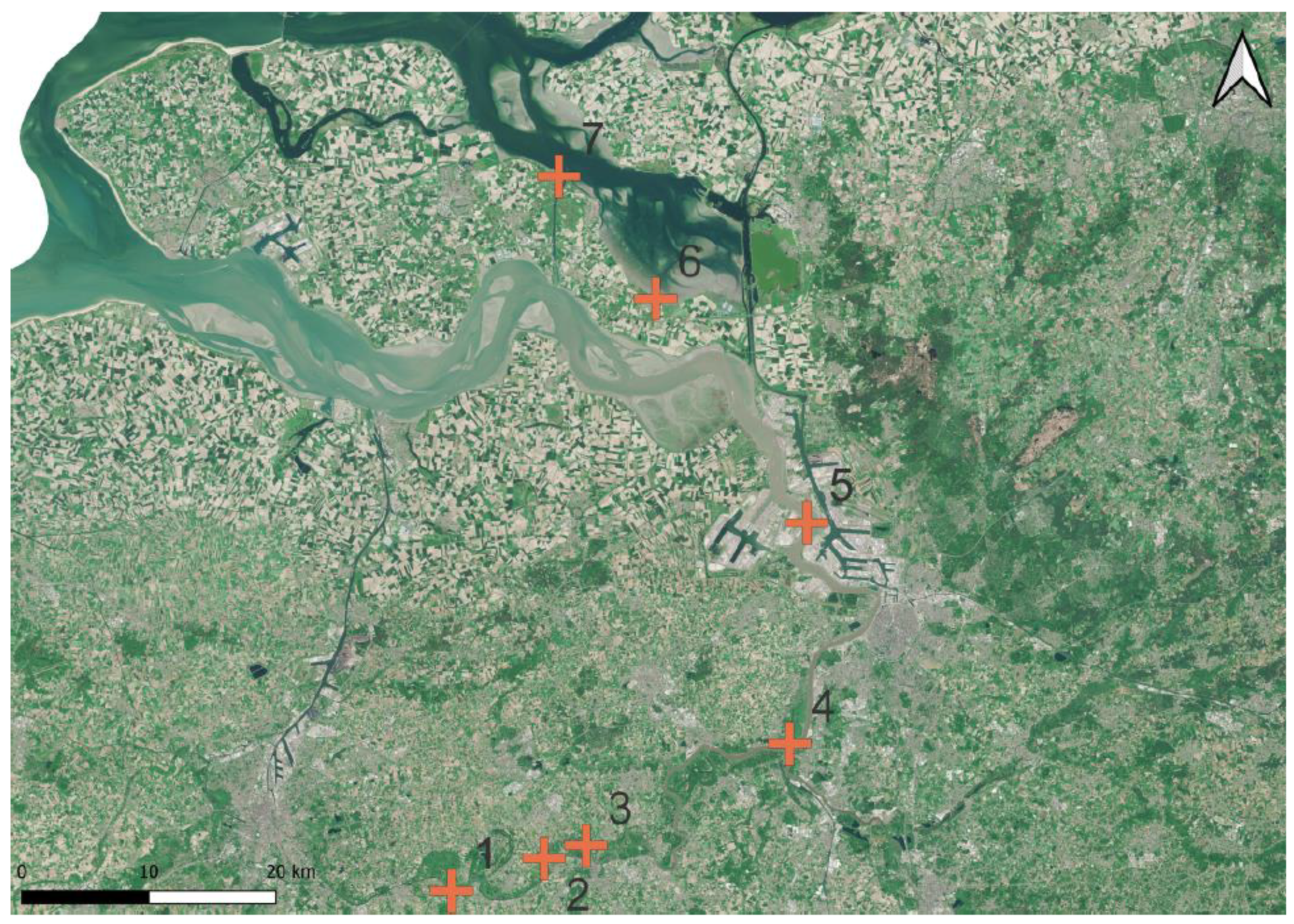
In the Western Scheldt, extended TWFF forest belts were found in aerial images along the estuarine stretch upstream from Antwerp, both in the mesohaline and the tidal freshwater stretch up to Gent (Figure 6, locations 1–5). These forest stands were confirmed as TWFF stands in the field. Downstream from Antwerp and along the brackish estuarine stretch, woodland fragments were found in tidal wetlands in aerial images and were confirmed as TWFF fragments at the site (Figure 7c). In the Eastern Scheldt, a small TWFF fragment was confirmed for the Eastern Scheldt in polyhaline conditions where S. alba and S. viminalis occurred high in the intertidal in front of the dike line (Figure 7d).
Figure 6.
Locations of TWFF in the Scheldt estuary; NL/B (red plus; numbers 1–7); numbers 1–7 from the upstream to the downstream Western Scheldt (1–5) to the Eastern Scheldt (6,7). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).
Figure 6.
Locations of TWFF in the Scheldt estuary; NL/B (red plus; numbers 1–7); numbers 1–7 from the upstream to the downstream Western Scheldt (1–5) to the Eastern Scheldt (6,7). Map created with QGIS v.3.22.5 based on Sentinel-2 Data (2018).

Figure 7.
Tidal willow floodplain forest in the Elbe estuary along the oligohaline stretch (a); Along the tidal freshwater stretch near the shipyard at Papenburg, Ems (GER/NL) (b); Along the brackish stretch downstream of the Antwerp port, Western Scheldt (B) (c); In the foreshore of the polyhaline Eastern Scheldt (NL) (d).
Figure 7.
Tidal willow floodplain forest in the Elbe estuary along the oligohaline stretch (a); Along the tidal freshwater stretch near the shipyard at Papenburg, Ems (GER/NL) (b); Along the brackish stretch downstream of the Antwerp port, Western Scheldt (B) (c); In the foreshore of the polyhaline Eastern Scheldt (NL) (d).


The literature study showed that detailed estuarine management plans for the Elbe and the Ems estuary, the Dutch Delta estuaries and the Scheldt estuary exist, e.g., the integrated management plan for the Elbe estuary (IBP 2012; [33]), the Delta Plan for the Dutch Delta including estuaries, and the Belgium Sigma Plan for the Scheldt estuary (1977; updated 2005). The Dutch Delta Plan aims for coastal protection via the closure of most of the Dutch Delta estuaries (except Nieuwe Waterweg and Western Scheldt for the accessibility of Rotterdam and Antwerp ports) and a reduction in the overall length of the coastline from 700 to 80 km; the Sigma Plan aims to achieve a similar protection level, including the construction of flood control areas, low laying polders next to the estuary surrounded by a lowered ring dike to allow water overflow, and storage and energy dissipation [34]. The Delta Plan and the Sigma Plan were developed in response to a dramatic storm surge in 1953 when large areas of the Scheldt and Dutch Delta estuaries were flooded, and dike breaching led to inundations and the loss of more than 1800 people. Suggestions for ecosystem restoration are lacking in both the Delta Plan and the Sigma Plan. However, the European Habitats Directive [29] protects the biodiversity of species and habitats, and today, most of the European estuaries are designated as protected Natura 2000 sites. Floodplain forests are listed as priority habitats (HD annex I), including tidal willow floodplain forests, and a good conservation status has to be maintained or must be restored. Accordingly, a strategy for improving the conservation status under the HD and measures is being implemented along the Elbe estuary [30]. However, in estuarine locations of large ports (e.g., Antwerp, Hamburg, Rotterdam), integrative management to maintain ecosystems and undertake restoration paralleled by flood risk reduction and harbour activities is challenging. The IBP for the Elbe estuary [33] was the only detailed plan found that suggested measures to improve the conservation status of estuarine habitats with respect to flood control and navigation. Comparable to the dramatic flooding in the Dutch Delta and Scheldt estuary, a storm surge in 1962 and consecutive flooding in the Elbe estuary were regarded as a disaster and marked a turning point in Hamburg’s history, even on the national level. More than 300 people died in the disaster, and 20,000 people lost their homes in Hamburg, making it an anchored memory, particularly for people who have to deal with natural hazards in administration [35]. As such, flood control has to be integrated into all planning at the Elbe estuary as well as in the IBP for the Elbe estuary. Since the IBP [34] is legally based on the HD, TWFF should be developed in the tidal freshwater wetlands from the Hamburg harbour up to the weir, which represents the upstream estuarine border. Thus, the Elbe estuary was selected as a study area for the long-term study of the diversity and structure of the TWFF restoration.
For TWFF restoration along the Elbe estuary upstream stretch, a development concept has been conducted according to the IBP [34], the landscape program (LAPRO), and the Species and Biotope protection program (AUBS) under the evaluation of the biotope mapping and mapping of the Natura 2000 habitats. The results were concealed with the protected areas and protected habitats regulations and aims. The site elevation was analysed based on the digital elevation model “Geoportal online”. A field study on the existing and potential vegetation at the site with regard to TWFF restoration was an integral part of the concept. Finally, the ownership structure was included in developing the concept. The study resulted in the suggestion of 14 sites for restoring TWFF in tidal freshwater wetlands along the 20 km stretch from the upstream estuarine border to the urban area of Hamburg at the Elbe estuary (see Figure 3 for the estuarine upstream stretch).
The restoration of TWFF according to the development plan is being performed one after the other, e.g., by the Elbe Habitat Foundation and the Ministry for Environment in Hamburg. The implementation of TWFF restoration according to the development concept started in 2018. However, a pilot project in TWFF restoration was started in 2014 (see Markus-Michalczyk and Michalczyk [30] for details on the first implementation phase). With this study, the results of a long-term study (2014–2021) on the pilot project in TWFF restoration are provided.
3.2. Long-Term Structure Analysis of Tidal Floodplain Forest Restoration
The long-term study on the TWFF restoration was conducted at the estuarine upstream site (53°24′25.00′′ N 10°14′46.10′′ E; Figure 3 upstream from Hamburg), which corresponds to site five in the tidal willow forest development concept. Here, juvenile willows (50 S. alba, 100 S. fragilis, 100 S. purpurea, 110 S. triandra, and 110 S. viminalis individuals) were placed at a 2 m distance among them and planted in groups at the site (Figure 8). The establishment and structure of Salix species at the estuarine restoration site were first recorded in September 2015 and in August 2016. In comparison to 470 initially (April 2014) planted rooted willow cuttings, 117 juvenile willows were established in 2015; among them were 2 S. alba, 36 S. fragilis, 35 S. purpurea, 1 S. triandra, and 43 S. viminalis individuals. In 2016, the number decreased slightly to 113 willows, including 2 S. alba, 34 S. fragilis, 33 S. purpurea, 1 S. triandra, and 43 S. viminalis individuals (see Markus-Michalczyk and Michalczyk [30] for details on the plantation in 2014 and on the early establishment in 2015 and 2016).
Based on these findings, in spring 2017, a substitution plantation of 100 Salix specimens was carried out at places where early establishment had failed. In autumn 2017, an initial study on the early establishment of the newly planted Salix specimens was conducted. On 2 October, the establishment of 48 of the 100 newly planted Salix specimens was recorded. In total, 15 S. alba and 34 S. viminalis specimens were confirmed. However, it was not possible to record the other newly planted Salix specimen since they were no more visible due to high herbaceous cover and cover by hedge bindweed (Calystegia sepium).
In spring 2019, a second study on the substitution plantation of Salix specimens was carried out. The mean height of the S. alba was 3.58 m (0.69 m ± sd). This indicated a mean height increase of 1.5 m. The height of the S. alba varied between 2 m and 4.3 m. The trunk shape was predominantly straight, and the trunk position was erect.
In spring 2021, a final study on the Salix specimen of the initial plantation (2014) and the substitution plantation (2017) was conducted to record the structure of the TWFF at the restoration site in the tidal freshwater wetlands of the Elbe estuary (Figure 9).
In the study on the initial (2014) planted Salix, 34 S. fragilis individuals, 34 S. viminalis, and 31 S. purpurea individuals were established in the spring of 2021 (Table 2). However, no S. triandra and just two S. alba specimen that was planted in 2014 for restoration in tidal freshwater wetlands at the restoration site in the Elbe estuary were established. The mean height of S. fragilis was 9.2 m, with a maximum height of 11.5 m. The mean diameter was 91 cm, with 118 cm as the maximum. The trunk position was, on average erect, and the shape was between straight up and inclined. Salix viminalis showed a mean height of 5.3 m with a maximum height of 7.5 m and a mean diameter of 95 cm with 89 cm as the maximum. The trunk shape was, on average straight, and the position was inclined. Salix purpurea showed a mean height of 4.1 m with a maximum height of 6 m. The mean diameter was 47 cm, with 105 cm as the maximum. The trunk shape was, on average arched, and the position was prostrate. The mean height of S. alba was 11 m, with a maximum height of 11.5 m and a mean diameter of 80 cm, with 83 cm as the maximum. The trunk shape was straight, and the position was erect.
In the study on the substitution plantation carried out in 2017—due to the establishment failure of the key species S. alba after their initial plantation in 2014—19 S. alba and 25 S. viminalis of the newly planted individuals were found to be established in our study in 2021 (Table 3). The mean height of S. alba was 6.1 m, with a maximum height of 8.5 m. The mean diameter was 35 cm, with 60 cm as the maximum. The trunk position was erect, and the shape was straight. Salix viminalis showed a mean height of 4.4 m with a maximum height of 6.5 m and a mean diameter of 30 cm with 46 cm as the maximum. The trunk shape was, on average straight, and the position was inclined.
4. Discussion
4.1. Estuarine Sites and Design for Tidal Willow Floodplain Forest Restoration
The study showed a limited extension of TWFF in the estuarine environment with small and fragmented patches. This corresponds to the estimation of the UNEP World Conservation Monitoring Centre of more than a 99% loss of the natural distribution of European floodplain forests comprising Salix species [10]. Flow regulation due to dams and dikes restricting both space and flow-induced disturbances is responsible for reduced willow floodplain forest regeneration. Deforestation for pastures further reduced willow floodplain forests and resulted in a lack of bare ground for Salix establishment [36].
In the Elbe estuary, human impact has led to losses in the extent of tidal wetland areas due to dike construction for almost 1000 years. The remaining willow stands were composed of coppiced willows. Since the 1960s, further flood protection measures have resulted in a loss of about 70% of the tidal wetlands at the tidal freshwater stretch [31]. This explains the limitation of TWFF in nature protection areas, e.g., Zollenspieker, Heuckenlock, and Neßsand in the state of Hamburg and along the Elbe estuarine oligohaline stretch on the Elbe islands, including Pagensand and Rhinplate as well as in the nature protection areas of Haseldorfer Binnenelbe mit Elbvorland and Bielenberger Wäldchen (Figure 3 and Figure 7a). Only small TWFF fragments occurred at some locations at the mesohaline stretch. floodplains along the estuarine gradient are categorised into tidal freshwater wetlands, brackish, and salt marshes, with vertical plant species zonation patterns formed according to the estuarine flooding gradient [37], where glycophytes are excluded in saline conditions.
In the Ems estuary, heavy dredging activities during the last decades have led to significantly increased suspended matter concentrations by an order of magnitude and have pushed the estuarine turbidity maximum up the river. The fairway adjustment of the estuary mainly in favour of a big shipyard resulted in a further change in the spatial suspended particulate matter distribution with negative impacts on biota [38]. Furthermore, the tidal influence is largely controlled by a downstream weir to reduce disturbance. Fairway adjustments and flood control measures affect estuarine hydrology and sedimentation-erosion processes ([35,39,40]). On the contrary, hydrodynamics (e.g., waves, currents), ice scour, as well as sediment dynamics have caused high disturbance rates in estuarine environments [41] and supported bare grounds as suitable sites for Salix seedlings’ establishment. Floodplain forest willow species can colonise only bar sediments by abundant seeds and vegetative propagules, where they grow rapidly when high moisture and high radiation is present [7].
The outstanding finding of an extended TWFF downstream of the Dollart in the Ems estuarine mouth close to the open sea suggests that TWFF, which established itself directly in front of the dike line, is resilient to saltwater intrusion. Similarly, the recorded TWFF of the Eastern Scheldt indicates a resilience of TWFF to the conditions in the foreshore in front of the dike line of the polyhaline Eastern Scheldt (Figure 7d). Shifts in salinity are a principal stressor that estuarine organisms have to cope with [41]. The estuarine vegetation of tidal wetlands shows distinct plant distribution patterns along the salinity gradient, with predominantly halophytic species in salt marshes and predominantly glycophytes in tidal freshwater wetlands [37]. Mature S. alba and S. viminalis tolerate oligohaline conditions in the field, and experimental mesocosms studies showed a similar salinity tolerance of juvenile willows up to oligohaline conditions [28]. However, the question of how juvenile and mature Salix specimens respond to short-term harsh salinity pulses during storm surges that occur during their dormant stage during winter remains to be discovered.
Similar to the Ems estuary, the Western Scheldt estuary at the Dutch–Belgium border is characterised by reduced productivity due to limited light availability and heightened turbidity [34]. However, contrary to the Dutch Delta Rhine-Meuse estuaries and the Thames estuary, the Western Scheldt estuary is not dammed by storm surge barriers and thus allows for higher disturbance due to waves and currents. This may have induced the establishment of a TWFF at the brackish stretch downstream of the Antwerp port (Figure 7c). The Scheldt estuary is shaped by narrowed floodplains, embankments, and the major effects of storm surges. Moreover, the deepening of the fairway for free accessibility to the port of Antwerp has led to a largely increased tidal range resulting in the macrotidal Scheldt estuary [34]. Although a concept or implementation of TWFF was neither recorded in the literature nor in the field, these ecosystems covered much of the narrow floodplains in front of the dike line upstream of Antwerp. Moreover, the observed Salix patches in the estuarine brackish stretch that developed by succession indicate the restoration potential for TWFF restoration in the Scheldt estuary. However, clear objectives for TWFF restoration have to be implicated since, until today, most tidal freshwater wetland projects have focused on restoring wetlands dominated by herbaceous plants (marshes) [42].
Along the Rhine-Meuse estuaries in the Netherlands, the Delta Plan for coastal protection led to the closure of most of the estuaries by building dams and barriers. Thus, tidal ranges decreased, and today, predominantly microtidal conditions characterise Rhine-Meuse estuaries in the Dutch Delta [34]. Flood defense in the low-lying areas of the Netherlands is challenging, and innovative flood defense schemes were suggested by Borsje et al. [25], e.g., TWFF restoration was implemented as a supplemental flood protection measure in a pilot project in a former floodplain forest habitat in the Dutch Rhine-Meuse Delta (Figure 1b). Here, willow floodplain forest plantation was used to reduce the overtopping of dykes at the Norwaard polder and, in parallel, maintain landscape value close to the tidal freshwater wetlands of the National Park De Biesbosch [25]. In that study, just the plants of one Salix species with an equal structure were used. However, TWFF consists of high Salix diversity on both the species and the genetic level [7]. Along the hydrologic gradients, Salix specimens were phenotypically clustered at wet sites exhibiting ecological specialisation to the habitat [43]. Thus, diverse Salix specimens and site-adapted plant material should be used for TWFF restoration. This corresponds to Thomas and Nisbet [19], and Wesenbreeck et al. [24], who highlight that caution should be taken to promote trees as a generic solution for flood mitigation since strategically applied and site-adapted solutions should be developed. In addition to flood mitigation criteria, current land use and ownership, as well as the conservation status, should be analysed before defining sites for floodplain forest initialisation [20]. Wesenbreek et al. [24] found that wave-attenuation capacity under extreme conditions was found for coppiced S. alba trees with three-year-old twigs only. Their suggestion that natural and more diverse willow forest ecosystems can play a significant role in flood risk reduction services has not yet been proven. Moreover, Salix provides further services for the estuarine environment since Salix can absorb both organic and inorganic pollutants making Salix an efficient tool for bioremediation [21]. Thus, investigations on more diverse TWFF in structure and function both in the flume and in selected field conditions are suggested. The results of our study may serve as a first suggestion for site selection but are not limited to the Elbe, Ems, Dutch delta, and the Scheldt estuary.
4.2. Long-Term Structure Analysis of Salix Species Diversity in Estuarine Restoration
The long-term study on the structure and function of diverse Salix species that were used in TWFF restoration at an estuarine upstream site (Figure 3, site 5) showed differences in Salix species establishment (Figure 8 and Figure 9; Table 2 and Table 3 ). The initial plantation in 2014 resulted in the establishment of one-quarter of the planted specimen after two years. More than 100 specimens reached a mean height of more than three meters. In 2021, more than 100 initially planted Salix specimens were verified with an increased mean height of more than six meters. This corresponds to the resilience of storm surges and tolerance to eroded sediment, provided that the roots are fully or partially embedded in the sediment [44]. However, the willow species differed in performance. The treelike species S. fragilis reached a mean height of more than 9 m with a trunk diameter greater than 90 cm; S. alba even reached 11 m with an 80 cm trunk diameter, whereas the shrubby species S. viminalis showed a mean height of more than 5 m, more than 70 cm in diameter, and S. purpurea showed a mean height of more than 4 m with a trunk diameter greater than 40 cm. Moreover, the treelike species showed straight and erect trunks, whereas the shrubby willow species showed inclined, arching, and prostrate trunks. For the salt marsh species, hydrodynamic results of a flume study demonstrated that plant species traits interact with environmental conditions resulting in scale-dependent feedback, which explains why the effects of vegetation on landscape formation are species-specific [4]. Accordingly, the shrubby and treelike Salix species could create different feedback on estuarine floodplain formation due to their difference in structure (height, trunk diameter, trunk shape, and trunk position). Resistance values to hydrodynamics generally increased with age and stem diameter. Thus, both juvenile shrubby and treelike floodplain willows with flexible branches of up to five years of age may be more appropriate for use in bioengineering [45] since juvenile twigs show a high bending stability, whereas older branches easily break and are fragmented by floods [7] and may even damage dikes. Stiff and dense mature woody vegetation serves to reduce current velocities but does not bend, and large woods may thus cause turbulence and flooding. On the contrary, complex, flexible, and dense woody vegetation bends in response to currents and reduces velocities, thus inducing lower turbulence. Thus, this vegetation type may be regarded as suitable for bank stabilisation and erosion control, especially when applied in high density [45], and TWFF may act as a NbS, which links NFM and SUDs [13].
It is estimated that if mangroves in the tropics were lost, 15 million more people would be flooded annually across the world [17]. However, it has not yet been quantified which flood control value TWFF provides best for humans in lowland estuaries in the temperate climate. However, with our study, part of the potential of TWFF in lowland estuaries has been shown. Moreover, a multi-criteria evaluation for forest initialisation and management in floodplains with the spatial analysis of various datasets on topography, physical conditions, land use, ownership, and conservation status to define the appropriate sites is required [20] and was applied in our study. To conclude, the studied tidal floodplain forest consisting of diverse Salix species was a successful restoration and a potential design for application as a NbS for flood control in estuaries. However, the quantification of the effects of complex and diverse natural floodplain vegetation for flood control measures is difficult and sites for Salix TWFF restoration and flood control should be selected with caution. More research on diverse Salix species corresponding to natural conditions and in interaction with other estuarine ecosystems is needed to optimise NFM and TWFF restoration and services.
Author Contributions
Conceptualization, H.M.-M.; methodology, H.M.-M. and M.M.; software, H.M.-M. and M.M.; validation, H.M.-M.; formal analysis, H.M.-M. and M.M.; investigation, H.M.-M. and M.M.; resources, H.M.-M.; data curation, H.M.-M. and M.M.; writing—original draft preparation, H.M.-M.; writing—review and editing, H.M.-M. and M.M.; visualization, H.M.-M. and M.M.; supervision, H.M.-M.; project administration, H.M.-M.; funding acquisition, H.M.-M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data are available upon request from the author.
Acknowledgments
Thank goes to the nature conservation authorities for communication and research permission.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schwarz, C.; Gourgue, O.; van Belzen, J.; Zhu, Z.; Bouma, T.J.; van de Koppel, J.; Ruessink, G.; Claude, N. Self-organization of a biogeomorphic landscape controlled by plant life-history traits. Nat. Geosci. 2018, 11, 672–677. [Google Scholar] [CrossRef]
- Corenblit, D.; Steiger, J.; Gurnell, A.M.; Tabacchi, E.; Roques, L. Control of sediment dynamics by vegetation as a key function driving biogeomorphic succession within fluvial corridors. Earth Surf Process. Landf. 2009, 34, 1790–1810. [Google Scholar] [CrossRef]
- Corenblit, D. Species signatures in landscapes. Nat. Geosci. 2018, 11, 621–622. [Google Scholar] [CrossRef]
- Bouma, T.J.; Temmerman, S.; van Duren, L.A.; Martini, E.; Vandenbruwaene, W.; Callaghan, D.P.; Balke, T.; Biermans, G.; Klaassen, P.C.; van Steeg, P.; et al. Organism traits determine the strength of scale-dependent bio-geomorphic feedbacks: A flume study on three intertidal plant species. Geomorphology 2013, 180–181, 57–65. [Google Scholar] [CrossRef]
- Rupprecht, F.; Möller, I.; Paul, M.; Kudella, T.; Spencer, T.; van Wesenbeeck, B.K.; Wolters, G.; Jensen, K.; Bouma, T.J.; Miranda-Lange, M.; et al. Vegetation-wave interactions in salt marshes under storm surge conditions. Ecol. Eng. 2017, 100, 301–315. [Google Scholar] [CrossRef] [Green Version]
- Lotze, H.K.; Lenihan, H.S.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.G.; Kay, M.C.; Kidwell, S.M.; Kirby, M.X.; Petersen, C.H.; Jackson, J.B. Depletion. degradation. and recovery potential of estuaries and coastal seas. Science 2006, 312, 1806–1809. [Google Scholar] [CrossRef]
- Karrenberg, S.; Edwards, P.J.; Kollmann, J. The life history of Saliacaceae living in the active zone of floodplains. Freshw. Biol. 2002, 47, 733–748. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas mit den Alpen. 2010. Sixth ed. Ulmer. Stuttgart.
- Dickmann, D.I.; Kuzovkina, Y.A. Poplars and Willows of the World. With Emphasis on Silvicultural Important Species. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; CAB International and FAO Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2014; pp. 8–91. [Google Scholar]
- UNEP-WCMC. European Forests and Protected Areas: Gap Analysis; United Nations Environmental Program—World Conservation Monitoring Centre: Cambridge, UK, 2000. [Google Scholar]
- Duarte, C.M.; Losada, I.J.; Hendriks, I.; Mazarrasa, I.; Marbà, N. The role of coastal plant communities for climate change mitigation and adaptation. Nat. Clim. Change 2013, 3, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, H.O.; Roberts, D.C.; Adams, H.; Adler, C.; Aldunce, P.; Ali, E.; Begum, R.A.; Betts, R.; Kerr, R.B.; Biesbroek, R.; et al. Climate change 2022: Impacts, Adaptation and Vulnerability; IPCC Sixth Assessment Report; Cambridge University Press: Cambridge, UK; New York, NY, USA, 2022; pp. 37–118. [Google Scholar]
- Lashford, C.; Lavers, T.; Reaney, S.; Charlesworth, S.; Burgess-Gamble, L.; Dale, J. Sustainable Catchment-Wide Flood Management: A Review of the Terminology and Application of Sustainable Catchment Flood Management Techniques in the UK. Water 2022, 14, 1204. [Google Scholar] [CrossRef]
- Costanza, R.; Anderson, S.J.; Sutton, P.; Mulder, K.; Mulder, O.; Kubiszewski, I.; Wang, X.; Liu, X.; Pérez-Maqueo, O.; Martinez, M.L.; et al. The global value of coastal wetlands for storm protection. Glob. Environ. Change 2021, 70, 102328. [Google Scholar] [CrossRef]
- Temmerman, S.; Meire, P.; Bouma, T.J.; Herman, P.M.J.; Ysebaert, T.; De Vriend, H.J. Ecosystem-based coastal defense in the face of global change. Nature 2013, 504, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Ducrotoy, J.; Elliott, M.; Cutts, N.; Franco, A.; Little, S.; Mazik, K.; Wilkinson, M. Temperate Estuaries: Their ecology under future environmental changes. In Coasts and Estuaries: The Future; Wolansk, E., Day, J., Elliott, M., Ramesh, R., Eds.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Menendez, P.; Losada, I.J.; Torres-Ortega, S.; Narayan, S.; Beck, M.W. The global flood protection benefits of mangroves. Sci. Rep. 2020, 10, 4404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isebrands, J.G.; Richardson, J. Introduction. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; CAB International and Food and FAO Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2014; pp. 1–7. [Google Scholar]
- Thomas, H.; Nisbet, T.R. An assessment of the impact of floodplain woodland on flood flows. Water Environ. J. 2007, 21, 114–126. [Google Scholar] [CrossRef]
- Tzioutzios, C.; Kastridis, A. Multi-Criteria Evaluation (MCE) method for the management of woodland plantations in floodplain areas. ISPRS Int. J. Geo.-Inf. 2020, 9, 725. [Google Scholar] [CrossRef]
- Sandil, S.; Gowala, N. Willows: Cost-Effective Tools for Bioremediation of Contaminated Soils. In Advances in Bioremediation and Phytoremediation for Sustainable Soil Management; Malik, J.A., Ed.; Springer: Cham, Switzerland, 2022. [Google Scholar]
- Stanton, B.J.; Serapiglia, M.J.; Smart, L.B. The domestication and Conservation of Poplar and willow genetic resources. In Poplars and Willows: Trees for Society and the Environment; Isebrands, J.G., Richardson, J., Eds.; CAB International and FAO Food and Agriculture Organisation of the United Nations (FAO): Rome, Italy, 2014. [Google Scholar]
- Markus-Michalczyk, H.; Zhu, Z.; Bouma, T.J. Morphological and biomechanical responses of floodplain willows to tidal flooding and salinity. Freshw. Biol. 2019, 64, 913–925. [Google Scholar] [CrossRef]
- van Wesenbeeck, B.K.; Wolters, G.; Antolínez, J.A.; Kalloe, S.A.; Hofland, B.; de Boer, W.P.; Çete, C.; Bouma, T.J. Wave attenuation through forests under extreme conditions. Sci. Rep. 2022, 12, 1884. [Google Scholar] [CrossRef] [PubMed]
- Borsje, B.W.; van Wesenbeeck, B.K.; Dekker, F.; Paalvast, P.; Bouma, T.J.; van Katwijk, M.M.; de Vries, M.B. How ecological engineering can serve in coastal protection. Ecol. Eng. 2011, 37, 113–122. [Google Scholar] [CrossRef]
- Markus-Michalczyk, H.; de Smit, J.; Zhu, Z.; Mchedlishvili, A.; van Bree, J.; Bouma, T.J. Seasonality and spacing determine the effect of juvenile floodplain willows (Salix alba and Salix viminalis) on water current velocities. Estuar. Coast Shelf Sci. 2020, 238, 106697. [Google Scholar] [CrossRef]
- Struyf, E.; Jacobs, S.; Meire, P.; Jensen, K.; Barendregt, A.; Studies, P.C.; Hall, J.V. Plant Communities of European Tidal Freshwater Wetlands; Barendregt, A., Whigham, D.F., Baldwin, A.H., Eds.; Tidal Freshwater Wetlands; Backhuys Publishers: Leiden, The Netherlands, 2009; pp. 59–70. [Google Scholar]
- Markus-Michalczyk, H.; Hanelt, D.; Ludewig, K.; Müller, D. Salt intrusion in tidal wetlands: European willow species tolerate oligohaline conditions. Estuar. Coast Shelf Sci. 2014, 136, 35–42. [Google Scholar] [CrossRef]
- European Habitats Directive. Council Directive 92/43/EEC of 21 May 1992 on the Conservation of Natural Habitats and of Wild Fauna and Flora. Off. J. Eur. Union 1992, 206, 7–50. [Google Scholar]
- Markus-Michalczyk, H.; Michalczyk, C. "Make me a willow cabin at your gate": Legislation and implementation of tidal forest restoration at estuarine upstream sites. Estuar. Coast Shelf Sci. 2018, 210, 1–6. [Google Scholar] [CrossRef]
- Bertram, H.; Brandt, I.; Poppendieck, H.-H.; Preisinger, H.; Prondzinki, J.; Ringenberg, J.; Wiedemann, D. Verbreitungsatlas und Lexikon der Wildpflanzen in Hamburgl; Poppendieck, H.-H., Bertram, H., Brandt, I., Engelschall, B., Prondzinski, J.V., Eds.; Der Hamburger Pflanzenatlas; Dölling und Galtz: München/Hamburg, Germany, 2010; pp. 417–422. [Google Scholar]
- Gockel, H.A. Soziale und Qualitative Entwicklung Sowie Baumhäufigkeiten in Eichenjungbeständen. Master’s Thesis, Universität Göttingen, Göttingen, Germany, 1994. [Google Scholar]
- Integrated Management Plan for the Elbe Estuary. 2012. Available online: http://www.natura2000-unterelbe.de/links-Gesamtplan.php (accessed on 12 December 2022).
- Meire, P.; Dauwe, W.; Maris, T.; Peeters, P.; Coen, L.; Deschamps, M.; Rutten, J.; Temmerman, S. The recent “Saint Nicholas” storm surge in the Scheldt estuary: The Sigma plan proves its efficiency! ECSA Bull. 2014, 62, 19–23. [Google Scholar]
- De Guttry, C.; Ratter, B. Expiry date of a disaster: Memory anchoring and the storm surge 1962 in Hamburg, Germany. ISPRS Int. J. Geo-Inf. 2022, 70, 102719. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J.A. Riverine flood plains: Present state and future trends. Env. Conserv 2002, 29, 8–330. [Google Scholar] [CrossRef] [Green Version]
- Engels, J.G.; Rink, F.; Jensen, K. Stress tolerance and biotic interactions determine plant zonation patterns in estuarine marshes during seedling emergence and early establishment. J. Ecol. 2010, 99, 277–287. [Google Scholar] [CrossRef]
- de Jonge, V.N.; Schuttelaars, H.M.; van Beusekom, J.E.E.; Talke, S.A.; de Swart, H.E. The influence of channel deepening on estuarine turbidity levels and dynamics. as exemplified by the Ems estuary. Estuar. Coast. Shelf. Sci. 2014, 139, 46–59. [Google Scholar] [CrossRef]
- de Jonge, V.N.; Schückel, U. Exploring the effects of dredging and organic waste on the quantitative structure and functioning of the Ems estuary ecosystem by Ecological Network Analysis. Ocean Coast. Manag. 2019, 174, 38–54. [Google Scholar] [CrossRef]
- Elliott, M.; Day, J.W.; Ramachandran, R.; Wolanski, E. 2 A synthesis: What is the future for coasts, estuaries, deltas and other transitional habitats in 2050 and beyond? In Coasts and Estuaries; Wolanski, E., Elliott, M., Ramachandran, R., Eds.; Elsevier: Burlington, UK, 2019; pp. 1–28. [Google Scholar]
- McLusky, D.S.; Elliott, M. The Estuarine Ecosystem, 3rd ed.; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Baldwin, A.H.; Hammerschlag, R.S.; Cahoon, D.R. Evaluating Restored Tidal Freshwater Wetlands. In Coastal Wetlands, 2nd ed.; Perillo, G.M.E., Wolanski, E., Cahoon, D.R., Hopkinson, C.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 889–912. [Google Scholar]
- Savage, J.A.; Cavender-Bares, J.M. Contrasting drought survival strategies of sympatric willows (genus: Salix): Consequence for coexistence and habitat specialization. Tree Physiol. 2011, 31, 604–614. [Google Scholar] [CrossRef]
- Asaeda, T.; Gomes, P.I.; Sakamoto, K.; Rashid, M.H. Tree colonization trends on a sediment bar after a major flood. River Res. Appl. 2011, 27, 976–984. [Google Scholar] [CrossRef]
- Florineth, F. Pflanzen statt Beton. In Handbuch zur Ingenieurbiologie und Vegetationstechnik; Patzer: Berlin/Hannover, Germany, 2004; ISBN 3-87617-107-5. [Google Scholar]
Figure 1.
Juvenile Salix specimen for flood protection along the dike line and for tidal willow floodplain forest restoration in tidal freshwater wetlands of the Elbe estuary (a); Salix specimen grown from stumps for flood protection on banks along the dike line in a tidal freshwater wetland in the Dutch delta system near De Biesbosch (b).
Figure 1.
Juvenile Salix specimen for flood protection along the dike line and for tidal willow floodplain forest restoration in tidal freshwater wetlands of the Elbe estuary (a); Salix specimen grown from stumps for flood protection on banks along the dike line in a tidal freshwater wetland in the Dutch delta system near De Biesbosch (b).

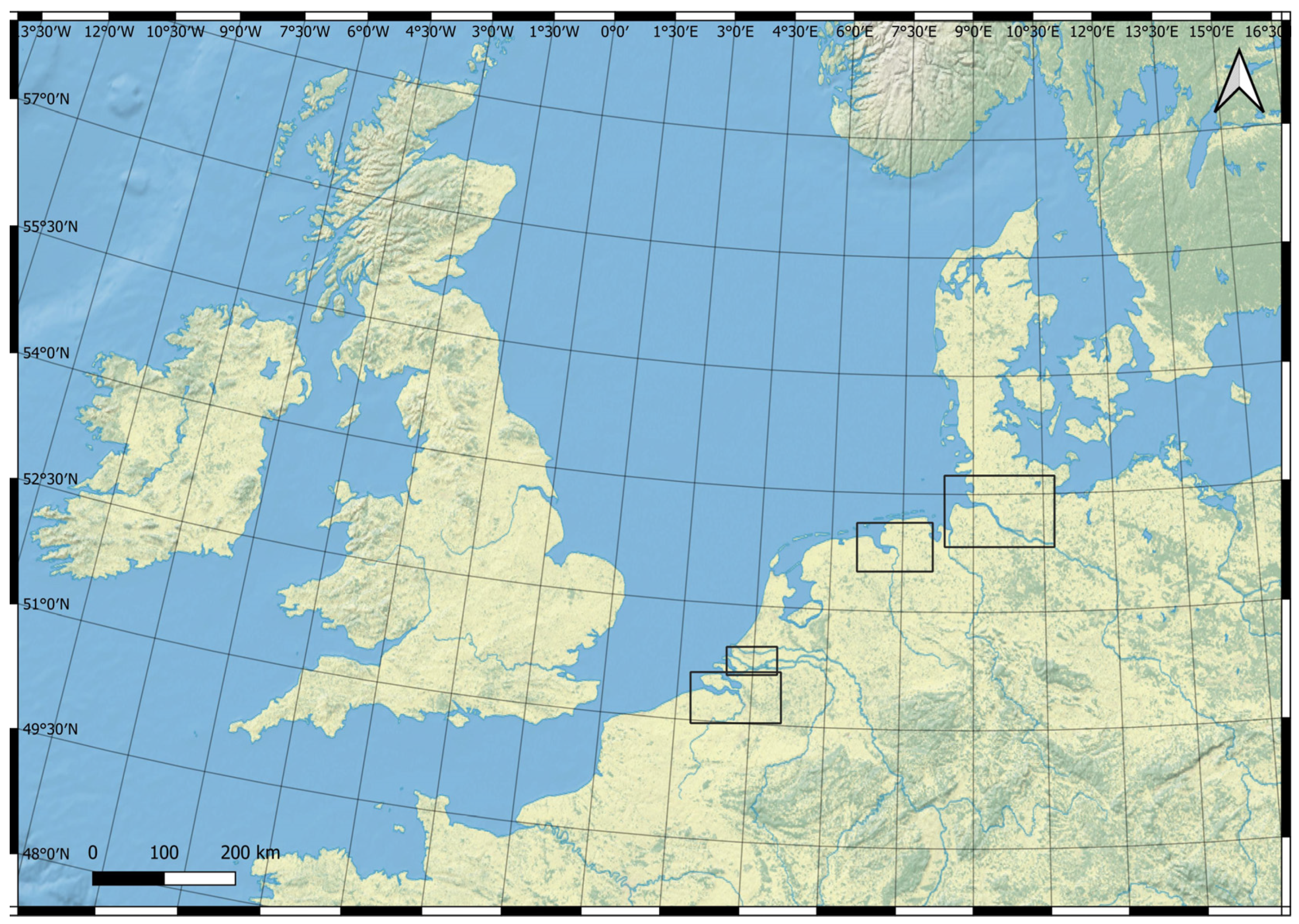
Figure 2.
Study area showing investigated European lowland estuaries along the southern North Sea (black rectangles from north/east to south/west): Elbe estuary (GER), Ems estuary (GER/NL), Dutch Delta estuaries (NL), and Scheldt estuary (NL/B) (Made with Natural Earth. Free vector and raster map data @ naturalearthdata.com, accessed on 4 January 2023).
Figure 2.
Study area showing investigated European lowland estuaries along the southern North Sea (black rectangles from north/east to south/west): Elbe estuary (GER), Ems estuary (GER/NL), Dutch Delta estuaries (NL), and Scheldt estuary (NL/B) (Made with Natural Earth. Free vector and raster map data @ naturalearthdata.com, accessed on 4 January 2023).

Figure 8.
Early establishment of tidal willow floodplain forest six month after initial plantation (2014) in tidal freshwater wetlands in the Elbe estuary.
Figure 8.
Early establishment of tidal willow floodplain forest six month after initial plantation (2014) in tidal freshwater wetlands in the Elbe estuary.

Figure 9.
Established tidal floodplain forest seven years after initial plantation including substitution plantations in tidal freshwater wetlands in the Elbe estuary.
Figure 9.
Established tidal floodplain forest seven years after initial plantation including substitution plantations in tidal freshwater wetlands in the Elbe estuary.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Biological diversity of tidal willow floodplain forests (TWFF).
| Scientific Name | Common Name | Growth Form |
|---|---|---|
| Salix alba L. | White willow | Large tree |
| Salix fragilis agg. | Crack willow | Large tree |
| Salix rubens Schrank | Brittle willow | Large tree |
| Salix caprea L. | Pussy willow | Shrub up to small tree |
| Salix triandra L. | German willow | Shrub up to small tree |
| Salix viminalis L. | Purple willow | Shrub up to small tree |
| Salix dasyclados Wimm. | Almond willow | Shrub |
| Salix purpurea L. | Basket willow | Shrub |
Table 2.
Establishment of TWFF in the Elbe estuary in 2016 (A) and 2021 (B): height (m); diameter (cm); trunk shape (1: straight; 2: arching; 3: crooked); trunk position (1: erect; 2: inclined; 3: prostrate) in S. fragilis, S. viminalis, S. triandra, S. purpurea, and S. alba; N: 34, 34, 3, 31, 2; plantation 2014.
Table 2.
Establishment of TWFF in the Elbe estuary in 2016 (A) and 2021 (B): height (m); diameter (cm); trunk shape (1: straight; 2: arching; 3: crooked); trunk position (1: erect; 2: inclined; 3: prostrate) in S. fragilis, S. viminalis, S. triandra, S. purpurea, and S. alba; N: 34, 34, 3, 31, 2; plantation 2014.
| Salix Species | Height (m) 2016 | Height (m) 2021 | Diameter (cm) 2021 | Trunk Shape 2021 | Trunk Position 2021 |
|---|---|---|---|---|---|
| S. fragilis Mean | 2.5 | 9.2 | 91 | 1.18 | 1.56 |
| Max | 4 | 11.5 | 118 | 3 | 3 |
| S. viminalis mean | 2.0 | 5.3 | 76 | 1.14 | 1.95 |
| max | 3 | 7.5 | 130 | 3 | 3 |
| S. triandra mean | 1.7 | - | - | - | - |
| max | 2 | ||||
| S. purpurea mean | 2.3 | 4.1 | 47 | 1.8 | 2.9 |
| max | 3 | 6 | 105 | 3 | 3 |
| S. alba mean | 1.8 | 11 | 80 | 1 | 1 |
| max | 2.2 | 11.5 | 83 | 1 | 1 |
Table 3.
Establishment of Salix alba (n = 19) and Salix viminalis (n = 25): height (m); diameter (cm); trunk shape (1: straight; 2: arching; 3: crooked); trunk position (1: erect; 2: inclined; 3: prostrate) four years after substitution plantation for TWFF restoration in tidal freshwater wetlands in the Elbe estuary.
Table 3.
Establishment of Salix alba (n = 19) and Salix viminalis (n = 25): height (m); diameter (cm); trunk shape (1: straight; 2: arching; 3: crooked); trunk position (1: erect; 2: inclined; 3: prostrate) four years after substitution plantation for TWFF restoration in tidal freshwater wetlands in the Elbe estuary.
| Salix Species | Height (m) | Diameter (cm) | Trunk Shape | Trunk Position |
|---|---|---|---|---|
| S. alba mean | 6.1 | 39 | 1 | 1.05 |
| max | 8.5 | 60 | 1 | 2 |
| S. viminalis Mean | 4.4 | 29.6 | 1.04 | 1.68 |
| Max | 6.5 | 46 | 2 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Markus-Michalczyk, H.; Michalczyk, M. Floodplain Forest Restoration as a Nature-Based Solution to Create Climate-Resilient Communities in European Lowland Estuaries. Water 2023, 15, 440. https://doi.org/10.3390/w15030440
AMA Style
Markus-Michalczyk H, Michalczyk M. Floodplain Forest Restoration as a Nature-Based Solution to Create Climate-Resilient Communities in European Lowland Estuaries. Water. 2023; 15(3):440. https://doi.org/10.3390/w15030440
Chicago/Turabian StyleMarkus-Michalczyk, Heike, and Matthias Michalczyk. 2023. "Floodplain Forest Restoration as a Nature-Based Solution to Create Climate-Resilient Communities in European Lowland Estuaries" Water 15, no. 3: 440. https://doi.org/10.3390/w15030440
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.