
Arsenite Methyltransferase Is an Important Mediator of Hematotoxicity Induced by Arsenic in Drinking Water
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Mice
2.3. Drinking Water Exposures
2.4. Collection of Blood, Preparation of Serum, and Hematology Analysis
2.5. Isolation of Primary Mouse Bone Marrow Cells
2.6. Isolation of Primary Mouse Spleen Cells
2.7. Bone Marrow BFU-E
2.8. Flow Cytometry
2.9. Mouse EPO ELISA
3. Arsenic Speciation by Hydride Generation-Cryotrapping-Inductively Coupled Plasma-Mass Spectrometry
Statistics
4. Results
4.1. AsIII Exposure Does Not Significantly Modify Mouse Body Weights, Drinking Water Consumption, Bone Marrow or Spleen Cell Recoveries, or Spleen Weights in Wild-Type and As3mt-KO Mice
4.2. Hematological Indicators of Anemia Are Reduced Following AsIII Exposure in Wild-Type, but Not As3mt-KO Mice
4.3. Abundance and Distribution of Arsenic Species in the Bone Marrow, Spleen, and Plasma of AsIII Exposed Wild-Type and As3mt-KO Mice
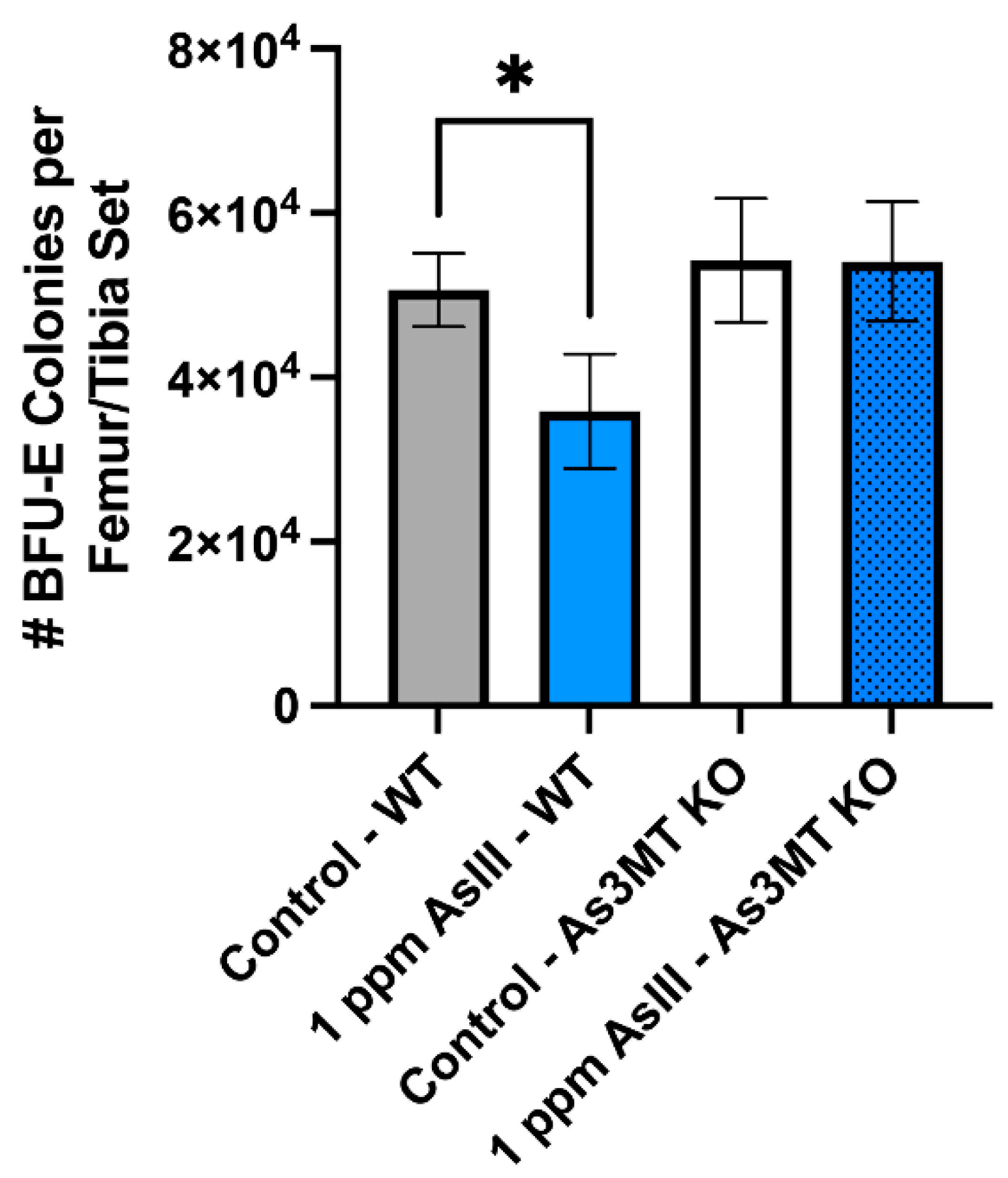
4.4. Attenuation of BFU-E Colony Formation in Wild-Type, but Not As3mt-KO Mice Following 60-Day AsIII Exposure
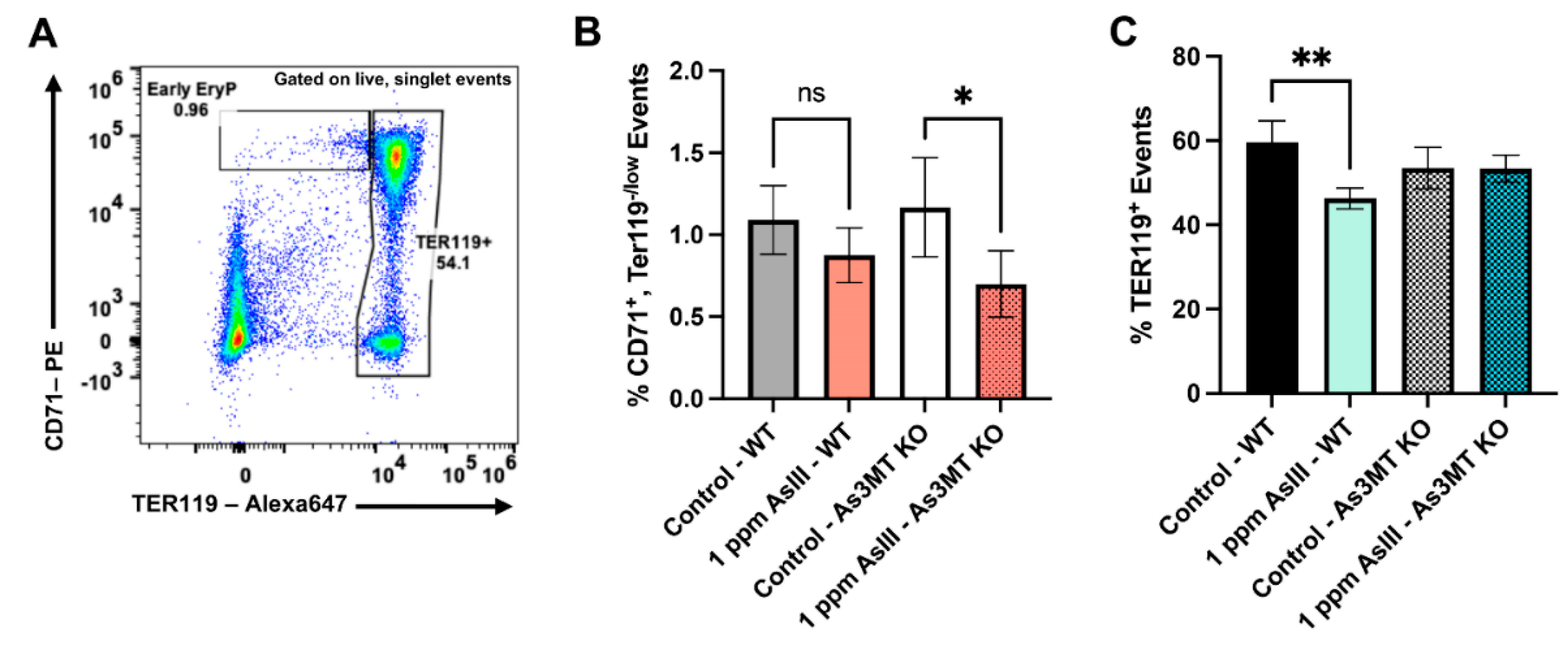
4.5. Early and Late-Stage Erythroblast Subsets Are Altered in the Bone Marrow of Wild-Type and As3mt-KO Mice Following Exposure to AsIII
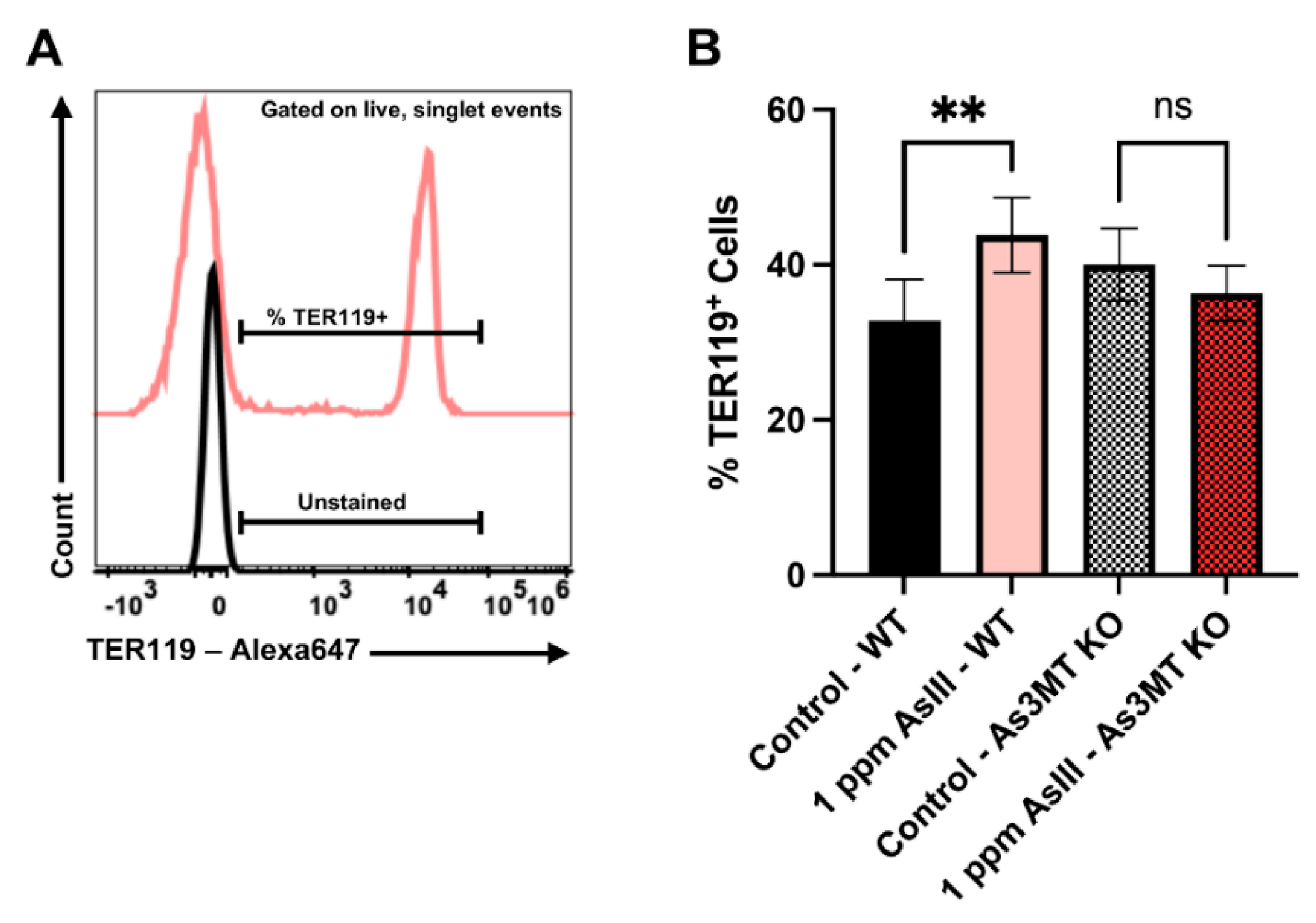
4.6. AsIII Exposure Increases Late-Stage, TER119+ Erythroblasts in the Spleens of Wild-Type, but Not As3mt-KO Mice
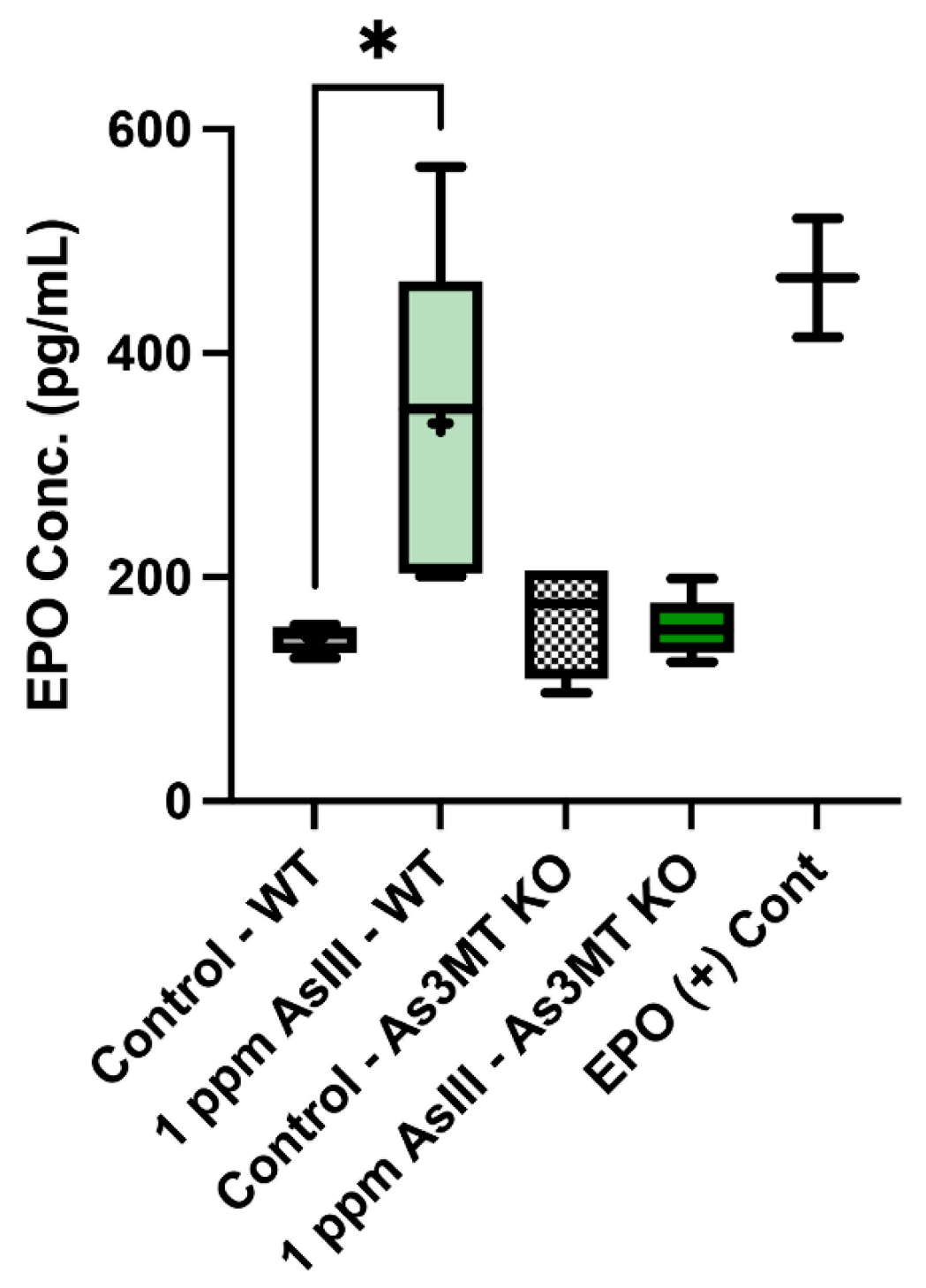
4.7. Levels of Circulating EPO Are Increased Following AsIII Exposure in Wild-Type, but Not As3mt-KO Mice
5. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Arsenic in Drinking-Water. 2019. Available online: https://www.who.int/publications/i/item/arsenic-in-drinking-water-background-document-for-development-of-who-guidelines-for-drinking-water-quality (accessed on 1 October 2022).
- Ferrario, D.; Gribaldo, L.; Hartung, T. Arsenic Exposure and Immunotoxicity: A Review Including the Possible Influence of Age and Sex. Curr. Environ. Health Rep. 2016, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Heck, J.E.; Chen, Y.; Grann, V.R.; Slavkovich, V.; Parvez, F.; Ahsan, H. Arsenic exposure and anemia in Bangladesh: A population-based study. J. Occup. Environ. Med. 2008, 50, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.F. Arsenic toxicity and potential mechanisms of action. Toxicol. Lett. 2002, 133, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Naujokas, M.F.; Anderson, B.; Ahsan, H.; Aposhian, H.V.; Graziano, J.H.; Thompson, C.; Suk, W.A. The broad scope of health effects from chronic arsenic exposure: Update on a worldwide public health problem. Environ. Health Perspect. 2013, 121, 295–302. [Google Scholar] [CrossRef]
- Tyler, C.R.; Allan, A.M. The Effects of Arsenic Exposure on Neurological and Cognitive Dysfunction in Human and Rodent Studies: A Review. Curr. Environ. Health Rep. 2014, 1, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Kile, M.L.; Faraj, J.M.; Ronnenberg, A.G.; Quamruzzaman, Q.; Rahman, M.; Mostofa, G.; Afroz, S.; Christiani, D.C. A cross sectional study of anemia and iron deficiency as risk factors for arsenic-induced skin lesions in Bangladeshi women. BMC Public Health 2016, 16, 158. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, K.K.; Guha Mazumder, D.N.; Ghose, N.; Ghose, A.; Lahiri, S. Systemic manifestations in chronic arsenic toxicity in absence of skin lesions in West Bengal. Indian J. Med. Res. 2009, 129, 75–82. [Google Scholar] [PubMed]
- Surdu, S.; Bloom, M.S.; Neamtiu, I.A.; Pop, C.; Anastasiu, D.; Fitzgerald, E.F.; Gurzau, E.S. Consumption of arsenic-contaminated drinking water and anemia among pregnant and non-pregnant women in northwestern Romania. Environ. Res. 2015, 140, 657–660. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. The Global Prevalence of Anaemia in 2011. 2015. Available online: https://apps.who.int/iris/handle/10665/177094 (accessed on 1 October 2022).
- Hopenhayn, C.; Bush, H.M.; Bingcang, A.; Hertz-Picciotto, I. Association between arsenic exposure from drinking water and anemia during pregnancy. J. Occup. Environ. Med. 2006, 48, 635–643. [Google Scholar] [CrossRef]
- Kassebaum, N.J.; Jasrasaria, R.; Naghavi, M.; Wulf, S.K.; Johns, N.; Lozano, R.; Regan, M.; Weatherall, D.; Chou, D.P.; Eisele, T.P.; et al. A systematic analysis of global anemia burden from 1990 to 2010. Blood 2014, 123, 615–624. [Google Scholar] [CrossRef]
- Breton, C.V.; Houseman, E.A.; Kile, M.L.; Quamruzzaman, Q.; Rahman, M.; Mahiuddin, G.; Christiani, D.C. Gender-specific protective effect of hemoglobin on arsenic-induced skin lesions. Cancer Epidemiol. Biomark. Prev. 2006, 15, 902–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- United States Environmental Protection Agency. 2012 Edition of the Drinking Water Standards and Health Advisories Table; United States Environmental Protection Agency: Washington, DC, USA, 2012.
- Agency for Toxic Substances and Disease Registry. Toxicological Profile for Arsenic; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2007.
- Agency for Toxic Substances and Disease Registry. Addendum to the Toxicological Profile for Arsenic; Agency for Toxic Substances and Disease Registry: Atlanta, GA, USA, 2016.
- Styblo, M.; Del Razo, L.M.; Vega, L.; Germolec, D.R.; LeCluyse, E.L.; Hamilton, G.A.; Reed, W.; Wang, C.; Cullen, W.R.; Thomas, D.J. Comparative toxicity of trivalent and pentavalent inorganic and methylated arsenicals in rat and human cells. Arch. Toxicol. 2000, 74, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.J.; Styblo, M.; Lin, S. The cellular metabolism and systemic toxicity of arsenic. Toxicol. Appl. Pharmacol. 2001, 176, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Vahter, M. Mechanisms of arsenic biotransformation. Toxicology 2002, 181–182, 211–217. [Google Scholar] [CrossRef]
- Lin, S.; Shi, Q.; Nix, F.B.; Styblo, M.; Beck, M.A.; Herbin-Davis, K.M.; Hall, L.L.; Simeonsson, J.B.; Thomas, D.J. A novel S-adenosyl-L-methionine:arsenic(III) methyltransferase from rat liver cytosol. J. Biol. Chem. 2002, 277, 10795–10803. [Google Scholar] [CrossRef] [Green Version]
- Thomas, D.J.; Li, J.; Waters, S.B.; Xing, W.; Adair, B.M.; Drobna, Z.; Devesa, V.; Styblo, M. Arsenic (+3 oxidation state) methyltransferase and the methylation of arsenicals. Exp. Biol. Med. 2007, 232, 3–13. [Google Scholar]
- Douillet, C.; Huang, M.C.; Saunders, R.J.; Dover, E.N.; Zhang, C.; Styblo, M. Knockout of arsenic (+3 oxidation state) methyltransferase is associated with adverse metabolic phenotype in mice: The role of sex and arsenic exposure. Arch. Toxicol. 2017, 91, 2617–2627. [Google Scholar] [CrossRef]
- Engstrom, K.; Vahter, M.; Mlakar, S.J.; Concha, G.; Nermell, B.; Raqib, R.; Cardozo, A.; Broberg, K. Polymorphisms in arsenic(+III oxidation state) methyltransferase (AS3MT) predict gene expression of AS3MT as well as arsenic metabolism. Environ. Health Perspect. 2011, 119, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Vahter, M. Variation in human metabolism of arsenic. In Arsenic Exposure and Health Effects; Chappell, W.R., Abernathy, C.O., Calderon, R.L., Eds.; Elsevier Science Ltd.: Amsterdam, The Netherlands, 1999; pp. 267–279. [Google Scholar]
- Vahter, M.; Concha, G.; Nermell, B.; Nilsson, R.; Dulout, F.; Natarajan, A.T. A unique metabolism of inorganic arsenic in native Andean women. Eur. J. Pharmacol. 1995, 293, 455–462. [Google Scholar] [CrossRef]
- Dzierzak, E.; Philipsen, S. Erythropoiesis: Development and differentiation. Cold Spring Harb. Perspect. Med. 2013, 3, a011601. [Google Scholar] [CrossRef]
- Palis, J. Primitive and definitive erythropoiesis in mammals. Front. Physiol. 2014, 5, 3. [Google Scholar] [CrossRef] [PubMed]
- Orkin, S.H.; Zon, L.I. Hematopoiesis: An evolving paradigm for stem cell biology. Cell 2008, 132, 631–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rieger, M.A.; Schroeder, T. Hematopoiesis. Cold Spring Harb. Perspect. Biol. 2012, 4, a008250. [Google Scholar] [CrossRef] [PubMed]
- Medina, S.; Xu, H.; Wang, S.C.; Lauer, F.T.; Liu, K.J.; Burchiel, S.W. Low level arsenite exposures suppress the development of bone marrow erythroid progenitors and result in anemia in adult male mice. Toxicol. Lett. 2017, 273, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Medina, S.; Bolt, A.M.; Zhang, H.; Wan, G.; Xu, H.; Lauer, F.T.; Wang, S.C.; Burchiel, S.W.; Liu, K.J. Inhibition of red blood cell development by arsenic-induced disruption of GATA-1. Sci. Rep. 2020, 10, 19055. [Google Scholar] [CrossRef]
- Medina, S.; Bolt, A.M.; Zhou, X.; Wan, G.; Xu, H.; Lauer, F.T.; Liu, K.J.; Burchiel, S.W. Arsenite and monomethylarsonous acid disrupt erythropoiesis through combined effects on differentiation and survival pathways in early erythroid progenitors. Toxicol. Lett. 2021, 350, 111–120. [Google Scholar] [CrossRef]
- Medina, S.; Zhang, H.; Santos-Medina, L.V.; Wan, G.; Bolt, A.M.; Zhou, X.; Burchiel, S.W.; Liu, K.J. Arsenic impairs the lineage commitment of hematopoietic progenitor cells through the attenuation of GATA-2 DNA binding activity. Toxicol. Appl. Pharmacol. 2022, 452, 116193. [Google Scholar] [CrossRef]
- Wan, G.; Medina, S.; Zhang, H.; Pan, R.; Zhou, X.; Bolt, A.M.; Luo, L.; Burchiel, S.W.; Liu, K.J. Arsenite exposure inhibits the erythroid differentiation of human hematopoietic progenitor CD34(+) cells and causes decreased levels of hemoglobin. Sci. Rep. 2021, 11, 22121. [Google Scholar] [CrossRef]
- Drobna, Z.; Naranmandura, H.; Kubachka, K.M.; Edwards, B.C.; Herbin-Davis, K.; Styblo, M.; Le, X.C.; Creed, J.T.; Maeda, N.; Hughes, M.F.; et al. Disruption of the arsenic (+3 oxidation state) methyltransferase gene in the mouse alters the phenotype for methylation of arsenic and affects distribution and retention of orally administered arsenate. Chem. Res. Toxicol. 2009, 22, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Medina, S.; Lauer, F.T.; Castillo, E.F.; Bolt, A.M.; Ali, A.S.; Liu, K.J.; Burchiel, S.W. Exposures to uranium and arsenic alter intraepithelial and innate immune cells in the small intestine of male and female mice. Toxicol. Appl. Pharmacol. 2020, 403, 115155. [Google Scholar] [CrossRef]
- Vahter, M. Methylation of inorganic arsenic in different mammalian species and population groups. Sci. Prog. 1999, 82 Pt 1, 69–88. [Google Scholar] [CrossRef] [PubMed]
- Parvez, F.; Medina, S.; Santella, R.M.; Islam, T.; Lauer, F.T.; Alam, N.; Eunus, M.; Rahman, M.; Factor-Litvak, P.; Ahsan, H.; et al. Arsenic exposures alter clinical indicators of anemia in a male population of smokers and non-smokers in Bangladesh. Toxicol. Appl. Pharmacol. 2017, 331, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Ezeh, P.C.; Xu, H.; Wang, S.C.; Medina, S.; Burchiel, S.W. Evaluation of Toxicity in Mouse Bone Marrow Progenitor Cells. Curr. Protoc. Toxicol. 2016, 67, 18.9.1.–18.9.2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; McClain, S.; Medina, S.; Lauer, F.T.; Douillet, C.; Liu, K.J.; Hudson, L.G.; Styblo, M.; Burchiel, S.W. Differential sensitivities of bone marrow, spleen and thymus to genotoxicity induced by environmentally relevant concentrations of arsenite. Toxicol. Lett. 2016, 262, 55–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balarajan, Y.; Ramakrishnan, U.; Ozaltin, E.; Shankar, A.H.; Subramanian, S.V. Anaemia in low-income and middle-income countries. Lancet 2011, 378, 2123–2135. [Google Scholar] [CrossRef]
- Ezeh, P.C.; Xu, H.; Lauer, F.T.; Liu, K.J.; Hudson, L.G.; Burchiel, S.W. Monomethylarsonous acid (MMA+3) Inhibits IL-7 Signaling in Mouse Pre-B Cells. Toxicol. Sci. 2016, 149, 289–299. [Google Scholar] [CrossRef] [Green Version]
- Petrick, J.S.; Jagadish, B.; Mash, E.A.; Aposhian, H.V. Monomethylarsonous acid (MMA(III)) and arsenite: LD(50) in hamsters and in vitro inhibition of pyruvate dehydrogenase. Chem. Res. Toxicol. 2001, 14, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Medina, S.; Lauer, F.T.; Douillet, C.; Liu, K.J.; Styblo, M.; Burchiel, S.W. Genotoxicity induced by monomethylarsonous acid (MMA(+3)) in mouse thymic developing T cells. Toxicol. Lett. 2017, 279, 60–66. [Google Scholar] [CrossRef]
- Chen, B.; Arnold, L.L.; Cohen, S.M.; Thomas, D.J.; Le, X.C. Mouse arsenic (+3 oxidation state) methyltransferase genotype affects metabolism and tissue dosimetry of arsenicals after arsenite administration in drinking water. Toxicol. Sci. 2011, 124, 320–326. [Google Scholar] [CrossRef] [Green Version]
- Ahsan, H.; Chen, Y.; Kibriya, M.G.; Slavkovich, V.; Parvez, F.; Jasmine, F.; Gamble, M.V.; Graziano, J.H. Arsenic metabolism, genetic susceptibility, and risk of premalignant skin lesions in Bangladesh. Cancer Epidemiol. Biomark. Prev. 2007, 16, 1270–1278. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Parvez, F.; Gamble, M.; Islam, T.; Ahmed, A.; Argos, M.; Graziano, J.H.; Ahsan, H. Arsenic exposure at low-to-moderate levels and skin lesions, arsenic metabolism, neurological functions, and biomarkers for respiratory and cardiovascular diseases: Review of recent findings from the Health Effects of Arsenic Longitudinal Study (HEALS) in Bangladesh. Toxicol. Appl. Pharmacol. 2009, 239, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.C.; Guo, Y.L.; Su, H.J.; Hsueh, Y.M.; Smith, T.J.; Ryan, L.M.; Lee, M.S.; Chao, S.C.; Lee, J.Y.; Christiani, D.C. Arsenic methylation and skin cancer risk in southwestern Taiwan. J. Occup. Environ. Med. 2003, 45, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.K.; Pu, Y.S.; Chung, C.J.; Shiue, H.S.; Yang, M.H.; Chen, C.J.; Hsueh, Y.M. Plasma folate level, urinary arsenic methylation profiles, and urothelial carcinoma susceptibility. Food Chem. Toxicol. 2008, 46, 929–938. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Chen, W.J.; Huang, C.Y.; Shiue, H.S.; Su, C.T.; Ao, P.L.; Pu, Y.S.; Hsueh, Y.M. Polymorphisms of Arsenic (+3 Oxidation State) Methyltransferase and Arsenic Methylation Capacity Affect the Risk of Bladder Cancer. Toxicol. Sci. 2018, 164, 328–338. [Google Scholar] [CrossRef] [Green Version]
- Luo, L.; Li, Y.; Gao, Y.; Zhao, L.; Feng, H.; Wei, W.; Qiu, C.; He, Q.; Zhang, Y.; Fu, S.; et al. Association between arsenic metabolism gene polymorphisms and arsenic-induced skin lesions in individuals exposed to high-dose inorganic arsenic in northwest China. Sci. Rep. 2018, 8, 413. [Google Scholar] [CrossRef] [Green Version]
- Yu, R.C.; Hsu, K.H.; Chen, C.J.; Froines, J.R. Arsenic methylation capacity and skin cancer. Cancer Epidemiol. Biomark. Prev. 2000, 9, 1259–1262. [Google Scholar]
- Gamble, M.V.; Liu, X.; Ahsan, H.; Pilsner, R.; Ilievski, V.; Slavkovich, V.; Parvez, F.; Levy, D.; Factor-Litvak, P.; Graziano, J.H. Folate, homocysteine, and arsenic metabolism in arsenic-exposed individuals in Bangladesh. Environ. Health Perspect. 2005, 113, 1683–1688. [Google Scholar] [CrossRef] [Green Version]
- Vahter, M.; Concha, G. Role of metabolism in arsenic toxicity. Pharmacol. Toxicol. 2001, 89, 1–5. [Google Scholar] [CrossRef]
- Paulson, R.F.; Shi, L.; Wu, D.C. Stress erythropoiesis: New signals and new stress progenitor cells. Curr. Opin. Hematol. 2011, 18, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Socolovsky, M. Molecular insights into stress erythropoiesis. Curr. Opin. Hematol. 2007, 14, 215–224. [Google Scholar] [CrossRef]
- Kim, T.S.; Hanak, M.; Trampont, P.C.; Braciale, T.J. Stress-associated erythropoiesis initiation is regulated by type 1 conventional dendritic cells. J. Clin. Investig. 2015, 125, 3965–3980. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Wild-Type | As3mt-KO | |||
|---|---|---|---|---|
| Parameter | Control | 1 ppm AsIII | Control | 1 ppm AsIII |
| Body Wt. (g) | 32.1 ± 3.18 | 31.38 ± 1.12 | 29.50 ± 1.78 | 29.99 ± 3.23 |
| Water Consumption (mL/mouse/wk) | 21.00 ± 1.50 | 21.00 ± 4.00 | 17.00 ± 0.98 | 19.00 ± 2.00 |
| Bone Marrow Cell Recovery (Total viable cells per femur/tibia set, ×106) | 57.90 ± 6.38 | 51.14 ± 11.75 | 66.92 ± 8.24 | 63.26 ± 11.06 |
| Spleen Wt. (g) | 80.80 ± 8.20 | 99.80 ± 18.49 | 79.4 ± 4.89 | 76.4 ± 7.30 |
| Spleen Cell Recovery (Total viable cells, ×106) | 153 ± 15.3 | 150 ± 15.3 | 98.66 ± 15.30 | 109.24 ± 18.97 |
| Wild-Type | As3mt-KO | |||
|---|---|---|---|---|
| Parameter | Control | 1 ppm AsIII | Control | 1 ppm AsIII |
| RBC Count (×1012/L) | 12.17 ± 0.13 | 11.21 ± 0.22 *** | 11.66 ± 0.25 | 11.47 ± 0.75 |
| Hct (%) | 52.08 ± 0.51 | 49.1 ± 0.38 *** | 48.56 ± 1.26 | 48.12 ± 3.47 |
| Hgb (g/dL) | 16.7 ± 0.47 | 16.12 ± 0.30 * | 14.96 ± 2.21 | 15.66 ± 1.21 |
| MCH (pg) | 13.98 ± 0.53 | 14.38 ± 0.46 | 13.58 ± 0.29 | 13.66 ± 0.30 |
| MCV (fL) | 43 ± 0.71 | 43.8 ± 0.84 | 41.8 ± 0.45 | 41.8 ± 0.45 |
| Tissues | Genotype | Exposure | As Amount by Species (pg) | |||
|---|---|---|---|---|---|---|
| iAs | MAs | DMAs | Total As | |||
| Bone Marrow | Wild-type | Control | 1.50 ± 0.82 | 0.22 ± 0.09 | 0.75 ± 0.15 | 2.46 ± 0.89 |
| 1 ppm AsIII | 4.80 ± 2.43 | 2.66 ± 2.14 * | 23.23 ± 9.71 **** | 30.69 ± 13.78 | ||
| As3mt-KO | Control | 11.63 ± 1.02 | 0.37 ± 0.12 | 0.25 ± 0.12 | 12.25 ± 1.06 | |
| 1 ppm AsIII | 134.27 ± 28.71 **** | 1.55 ± 0.24 | 0.23 ± 0.09 | 136.05 ± 28.50 **** | ||
| Spleen | Wild-type | Control | 6.68 ± 4.82 | 0.19 ± 0.13 | 0.26 ± 0.14 | 7.14 ± 4.97 |
| 1 ppm AsIII | 10.04 ± 3.32 | 0.75 ± 0.38 ** | 5.22 ± 1.61 **** | 16.00 ± 2.65 | ||
| As3mt-KO | Control | 30.14 ± 11.02 | 0.33 ± 0.13 | 0.21 ± 0.08 | 30.68 ± 11.18 | |
| 1 ppm AsIII | 89.33 ± 17.79 **** | 0.98 ± 0.20 ** | 0.14 ± 0.05 | 90.45 ± 17.90 **** | ||
| Plasma | Wild-type | Control | 12.83 ± 6.47 | 2.29 ± 0.91 | 218.98 ± 54.90 | 234.11 ± 61.23 |
| 1 ppm AsIII | 92.32 ± 39.29 | 117.6 ± 3.56 **** | 6956.65 ± 1495.13 **** | 7166.57 ± 1533.08 **** | ||
| As3mt-KO | Control | 243.74 ± 142.48 | 2.44 ± 1.32 | 1.49 ± 0.62 | 247.67 ± 144.19 | |
| 1 ppm AsIII | 1537.93 ± 257.47 **** | 26.33 ± 2.14 **** | 8.81 ± 4.67 | 1573.07 ± 254.08 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Medina, S.; Zhang, H.; Santos-Medina, L.V.; Yee, Z.A.; Martin, K.J.; Wan, G.; Bolt, A.M.; Zhou, X.; Stýblo, M.; Liu, K.J. Arsenite Methyltransferase Is an Important Mediator of Hematotoxicity Induced by Arsenic in Drinking Water. Water 2023, 15, 448. https://doi.org/10.3390/w15030448
Medina S, Zhang H, Santos-Medina LV, Yee ZA, Martin KJ, Wan G, Bolt AM, Zhou X, Stýblo M, Liu KJ. Arsenite Methyltransferase Is an Important Mediator of Hematotoxicity Induced by Arsenic in Drinking Water. Water. 2023; 15(3):448. https://doi.org/10.3390/w15030448
Chicago/Turabian StyleMedina, Sebastian, Haikun Zhang, Laura V. Santos-Medina, Zachary A. Yee, Kaitlin J. Martin, Guanghua Wan, Alicia M. Bolt, Xixi Zhou, Miroslav Stýblo, and Ke Jian Liu. 2023. "Arsenite Methyltransferase Is an Important Mediator of Hematotoxicity Induced by Arsenic in Drinking Water" Water 15, no. 3: 448. https://doi.org/10.3390/w15030448