
Environmental Factors Controlling Zooplankton Communities in Thermokarst Lakes of the Bolshezemelskaya Tundra Permafrost Peatlands (NE Europe)

Abstract
:1. Introduction
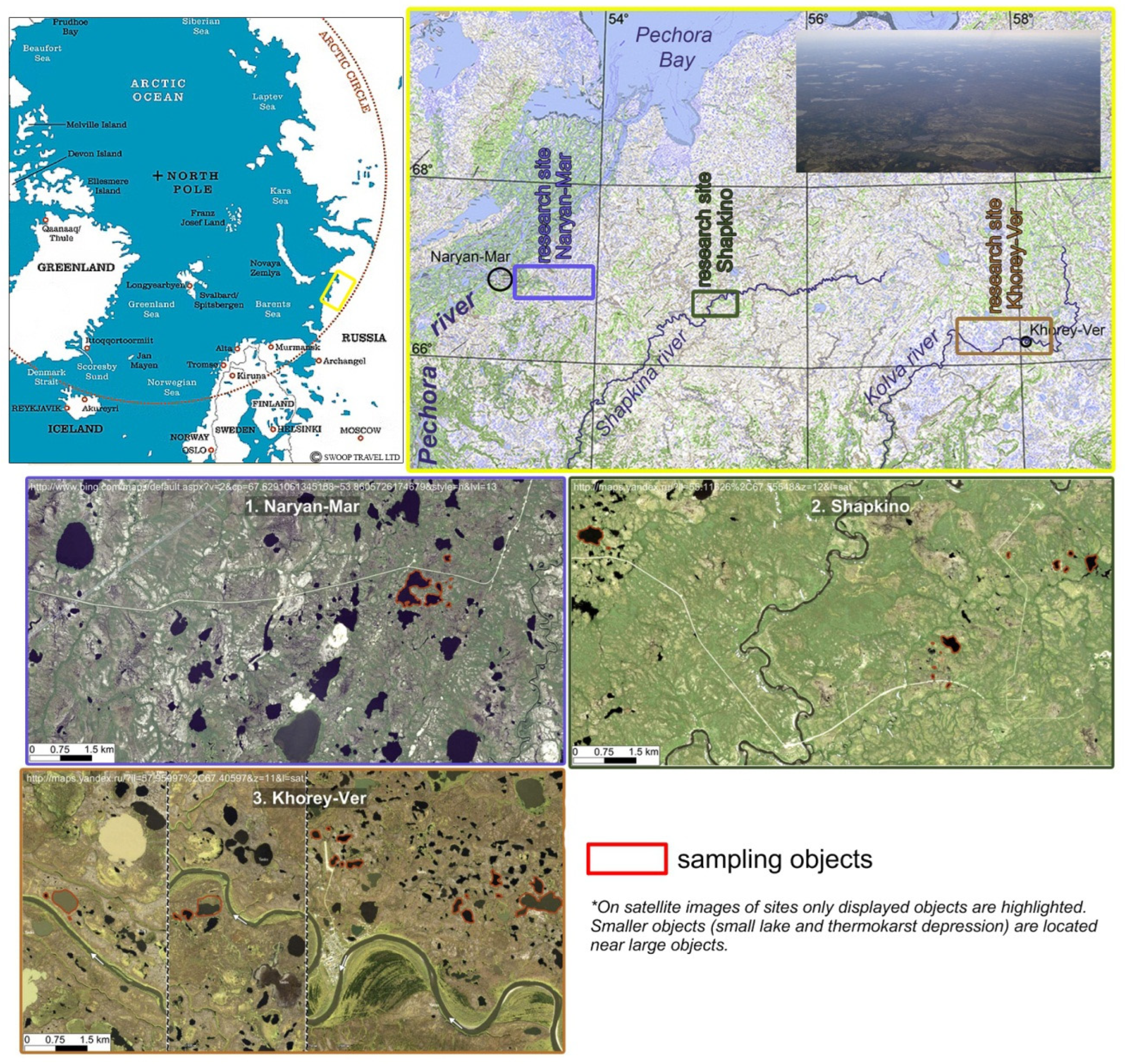
2. Materials and Methods
3. Results
3.1. Morphometric and Hydrochemical Indicators of the Studied Lakes
3.2. The Biodiversity of Zooplankton
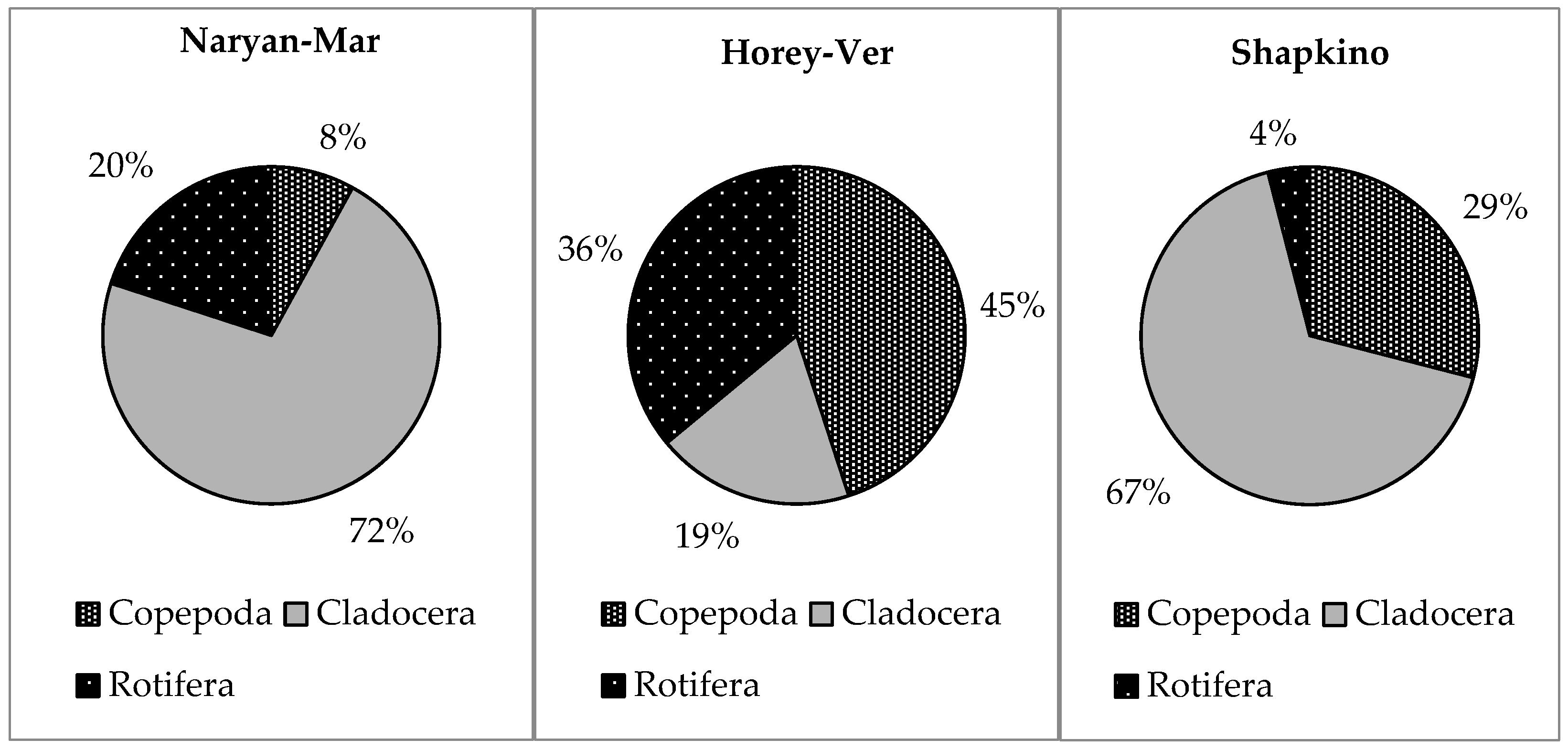
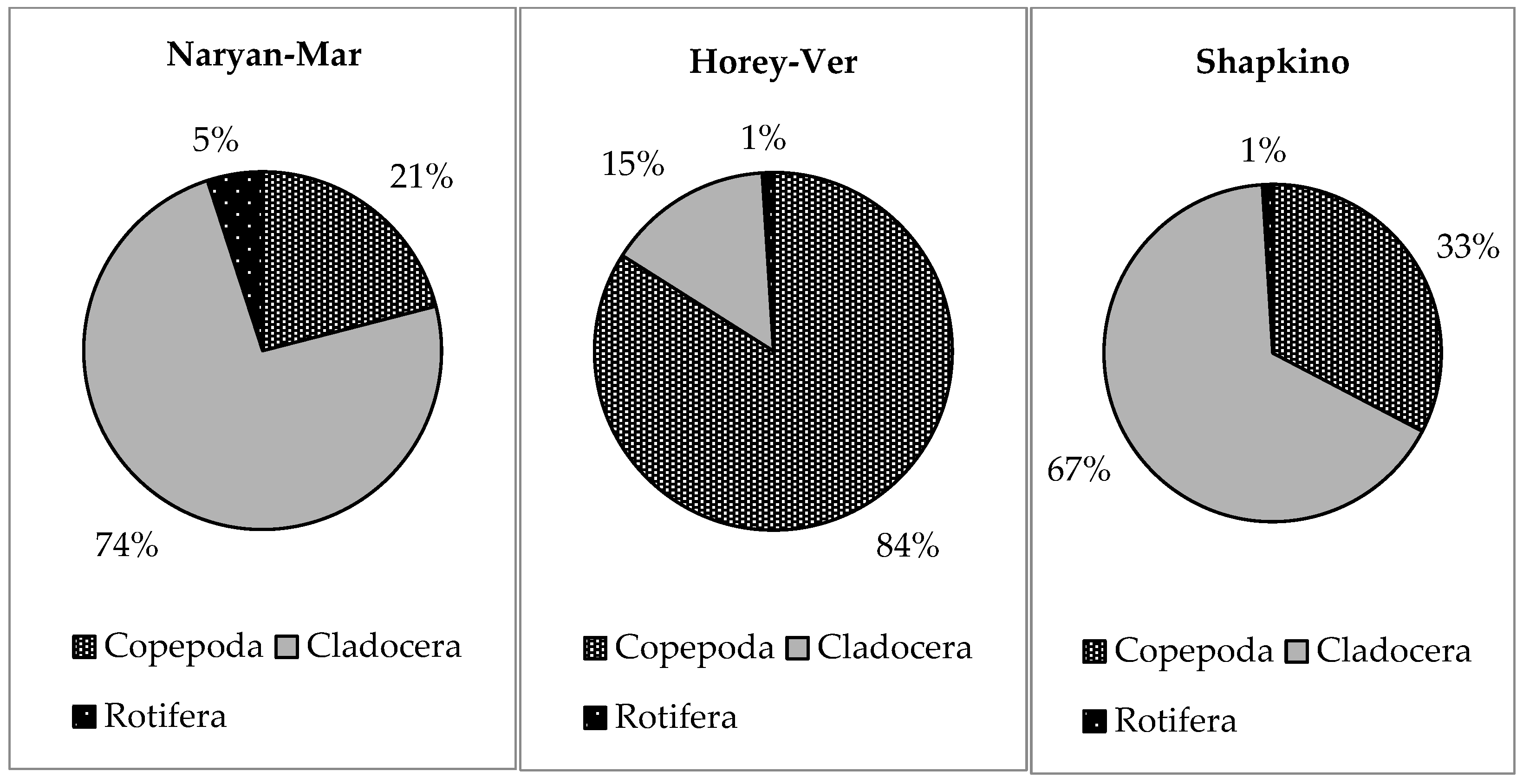
3.3. Trophic Structure
3.4. Quantitative Characteristics of Zooplankton
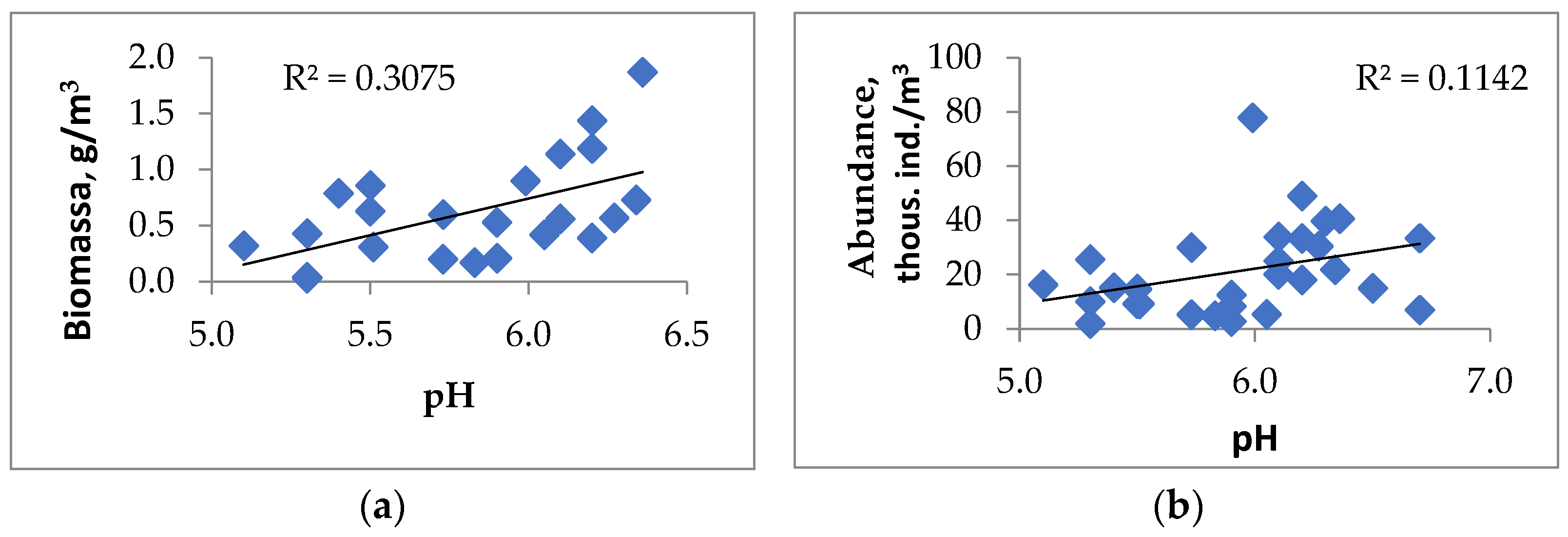
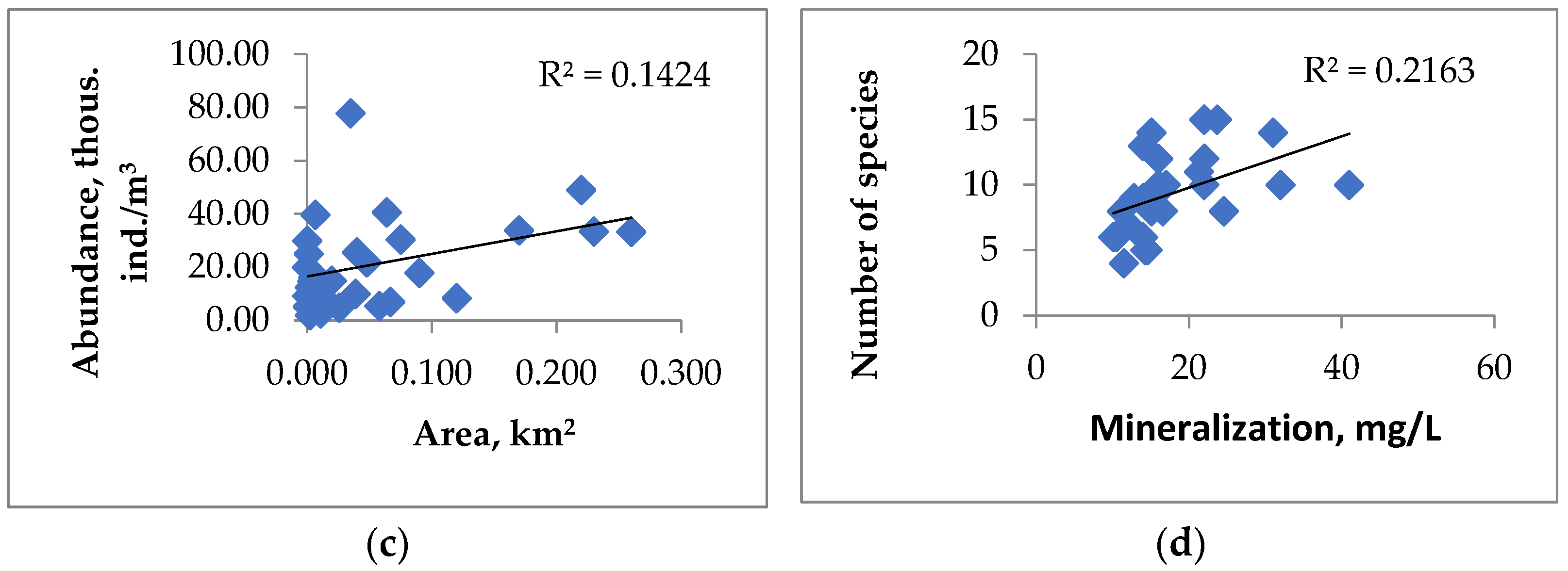
3.5. Assessment of the Influence of Abiotic and Biotic Factors on the Qualitative and Quantitative Characteristics of Zooplankton Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Culp, J.M.; Goedkoop, W.; Lento, J.; Christoffersen, K.S.; Frenzel, S.; Gudbergsson, G.; Liljaniemi, P.; Sandøy, S.; Svoboda, M.; Brittain, J.; et al. The Arctic Freshwater Biodiversity Monitoring Plan; CAFF Monitoring Series Report No. 7; CAFF International Secretariat: Akureyri, Iceland, 2012; ISBN 978-9935-431-19-6. [Google Scholar] [CrossRef]
- Tishkov, A.A. “Arctic vector” in the conservation of terrestrial ecosystems and biodiversity. Arct. Ecol. Econ. 2012, 2, 28–43. (In Russian) [Google Scholar]
- Vadadi-Fulop, C.; Sipkay, C.; Meszaros, G.; Hufnagel, L. Climate change and freshwater zooplankton: What does it boil down to? Aquat. Ecol. 2012, 46, 501–519. [Google Scholar] [CrossRef] [Green Version]
- Callaghan, T.V.; Jonasson, S. Arctic terrestrial ecosystems and environmental change. Philos. Trans. R. Soc. Lond. 1995, 352, 259–276. [Google Scholar] [CrossRef]
- Magnuson, J.J.; Robertson, D.M.; Benson, B.J.; Wynne, R.H.; Livingstone, D.M.; Arai, T.; Assel, R.A.; Barry, R.G.; Card, V.; Kuusisto, E.; et al. Historical Trends in Lake and River Ice Cover in the Northern Hemisphere. Science 2000, 289, 1743–1746. [Google Scholar] [CrossRef] [Green Version]
- Iglovsky, S.A.; Shvartsman, Y.G.; Bolotov, I.N. Kriolitozona Dvinsko-Mezen’skoi Ravniny i Poluostrova Kanin; IEPS UrO RAN: Arkhangelsk, Russia, 2010; p. 124. (In Russian) [Google Scholar]
- Rautio, M. Community structure of crustacean zooplankton in subarctic ponds—Effects of altitude and physical heterogeneity. Ecography 1988, 21, 327–335. [Google Scholar] [CrossRef]
- Rautio, M.; Dufresne, F.; Lauion, I.; Bonilla, S.; Vincent, W.F.; Christoffersen, K. Shallow freshwater ecosystems of the circumpolar Arctic. Ecoscience 2011, 18, 204–222. [Google Scholar] [CrossRef]
- Frolova, L.A.; Nazarova, L.B.; Pestryakov, L.A.; Hartsook, W. Analysis of the influence of climate-dependent factors on the formation of zooplankton communities in the Arctic lakes of the Anabar River basin. Sib. Ecol. J. 2013, 1, 3–15. (In Russian) [Google Scholar]
- Fefilova, E.B.; Kononova, O.N.; Dubovskaya, O.P.; Khokhlova, L.G. Current state of zooplankton of the Bolshezemelskaya tundra lake system. Biol. Inland Waters 2012, 4, 44–52. [Google Scholar] [CrossRef]
- Hessen, D.O.; Bakkestuen, V.; Walseng, B. Energy input and zooplankton species richness. Ecography 2007, 30, 749–758. [Google Scholar] [CrossRef]
- Sweetman, J.N.; Ruhland, K.M.; Smol, J.P. Environmental and spatial factors influencing the distribution of cladocerans in lakes across the central Canadian Arctic treeline region. J. Limnol. 2010, 69, 76–87. [Google Scholar] [CrossRef]
- Verbitskii, V.B.; Verbitskaya, T.; Kurbatova, S. Reactions of zooplankton on temperature effects. I. Influence of the nonperiodic temperature changes on the population dynamics of Cladocera. Trans. Papanin Inst. Biol. Inland Waters Russ. Acad. Sci. 2017, 78, 14–36. (In Russian) [Google Scholar]
- Korhola, A. Distribution patterns of Cladocera in Subarctic Fennoscandian lakes and their potential in environmental reconstruction. Ecography 1999, 22, 357–373. [Google Scholar] [CrossRef]
- Feniova, I.Y.; Razlutsky, V.I.; Palash, A.L. Temperature effects of interspecies competition between cladoceran species in experimental conditions. Biol. Inland Waters 2011, 1, 71–78. [Google Scholar] [CrossRef]
- Rautio, M.; Korhola, A. Effects of ultraviolet radiation and dissolved organic carbon on the survival of subarctic zooplankton. Polar Biol. 2002, 25, 460–468. [Google Scholar] [CrossRef]
- Adrian, R.; Reilly, C.O.; Zagarese, H.; Baines, S.; Hessen, D.O.; Keller, W.; Livingstone, D.; Sommaruga, R.; Straile, D.; Van Donk, E.; et al. Lakes as sentinels of climate change. Limnol. Oceanogr. 2009, 54, 2283–2297. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, E.; Davidson, T.A.; Meerhoff, M.; Trolle, D. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef] [Green Version]
- Carosi, A.; Ghetti, L.; Lorenzoni, M. The Role of Climate Changes in the Spread of Freshwater Fishes: Implications for Alien Cool and Warm-Water Species in a Mediterranean Basin. Water 2021, 13, 347. [Google Scholar] [CrossRef]
- Vekhov, N.V. Zooplankton of small lakes in the eastern part of the Bolshezemelskaya tundra. Sci. Rep. High. School. Biol. Sci. 1974, 2, 7–13. (In Russian) [Google Scholar]
- Vekhov, N.V. Zooplankton of lakes of the Bolshezemelskaya tundra. Zool. J. 1975, 54, 181–187. (In Russian) [Google Scholar]
- Hebert, P.D.N.; Hann, B.J. Patterns in the composition of Arctic tundra pond microcrustacean communities. Can. J. Fish. Aquat. Sci. 1986, 43, 1416–1425. [Google Scholar] [CrossRef]
- Baranovskaya, V.K. Crustacea (Systematic list of invertebrates from reservoirs of the Bolshezemelskaya tundra). In Flora and Fauna of Reservoirs of the European North; Getsen, M.V., Ed.; Nauka: Leningrad, Russia, 1978; pp. 174–177. (In Russian) [Google Scholar]
- Chengalath, R.; Koste, W. Composition and distributional patterns in arctic rotifers. Hydrobiologia 1989, 186, 191–200. [Google Scholar] [CrossRef]
- Dubovskaya, O.P.; Kotov, A.A.; Korvchinsky, N.N.; Smirnov, N.N.; Sinev, A.Y. Zooplankton of lakes of the spurs of the Putorana plateau and adjacent territories (north of the Krasnoyarsk region). Sib. Ecol. J. 2010, 4, 571–608. (In Russian) [Google Scholar]
- Kononova, O.N.; Dubovskaya, O.P.; Fefilova, E.B. Zooplankton and dead zooplankton in Kharbeyskie lakes of Bolshezemelskay tundra (period from 2009 to 2012). J. Sib. Fed. Univ. Biol. 2014, 3, 303–327. [Google Scholar] [CrossRef]
- De Smet, W.H.; Van Rompu, E.A.; Beyens, L. Contribution to the rotifer fauna of subarctic Greenland (Kangerlussuaq and Ammassalik area). Hydrobiologia 1993, 255/256, 463–466. [Google Scholar] [CrossRef]
- De Smet, W.H.; Beyens, L. Rotifers from the Canadian High Arctic (Devon Island, Northwest Territories). Hydrobiologia 1995, 313, 29–34. [Google Scholar] [CrossRef]
- Starobogatov, Y.I. Mollusk Fauna and Zoogeographic Zoning of Continental Reservoirs of the Globe; Nauka: Leningrad, Russia, 1970; p. 372. (In Russian) [Google Scholar]
- Flint, R.F. The Earth and Its History; Norton: New York, NY, USA, 1973; p. 359. [Google Scholar]
- Abdurakhmanov, G.M.; Krivolutsky, D.A.; Myalo, E.G.; Ogureeva, G.N. Biogeography; Higher Education; Akademiya: Moscow, Russia, 2003; p. 480. (In Russian) [Google Scholar]
- Kozhevnikov, Y.P.; Zheleznov-Chukotsky, N.K. Paleobiogeography of “Beringia” as it is Scientific Dialogue. Nat. Sci. 2014, 1, 30–83. (In Russian) [Google Scholar]
- Hopkins, D.M. The Paleogeography and Climatic History of Beringia during Late Cenozoic Time; Inter Nord: Paris, France, 1972; Volume 12, pp. 121–150. [Google Scholar]
- Pewe, T.L. Quaternary Geology of Alaska; U.S. Geological Survey: Reston, VA, USA, 1975; p. 145. [Google Scholar]
- Andreeva, S.M. Zyryanskoye Glaciation in the North of Middle Siberia. News Acad. Sci. USSR. Geogr. Ser. 1978, 5, 72–79. (In Russian) [Google Scholar]
- Hopkins, D.M.; Matthews, J.V., Jr.; Schweger, C.E.; Young, S.B. Paleoecology of Beringia; Academic Press: New York, NY, USA, 1982; p. 512. [Google Scholar]
- Weider, L.J.; Hobaek, A. Phylogeography and Arctic biodiversity: A review. Ann. Zool. Fenn. 2000, 37, 217–231. [Google Scholar]
- Weider, L.J.; Hobaek, A. Glacial refugia, haplotype distributions, and clonal richness of the Daphnia pulex complex in arctic Canada. Mol. Ecol. 2003, 12, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Novichkova, A.A.; Azovsky, A.I. Factors affecting regional diversity and distribution of freshwater microcrustaceans (Cladocera, Copepoda) at high latitudes. Polar Biol. 2017, 40, 185–198. [Google Scholar] [CrossRef]
- Andronikova, I.N. Structural and Functional Organization of Zooplankton in Lake Ecosystems of Different Trophic Types; Nauka: Saint Petersburg, Russia, 1996; p. 189. (In Russian) [Google Scholar]
- Sweetman, J.N.; Smol, J.P. A guide to the identification of cladoceran remains (Crustacea, Branchiopoda) in Alaskan lake sediments (with 118 figures in the text and 1 appendix). Arch. Fur Hydrobiol. 2006, 151, 353–394. [Google Scholar]
- Zvereva, O.S.; Vlasova, T.A.; Goldina, L.P. Vashutkinskie Lakes and Their History Research. In Hydrobiological Study and Fishery Development of Lakes of the Far North; Nauka: Moscow, Russia, 1966; pp. 4–21. (In Russian) [Google Scholar]
- Goldina, L.P. Geography of Lakes of the Bolshezemelskaya Tundra; Nauka: Moscow, Russia, 1972; p. 100. (In Russian) [Google Scholar]
- Vlasova, T.A.; Baranovskaya, V.K.; Getsen, M.V. Biological productivity of the Harbeyskie lakes of the Bolshezemelskaya tundra. In Production and Biological Studies of Freshwater Ecosystems; Belarusian State University: Minsk, Russia, 1973; pp. 147–163. (In Russian) [Google Scholar]
- Baranovskaya, V.K. Zooplankton of the Harbeyskie lakes of the Bolshezemelskaya tundra. In Productivity of Lakes of the Eastern Part of the Bolshezemelskaya Tundra; Vinberg, G.G., Vlasova, T.A., Eds.; Nauka: Leningrad, Russia, 1976; pp. 90–101. (In Russian) [Google Scholar]
- Kutikova, L.A. Rotatoria. In Flora and Fauna of Reservoirs of the European North (on the Example of Lakes of the Bolshezemelskaya Tundra); Getsen, M.V., Ed.; Nauka: Leningrad, Russia, 1978; pp. 48–51. (In Russian) [Google Scholar]
- Makartseva, E.S.; Prilezhaev, I.D. Zooplankton and its Products. In Features of the Structure of the Lakes of the Far North (on the Example of the Lakes of the Bolshezemelskaya Tundra); Drabkova, V.G., Trifonova, I.S., Eds.; Nauka: Saint Petersburg, Russia, 1994; pp. 146–168. (In Russian) [Google Scholar]
- Rakovskaya, E.M.; Davydova, M.I. Physical Geography of Russia; Humanit, Ed.; Part 1; Center VLADOS: Moscow, Russia, 2001; p. 288. (In Russian) [Google Scholar]
- Shamanova, I.I. Geocryological conditions of the central part of the Siberian uvalov. Izv. Akad. Nauk. SSSR. Ser. Is Geogr. 1975, 4, 109–116. (In Russian) [Google Scholar]
- Kozlov, S.A. Assessment of the stability of the hydrological environment at offshore hydrocarbon deposits in the Arctic. Neft. Delo. 2005, 2, 15–24. (In Russian) [Google Scholar]
- Smith, L.C.; Sheng, Y.; Macdonald, G.M.; Hinzman, L.D. Disappearing Arctic lakes. Science 2005, 308, 1429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zabelina, S.A.; Shirokova, L.S.; Klimov, S.I.; Chupakov, A.V.; Lim, A.G.; Polishchuk, Y.M.; Polishchuk, V.Y.; Bogdanov, A.N.; Muratov, I.N.; Guerin, F.; et al. Carbon emission from thermokarst lakes in NE European tundra. Limnol. Oceanogr. 2020, 66, S216–S230. [Google Scholar] [CrossRef]
- Papchenkov, V.G. Vegetation Cover of Reservoirs and Watercourses of the Middle Volga Region; International University of Business and New Technologies (MUBiNT): Yaroslavl, Russia, 2001; p. 213. (In Russian) [Google Scholar]
- Vinberg, G.G.; Lavrentieva, G.M. Methodological Recommendations for the Collection and Processing of Materials for Hydrobiological Studies in Freshwater Reservoirs. In Zooplankton and Its Products; Nauka Publishing House: Leningrad, Russia, 1984; p. 33. [Google Scholar]
- Alekseyev, V.R.; Tsalolikhin, S.Y. Determinant of Zooplankton and Zoobenthos of Fresh Waters of European Russia. In Zooplankton; Association of Scientific Publications of the CMC: Moscow/Saint Peterburg, Russia, 2010; p. 474. (In Russian) [Google Scholar]
- Pesenko, Y.A. Principles and Methods of Quantitative Analysis in Faunal Studies; Nauka: Moscow, Russia, 1982; p. 287. (In Russian) [Google Scholar]
- Magurran, E. Ecological Diversity and Its Measurement; Mir: Moscow, Russia, 1992; p. 184. (In Russian) [Google Scholar]
- Chuikov, Y.S. Methods of ecological analysis of the composition and structure of aquatic animal communities. Ecological classification of invertebrates found in freshwater plankton. Ecology 1981, 3, 71–77. (In Russian) [Google Scholar]
- Kitaev, S.P. Ecological Basis of Biological Productivity of Lakes of Different Natural Zones; Nauka: Moscow, Russia, 1984; p. 207. (In Russian) [Google Scholar]
- Krylov, A.V. Zooplankton of Lowland Small Rivers; Nauka: Moscow, Russia, 2005; p. 264. (In Russian) [Google Scholar]
- Lakin, G.F. Biometrics; Higher School: Moscow, Russia, 1990; p. 351. (In Russian) [Google Scholar]
- Bashneva, V.S. Statistics in Questions and Answers; Prospect Publishing House: Moscow, Russia, 2004; p. 344. (In Russian) [Google Scholar]
- Shirokova, L.S.; Chupakov, A.V.; Zabelina, S.A.; Neverova, N.V.; Payandi-Rolland1, D.; Causseraund, C.; Karlsson, J.; Pokrovsky, O.S. Humic surface waters of frozen peat bogs (permafrost zone) are highly resistant to bio and photodegradation. Biogeosciences 2019, 16, 2511–2526. [Google Scholar] [CrossRef] [Green Version]
- Derevenskaya, O.Y. Trophic structure of zooplankton in lakes of the Middle Volga region. Biol. Inland Waters 2002, 2, 46–50. (In Russian) [Google Scholar]
- Luecke, C.; O’Brien, W.J. The effect of Heterocope predation on zooplankton communities in arctic ponds. Limnol. Oceanogr. 1983, 28, 367–377. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, W.J.; Luecke, C. Zooplankton Community Structure in Arctic Ponds: Shifts Related to Pond Size. Arctic 2011, 64, 483–487. [Google Scholar] [CrossRef] [Green Version]
- Kotov, A.A.; Karabanov, D.P.; Damme, K.V. Non-Indigenous Cladocera (Crustacea: Branchiopoda): From a Few Notorious Cases to a Potential Global Faunal Mixing in Aquatic Ecosystems. Water 2022, 14, 2806. [Google Scholar] [CrossRef]
- Frenkel, S.E. Zooplankton of pelagial estuarine reservoirs of the Kamchatka river in 2009–2011. Stud. Aquat. Biol. Resour. Kamchatka North West. Part Pac. Ocean. 2013, 31, 74–88. [Google Scholar]
- Bogdanova, E.N. To the study of zooplankton of Yamal (zooplankton of the Kharasavey-Yakhi River basin, Middle Yamal). Sci. Bull. Yamal Nenets Auton. Dist. 2009, 1, 56–62. [Google Scholar]
- Streletskay, E.A. Review of the fauna of Rotatoria, Cladocera and Copepoda of the basin of the Anadyr river. Sib. Ecol. J. 2010, 4, 649–662. (In Russian) [Google Scholar] [CrossRef]
- Cherevichko, A.V. Zooplankton of different types of reservoirs of the Malozemelskaya tundra. Bull. North. Arct. Fed. Univ. Ser. Nat. Sci. 2012, 3, 66–72. (In Russian) [Google Scholar]
- Ermolaeva, N.I. Zooplankton of different types of reservoirs of the Yamal Peninsula in 2015. Sci. Bull. Yamal Nenets Auton. Dist. 2016, 2, 56–62. (In Russian) [Google Scholar]
- Semenchenko, V.P.; Razlutsky, V.I.; Guseva, Z.F.; Palash, A.L. Zooplankton of the Littoral Zone of Lakes of Different Types; Belorusskaya Navuka: Minsk, Belarus, 2013; p. 181. (In Russian) [Google Scholar]
- Choi, J.-Y.; Jeong, K.-S.; La, G.-H.; Chang, K.-H.; Joo, G.-J. The influence of aquatic macrophytes on the distribution and feeding habits of two Asplanchna species (A. priodonta and A. herrickii) in shallow wetlands in South Korea. J. Limnol. 2015, 74, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zimbalevskaya, L.N. Phytophilic Invertebrates of Lowland Rivers and Reservoirs; Naukova Dumka: Kiev, Russia, 1981; p. 216. (In Russian) [Google Scholar]
- Kurbatova, S.A.; Yershov, I.Y. The importance of aquatic plants in maintaining zooplankton diversity and abundance: An experimental approach. Regul. Mech. Biosyst. 2020, 11, 531–535. [Google Scholar] [CrossRef]
- Alimov, A.F. Elements of the Theory of Functioning of Aquatic Ecosystems; Science: Saint Petersburg, Russia, 2001; p. 147. (In Russian) [Google Scholar]
- Pennak, R.W. Regional lake typology in northern Colorado, U.S.A. Verh. Der Int. Ver. Theor. Und Angew. Limnol. 1958, 13, 264–283. [Google Scholar] [CrossRef]
- Fryer, G. Crustacean diversity in relation to the size of water bodies: Some facts and problems. Freshw. Biol. 1985, 15, 347–361. [Google Scholar] [CrossRef]
- Shirokova, L.S.; Chupakov, A.V.; Ivanova, I.S.; Moreva, O.Y.; Zabelina, S.A.; Shutskiy, N.A.; Loiko, S.V.; Pokrosky, O.S. Lichen, moss and peat control of C, nutrient and trace metal regime in lakes of permafrost peatlands. Sci. Total Environ. 2021, 782, 146737. [Google Scholar] [CrossRef] [PubMed]
- Alimov, A.F. Svyaz biological diversity in continental reservoirs with their morphometry and water mineralization. Biol. Inland Waters 2008, 1, 3–8. (In Russian) [Google Scholar]
- Aleshina, O.A.; Kozlova, L.A.; Uslamin, D.V. Zonal distribution of zooplankton in fresh lakes of western Siberia (on the example of the Tyumen region). Tyumen State Univ. Socio. Econ. Leg. Res. 2012, 12, 148–159. (In Russian) [Google Scholar]
- Fetter, G.V.; Ermolaeva, N.I. Influence of abiotic factors on the zooplankton structure of small lakes in the South of Western Siberia. Izv. AO RGO 2018, 2, 95–103. (In Russian) [Google Scholar]
- Pokrovsky, O.S.; Manasypov, R.M.; Pavlova, O.A.; Shirokova, L.S.; Vorobyev, S.N. Carbon, nutrient and metal controls on phytoplankton concentration and biodiversity in thermokarst lakes of latitudinal gradient from isolated to continuous permafrost. Sci. Total Environ. 2022, 806, 3. [Google Scholar] [CrossRef]
- Derevenskaya, O.Y.; Unkovska, E.N.; Mingazova, N.M. Zooplankton of lakes in conditions of waterlogging and acidification. Sci. Notes Kazan Univ. Ser. Nat. Sci. 2019, 161, 521–537. (In Russian) [Google Scholar]
- Lazareva, V.I. Transformation of zooplankton communities in small lakes during acidification. In Structure and Functioning of Ecosystems of Acidic Lakes; Komov, V.T., Ed.; Nauka: Saint-Petersburg, Russia, 1994; pp. 150–169. (In Russian) [Google Scholar]
- Pinel-Alloul, B.; Chemli, A.; Taranu, Z.E.; Bertolo, A. Using the Diversity, Taxonomic and Functional Attributes of a Zooplankton Community to Determine Lake Environmental Typology in the Natural Southern Boreal Lakes (Québec, Canada). Water 2022, 14, 578. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Study Area | Degree of Overgrowth of the Lake | |||
|---|---|---|---|---|---|
| District Khorey-Ver | District Shapkino | District Naryan-Mar | Lightly Overgrown | Overgrown | |
| pH | 5.8 ± 0.3 | 6.2 ± 0.3 | 5.8 ± 0.6 | 6.1 ± 0.1 | 5.6 ± 0.1 |
| Dissolved O2, mg/L | 8.0 ± 0.6 | 8.7 ± 1.5 | 8.4 ± 1.1 | 8.5 ± 0.3 | 7.9 ± 0.2 |
| Conductivity, µS | 14.1 ± 3.4 | 33.0 ± 11.4 | 20.6 ± 6.2 | 20.3 ± 3.1 | 16.1 ± 2.2 |
| Color, °Cr-Co | 133 ± 28 | 143 ± 23 | 190 ± 7.8 | 135 ± 5.7 | 178 ± 19 |
| DOC, ppm | 13.6 ± 2.4 | 19.0 ± 3.4 | 17.5 ± 7.9 | – | – |
| Fe, µg L−1 (min–max) | 204 (74–388) | 519 (141–1220) | 290 (107–706) | – | – |
| Total-P, µg L−1 (min–max) | 21.2 (6.9–64) | 18.4 (12.9–29) | 14.3 (8.8–18) | – | – |
| Total-N, µg L−1 (min–max) | 617 (263–923) | 420 (321–625) | 367 (136–488) | – | – |
| Number of lakes | 16 | 7 | 6 | 15 | 14 |
| Parameter | District Naryan-Mar (n = 6) | District Shapkino (n = 7) | District Khorey-Ver (n = 16) |
|---|---|---|---|
| Number of species | 11.3 ± 1.0 | 10.1 ± 1.0 | 7.8 ± 0.6 |
| Average (min–max) abundance, thousand ind./m3 | 22.35 (2.04–33.88) | 19.61 (7.00–39.76) | 21.58 (27.44–77.86) |
| Average biomass (min–max), g/m3 | 0.44 (0.04–0.79) | 2.94 (0.21–8.25) | 0.61 (0.03–1.87) |
| N Rot:Clad:Cop,% | 20:73:7 | 4:72:24 | 23:22:55 |
| B Rot:Clad:Cop,% | 6:71:23 | 1:74:25 | 3:19:78 |
| Nclad/Ncop | 9.10 | 2.30 | 0.43 |
| Bcycl/Bcal | 0.02 | 0.05 | 0.02 |
| B3/B2 | 0.34 | 0.08 | 0.17 |
| W, mg | 0.020 ± 0.006 | 0.130 ± 0.046 | 0.040 ± 0.005 |
| Shannon index by abundance (HN) | 2.50 ± 0.12 | 2.27 ± 0.16 | 1.92 ± 0.18 |
| Index of uniformity Pielou (I) | 0.71 ± 0.03 | 0.69 ± 0.04 | 0.62 ± 0.05 |
| Berger–Parker dominance index (Ib/p) | 2.75 ± 0.29 | 2.06 ± 0.09 | 1.96 ± 0.18 |
| Simpson index by abundance (Is) | 0.23 ± 0.02 | 0.28 ± 0.03 | 0.39 ± 0.05 |
| Parameter | Structural Indicators | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| N Cop | B Cop | N Clad | B Clad | N Rot | B Rot | Total Abundance, ind./m3 | Total Biomass, g/m3 | Number of Species | |
| Area (S) | 0.13 | 0.13 | −0.01 | −0.07 | 0.40 | 0.29 | 0.42 | 0.12 | −0.13 |
| Water temperature | 0.14 | 0.04 | −0.21 | −0.30 | 0.03 | 0.10 | −0.08 | −0.19 | −0.11 |
| pH | 0.25 | 0.30 | 0.31 | 0.37 | −0.05 | −0.14 | 0.44 | 0.5 | 0.00 |
| Dissolved O2 | 0.00 | 0.04 | 0.11 | 0.10 | −0.08 | −0.11 | 0.13 | 0.05 | 0.11 |
| Mineralization | 0.29 | 0.15 | 0.65 | 0.70 | −0.34 | −0.27 | 0.04 | 0.25 | 0.45 |
| Color Cr-Co | 0.33 | 0.26 | 0.11 | 0.14 | −0.10 | −0.13 | −0.15 | −0.14 | 0.06 |
| Parameter | The Dominant Species of Zooplankton | |||||
|---|---|---|---|---|---|---|
| E. gracilis | P. pediculus | D. middendorffiana | B. longispina | C. unicornis | K. longispina | |
| Area (S) | 0.34 | −0.26 | −0.27 | 0.03 | 0.41 | 0.15 |
| pH | 0.11 | 0.18 | 0.60 | 0.57 | 0.1 | −0.34 |
| Mineralization | −0.21 | 0.70 | 0.56 | 0.67 | −0.37 | −0.14 |
| Water temperature | 0.24 | −0.45 | 0.25 | 0.02 | 0.04 | 0.06 |
| E. gracilis | −0.56 | −0.09 | −0.28 | 0.27 | 0.12 | |
| P. pediculus | 0.15 | 0.55 | −0.25 | −0.09 | ||
| D. middendorffiana | 0.56 | 0.00 | −0.15 | |||
| B. longispina | −0.28 | −0.05 | ||||
| C. unicornis | 0.06 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobko, E.I.; Shirokova, L.S.; Klimov, S.I.; Chupakov, A.V.; Zabelina, S.A.; Shorina, N.V.; Moreva, O.Y.; Chupakova, A.A.; Vorobieva, T.Y. Environmental Factors Controlling Zooplankton Communities in Thermokarst Lakes of the Bolshezemelskaya Tundra Permafrost Peatlands (NE Europe). Water 2023, 15, 511. https://doi.org/10.3390/w15030511
Sobko EI, Shirokova LS, Klimov SI, Chupakov AV, Zabelina SA, Shorina NV, Moreva OY, Chupakova AA, Vorobieva TY. Environmental Factors Controlling Zooplankton Communities in Thermokarst Lakes of the Bolshezemelskaya Tundra Permafrost Peatlands (NE Europe). Water. 2023; 15(3):511. https://doi.org/10.3390/w15030511
Chicago/Turabian StyleSobko, Elena I., Liudmila S. Shirokova, Sergey I. Klimov, Artem V. Chupakov, Svetlana A. Zabelina, Natalia V. Shorina, Olga Yu. Moreva, Anna A. Chupakova, and Taissia Ya. Vorobieva. 2023. "Environmental Factors Controlling Zooplankton Communities in Thermokarst Lakes of the Bolshezemelskaya Tundra Permafrost Peatlands (NE Europe)" Water 15, no. 3: 511. https://doi.org/10.3390/w15030511