

Natural Regeneration of Morus alba in Robinia pseudoacacia Plantation and the Mechanism of Seed Germination and Early Seedling Growth Restriction in the Yellow River Delta
 and
and
Abstract
:1. Introduction
2. Materials and Methods
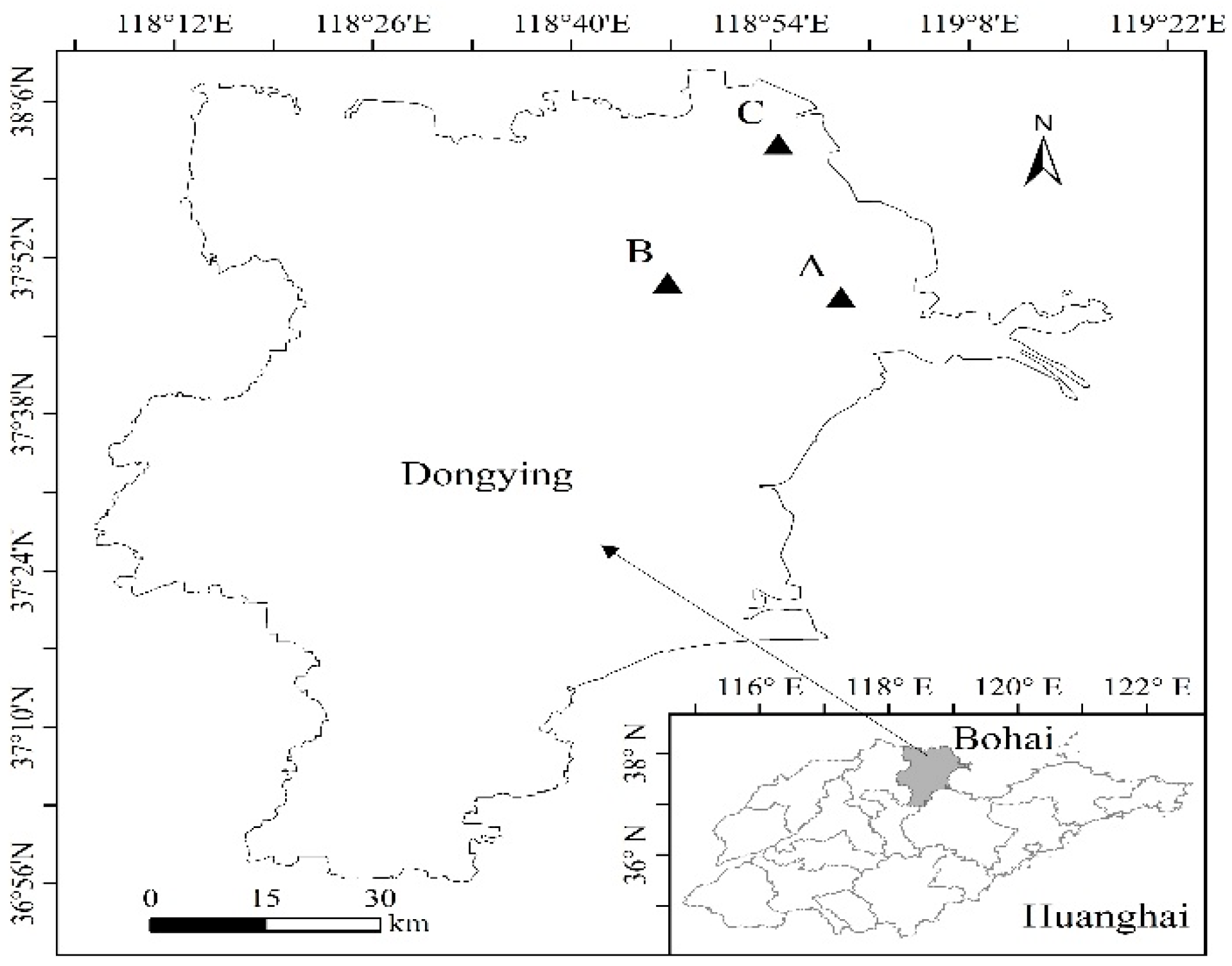
2.1. Study Area
2.2. Experimental Design
2.2.1. Survey of R. pseudoacacia Planted Forests
2.2.2. Collection of M. alba Seeds
2.2.3. Seed Germination and Seedling Growth Tests with Different Soil Salt Content, Light Intensity, and Maternal Tree Habitat
2.3. Data Analysis
3. Results
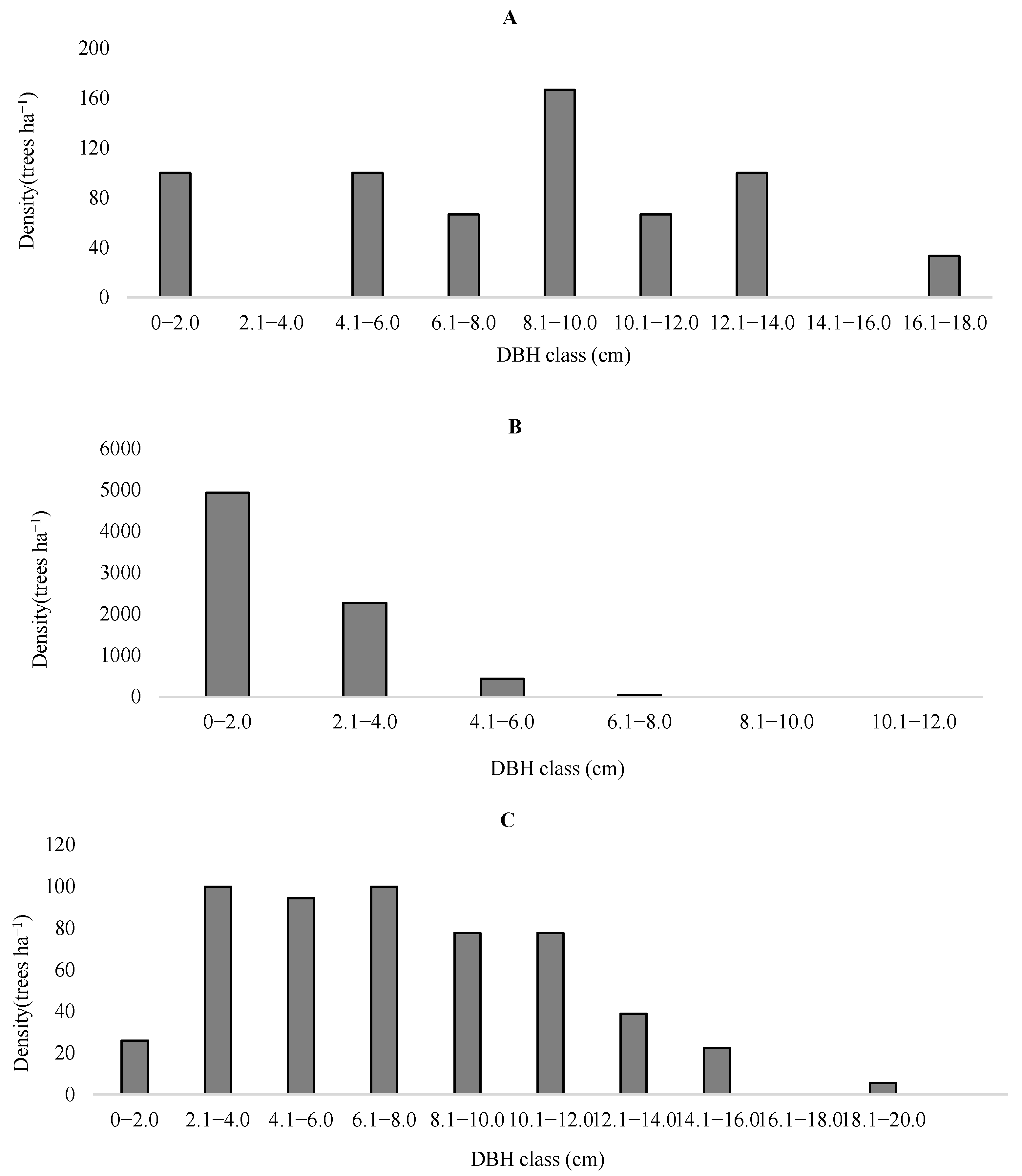
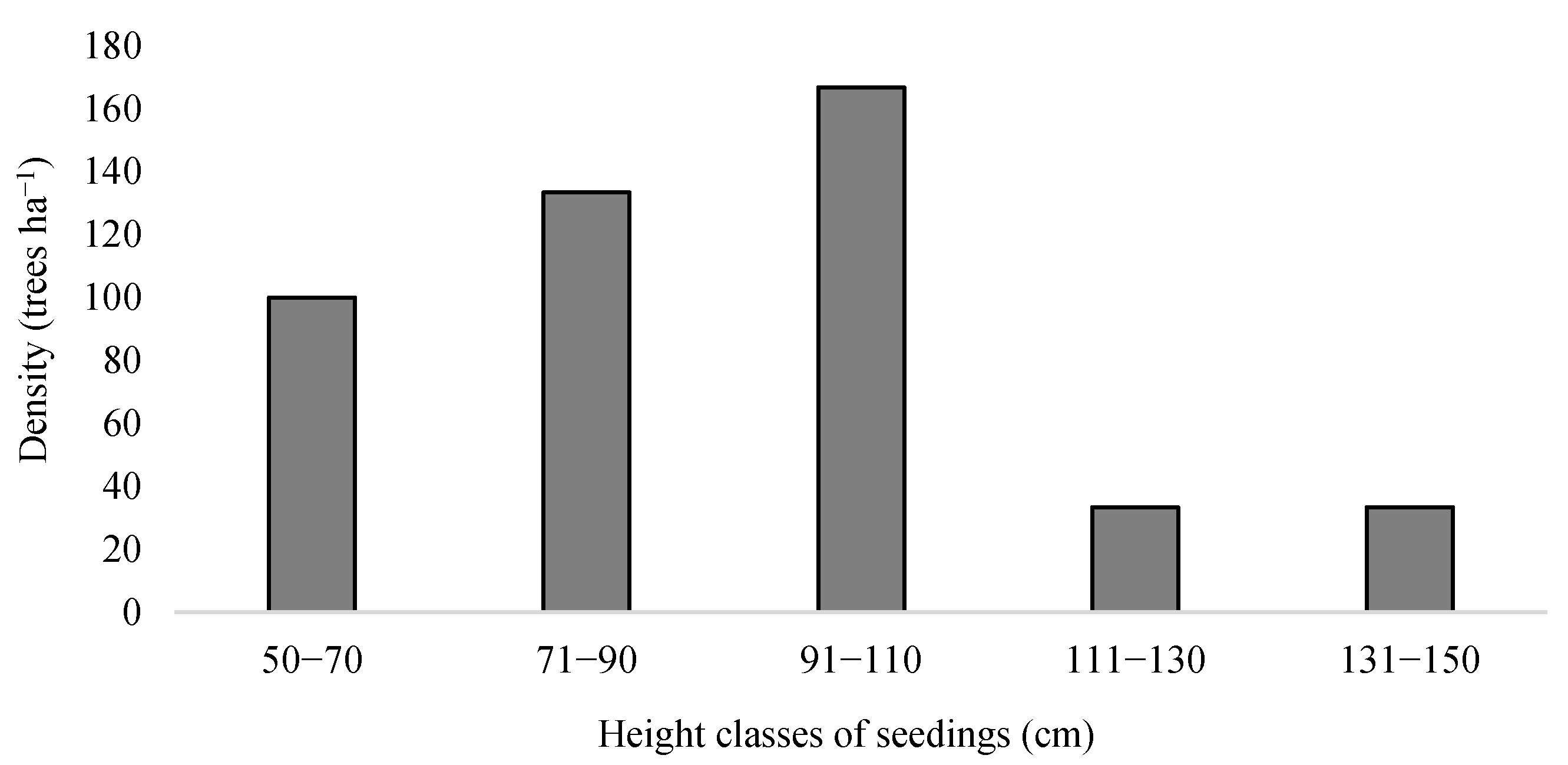
3.1. Structural Characteristics of M. alba in the R. pseudoacacia Plantation
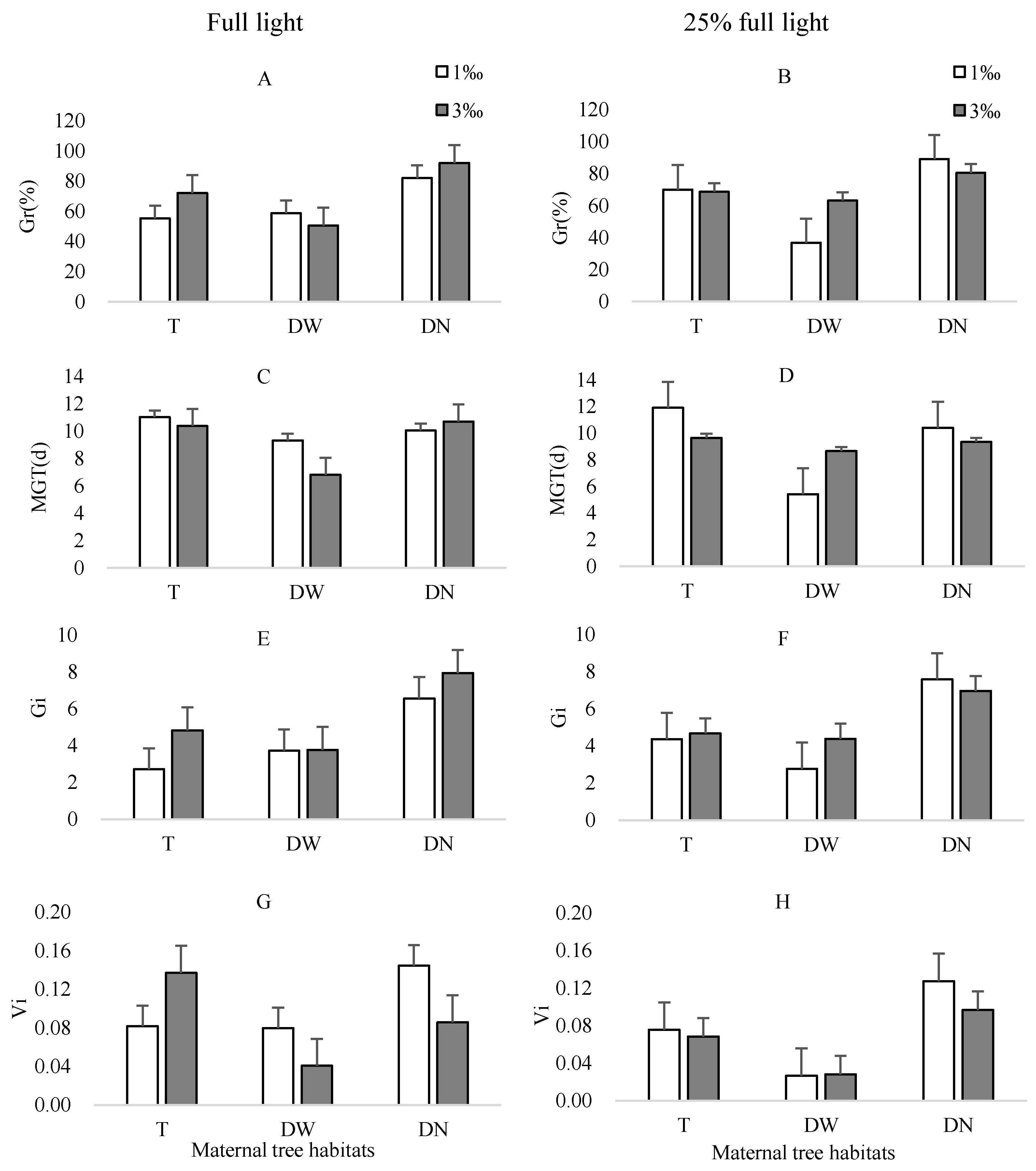
3.2. Effect of Light Intensity and Soil Salinity on Seed Germination and Seedling Growth of M. alba Seeds
3.2.1. Seed Germination
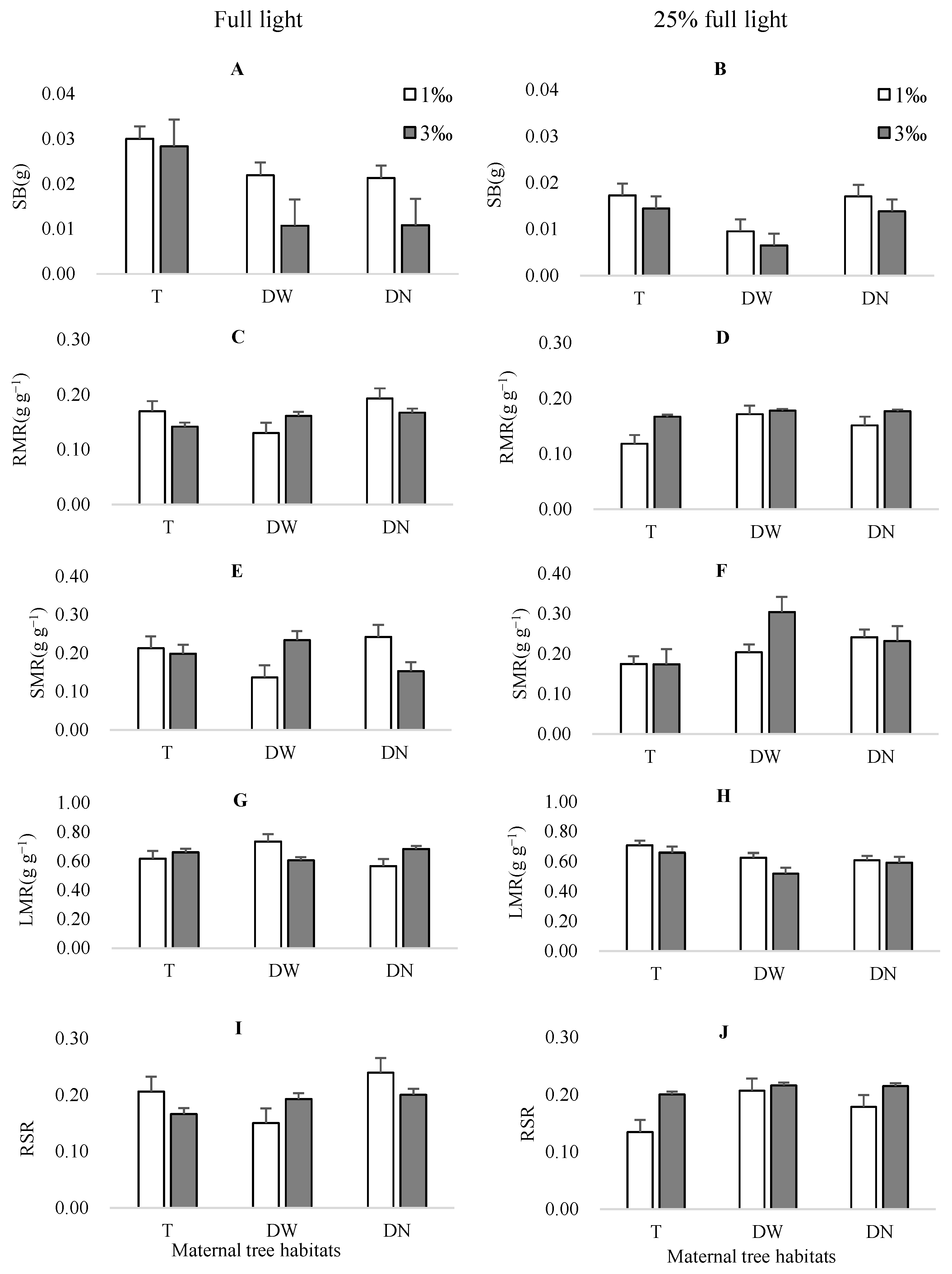
3.2.2. Seedling Biomass Traits
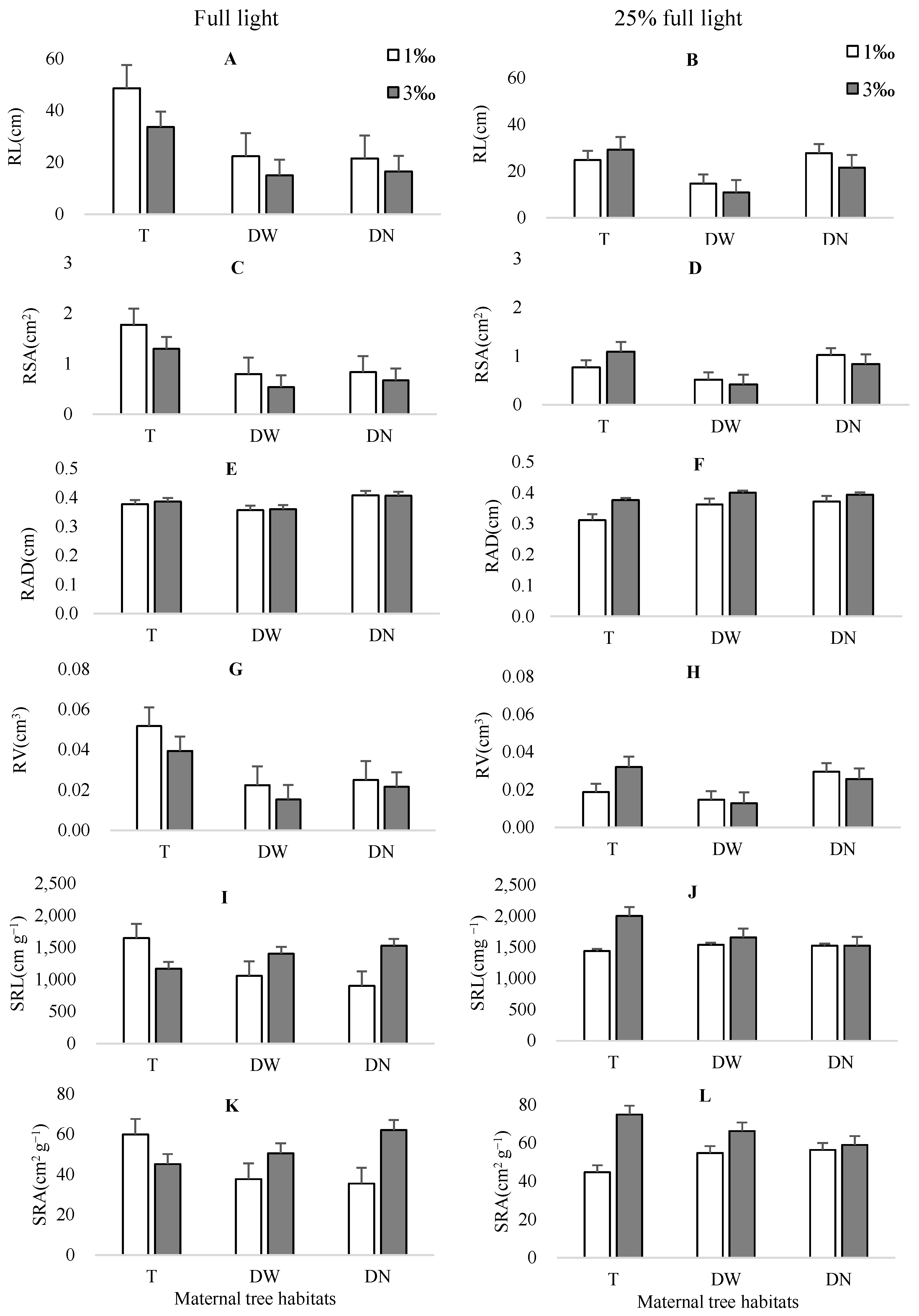
3.2.3. Seedling Root Morphology
4. Discussion
4.1. Natural Regeneration of M. alba in R. pseudoacacia Plantation in the Yellow River Delta
4.2. Comparison of M. alba Seeds in Saline Alkali Land and Nonsaline Alkali Land during Seed Germination and Early Seedling Growth
4.3. Effect of Light Intensity and Soil Salinity on Seed Germination and Seedling Growth of M. alba
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Payn, T.; Carnus, J.M.; Freer-Smith, P.; Kimberley, M.; Kollert, W.; Liu, S.; Orazio, C.; Rodriguez, L.; Silva, L.N.; Wingfifield, M.J. Changes in planted forests and future global implications. For. Ecol. Manag. 2015, 352, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Yang, Y.; Wang, H. Development strategy and management countermeasures of planted forests in China: Transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services. Acta Ecol. Sin. 2018, 3, 1–10. [Google Scholar]
- Petritan, A.M.; Biris, I.A.; Merce, O.; Turcu, D.O.; Petritan, I.C. Structure and diversity of a natural temperate sessile oak (Quercus petraea L.)—European Beech (Fagus sylvatica L.) forest. For. Ecol. Manag. 2012, 280, 140–149. [Google Scholar] [CrossRef]
- Pröll, G.; Darabant, A.; Gratzer, G.; Katzensteiner, K. Unfavourable microsites, competing vegetation and browsing restrict post-disturbance tree regeneration on extreme sites in the Northern Calcareous Alps. Eur. J. For. Res. 2015, 134, 293–308. [Google Scholar] [CrossRef]
- Valladares, F.; Niinemets, Ü. Shade tolerance, a key plant feature of complex nature and consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Laanisto, L.; Niinemets, Ü.; Zavala, M.A. Shedding light on shade: Ecological perspectives of understorey plant life. Plant Ecol. Divers. 2016, 9, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Szwagrzyk, J.; Szewczyk, J.; Bodziarczyk, J. Dynamics of seedling banks in beech forest: Results of a 10-year study on germination, growth and survival. For. Ecol. Manag. 2001, 141, 237–250. [Google Scholar] [CrossRef]
- Guo, L.; Ni, R.; Kan, X.; Lin, Q.; Mao, P.; Cao, B.; Gao, P.; Dong, J.; Mi, W.; Zhao, B. Effects of Precipitation and Soil Moisture on the Characteristics of the Seedling Bank under Quercus acutissima Forest Plantation in Mount Tai, China. Forests 2022, 13, 545. [Google Scholar] [CrossRef]
- Madsen, P.; Hahn, K. Natural regeneration in a beech-dominated forest managed by close-to-nature principles—A gap cutting based experiment. Can. J. For. Res. 2008, 38, 1716–1729. [Google Scholar] [CrossRef]
- Agestam, E.; Ekö, P.M.; Nilsson, U.; Welander, N.T. The effects of shelterwood density and site preparation on natural regeneration of Fagus sylvatica in southern Sweden. For. Ecol. Manag. 2003, 176, 61–73. [Google Scholar] [CrossRef]
- González-Rodríguez, V.; Villar, R.; Navarro-Cerrillo, R.M. Maternal influences on seed mass effect and initial seedling growth in four Quercus species. Acta Oecol. 2011, 37, 1–9. [Google Scholar] [CrossRef]
- Domic, A.I.; Capriles, J.M.; Camilo, G.R. Evaluating the fitness effects of seed size and maternal tree size on Polylepis tomentella (Rosaceae) seed germination and seedling performance. J. Trop. Ecol. 2020, 36, 115–122. [Google Scholar] [CrossRef]
- Mousseau, T.A.; Fox, C.W. The adaptive significance of maternal effects. Trends Ecol. Evol. 1998, 13, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Yuan, Z.; Wei, Z.; Hu, X. Effect of maternal environment on seed germination and seed yield components of Thlaspi arvense. Ind. Crop Prod. 2022, 181, 114790. [Google Scholar] [CrossRef]
- Adji, B.I.; Akaffou, D.S.; Reffye, P.D.; Sabatier, S. Maternal environment and seed size are important for successful germination and seedling establishment of Pterocarpus erinaceus (Fabaceae). J. For. Res. 2022, 33, 977–990. [Google Scholar] [CrossRef]
- Galloway, L.F.; Etterson, J.R. Transgenerational plasticity is adaptive in the wild. Science 2007, 318, 1134–1136. [Google Scholar] [CrossRef] [Green Version]
- Sionit, N.; Kramer, P.J. Effect of water stress during different stages of growth of soybean. Agron. J. 1977, 69, 274–278. [Google Scholar] [CrossRef]
- Zolfaghari, R.; Dalvand, F.; Fayyaz, P.; Solla, A. Maternal drought stress on persian oak (Quercus brantii Lindl.) affects susceptibility to single and combined drought and biotic stress in offspring. Environ. Exp. Bot. 2022, 194, 104716. [Google Scholar] [CrossRef]
- Mohamed, H.A.; Clark, J.A.; Ong, C.K. The influence of temperature during seed development on the germination characteristics of millet seeds. Plant Cell Environ. 1985, 8, 361–362. [Google Scholar] [CrossRef]
- Herman, J.J.; Sultan, S.E.; Tim, H.K.; Riggs, C. Adaptive transgenerational plasticity in an annual plant: Grandparental and parental drought stress enhance performance of seedlings in dry soil. Integ. Comp. Biol. 2012, 52, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Herman, J.J.; Sultan, S.E. Adaptive Transgenerational pasticity in plants: Case studies, mechanisms, and implications for natural populations. Front. Plant Sci. 2011, 2, 102. [Google Scholar] [CrossRef] [PubMed]
- Yakovlev, I.; Fossdal, C.G.; Skrøppa, T.; Olsen, J.E.; Jahren, A.H.; Johnsen, O. An adaptive epigenetic memory in conifers with important implications for seed production. Seed Sci. Res. 2012, 22, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.Z.; Khan, M.A. Tolerance and recovery responses of playa halophytes to light, salinity and temperature stresses during seed germination. Flora 2010, 205, 764–771. [Google Scholar] [CrossRef]
- Wang, D.; Jia, Y.; Cui, J. Research progress on effects of salt stress on plants and salt adaptability of plants. Chin. Agric. Sci. Bull. 2009, 25, 124–128. [Google Scholar]
- Zhang, G.Y.; Tan, J.Z.; Liu, M.J. Effects of salt stress on mulberry seed germination and physiological and biochemical characteristics of seedlings. Acta Sericologica Sin. 2004, 02, 191–194. [Google Scholar]
- Zhang, T.Y.; Sheng, J.; Wang, R.Y.; Li, H.Y.; Yang, Y.F. Effects of saline alkali stress and temperature on seed germination of Artemisia selengensis. Acta Agrestia Sin. 2019, 27, 581–588. [Google Scholar]
- Cheng, L.; Li, Z.; Han, Z.; Shi, X. Response of seed germination of Aralia salina to temperature, light and salt drought stress. Pratacultural Sci. 2015, 32, 961–966. [Google Scholar]
- Sendall, K.M.; Reich, P.B.; Lusk, C.H. Size-related shifts in carbon gain and growth responses to light differ among rainforest evergreens of contrasting shade tolerance. Oecologia 2018, 187, 609–623. [Google Scholar] [CrossRef]
- Cavalcanti, F.R.; Santos Lima, J.P.M.; Ferreira-Silva, S.L.; Viegas, R.A.; Silveira, J.A.G. Roots and leaves display contrasting oxidative response during salt stress and recovery in cowpea. J. Plant Physiol. 2007, 164, 591–600. [Google Scholar] [CrossRef]
- Mao, P.; Zhang, Y.; Cao, B.; Guo, L.M.; Shao, H.; Cao, Z.; Jiang, Q.; Wang, X. Effects of salt stress on eco-physiological characteristics in Robinia pseudoacacia based on salt-soil rhizosphere. Sci. Total Environ. 2016, 568, 118–123. [Google Scholar] [CrossRef]
- Lama, G.F.C.; Crimaldi, M. Assessing the role of Gap Fraction on the Leaf Area Index (LAI) estimations of riparian vegetation based on Fisheye lenses. In Proceedings of the 29th European Biomass Conference and Exhibition, Marseille, France, 26–29 April 2021; pp. 1172–1176. [Google Scholar] [CrossRef]
- Crimaldi, M.; Lama, G.F.C. Impacts of riparian plants biomass assessed by UAV-acquired multispectral images on the hydrodynamics of vegetated streams. In Proceedings of the 29th European Biomass Conference and Exhibition, Marseille, France, 26–29 April 2021; pp. 1157–1161. [Google Scholar] [CrossRef]
- Pirone, D.; Cimorelli, L.; Del Giudice, G.; Pianese, D. Short-term rainfall forecasting using cumulative precipitation fields from station data: A probabilistic machine learning approach. J. Hydrol. 2023, 617, 128949. [Google Scholar] [CrossRef]
- Yao, L.; Liu, G.; Liu, Q.; Fei, L. Remote sensing monitoring the health of artificial Robinia pseudoacacia forest. Geomat. Inf. Sci. Wuhan Univ. 2010, 35, 863–867. [Google Scholar]
- Ke, Y. Study on Salt Resistance of Mulberry and its Application in Saline Alkali Land; China Academy of Forestry Sciences: Beijing, China, 2008. [Google Scholar]
- Cheng, J.L.; Duan, J.L.; Wang, N.; Xiao, L.Y. Effect of polyethylene glycol simulated water stress on mulberry seed germination and physiology. J. Anhui Agric. Sci. 2006, 24, 6420–6422. [Google Scholar]
- Zhang, J.; Xing, S.; Sun, Q.; Chi, J.; Song, Y. Study on flora resources and its features in Yellow River Delta region. Res. Soil Water Conserv. 2006, 13, 100–102. [Google Scholar]
- Mao, P.; Kan, X.; Pang, Y.; Ni, R.; Cao, B.; Wang, K.; Zhang, J.; Tan, C.; Geng, Y.; Cao, X.; et al. Effects of forest gap and seed size on germination and early seedling growth in Quercus acutissima plantation in Mount Tai, China. Forests 2022, 13, 1025. [Google Scholar] [CrossRef]
- Diaci, J.; Adamic, T.; Rozman, A. Gap recruitment and partitioning in an old-growth beech forest of the Dinaric Mountains: Influences of light regime, herb competition and browsing. For. Ecol. Manag. 2012, 285, 20–28. [Google Scholar] [CrossRef]
- O’Brien, M.J.; O’Hara, K.L.; Erbilgin, N.; Wood, D.L. Overstory and shrub effects on natural regeneration processes in native Pinus radiata stands. For. Ecol. Manag. 2007, 240, 178–185. [Google Scholar] [CrossRef]
- Baskin, J.M.; Lu, J.J.; Baskin, C.C.; Tan, D.Y.; Wang, L. Diaspore dispersal ability and degree of dormancy in heteromorphic species of cold deserts of northwest China: A review. Perspect. Plant Ecol. Evol. Syst. 2014, 16, 93–99. [Google Scholar] [CrossRef]
- Larios, E.; Búrquez, A.; Becerra, J.X.; Venable, D.L. Natural selection on seed size through the life cycle of a desert annual plant. Ecology 2014, 95, 3213–3220. [Google Scholar] [CrossRef] [Green Version]
- Vivas, M.; Zas, R.; Sampedro, L.; Solla, A. Environmental maternal effects mediate the resistance of maritime pine to biotic stress. PLoS ONE 2013, 8, e70148. [Google Scholar] [CrossRef] [Green Version]
- Yin, K. Effects of the Age of Mother Tree and Site Conditions on the Salt Resistant Germination of Robinia Pseudoacacia Seeds; Shandong Agricultural University: Tai’an, China, 2019. [Google Scholar]
- Mao, P.; Guo, L.; Gao, Y.; Qi, L.; Cao, B. Effects of seed size and sand burial on germination and early growth of seedlings for coastal Pinus thunbergii Parl in the Northern Shandong Peninsula, China. Forests 2019, 10, 281. [Google Scholar] [CrossRef] [Green Version]
- Quero, J.L.; Villar, R.; Marañón, T.; Zamora, R.; Poorter, L. Seed-mass effects in four Mediterranean quercus species (Fagaceae) growing in contrasting light environments. Am. J. Bot. 2007, 94, 1795–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C. Characteristics of Temporal and Spatial Variation of Water Recourse in the Yellow River Delta under Conditions of Climate Change; Nanjing University of Information Science & Technology: Nanjing, China, 2012. [Google Scholar]
- Guo, L.; Cao, B.; Mao, P.; Li, Z.; Hao, M.; Wang, T.; Jiang, F. Fine root vertical-seasonal distribution of Robinia pseudoacacia in relation to abioticfactors in a chronosequence in coastal saline alkali land of the Yellow River Delta, China. Turk. J. Agric. For. 2021, 45, 750–765. [Google Scholar] [CrossRef]
- Feng, L.; Liu, J.T.; Han, G.X.; Zhang, Q.H.; Peng, L. Effects of grounder level fluctuation on characteristics of soil seed banks in coastal wetlands of the Yellow River Delta. Acta Ecol. Sin. 2021, 41, 3826–3835. [Google Scholar]
- Huang, H.; Fu, J.; Cao, B.; Zhao, W.; Mao, P.; Wang, T.; Hao, Y.; Zhang, X.; Tan, H. Effects of salt stress and temperature on seed germination and seedling growth of mulberry. J. Southwest For. Univ. 2021, 41, 30–37. [Google Scholar]
- Poorter, L. Growth responses of 15 rainforest tree species to a light gradient: The relative importance of morphological and physiological traits. Funct. Ecol. 1999, 13, 396–410. [Google Scholar] [CrossRef]
- Bloor, J.M.G.; Grubb, P.J. Morphological plasticity of shade-tolerant tropical rainforest tree seedlings exposed to light changes. Funct. Ecol. 2004, 18, 337–348. [Google Scholar] [CrossRef]
- Rey, P.J.; Alcántara, J.M. Recruitment dynamics of a fleshy-fruited plant (Olea europaea): Connecting patterns of seed dispersal to seedling establishment. J. Ecol. 2000, 88, 622–633. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Indicators | Stand Age (Years) | Density of Standing Timber (Trees ha−1) | Average DBH (cm) | Dead Wood Density (Trees ha−1) | Soil Salinity (‰) | Soil pH |
|---|---|---|---|---|---|---|
| Jiufenchang | 19 | 1033 | 8.33 ± 4.32 | 0 | 0.19 | 8.01 |
| Shenxiangou | 43 | 900 | 19.12 ± 2.88 | 100 | 0.21 | 7.92 |
| Reserve | 31 | 0 | 0 | 1200 | 0.23 | 7.86 |
| Indicators | Density (Trees ha−1) | Chest Height Section Area (Trees ha−1) | Average Diameter at Breast Height (cm) | Seedling Density (Trees ha−1) |
|---|---|---|---|---|
| Jiufenchang | 633 | 4.35 | 8.33 ± 4.32 | No seedlings |
| Shenxiangou | 7667 | 3.04 | 2.02 ± 0.10 | 467 |
| Protected Areas | 656 | 1.57 | 8.56 ± 3.91 | 26 |
| Indicators | Light (A) | Salt (B) | Maternal Tree Habitat (C) | A × B | A × C | B × C | A × B × C | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| F | p | F | p | F | p | F | p | F | p | F | p | F | p | |
| Gr | 0.03 | >0.05 | 5.26 | <0.05 | 54.58 | <0.01 | 0.01 | >0.05 | 1.66 | >0.05 | 0.90 | >0.05 | 12.35 | <0.01 |
| MGT | 0.87 | >0.05 | 0.68 | >0.05 | 14.85 | <0.01 | 0.60 | >0.05 | 0.41 | >0.05 | 1.14 | >0.05 | 5.76 | <0.01 |
| Gi | 0.64 | >0.05 | 9.86 | <0.01 | 69.90 | <0.01 | 2.09 | >0.05 | 1.34 | >0.05 | 0.84 | >0.05 | 5.29 | <0.01 |
| Vi | 8.76 | <0.01 | 2.55 | >0.05 | 24.16 | <0.01 | 0.02 | >0.05 | 1.55 | >0.05 | 5.89 | <0.01 | 4.21 | <0.05 |
| RMR | 0.00 | >0.05 | 1.56 | >0.05 | 2.92 | >0.05 | 5.10 | <0.05 | 3.78 | <0.05 | 0.50 | >0.05 | 4.36 | <0.05 |
| SMR | 1.95 | >0.05 | 0.60 | >0.05 | 1.19 | >0.05 | 0.79 | >0.05 | 2.93 | >0.05 | 5.83 | <0.01 | 0.40 | >0.05 |
| LMR | 1.70 | >0.05 | 1.48 | >0.05 | 2.62 | >0.05 | 2.96 | >0.05 | 4.95 | <0.05 | 6.41 | <0.01 | 1.39 | >0.05 |
| RSR | 0.00 | >0.05 | 1.34 | >0.05 | 2.72 | >0.05 | 5.13 | <0.05 | 3.65 | <0.05 | 0.49 | >0.05 | 4.10 | <0.05 |
| SB | 46.61 | <0.01 | 24.72 | <0.01 | 34.83 | <0.01 | 4.86 | <0.05 | 10.96 | <0.01 | 2.32 | >0.05 | 2.06 | >0.05 |
| RL | 1.60 | >0.05 | 2.08 | >0.05 | 9.02 | <0.01 | 0.93 | >0.05 | 2.20 | >0.05 | 0.00 | >0.05 | 0.71 | >0.05 |
| RSA | 2.60 | >0.05 | 1.22 | >0.05 | 9.99 | <0.01 | 1.48 | >0.05 | 3.01 | >0.05 | 0.07 | >0.05 | 0.98 | >0.05 |
| RAD | 1.13 | >0.05 | 3.53 | >0.05 | 2.40 | >0.05 | 2.50 | >0.05 | 2.49 | >0.05 | 0.40 | >0.05 | 0.15 | >0.05 |
| RV | 3.71 | >0.05 | 0.49 | >0.05 | 10.46 | <0.01 | 2.03 | >0.05 | 3.94 | <0.05 | 0.20 | >0.05 | 1.24 | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Zhu, X.; Liu, W.; Wang, K.; Tan, C.; Liu, G.; Mao, P.; Cao, B.; Jia, B.; Pang, Y.; et al. Natural Regeneration of Morus alba in Robinia pseudoacacia Plantation and the Mechanism of Seed Germination and Early Seedling Growth Restriction in the Yellow River Delta. Water 2023, 15, 546. https://doi.org/10.3390/w15030546
Wang M, Zhu X, Liu W, Wang K, Tan C, Liu G, Mao P, Cao B, Jia B, Pang Y, et al. Natural Regeneration of Morus alba in Robinia pseudoacacia Plantation and the Mechanism of Seed Germination and Early Seedling Growth Restriction in the Yellow River Delta. Water. 2023; 15(3):546. https://doi.org/10.3390/w15030546
Chicago/Turabian StyleWang, Mengyao, Xiangyuan Zhu, Wei Liu, Kexin Wang, Chunxia Tan, Guangfang Liu, Peili Mao, Banghua Cao, Bo Jia, Yuanxiang Pang, and et al. 2023. "Natural Regeneration of Morus alba in Robinia pseudoacacia Plantation and the Mechanism of Seed Germination and Early Seedling Growth Restriction in the Yellow River Delta" Water 15, no. 3: 546. https://doi.org/10.3390/w15030546