
Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle
 , ,
, ,  ,
,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Seawater Sampling by PROTEUS
2.2. The PROTEUS Unmanned Autonomous Vehicle
2.3. Physical and Chemical Parameters
2.4. Microbial Abundances
2.4.1. Prokaryotic Cell Abundance, Volume and Biomass by Image Analysis
2.4.2. Quantification of Viable (Live/Dead) and Respiring (CTC+) Prokaryotic Cells
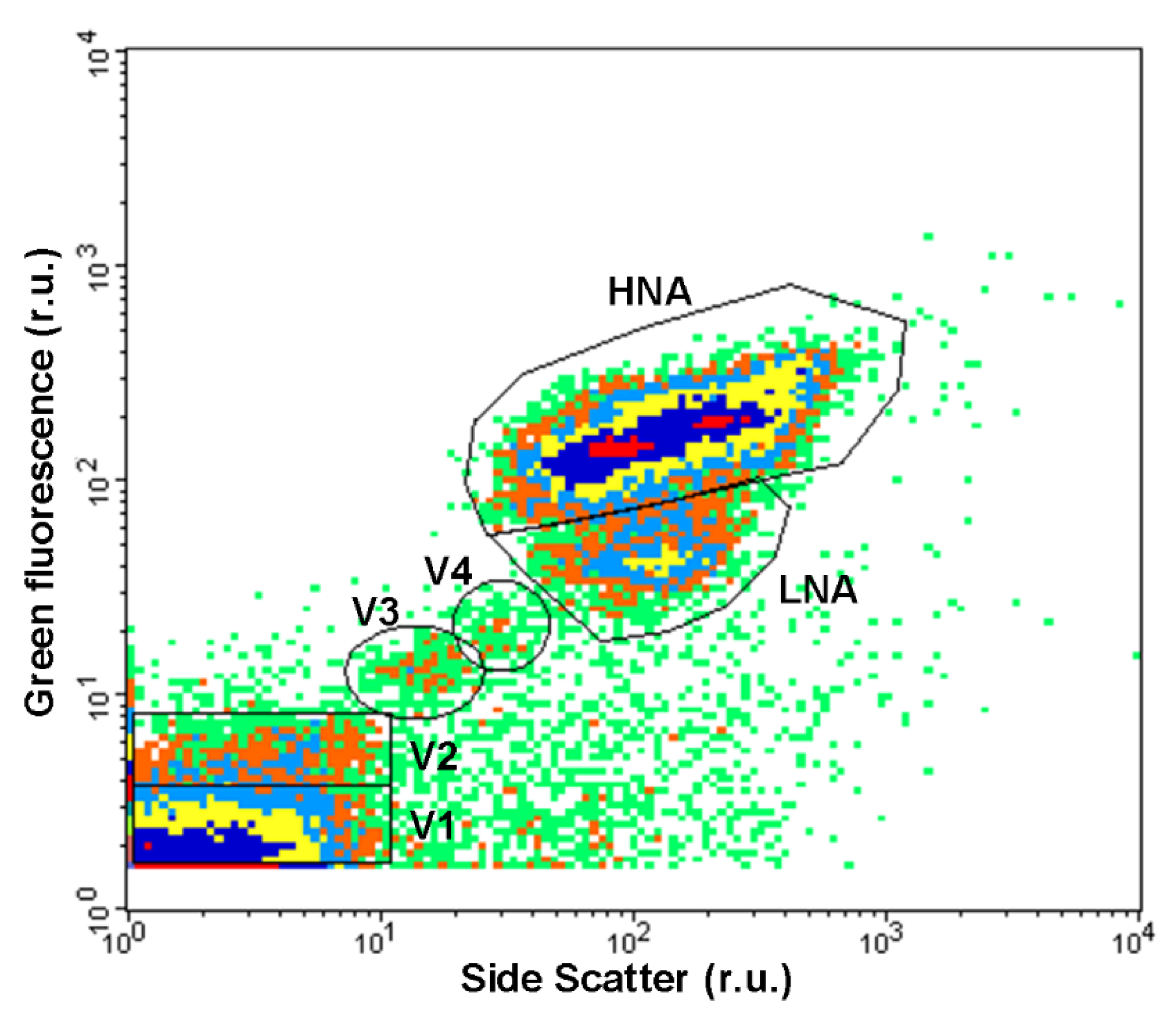
2.4.3. Virus-like Particles, Autotrophic and Prokaryotic Cells by Flow Cytometry
2.5. Culturable Heterotrophic Bacteria
2.6. Prokaryotic Community Composition
2.7. Microbial Metabolism
2.7.1. Physiological Profiles of Microbial Community
2.7.2. Enzymatic Activities
2.8. Statistical Analyses
3. Results
3.1. Environmental Parameters
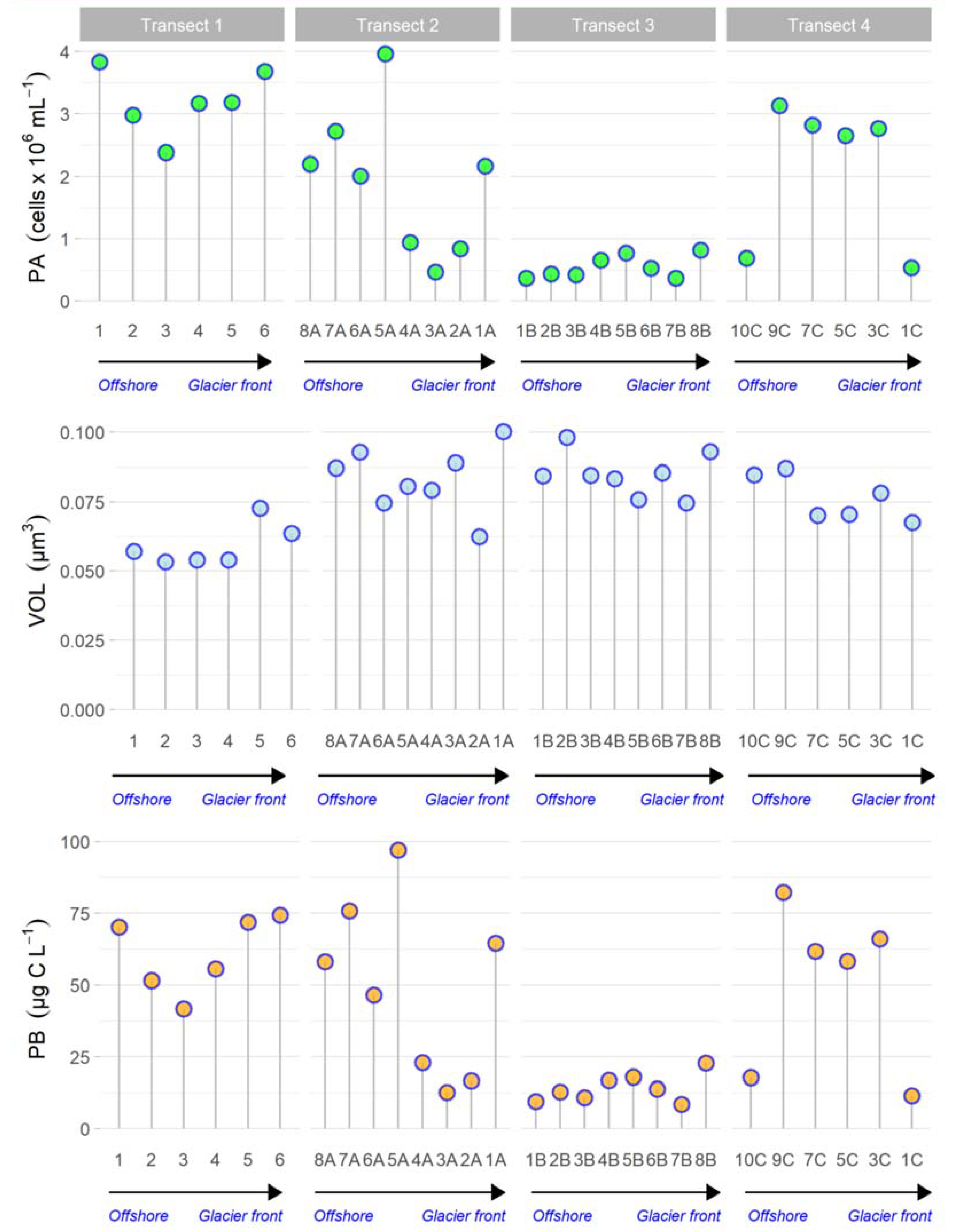
3.2. Microbial Cell Abundances
3.2.1. Prokaryotic Cell Abundance, Volume and Biomass by Image Analysis
3.2.2. Evaluation of Viable (Live/Dead) and Respiring Prokaryotic Cells (CTC+)
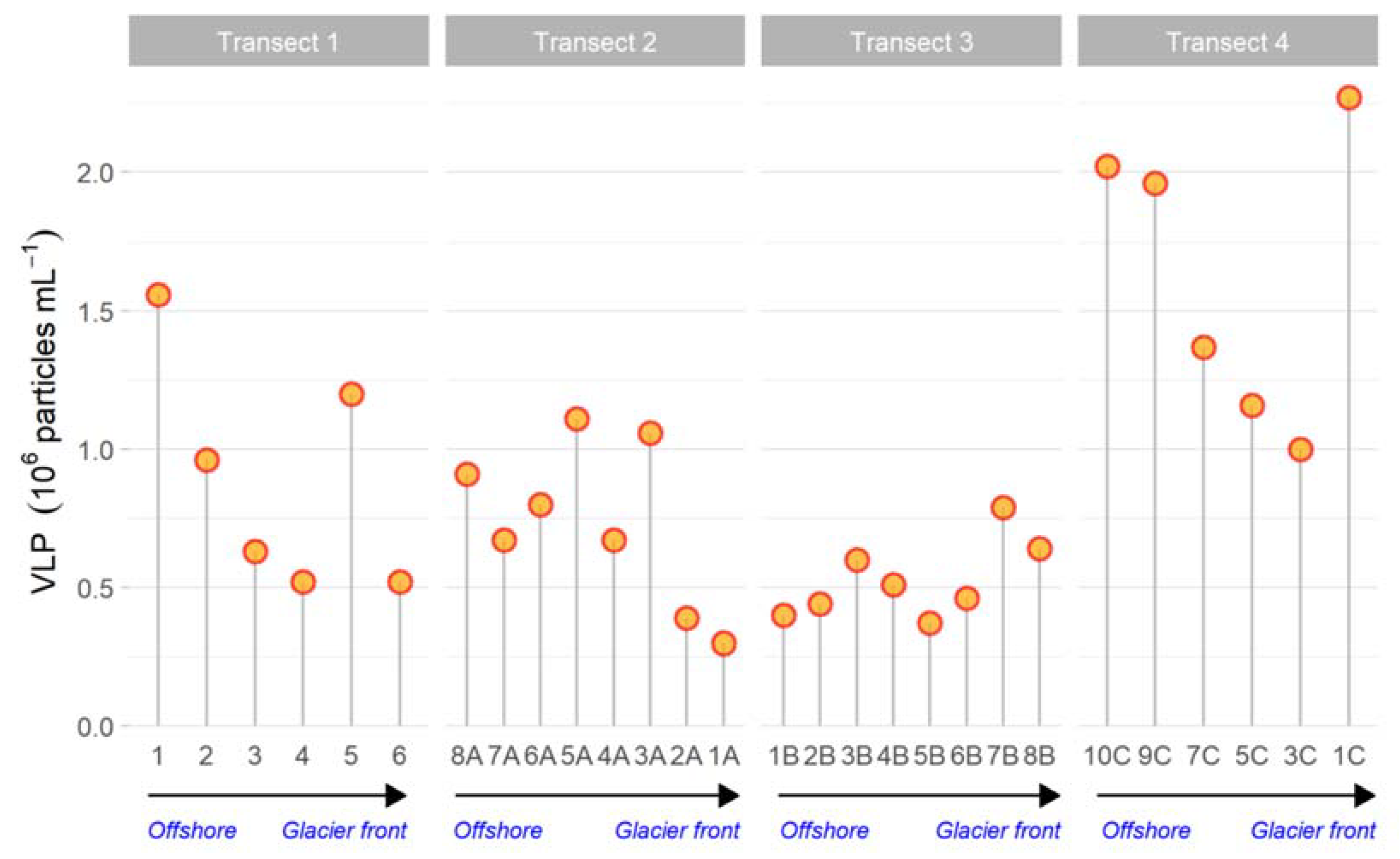
3.2.3. Virus-like Particles, Autotrophic and Prokaryotic Cells by Flow Cytometry
3.3. Culturable Heterotrophic Bacteria
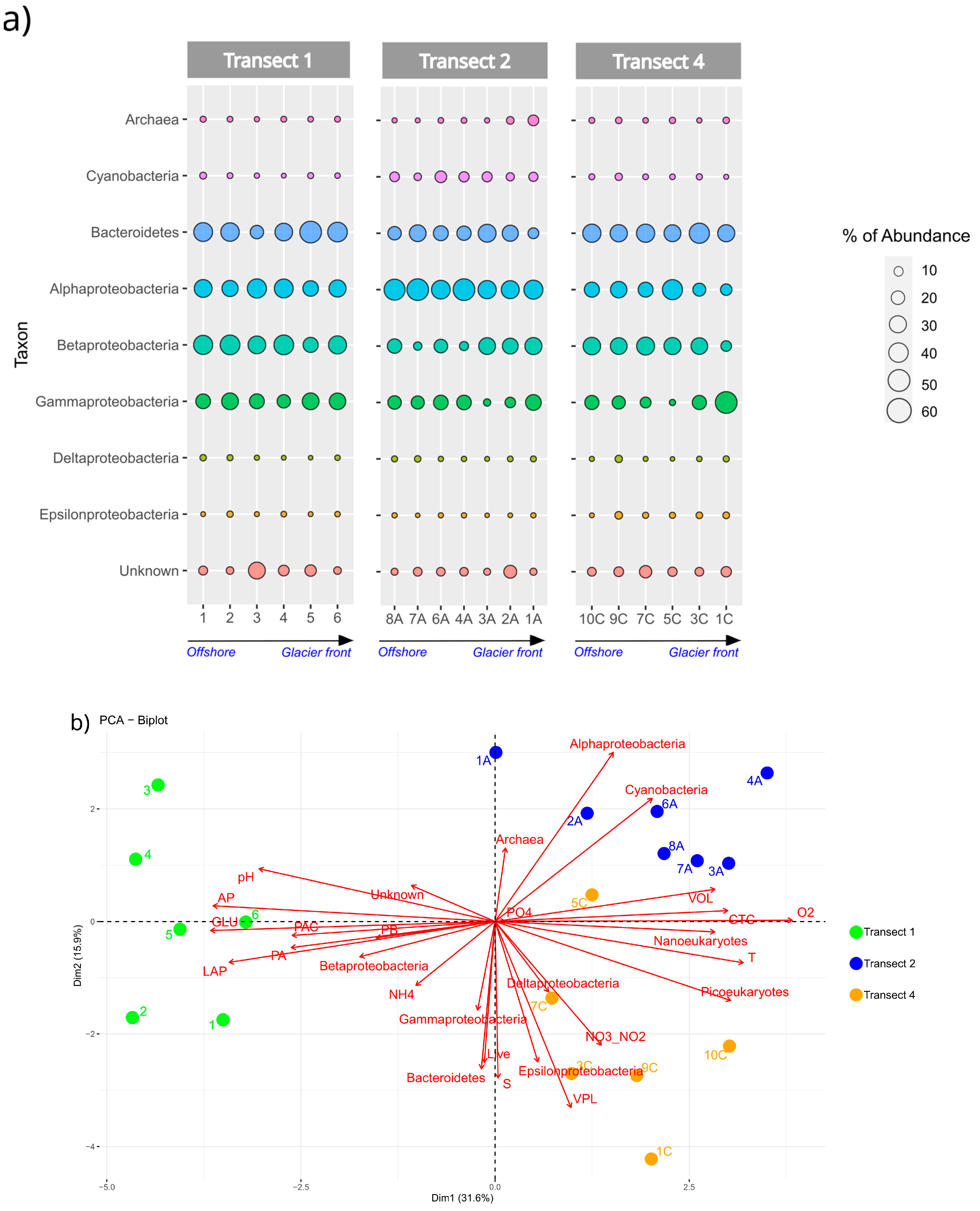
3.4. Prokaryotic Community Composition
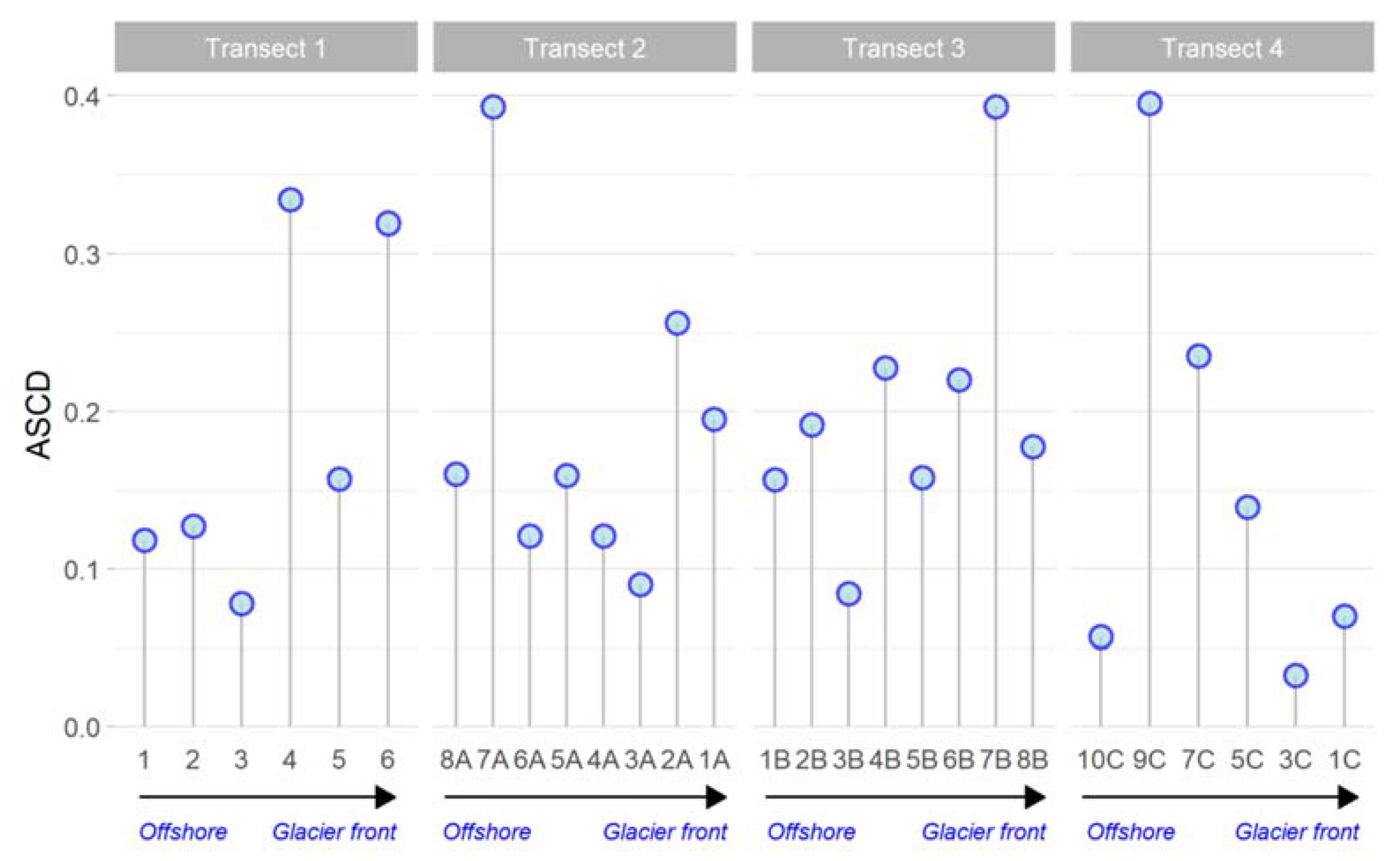
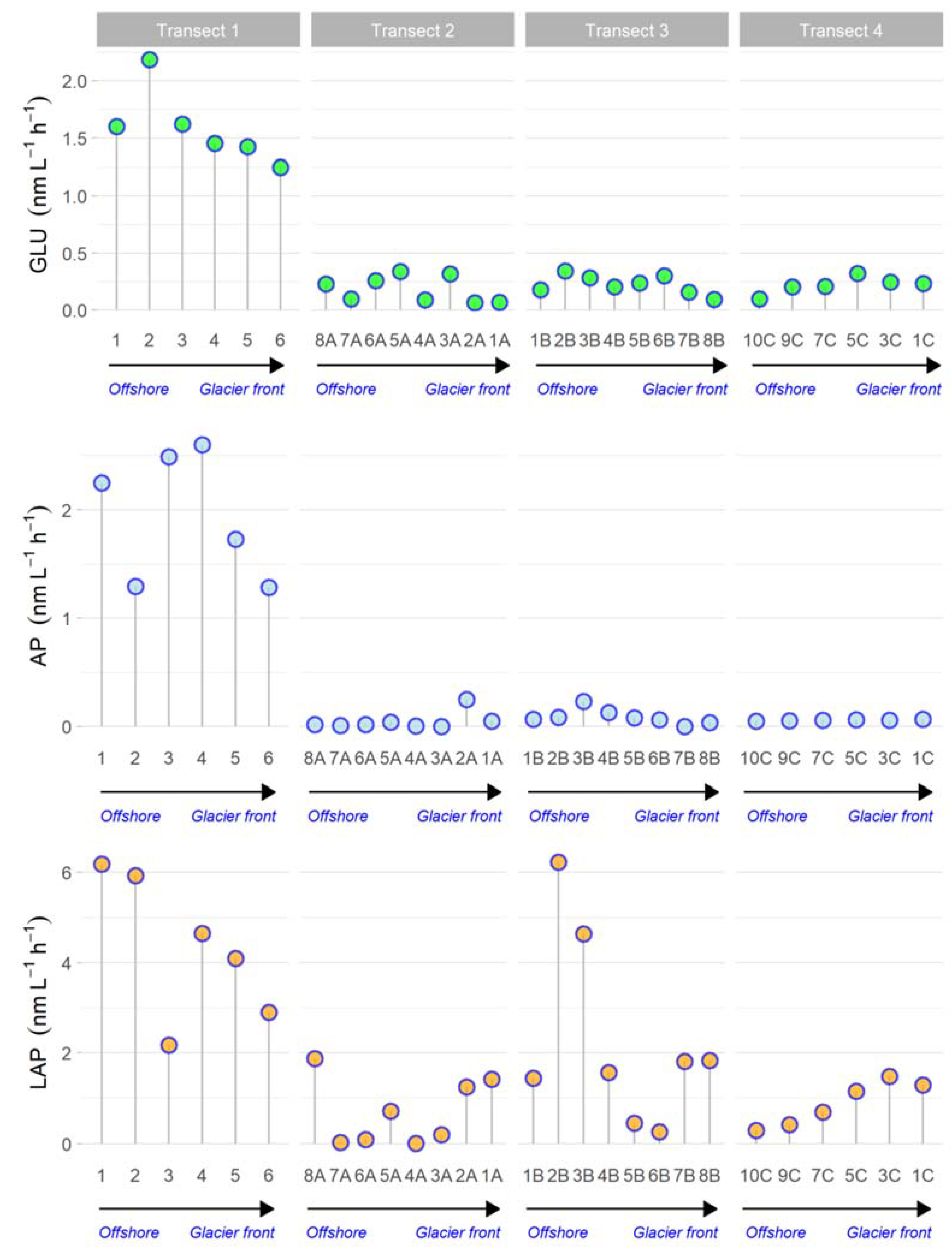
3.5. Microbial Metabolism
3.5.1. Physiological Profiles of Microbial Community
3.5.2. Enzymatic Activities
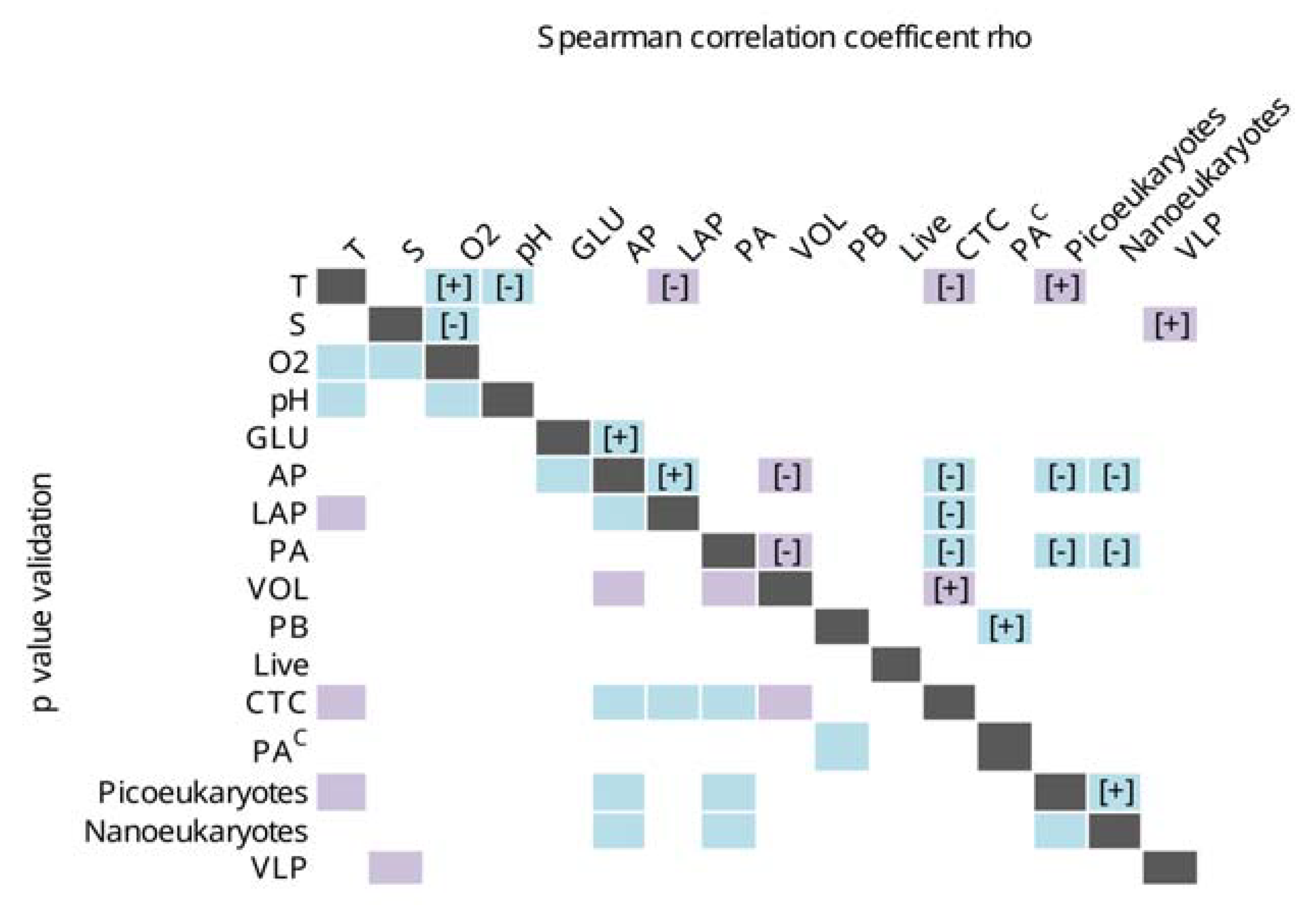
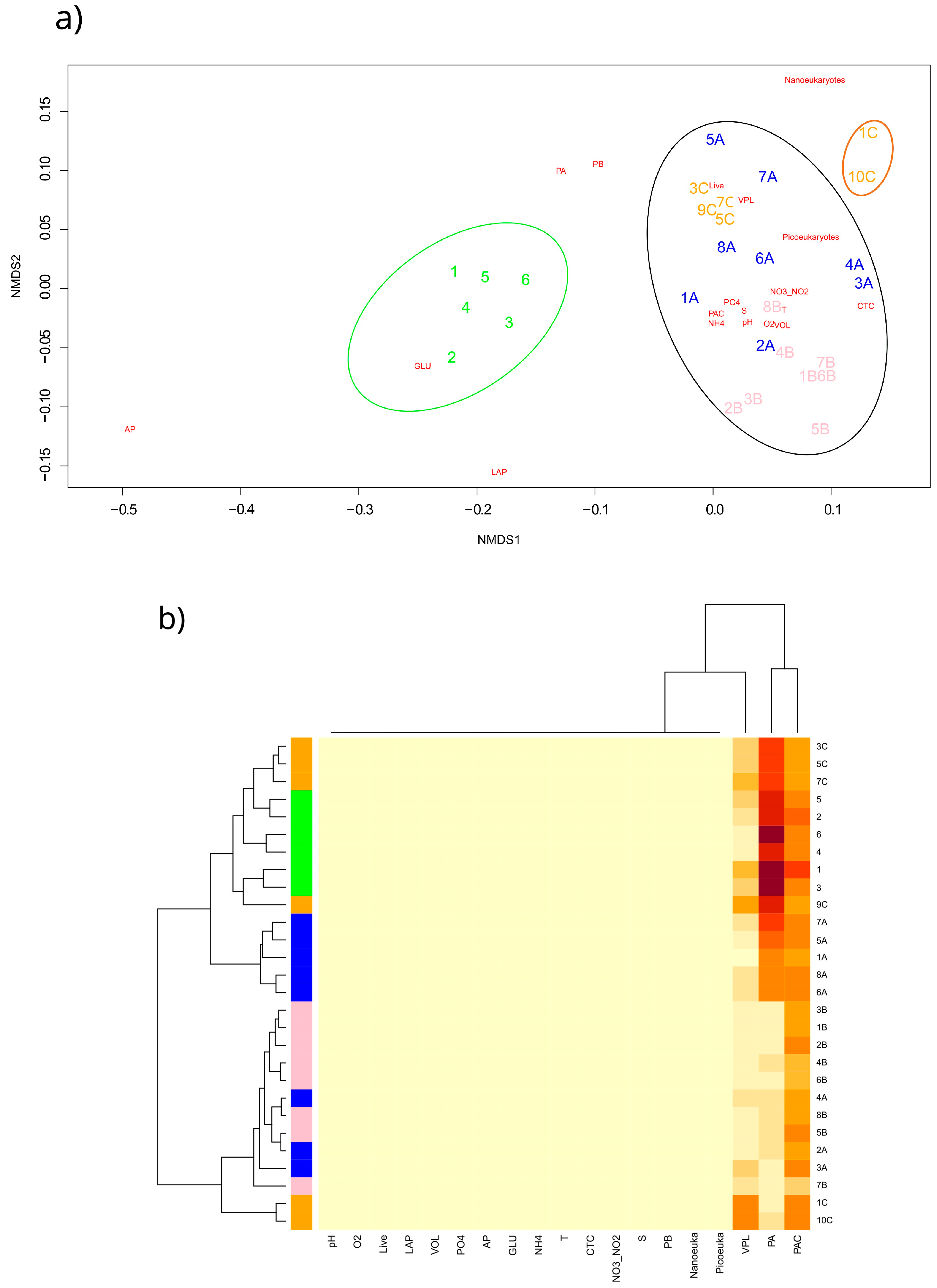
3.6. Statistical Analysis
4. Discussion
5. Conclusions
- −
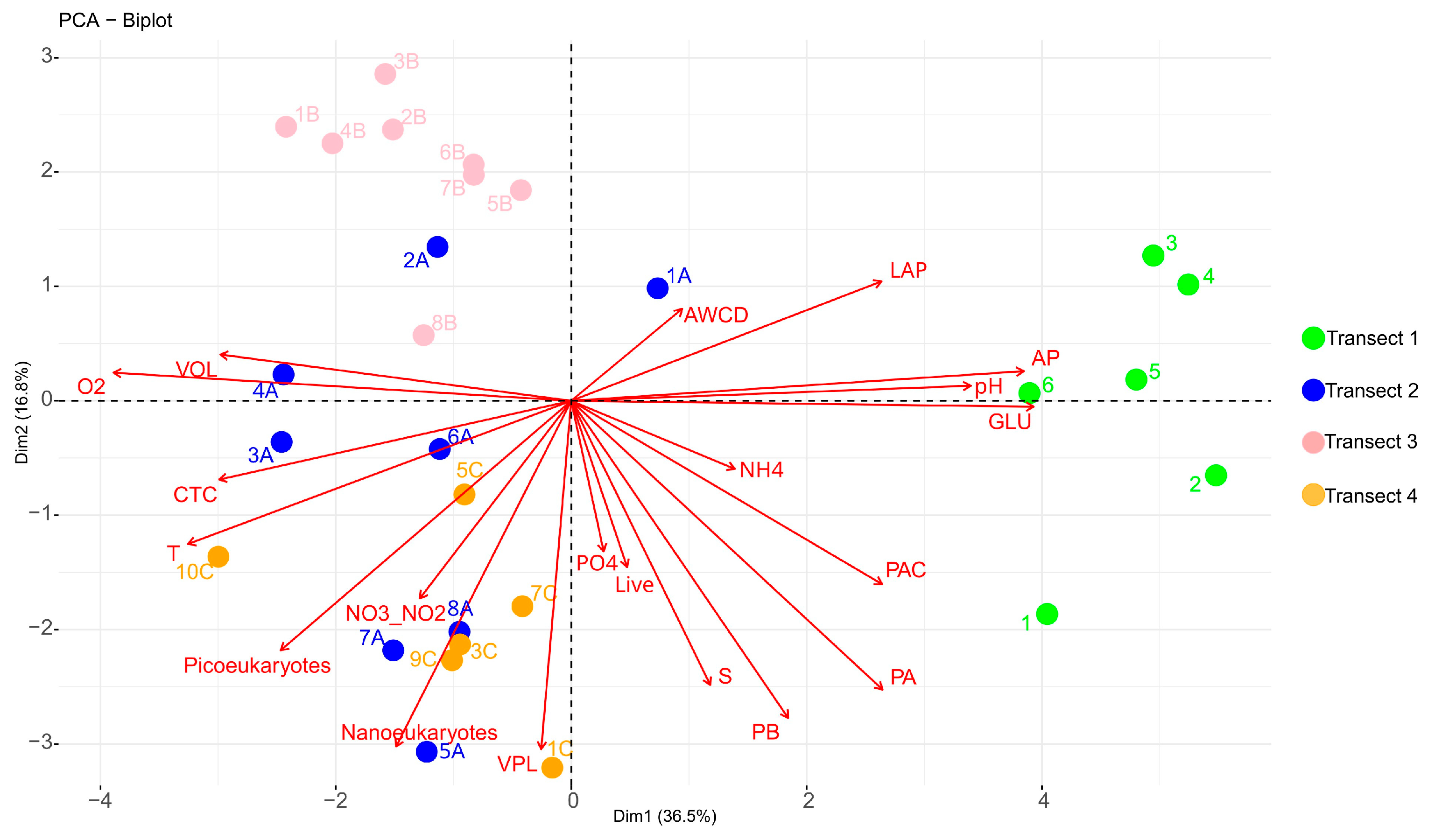
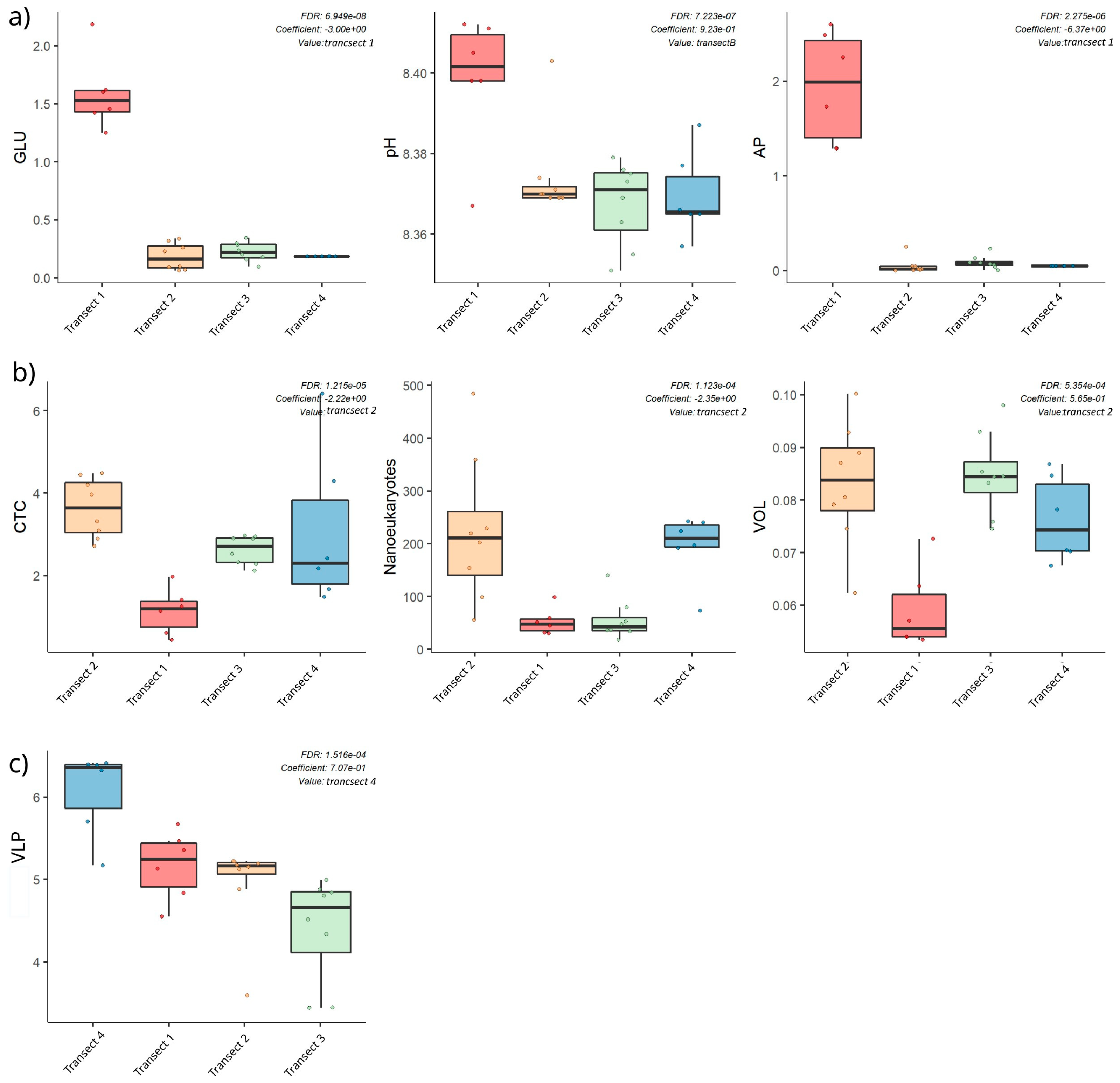
- Transect 1 was totally different from the others (2, 3, 4) that were mostly well-characterised in their distinctive properties (even if with some anomalous samples). The regression also confirmed these significant differences between Transect 1 and the others, as well as between Transects 3 and 4;
- −
- Transect 2 was characterised by large-sized microorganisms, actively respiring cells and terrigenous inputs (i.e., Nanoeukaryotes, VOL, CTC, PO43−);
- −
- In Transect 4, the microbial community appeared to be controlled by viruses and characterised by small-sized cells (Picoeukaryotes) due to the reduced temperature and dilution of glacier water (low salinity);
- −
- Differences in microbial abundance and metabolism were attributable to complex environmental conditions like ice melting or Atlantic water inflow.
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pedrós-Alió, C.; Potvin, M.; Lovejoy, C. Diversity of planktonic microorganisms in the Arctic Ocean. Prog. Oceanogr. 2015, 139, 233–243. [Google Scholar] [CrossRef]
- Azam, F.; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol. 2007, 5, 782–791. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.; Almeida, A.; Coelho, F.J.R.C.; Gomes, N.C.M.; Oliveira, V.; Santos, A.L. Bacterial extracellular enzymatic activity in globally changing aquatic ecosystems. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 124–135. [Google Scholar]
- Alonso-Sáez, L.; Sánchez, O.; Gasol, J.M.; Balagué, V.; Pedrós-Alio, C. Winter-to-summer changes in the composition and single-cell activity of near-surface Arctic prokaryotes. Environ. Microbiol. 2008, 10, 2444–2454. [Google Scholar] [CrossRef]
- Müller, O.; Wilson, B.; Paulsen, M.L.; Ruminska, A.; Armo, H.R.; Bratbak, G.; Øvreås, L. Spatio temporal dynamics of ammonia-oxidizing Thaumarchaeota in distinct Arctic water masses. Front. Microbiol. 2018, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Caruso, G.; Madonia, A.; Bonamano, S.; Miserocchi, S.; Giglio, F.; Maimone, G.; Azzaro, F.; Decembrini, F.; La Ferla, R.; Piermattei, V.; et al. Microbial abundance and enzyme activity patterns: Response to changing Environmental Characteristics along a Transect in Kongsfjorden (Svalbard Islands). J. Mar. Sci. Eng. 2020, 8, 824. [Google Scholar] [CrossRef]
- Fadeev, E.; Cardozo-Mino, M.G.; Rapp, J.Z.; Bienhold, C.; Salter, I.; Salman-Carvalho, V.; Molari, M.; Tegetmeyer, H.E.; Buttigieg, P.; Boetius, A. Comparison of two 16S rRNA primers (V3–V4 and V4–V5) for studies of Arctic microbial communities. Front. Microbiol. 2021, 12, 637526. [Google Scholar] [CrossRef] [PubMed]
- Carter-Gates, M.; Balestreri, C.; Thorpe, S.E.; Cottier, F.; Baylay, A.; Bibby, T.S.; Moore, C.M.; Schroeder, D.C. Implications of increasing Atlantic influence for Arctic microbial community structure. Sci. Rep. 2020, 10, 19262. [Google Scholar] [CrossRef]
- Boetius, A.; Anesio, A.M.; Deming, J.W.; Mikucki, J.A.; Rapp, J.Z. Microbial ecology of the cryosphere: Sea ice and glacial habitats. Nat. Rev. Microbiol. 2015, 13, 677–690. [Google Scholar] [CrossRef]
- Malone, T.; Azzaro, M.; Bode, A.; Brown, E.; Duce, R.; Kamykowski, D.; Kang, S.H.; Kedong, Y.; Thorndyke, M.; Wang, J.; et al. Primary Production, Cycling of Nutrients, Surface Layer and Plankton. In First Global Integrated Marine Assessment, Also Known as the First World Ocean Assessment: World Ocean Assessment I; Cambridge University Press: Cambridge, United Nations, 2017; United Nations Chapter 6. [Google Scholar]
- Azzaro, M.; Aliani, S.; Maimone, G.; Decembrini, F.; Caroppo, C.; Giglio, F.; Langone, L.; Miserocchi, S.; Cosenza, A.; Azzaro, F.; et al. Short-term dynamics of nutrients, planktonic abundances and microbial respiratory activity in the Arctic Kongsfjorden (Svalbard, Norway). Polar Biol. 2021, 44, 361–378. [Google Scholar] [CrossRef]
- Zappalà, G.; Bruzzone, G.; Caruso, G.; Azzaro, M. Development of an automatic sampler for extreme polar environments: First in situ application in Svalbard Islands. Rend. Fis. Acc. Lincei 2016, 27, 251–259. [Google Scholar] [CrossRef]
- Zappalà, G.; Bruzzone, G.; Azzaro, M.; Caruso, G. New advanced technology devices for operational oceanography in extreme conditions. Int. J. Sustain. Dev. Plan. 2017, 12, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Wietz, M.; Bienhold, C.; Metfies, K.; Torres-Valdés, S.; von Appen, W.J.; Salter, I.; Boetius, A. The polar night shift; seasonal dynamics and drivers of Arctic Ocean microbiomes revealed by autonomous sampling. ISME Comm. 2021, 1, 76. [Google Scholar] [CrossRef]
- Bruzzone, G.; Odetti, A.; Caccia, M. Remote data collection near marine glacier fronts. Sea Technol. 2018, 3, 22–24. [Google Scholar]
- Bruzzone, G.; Odetti, A.; Caccia, M.; Ferretti, R. Monitoring of sea-ice-atmosphere interface in the proximity of Arctic tidewater glaciers: The contribution of marine robotics. Remote Sens. 2020, 12, 1707. [Google Scholar] [CrossRef]
- Svendsen, H.; Beszczynska-Møller, A.; Hagen, J.O.; Lefauconnier, B.; Tverberg, V.; Gerland, S.; Børre Ørbæk, J.; Bischof, K.; Papucci, C.; Zajaczkowski, M.; et al. The physical environment of Kongsfjorden–Krossfjorden, an Arctic fjord system in Svalbard. Polar Res. 2002, 21, 133–166. [Google Scholar] [CrossRef] [Green Version]
- Strickland, J.D.H.; Parsons, T.R. A practical handbook of seawater analysis. J. Fish. Res. Board Can. Bull. 1972, 157, 1–310. [Google Scholar]
- Aminot, A.; Chaussepied, M. Manuel des Analyses Chimiques en Milieu Marin; CNEXO, Editions Louvre: Paris, France, 1983; p. 395. [Google Scholar]
- Porter, K.G.; Feig, Y.S. The use of DAPI for identifying and counting aquatic microflora. Limnol. Oceanogr. 1980, 25, 943–948. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Azzaro, M.; Conversano, F.; Brunet, C.; Cabral, A.S.; Paranhos, R. Vertical distribution of the prokaryotic cell size in the Mediterranean Sea. Helgol. Mar. Res. 2012, 66, 635–650. [Google Scholar] [CrossRef] [Green Version]
- La Ferla, R.; Azzaro, M.; Michaud, L.; Caruso, G.; Lo Giudice, A.; Paranhos, R.; Cabral, A.S.; Conte, A.; Cosenza, A.; Maimone, G.; et al. Prokaryotic abundance and activity in permafrost of the northern Victoria Land and upper Victoria Valley (Antarctica). Microb. Ecol. 2017, 74, 402–415. [Google Scholar] [CrossRef]
- Gasol, J.M.; del Giorgio, P.A. Using flow cytometry for counting natural planktonic bacteria and understanding the structure of planktonic bacterial communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef] [Green Version]
- Brussaard, C.P.D. Viral control of phytoplankton population—A review. J. Eukaryot. Microbiol. 2004, 51, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Marie, D.; Simon, N.; Vaulot, D. Phytoplankton cell counting by flow cytometry. In Algal Culturing Techniques; Andersen, R.A., Ed.; Physiological Society of America: Oxford, UK, 2005; pp. 253–267. [Google Scholar]
- Teira, E.; Reinthaler, T.; Pernthaler, A. Combining catalysed reporter deposition-fluorescence in situ hybridization and microautoradiography to detect substrate utilization by bacteria and Archaea in the deep ocean. Appl. Environ. Microbiol. 2004, 70, 4411–4414. [Google Scholar] [CrossRef] [PubMed]
- Pernthaler, A.; Pernthaler, J.; Amann, R. Sensitive multi-color fluorescence in situ hybridization for the identification of environmental microorganisms. In Molecular Microbial Ecology Manual, 2nd ed.; Kowalchuk, G., Bruijn, F.J., Head, I.M., Akkermans, A.D., Elsas, J.D., Eds.; Kluwer Academic Press: Dordrecht, The Netherlands, 2004; Volume 11, pp. 711–726. [Google Scholar]
- Wallner, G.; Amann, R.; Beisker, W. Optimizing fluorescent in situ hybridization of suspended cells with rRNA-targeted oligonucleotide probes for the flow cytometric identification of microorganisms. Cytometry 1993, 14, 136–143. [Google Scholar] [CrossRef] [PubMed]
- Amann, R.I.; Binder, B.J.; Olson, R.J.; Chisholm, S.W.; Devereux, R.; Stahl, D.A. Combination of 16S rRNA-targeted oligonucleotide probes with flow cytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol. 1990, 56, 1919–1925. [Google Scholar] [CrossRef] [Green Version]
- Daims, H.; Brühl, A.; Amann, R.; Schleifer, K.H.; Wagner, M. The domain-specific probe EUB338 is insufficient for the detection of all Bacteria: Development and evaluation of a more comprehensive probe set. Syst. Appl. Microbiol. 1999, 22, 434–444. [Google Scholar] [CrossRef]
- Neef, A. Application of In Situ Identification of Bacteria to Population Analysis in Complex Microbial Communities. Ph.D. Thesis, Technical University of Munich, Munich, Germany, 1997. [Google Scholar]
- Manz, W.; Amann, R.; Ludwig, W.; Wagner, M.; Schleifer, K.-H. Phylogenetic oligodeoxynucleotide probes for the major subclasses of proteobacteria: Problems and solutions. Syst. Appl. Microbiol. 1992, 15, 593–600. [Google Scholar] [CrossRef]
- Loy, A.; Lehner, A.; Lee, N.; Adamczyk, J.; Meier, H.; Ernst, J.; Schleifer, K.H.; Wagner, M. Oligonucleotide microarray for 16S rRNA gene based detection of all recognized lineages of sulfate reducing prokaryotes in the environment. Appl. Environ. Microbiol. 2002, 68, 5064–5081. [Google Scholar] [CrossRef] [Green Version]
- Loy, A.; Maixner, F.; Wagner, M.; Horn, M. ProbeBase: An online resource for rRNA-targeted oligonucleotide probes. Nucleic Acids Res. 2007, 35, 800–804. [Google Scholar] [CrossRef] [Green Version]
- Manz, W.; Amann, R.; Ludwig, W.; Vancanneyt, M.; Schleifer, K.H. Application of a suite of 16S rRNA specific oligonucleotide probes designed to investigate bacteria of the phylum Cytophaga-Flavobacter-Bacteroides in the natural environment. Microbiology 1996, 142, 1097–1106. [Google Scholar] [CrossRef] [Green Version]
- Stahl, D.A.; Amann, R. Development and application of nucleic acid probes. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrand, E., Goodfellow, M., Eds.; Wiley: Chichester, UK, 1991; pp. 205–248. [Google Scholar]
- Garland, J.L.; Mills, A.L. Classification and characterization of heterotrophic microbial communities on the basis of patterns of community-level sole-carbon-source utilization. Appl. Environ. Microbiol. 1991, 57, 2351–2359. [Google Scholar] [CrossRef] [Green Version]
- Garland, J.L. Analytical approaches to the characterization of samples of microbial communities using patterns of potential C source utilization. Soil Biol. Biochem. 1996, 28, 213–221. [Google Scholar] [CrossRef]
- Sala, M.M.; Arin, L.; Balagué, V.; Felipe, J.; Guadayol, O.; Vaqué, D. Functional diversity of bacterioplankton assemblages in Western Antarctic seawaters during late spring. Mar. Ecol. Progr. Ser. 2005, 292, 13–21. [Google Scholar] [CrossRef]
- Sala, M.M.; Estrada, M.; Gasol, J.M. Seasonal changes in the functional diversity of bacterioplankton in contrasting coastal environments of the NW Mediterranean. Aquat. Microb. Ecol. 2006, 44, 1–9. [Google Scholar] [CrossRef]
- Hoppe, H.G. Use of fluorogenic model substrates for extracellular enzyme activity (EEA) measurement of bacteria. In Handbook of Methods in Aquatic Microbial Ecology; Kemp, P.F., Sherr, B.F., Sherr, E.B., Cole, J.J., Eds.; Lewis Publisher: Boca Raton, FL, USA, 1993; pp. 423–432. [Google Scholar]
- Clarke, K.R.; Gorley, R.N.; Somerfield, P.J.; Warwick, R.M. Change in Marine Communities: An Approach to Statistical Analysis and Interpretation, 3rd ed.; PRIMER-E Ltd.: Plymouth, UK, 2014. [Google Scholar]
- Piermattei, V.; Madonia, A.; Bonamano, S.; Martellucci, R.; Bruzzone, G.; Ferretti, R.; Odetti, A.; Azzaro, M.; Zappalà, G.; Marcelli, M. Cost-Effective technologies to study the Arctic Ocean environment. Sensors 2018, 18, 2257. [Google Scholar] [CrossRef] [Green Version]
- Pasculli, L.; Piermattei, V.; Madonia, A.; Bruzzone, G.; Caccia, M.; Ferretti, R.; Odetti, A.; Marcelli, M. New cost-effective technologies applied to the study of the glacier melting influence on physical and biological processes in Kongsfjorden area (Svalbard). J. Mar. Sci. Eng. 2020, 8, 593. [Google Scholar] [CrossRef]
- Tam, L.; Kevan, P.G.; Trevors, J.T. Viable bacterial biomass and functional diversity in fresh and marine waters in the Canadian Arctic. Polar Biol. 2003, 26, 287–294. [Google Scholar] [CrossRef]
- Jankowska, K.; Wiodarska-Kowalczuk, M.; Wieczorek, P. Abundance and biomass of bacteria in two glacial fjords. Pol. Polar Res. 2005, 26, 77–84. [Google Scholar]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Herndl, G.J. Changes in viral and bacterial communities during the ice-melting season in the coastal Arctic (Kongsforden, Ny-Ålesund). Environ. Microbiol. 2011, 13, 1827–1841. [Google Scholar] [CrossRef]
- Rokkan Iversen, K.; Seuthe, L. Seasonal microbial processes in a high-latitude fjord (Kongsfjorden, Svalbard): I. Heterotrophic bacteria, picoplankton and nanoflagellates. Polar Biol. 2011, 34, 731–749. [Google Scholar] [CrossRef] [Green Version]
- Sinha, R.K.; Krishnan, K.P.; Kerkar, S.; David, T.D. Spatio-temporal monitoring and ecological significance of retrievable pelagic heterotrophic bacteria in Konsfjorden, an Arctic Fjord. Indian J. Microbiol. 2016, 57, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Kühn, D.; Oye, V.; Albaric, J.; Harris, D.; Hillers, G.; Braathen, A.; Olaussen, S. Preparing for CO2 storage in the Arctic—Assessing background seismic activity and noise characteristics at the CO2 Lab site, Svalbard. Energy Procedia 2014, 63, 4313–4322. [Google Scholar] [CrossRef] [Green Version]
- Papale, M.; Conte, A.; Mikkonen, A.; Michaud, L.; La Ferla, R.; Azzaro, M.; Caruso, G.; Paranhos, R.; Cabral, A.S.; Maimone, G.; et al. Prokaryotic assemblages within permafrost active layer at Edmonson Point (Northern Victoria Land, Antarctica). Soil Biol. Biochem. 2018, 123, 165–179. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Caruso, G.; Azzaro, F.; Azzaro, M.; Decembrini, F.; Cosenza, A.; Leonardi, M.; Paranhos, R. Are prokaryotic cell shape and size suitable to ecosystem characterization? Hydrobiologia 2014, 726, 65–80. [Google Scholar] [CrossRef]
- La Ferla, R.; Maimone, G.; Lo Giudice, A.; Azzaro, F.; Cosenza, A.; Azzaro, M. Cell size and other phenotypic traits of prokaryotic cells in pelagic areas of the Ross Sea (Antarctica). Hydrobiologia 2015, 761, 181–194. [Google Scholar] [CrossRef]
- Kuznetsovaa, E.V.; Kosolapova, D.B.; Krylova, A.V. Changes in size-morphological structure of bacterioplankton in freshwater environments of Svalbard. Contemp. Probl. Ecol. 2022, 15, 139–146. [Google Scholar] [CrossRef]
- Sherr, B.F.; delGiorgio, P.; Sherr, E.B. Estimating abundance and single-cell characteristics of respiring bacteria via the redox dye CTC. Aquat. Microb. Ecol. 1999, 18, 117–131. [Google Scholar] [CrossRef]
- Howard-Jones, M.H.; Ballard, V.D.; Allen, A.E.; Frischer, M.E.; Verity, P.G. Distribution of bacterial biomass and activity in the marginal ice zone of the central Barents Sea during summer. J. Mar. Syst. 2002, 38, 77–91. [Google Scholar] [CrossRef]
- Jiang, X.; He, J.; Cai, M. Abundance and biomass of heterotrophic microbes in the Kongsfjorden, Svalbard. Acta Oceanol. Sin. 2005, 24, 143–152. [Google Scholar]
- Wommack, E.K.; Nasko, D.J.; Chopyk, J.; Sakowski, G.E. Counts and sequences, observations that continue to change our understanding of viruses in nature. J. Microbiol. 2015, 53, 181–192. [Google Scholar] [CrossRef]
- Knowles, B.; Silveira, C.B.; Bailey, B.A.; Barott, K.; Cantu, V.A.; Cobián-Güemes, A.G.; Coutinho, F.H.; Dinsdale, E.A.; Felts, B.; Furby, K.A.; et al. Lytic to temperate switching of viral communities. Nature 2016, 531, 466–470. [Google Scholar] [CrossRef]
- Wigington, C.H.; Sonderegger, D.; Brussaard, C.P.; Buchan, A.; Finke, J.F.; Fuhrman, J.A.; Lennon, J.T.; Middelboe, M.; Suttle, C.A.; Stock, C.; et al. Re-examination of the relationship be-tween marine virus and microbial cell abundances. Nat. Microbiol. 2016, 1, 15024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabral, A.S.; Lessa, M.M.; Mello, M.P.; Martins, R.M.; Pulgati, F.H.; Paranhos, R. Occurrence and role of virioplankton in a tropical estuarine system. Hydrobiologia 2020, 847, 4125–4140. [Google Scholar] [CrossRef]
- Flombaum, P.; Gallegos, J.L.; Gordillo, R.A.; Rincón, J.; Zabala, L.L.; Jiao, N.; Karl, D.M.; Li, W.K.W.; Lomas, M.W.; Veneziano, D.; et al. Present and future global distributions of the marine Cyanobacteria Prochlorococcus and Synechococcus. Proc. Natl. Acad. Sci. USA 2013, 110, 9824–9829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; He, J.; Zhao, Y.; Zhang, F.; Cai, M. Flow cytometry investigation of picoplankton across latitudes and along the circum Antarctic Ocean. Acta Oceanol. Sin. 2012, 31, 134–142. [Google Scholar] [CrossRef]
- Brussaard, C.P.D.; Payet, J.P.; Winter, C.; Weinbauer, M.G. Quantification of aquatic viruses by flow cytometry. In Manual of Aquatic Viral Ecology; Wilhelm, S.W., Weinbauer, M.G., Suttle, C.A., Eds.; American Society of Limnology and Oceanography: Waco, TX, USA, 2010; Chapter 11; pp. 102–109. [Google Scholar] [CrossRef]
- Mojica, K.D.A.; Huisman, J.; Wilhelm, S.W.; Brussaard, C. Latitudinal variation in virus-induced mortality of phytoplankton across the North Atlantic Ocean. ISME J. 2016, 10, 500–513. [Google Scholar] [CrossRef] [Green Version]
- Papale, M.; Lo Giudice, A.; Conte, A.; Rizzo, C.; Rappazzo, A.C.; Maimone, G.; Caruso, G.; La Ferla, R.; Azzaro, M.; Gug-liandolo, C.; et al. Microbial assemblages in pressurized Antarctic brine pockets (Tarn Flat, Northern Victoria Land): A hotspot of biodiversity and activity. Microorganisms 2019, 7, 333. [Google Scholar] [CrossRef] [Green Version]
- Zaccone, R.; Caruso, G. Microbial enzymes in the Mediterranean Sea: Relationship with climate changes. AIMS Microbiol. 2019, 5, 251–271. [Google Scholar] [CrossRef]
- Delpech, L.-M.; Vonnahme, T.R.; McGovern, M.; Gradinger, R.; Præbel, K.; Poste, A.E. Terrestrial inputs shape coastal Bacterial and Archaeal communities in a High Arctic Fjord (Isfjorden, Svalbard). Front. Microbiol. 2021, 12, 614634. [Google Scholar] [CrossRef]
- Han, D.; Richter-Heitmann, T.; Kim, I.N.; Choy, E.; Park, K.T.; Unno, T.; Kim, J.; Nam, S.I. Survey of Bacterial Phylogenetic Diversity During the Glacier Melting Season in an Arctic Fjord. Microb. Ecol. 2021, 81, 579–591. [Google Scholar] [CrossRef]
- Zhang, X.; Ma, X.; Wang, N.; Yao, T. New subgroup of Bacteroidetes and diverse microorganisms in Tibetan plateau gla-cial ice provide a biological record of environmental conditions. FEMS Microbiol. Ecol. 2009, 67, 21–29. [Google Scholar] [CrossRef]
- Hell, K.; Edwards, A.; Zarsky, J.; Podmirseg, S.M.; Girdwood, S.; Pachebat, J.A.; Insam, H.; Sattler, B. The dynamic bacterial communities of a melting high Arctic glacier snowpack. ISME J. 2013, 7, 1814–1826. [Google Scholar] [CrossRef] [PubMed]
- Musilova, M.; Tranter, M.; Bennett, S.A.; Wadham, J.; Anesio, A.M. Stable microbial community composition on the Greenland icesheet. Front. Microbiol. 2015, 6, 193. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Krishnan, K.P. Differences in free-living and particle associated bacterial communities and their spatial variation in Kongsfjorden, Arctic. J. Basic Microbiol. 2017, 57, 827–838. [Google Scholar] [CrossRef] [PubMed]
- De Corte, D.; Sintes, E.; Yokokawa, T.; Herndl, G.J. Comparison between MICRO–CARD–FISH and 16 S rRNA gene clone libraries to assess the active versus total bacterial community in the coastal Arctic. Environ. Microbiol. Rep. 2013, 5, 272–281. [Google Scholar] [CrossRef]
- Piquet, A.-T.; Maat, D.; Confurius-Guns, V.; Sintes, E.; Herndl, G.; van de Poll, W.; Wiencke, C.; Buma, A.; Bolhuis, H. Springtime dynamics, productivity and activity of prokaryotes in two Arctic fjords. Polar Biol. 2016, 39, 1749–1763. [Google Scholar] [CrossRef]
- Piquet, A.-T.; Scheepens, J.; Bolhuis, H.; Wiencke, C.; Buma, A. Variability of protistan and bacterial communities in two Arctic fjords (Spitsbergen). Polar Biol. 2010, 33, 1521–1536. [Google Scholar] [CrossRef] [Green Version]
- Harding, T.; Jungblut, A.D.; Lovejoy, C.; Vincent, W.F. Microbes in high Arctic snow and implications for the cold biosphere. Appl. Environ. Microbiol. 2011, 77, 3234–3243. [Google Scholar] [CrossRef] [Green Version]
- Zarsky, J.D.; Stibal, M.; Hodson, A.; Sattler, B.; Schostag, M.; Hansen, L.H.; Jacobsen, C.S.; Psenner, R. Large cryoconite aggregates on a Svalbard glacier support a diverse microbial community including ammonia-oxidizing archaea. Environ. Res. Lett. 2013, 8, 35044. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transect 1 | Transect 2 | Transect 3 | Transect 4 | ||||
|---|---|---|---|---|---|---|---|
| Stations | Glacier Distance (m) | Stations | Glacier Distance (m) | Stations | Glacier Distance (m) | Stations | Glacier Distance (m) |
| 6 | 286.2 | 1A | 6.7 | 8B | 216.6 | 1C | 0.8 |
| 5 | 322.54 | 2A | 10.91 | 7B | 247.47 | 3C | 49.13 |
| 4 | 381.74 | 3A | 45.16 | 6B | 309.89 | 5C | 177.52 |
| 3 | 418.71 | 4A | 82.4 | 5B | 378.52 | 7C | 351.89 |
| 2 | 423.69 | 5A | 133.21 | 4B | 405.7 | 9C | 462.47 |
| 1 | 483.30 | 6A | 167.24 | 3B | 361.69 | 10C | 562.85 |
| 7A | 222.13 | 2B | 388.48 | ||||
| 8A | 321.8 | 1B | 403.38 | ||||
| Probes | Target Group | Probe Sequence (5′–3′) | References |
|---|---|---|---|
| EUB338I | Most, but not all, bacteria | GCTGCCTCCCGTAGGAGT | [29] |
| EUB338II | Planctomycetes | GCAGCCACCCGTAGGTGT | [30] |
| EUB338III | Verrucomicrobiales | GCTGCCACCCGTAGGTGT | [30] |
| ALF968 | Alphaproteobacteria | GGTAAGGTTCTGCGCGTT | [31] |
| BET42a | Betaproteobacteria | GCCTTCCCACTTCGTTT | [32] |
| GAM42a | Gammaproteobacteria | GCCTTCCCACATCGTTT | [32] |
| DELTA495a | Most Deltaproteobacteria | AGTTAGCCGGTGCTTCCT | [33] |
| DELTA495b | Deltaproteobacteria | AGTTAGCCGGCGCTTCCT | [33] |
| DELTA495c | Deltaproteobacteria | AATTAGCCGGTGCTTCCT | [33] |
| EPSY914 | Epsilonproteobacteria | GGTCCCCGTCTATTCCTT | [34] |
| CF319a | Bacteroidetes | TGGTCCGTGTCTCAGTAC | [35] |
| ARCH915 | Archaea | GTGCTCCCCCGCCAATTCCT | [36] |
| Stations | T | S | O2 | pH | PO43− | NH4+ | NO2− + NO3− | |
|---|---|---|---|---|---|---|---|---|
| °C | psu | ppm | µM | µM | µM | |||
| 6 | 4.31 | 31.967 | 9.68 | 8.39 | 0.30 | 2.16 | 1.81 | |
| 5 | 3.69 | 31.64 | 9.53 | 8.41 | 0.33 | 1.51 | 1.69 | |
| Transect 1 | 4 | 4.20 | 31.49 | 9.63 | 8.40 | 0.23 | 1.38 | 1.86 |
| 3 | 3.42 | 30.97 | 9.33 | 8.41 | 0.27 | 1.49 | 1.79 | |
| 2 | 3.98 | 31.43 | 9.60 | 8.39 | 0.29 | 2.60 | 3.87 | |
| 1 | 5.67 | 32.11 | 10.04 | 8.37 | 0.43 | 1.36 | 6.62 | |
| 1A | 3.59 | 31.21 | 10.99 | 8.40 | 0.20 | 2.69 | 2.00 | |
| 2A | 6.20 | 31.08 | 11.74 | 8.37 | 0.22 | 1.23 | 3.00 | |
| 3A | 6.23 | 31.23 | 11.70 | 8.37 | 0.34 | 1.09 | 5.00 | |
| Transect 2 | 4A | 6.27 | 30,99 | 11.79 | 8.37 | 0.24 | 0.90 | 3.38 |
| 5A | 6.28 | 30.90 | 11.78 | 8.37 | 0.23 | 0.93 | 2.99 | |
| 6A | 6.25 | 31.11 | 11.74 | 8.37 | 0.26 | 1.70 | 3.14 | |
| 7A | 6.17 | 31.11 | 11.76 | 8.37 | 0.64 | 1.31 | 4.37 | |
| 8A | 5.93 | 31.43 | 11.63 | 8.37 | 0.41 | 1.61 | 4.62 | |
| 8B | 5.65 | 31.92 | 10.53 | 8.38 | 0.2 | 1.47 | 3.64 | |
| 7B | 4.61 | 31.66 | 11.09 | 8.38 | 0.32 | 1.35 | 3.28 | |
| 6B | 4.61 | 31.64 | 11.09 | 8.37 | 0.28 | 1.36 | 3.17 | |
| Transect 3 | 5B | 6.01 | 31.63 | 11.68 | 8.37 | 0.25 | 1.78 | 3.01 |
| 4B | 5.97 | 24.95 | 12.34 | 8.37 | 0.36 | 1.42 | 4.22 | |
| 3B | 5.48 | 25.52 | 12.18 | 8.35 | 0.22 | 1.43 | 3.32 | |
| 2B | 6.126 | 25.33 | 12.29 | 8.36 | 0.24 | 1.56 | 2.81 | |
| 1B | 5.94 | 25.81 | 12.21 | 8.35 | 0.26 | 1.13 | 4.64 | |
| 1C | 5.17 | 31.78 | 11.19 | 8.39 | 0.17 | 1.44 | 6.7 | |
| 3C | 6.32 | 31.80 | 11.53 | 8.37 | 0.39 | 2.61 | 4.07 | |
| Transect 4 | 5C | 6.41 | 31.67 | 11.53 | 8.36 | 0.22 | 1.21 | 2.04 |
| 7C | 5.70 | 31.98 | 11.48 | 8.38 | 0.32 | 1.81 | 4.17 | |
| 9C | 6.39 | 31.81 | 11.66 | 8.36 | 0.27 | 2.145 | 4.25 | |
| 10C | 6.39 | 33.12 | 11.59 | 8.36 | 0.21 | 1.625 | 3.14 |
| Stations | Live | CTC | |
|---|---|---|---|
| % | % | ||
| 6 | 1.81 | 1.97 | |
| 5 | 1.17 | 0.44 | |
| Transect 1 | 4 | 1.41 | 0.61 |
| 3 | 0.64 | 1.41 | |
| 2 | 15.8 | 1.25 | |
| 1 | 198 | 1.14 | |
| 1A | 0.99 | 3.09 | |
| 2A | 1.68 | 3.31 | |
| 3A | 2.07 | 4.48 | |
| Transect 2 | 4A | 1.31 | 4.44 |
| 5A | 0.51 | 4.2 | |
| 6A | 1.47 | 3.97 | |
| 7A | 3.21 | 2.89 | |
| 8A | 1.10 | 2.72 | |
| 8B | 0.70 | 2.90 | |
| 7B | 1.60 | 2.89 | |
| 6B | 1.09 | 2.97 | |
| Transect 3 | 5B | 0.56 | 2.32 |
| 4B | 0.91 | 2.52 | |
| 3B | 0.90 | 2.94 | |
| 2B | 5.84 | 2.28 | |
| 1B | 0.99 | 2.11 | |
| 1C | 20.8 | 2.17 | |
| 3C | 13.8 | 4.29 | |
| Transect 4 | 5C | 0.96 | 1.48 |
| 7C | 3.74 | 1.67 | |
| 9C | 0.89 | 2.42 | |
| 10C | 0.61 | 6.41 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Papale, M.; Caruso, G.; Maimone, G.; La Ferla, R.; Lo Giudice, A.; Rappazzo, A.C.; Cosenza, A.; Azzaro, F.; Ferretti, R.; Paranhos, R.; et al. Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle. Water 2023, 15, 556. https://doi.org/10.3390/w15030556
Papale M, Caruso G, Maimone G, La Ferla R, Lo Giudice A, Rappazzo AC, Cosenza A, Azzaro F, Ferretti R, Paranhos R, et al. Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle. Water. 2023; 15(3):556. https://doi.org/10.3390/w15030556
Chicago/Turabian StylePapale, Maria, Gabriella Caruso, Giovanna Maimone, Rosabruna La Ferla, Angelina Lo Giudice, Alessandro Ciro Rappazzo, Alessandro Cosenza, Filippo Azzaro, Roberta Ferretti, Rodolfo Paranhos, and et al. 2023. "Microbial Community Abundance and Metabolism Close to the Ice-Water Interface of the Blomstrandbreen Glacier (Kongsfjorden, Svalbard): A Sampling Survey Using an Unmanned Autonomous Vehicle" Water 15, no. 3: 556. https://doi.org/10.3390/w15030556