
Oxidative Stress Response in the Seaweed Padina pavonica Associated with the Invasive Halimeda incrassata and Penicillus capitatus
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Collection and Processing
2.2. Biochemical Analysis
2.3. Statistical Analyses
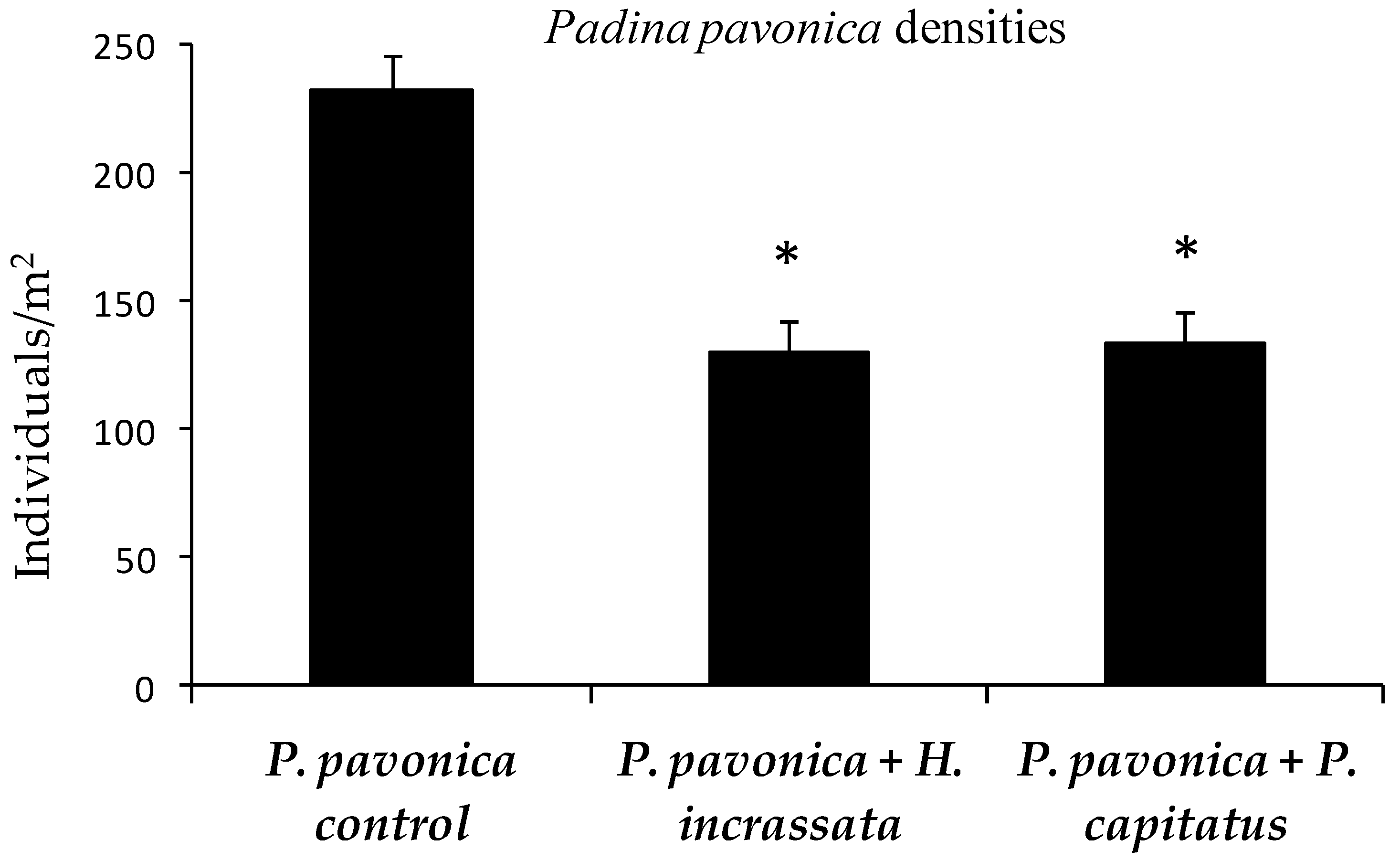
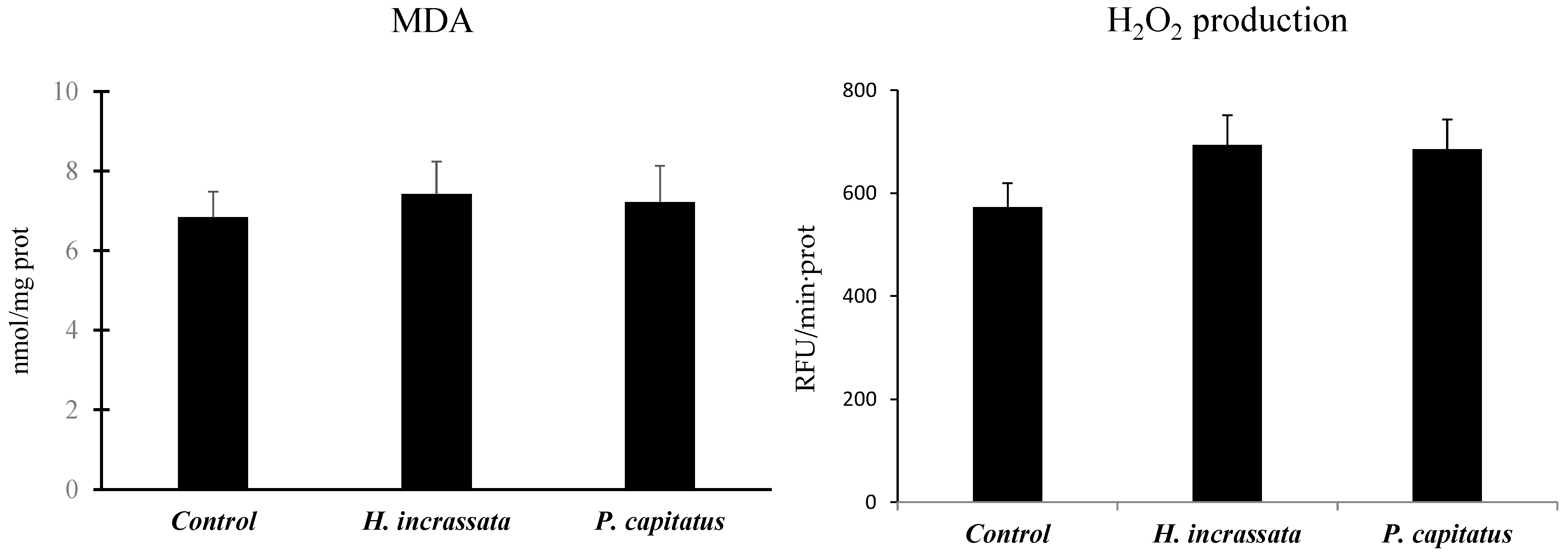
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Austen, M.C.; Crowe, T.P.; Elliott, M.; Paterson, D.M.; Peck, M.A.; Piraino, S. Vectors of change in the marine environment: Ecosystem and economic impacts and management implications. Estuar. Coast. Shelf Sci. 2018, 201, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Mineur, F.; Arenas, F.; Assis, J.; Davies, A.J.; Engelen, A.H.; Fernandes, F.; Malta, E.; Thibaut, T.; Van Nguyen, T.; Vaz-Pinto, F.; et al. European seaweeds under pressure: Consequences for communities and ecosystem functioning. J. Sea Res. 2015, 98, 91–108. [Google Scholar] [CrossRef]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of invasive alien marine species on ecosystem services and biodiversity: A pan-European review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. Mare Nostrum, mare quod invaditur—The history of bioinvasions in the mediterranean sea. In Histories of Bioinvasions in the Mediterranean; Queiroz, A., Pooley, S., Eds.; Springer: Cham, Denmark, 2018; Volume 8, pp. 21–49. [Google Scholar]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Thomsen, M.S.; Wernberg, T.; South, P.M.; Schiel, D.R. Non-native seaweeds drive changes in marine coastal communities around the world. In Seaweed Phylogeography; Hu, Z.M., Fraser, C., Eds.; Springer: Dordrecht, The Netherlands, 2016; pp. 147–185. [Google Scholar]
- Hillis-Colinvaux, L. Ecology and taxonomy of Halimeda: Primary producer of coral reefs. In Advances in Marine Biology; Blaxter, J.H.S., Russell, F.S., Yonge, M., Eds.; Academic Press: London, UK, 1980; pp. 1–327. [Google Scholar]
- Verbruggen, H.; Tyberghein, L.; Pauly, K.; Vlaeminck, C.; Nieuwenhuyze, K.V.; Kooistra, W.H.; Leliaert, F.; Clerck, O.D. Macroecology meets macroevolution: Evolutionary niche dynamics in the seaweed Halimeda. Glob. Ecol. Biogeogr. 2009, 18, 393–405. [Google Scholar] [CrossRef] [Green Version]
- Alós, J.; Tomas, F.; Terrados, J.; Verbruggen, H.; Ballesteros, E. Fast-spreading green beds of recently introduced Halimeda incrassata (Bryopsidales, Chlorophyta) invade the NW Mediterranean. Mar. Ecol. Prog. Ser. 2016, 558, 153–158. [Google Scholar] [CrossRef] [Green Version]
- Gallardo, T.; Garreta, A.G.; Ribera, M.A.; Cormaci, M.; Furnari, G.; Giaccone, G.; Boudouresque, C.F. Check-list of Mediterranean seaweeds. II. Chlorophyceae Wille sl. Bot. Mar. 1993, 36, 399–422. [Google Scholar] [CrossRef]
- Sureda, A.; Tejada, S.; Capó, X.; Melià, C.; Ferriol, P.; Pinya, S.; Mateu-Vicens, G. Oxidative stress response in the seagrass Posidonia oceanica and the seaweed Dasycladus vermicularis associated to the invasive tropical green seaweed Halimeda incrassata. Sci. Total Environ. 2017, 601, 918–925. [Google Scholar] [CrossRef]
- Guzzetti, E.; Salabery, E.; Ferriol, P.; Díaz, J.A.; Tejada, S.; Faggio, C.; Sureda, A. Oxidative stress induction by the invasive sponge Paraleucilla magna growing on Peyssonnelia squamaria algae. Mar. Environ. Res. 2019, 150, 104763. [Google Scholar] [CrossRef]
- Quetglas-Llabrés, M.M.; Tejada, S.; Capó, X.; Langley, E.; Sureda, A.; Box, A. Antioxidant response of the sea urchin Paracentrotus lividus to pollution and the invasive algae Lophocladia lallemandii. Chemosphere 2020, 261, 127773. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.N.; Cheung, P.C.K.; Ooi, V.E.C.; Ang, P.O. Evaluation of antioxidative activity of extracts from a brown seaweed, Sargassum siliquastrum. J. Agric. Food Chem. 2002, 50, 3862–3866. [Google Scholar] [CrossRef] [PubMed]
- Sureda, A.; Box, A.; Terrados, J.; Deudero, S.; Pons, A. Antioxidant response of the seagrass Posidonia oceanica when epiphytized by the invasive macroalgae Lophocladia lallemandii. Mar. Environ. Res. 2008, 66, 359–363. [Google Scholar] [CrossRef]
- Dring, M.J. Stress resistance and disease resistance in seaweeds: The role of reactive oxygen metabolism. Adv. Bot. Res. 2005, 43, 175–207. [Google Scholar]
- Tejada, S.; Sureda, A. Antioxidant response of the brown algae Dictyota dichotoma epiphyted by the invasive red macroalgae Lophocladia lallemandii. J. Coast. Life Med. 2014, 2, 362–366. [Google Scholar]
- Gennaro, P.; Piazzi, L.; Persia, E.; Porrello, S. Nutrient exploitation and competition strategies of the invasive seaweed Caulerpa cylindracea. Eur. J. Phycol. 2015, 50, 384–394. [Google Scholar] [CrossRef] [Green Version]
- Gniazdowska, A.; Krasuska, U.; Andrzejczak, O.; Soltys, D. Allelopathic compounds as oxidative stress agents: YES or NO. In Reactive Oxygen and Nitrogen Species Signaling and Communication in Plants; Gupta, K., Igamberdiev, A., Eds.; Springer: Cham, Switzerland, 2015; pp. 155–176. [Google Scholar]
- Del Monaco, C.; Hay, M.E.; Gartrell, P.; Mumby, P.J.; Diaz-Pulido, G. Effects of ocean acidification on the potency of macroalgal allelopathy to a common coral. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bedinger, L.A.; Bell, S.S.; Dawes, C.J. Rhizophytic algal communities of shallow, coastal habitats in Florida: Components above and below the sediment surface. Bull. Mar. Sci. 2013, 89, 437–460. [Google Scholar] [CrossRef]
- Benita, M.; Dubinsky, Z.; Iluz, D. Padina pavonica: Morphology and calcification functions and mechanism. Am. J. Plant Sci. 2018, 9, 1156–1168. [Google Scholar] [CrossRef] [Green Version]
- Silberfeld, T.; Bittner, L.; Fernández-García, C.; Cruaud, C.; Rousseau, F.; de Reviers, B.; Leliaert, F.; Payri, C.E.; De Clerck, O. Species diversity, phylogeny and large scale biogeographic patterns of the genus Padina (Phaeophyceae, Dictyotales). J. Phycol. 2013, 49, 130–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chbani, A.; Mawlawi, H.; Etahiri, S. Activité antibactérienne des extraits d’une algue brune Padina pavonica de la côte méditerranéenne au Liban. Phytothérapie 2011, 9, 283–286. [Google Scholar] [CrossRef]
- Khaled, N.; Hiba, M.; Asma, C. Antioxidant and antifungal activities of Padina pavonica and Sargassum vulgare from the Lebanese Mediterranean Coast. Adv. Environ. Biol. 2012, 6, 42–48. [Google Scholar]
- Orlando-Bonaca, M.; Pitacco, V.; Bajt, O.; Falnoga, I.; Hudobivnik, M.J.; Mazej, D.; Šlejkovec, Z.; Bonanno, G. Spatial and temporal distribution of trace elements in Padina pavonica from the northern Adriatic Sea. Mar. Pollut. Bull. 2021, 172, 112874. [Google Scholar] [CrossRef] [PubMed]
- Jordi, A.; Basterretxea, G.; Wang, D.P. Local versus remote wind effects on the coastal circulation of a microtidal bay in the Mediterranean Sea. J. Mar. Syst. 2011, 88, 312–322. [Google Scholar] [CrossRef]
- Rodríguez-Prieto, C.; Ballesteros, E.; Boisset, F.; Afonso-Carrillo, J. Guía de las Macroalgas y Fanerógamas Marinas del Mediterráneo Occidental; Omega SA: Barcelona, Spain, 2013; p. 656. [Google Scholar]
- Aebi, H. Catalase in Vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar]
- Flohé, L.; Ötting, F. Superoxide Dismutase Assays. Methods Enzymol. 1984, 105, 93–104. [Google Scholar]
- Pinya, S.; Tejada, S.; Capó, X.; Sureda, A. Invasive predator snake induces oxidative stress responses in insular amphibian species. Sci. Total Environ. 2016, 566, 57–62. [Google Scholar] [CrossRef]
- Kubiliene, L.; Laugaliene, V.; Pavilonis, A.; Maruska, A.; Majiene, D.; Barcauskaite, K.; Kubilius, R.; Kasparaviciene, G.; Savickas, A. Alternative preparation of propolis extracts: Comparison of their composition and biological activities. BMC Complement. Altern. Med. 2015, 15, 015–0677. [Google Scholar] [CrossRef] [Green Version]
- Pinya, S.; Renga, E.; Fernández, G.; Mateu-Vicens, G.; Tejada, S.; Capó, X.; Sureda, A. Physiological biomarkers in loggerhead turtles (Caretta caretta) as a tool for monitoring sanitary evolution in marine recovery centres. Sci. Total Environ. 2021, 757, 143930. [Google Scholar] [CrossRef]
- Picciotto, M.; Bertuccio, C.; Giacobbe, S.; Spanò, N. Caulerpa taxifolia var. distichophylla: A further stepping stone in the western Mediterranean. Mar. Biodivers. Rec. 2016, 9, 1–5. [Google Scholar]
- Trainito, E.; Doneddu, M.; Furfaro, G. Aliens in changing seascapes: A newly reported non-native sacoglossan (Mollusca, Heterobranchia) in the western Mediterranean Sea. Check List 2022, 18, 545–551. [Google Scholar] [CrossRef]
- Sangil, C.; Martín-García, L.; Afonso-Carrillo, J.; Barquín, J.; Sansón, M. Halimeda incrassata (Bryopsidales, Chlorophyta) reaches the Canary Islands: Mid-and deep-water meadows in the eastern subtropical Atlantic Ocean. Bot. Mar. 2018, 61, 103–110. [Google Scholar] [CrossRef]
- Davis, B.C.; Fourqurean, J.W. Competition between the tropical alga, Halimeda incrassata, and the seagrass, Thalassia testudinum. Aquat. Bot. 2001, 71, 217–232. [Google Scholar] [CrossRef]
- Delgado, O.; Lapointe, B.E. Nutrient-limited productivity of calcareous versus fleshy macroalgae in a eutrophic, carbonate-rich tropical marine environment. Coral Reefs 1994, 13, 151–159. [Google Scholar] [CrossRef]
- Mateo-Ramírez, Á.; Máñez-Crespo, J.; Royo, L.; Tuya, F.; Castejón-Silvo, I.; Hernan, G.; Pereda-Briones, L.; Terrados, J.; Tomas, F. A Tropical Macroalga (Halimeda incrassata) Enhances Diversity and Abundance of Epifaunal Assemblages in Mediterranean Seagrass Meadows. Front. Mar. Sci. 2022, 9, 1030. [Google Scholar] [CrossRef]
- Ross, C.; Kupper, F.C.; Jacobs, R.S. Involvement of reactive oxygen species and reactive nitrogen species in the wound response of Dasycladus vermicularis. Chem. Biol. 2006, 13, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Cavas, L.; Yurdakoc, K. A comparative study: Assessment of the antioxidant system in the invasive green alga Caulerpa racemosa and some macrophytes from the Mediterranean. J. Exp. Mar. Biol. Ecol. 2005, 321, 35–41. [Google Scholar] [CrossRef]
- Faulkner, D.J. Marine natural products. Nat. Prod. Rep. 2001, 18, 1–49. [Google Scholar] [CrossRef]
- Ansari, A.A.; Ghanem, S.M. Growth attributes and biochemical composition of Padina pavonica (L.) from the Red Sea, in response to seasonal alterations of Tabuk, Saudi Arabia. Egypt. J. Aquat. Res. 2019, 45, 139–144. [Google Scholar] [CrossRef]
- Celis-Plá, P.S.; Korbee, N.; Gómez-Garreta, A.; Figueroa, F.L. Seasonal photoacclimation patterns in the intertidal macroalga Cystoseira tamariscifolia (Ochrophyta). Sci. Mar. 2014, 78, 377–388. [Google Scholar] [CrossRef] [Green Version]
- Hakim, M.M.; Patel, I.C. A review on phytoconstituents of marine brown algae. Future J. Pharm. Sci. 2020, 6, 129. [Google Scholar] [CrossRef]
- Pinto, E.; Sigaud-kutner, T.C.; Leitao, M.A.; Okamoto, O.K.; Morse, D.; Colepicolo, P. Heavy metal–induced oxidative stress in algae 1. J. Phycol. 2003, 39, 1008–1018. [Google Scholar] [CrossRef]
- Chale-Dzul, J.; Freile-Pelegrín, Y.; Robledo, D.; Moo-Puc, R. Protective effect of fucoidans from tropical seaweeds against oxidative stress in HepG2 cells. J. Appl. Phycol. 2017, 29, 2229–2238. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| P. pavonica Control | P. pavonica + H. incrassata | P. pavonica + P. capitatus | |
|---|---|---|---|
| Temperature (°C) | 24.67 ± 0.04 | 24.63 ± 0.03 | 24.63 ± 0.04 |
| Conductivity (mS/cm) | 57.93 ± 0. 14 | 57.67 ± 0.09 | 57.90 ± 0.07 |
| Salinity (ppt) | 38.93 ± 008 | 38.77 ± 0.08 | 38.90 ± 0.07 |
| Oxygen saturation (%) | 89.20 ± 0.28 | 89.77 ± 0.15 | 89.43 ± 0.11 |
| Dissolved oxygen (mg/L) | 7.41 ± 0.02 | 7.46 ± 0.02 | 7.44 ± 0.02 |
| Sediment grain size distribution (%) | |||
| >2 mm | 1.2 ± 0.1 | 1.1 ± 0.2 | 1.3 ± 0.2 |
| <2 mm–>500 µm | 24.9 ± 1.9 | 24.6 ± 1.8 | 24.9 ± 1.4 |
| <500 µm–>125 µm | 73.8 ± 2.0 | 74.1 ± 2.0 | 73.7 ± 1.4 |
| <125 µm–>56 µm | 0.11 ± 0.01 | 0.1 ± 0.03 | 0.1 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerrato, M.; Mir-Rosselló, P.M.; Ferriol, P.; Gil, L.; Monserrat-Mesquida, M.; Tejada, S.; Pinya, S.; Sureda, A. Oxidative Stress Response in the Seaweed Padina pavonica Associated with the Invasive Halimeda incrassata and Penicillus capitatus. Water 2023, 15, 557. https://doi.org/10.3390/w15030557
Cerrato M, Mir-Rosselló PM, Ferriol P, Gil L, Monserrat-Mesquida M, Tejada S, Pinya S, Sureda A. Oxidative Stress Response in the Seaweed Padina pavonica Associated with the Invasive Halimeda incrassata and Penicillus capitatus. Water. 2023; 15(3):557. https://doi.org/10.3390/w15030557
Chicago/Turabian StyleCerrato, Marcello, Pere M. Mir-Rosselló, Pere Ferriol, Lorenzo Gil, Margalida Monserrat-Mesquida, Silvia Tejada, Samuel Pinya, and Antoni Sureda. 2023. "Oxidative Stress Response in the Seaweed Padina pavonica Associated with the Invasive Halimeda incrassata and Penicillus capitatus" Water 15, no. 3: 557. https://doi.org/10.3390/w15030557