
Water Stress Induced Changes in Root Traits and Yield of Irrigated Rice under Subtropical Condition

 ,
,  ,
, 
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site and Plant Materials
2.2. Experimental Design and Crop Management
2.3. Determination of Root Phenological and Physiological Traits
2.3.1. Root Number (RN)
2.3.2. Root Length (RL, cm)
2.3.3. Root Volume (RV, cm3 hill−1)
2.3.4. Root Porosity (RP, %)
2.3.5. Plant Physiological Traits
Crop Growth Rate (CGR, gm−2 day−1)
Relative Growth Rate (RGR, mg g−1 day−1)
Net Assimilation Rate (NAR, mg m−2 day−1)
Total Dry Matter (TDM)
2.4. Yield and Yield Components
2.5. Statistical Analysis
3. Results
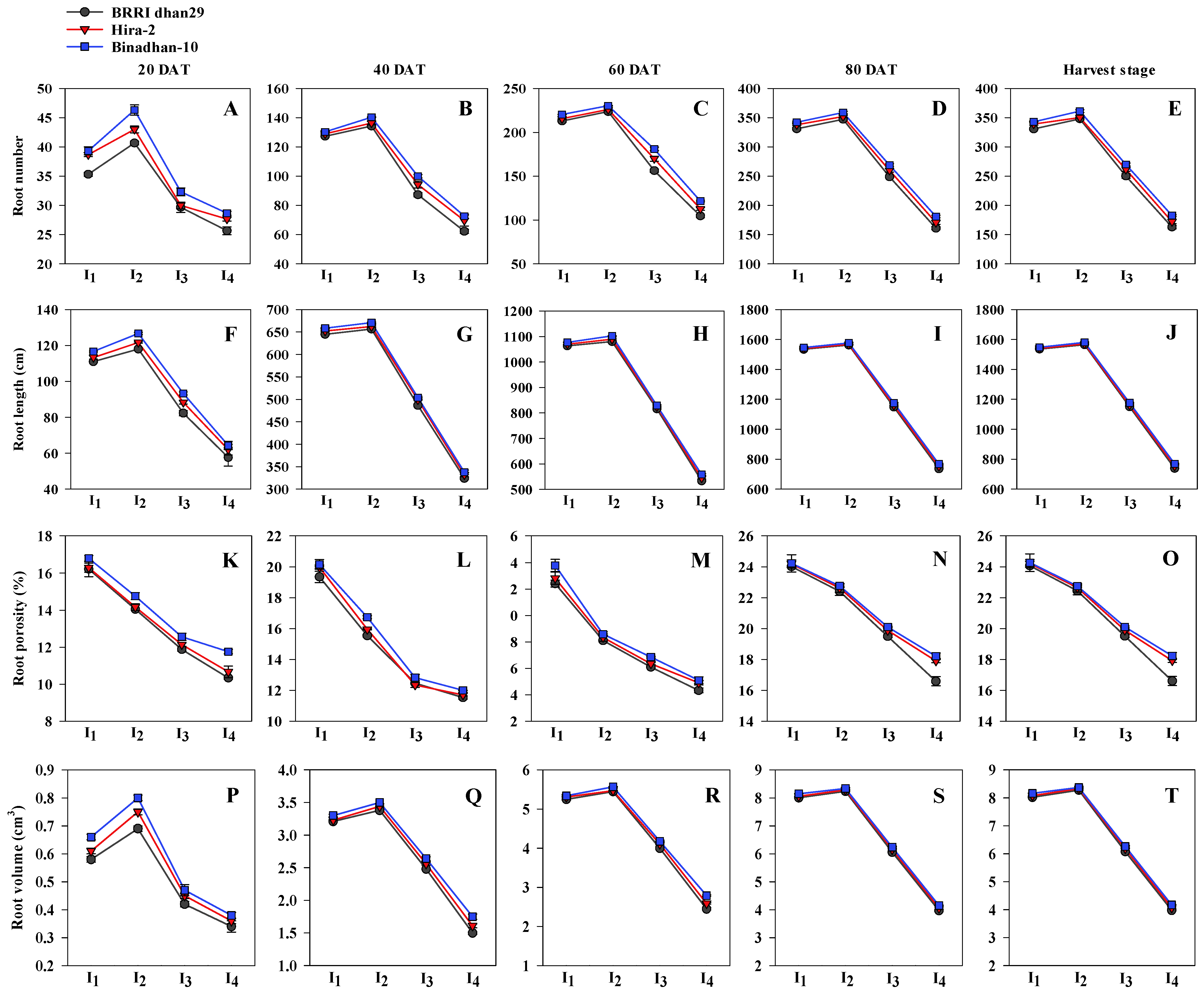
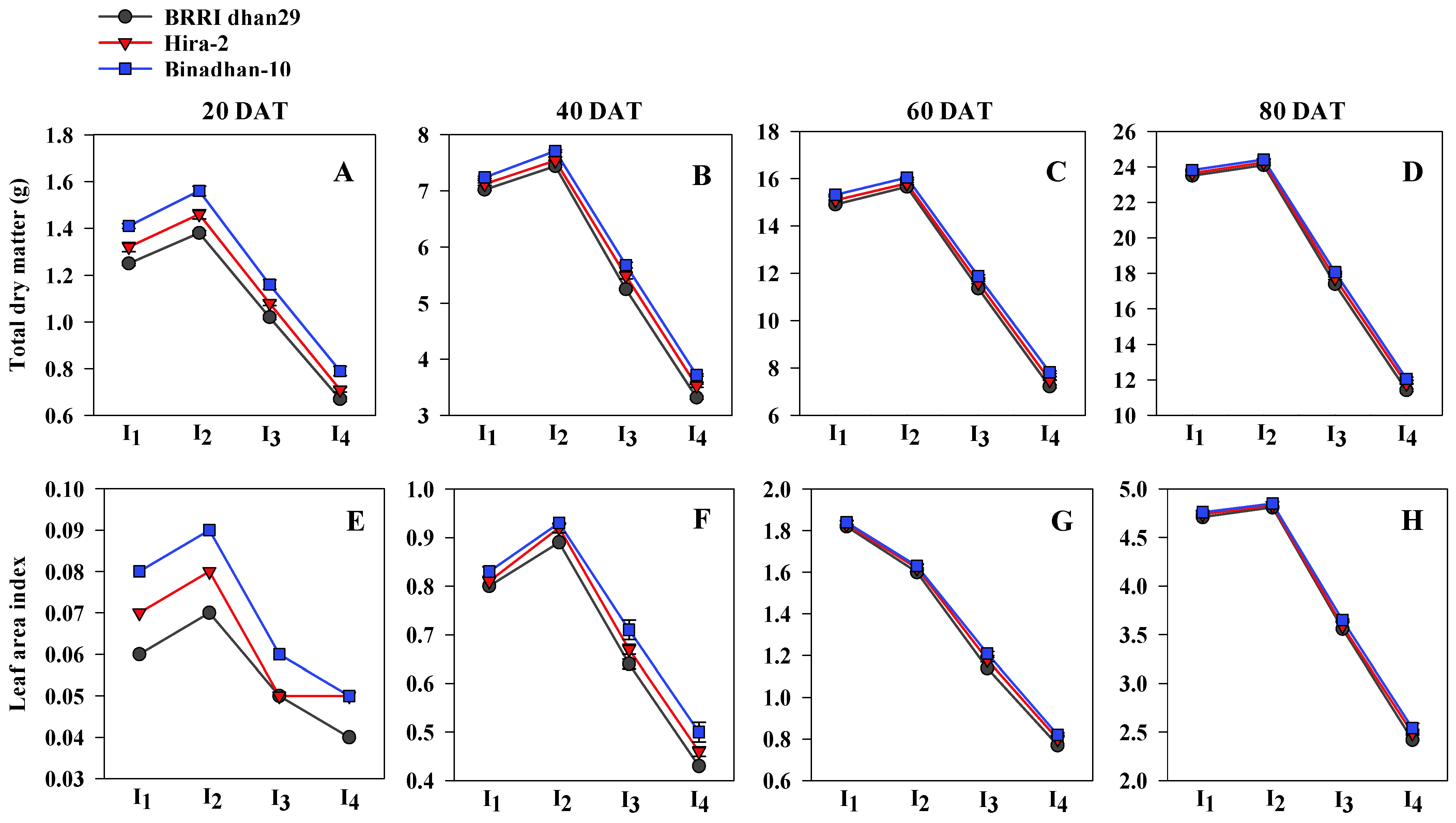
3.1. Root Morphological Traits, Total Dry Matter and Leaf Area Index
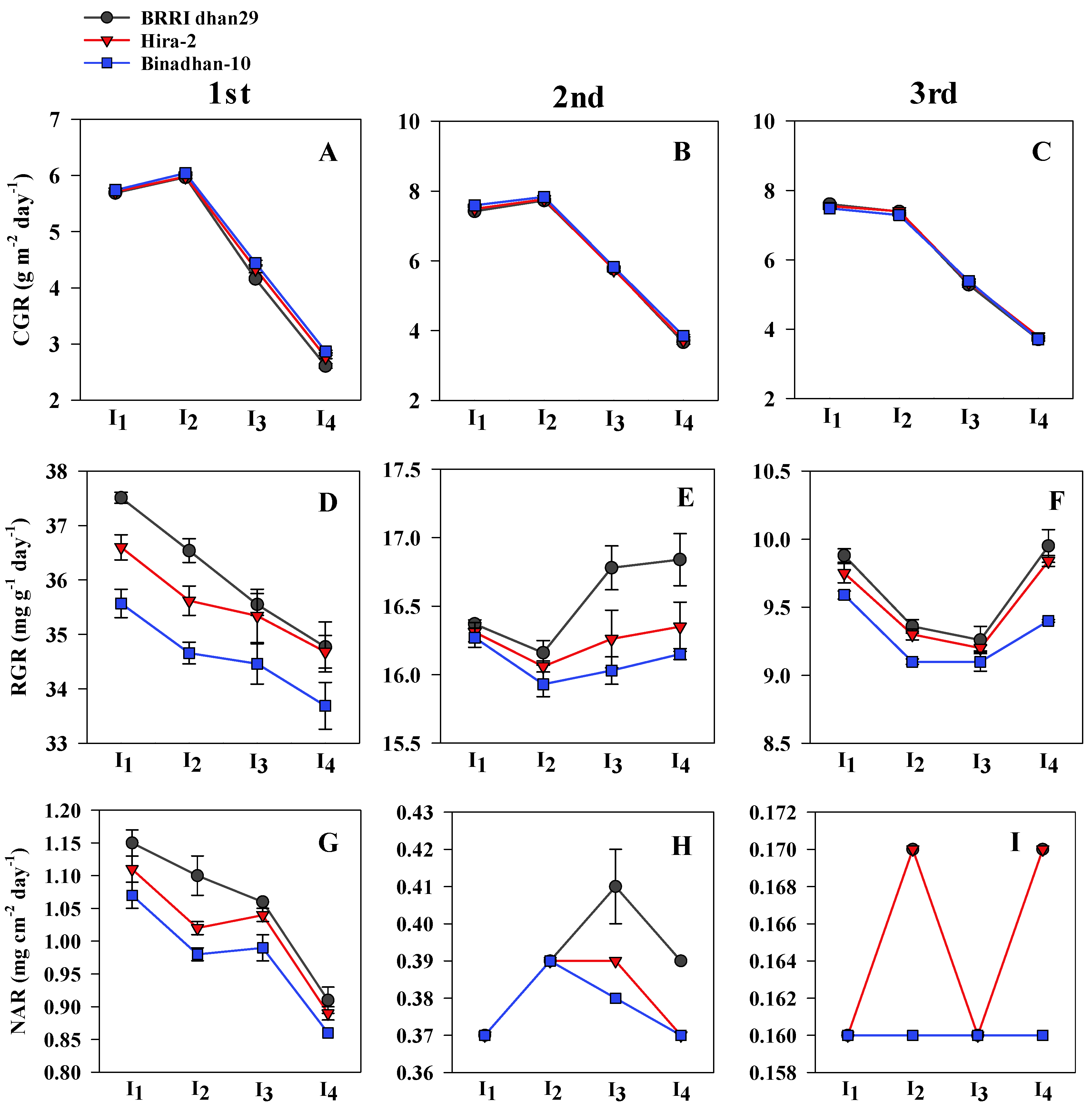
3.2. Growth Parameters
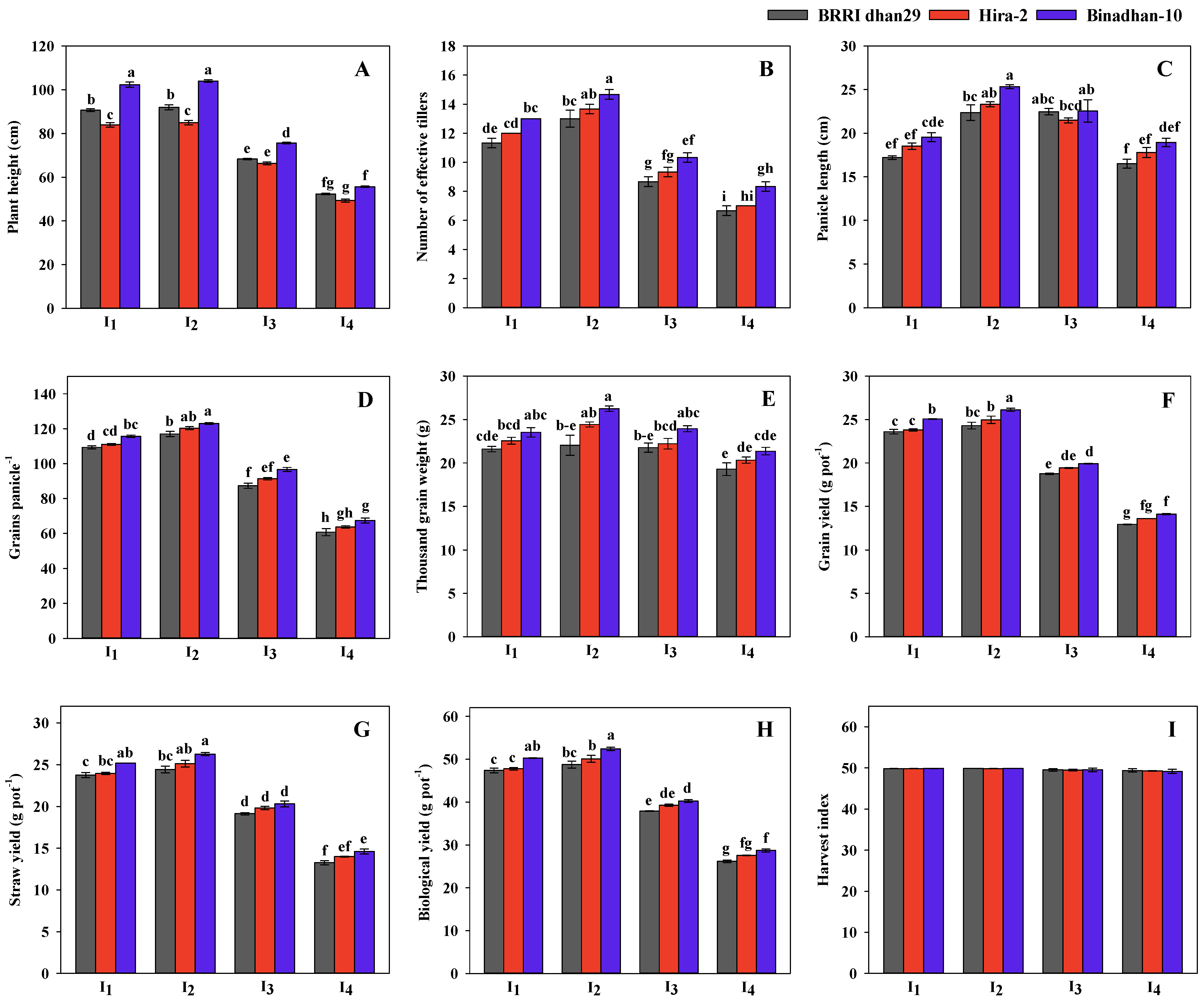
3.3. Grain Yield and Yield Component of the Rice Varieties
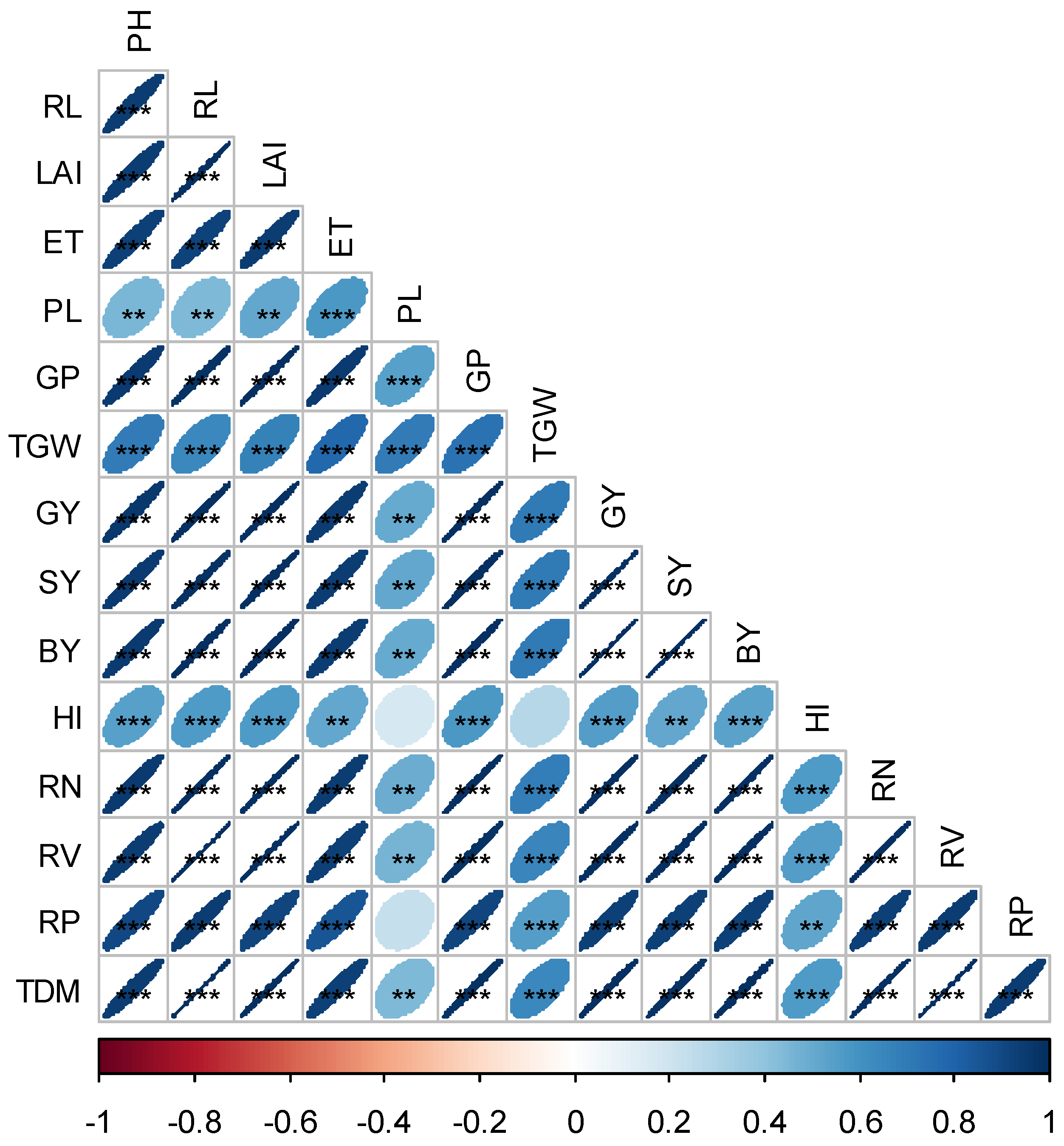
3.4. Relationship among Root Traits, Growth Parameters, Yield and Yield Attributes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aquastat. FAO Global Information System on Water and Agriculture. Available online: https://www.fao.org/aquastat/en/overview/ (accessed on 20 June 2022).
- Kang, S.; Hao, X.; Du, T.; Tong, L.; Su, X.; Lu, H.; Li, X.; Huo, Z.; Li, S.; Ding, R. Improving agricultural water productivity to ensure food security in China under changing environment: From research to practice. Agric. Water Manag. 2017, 179, 5–17. [Google Scholar] [CrossRef]
- Hassan, W.H.; Hussein, H.H.; Nile, B.K. The effect of climate change on groundwater recharge in unconfined aquifers in the western desert of Iraq. Groundw. Sustain. Dev. 2022, 16, 100700. [Google Scholar] [CrossRef]
- Hassan, S.F.; Hameed, K.A.; Ethafa, A.K.H.; Kadim, A.N.; Abbod, A.H.Y.; Ali, A.R.H.; Khalil, F.I. Response of three rice cultivars to the intermittent irrigation in Southern Iraq. Int. J. Appl. Agric. 2015, 1, 36–41. [Google Scholar] [CrossRef]
- Thakur, A.K.; Mohanty, R.K.; Patil, D.U.; Kumar, A. Impact of water management on yield and water productivity with System of Rice Intensification (SRI) and conventional transplanting system in rice. Paddy Water Environ. 2014, 12, 413–424. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, J. Crop management techniques to enhance harvest index in rice. J. Exp. Bot. 2010, 61, 3177–3189. [Google Scholar] [CrossRef]
- Khepar, S.; Yadav, A.; Sondhi, S.; Siag, M. Water balance model for paddy field under intermittent irrigation practices. Irrig. Sci. 2000, 19, 199–208. [Google Scholar] [CrossRef]
- Tuong, T.P.; Bouman, B.A.M.; Mortimer, M. More rice less water, integrated approaches for increasing water productivity in irrigated rice-based systems in Asia. Plant Prod. Sci. 2005, 8, 231–241. [Google Scholar] [CrossRef]
- Li, C.; Salas, W.; DeAngelo, B.; Rose, S. Assessing alternatives for mitigating net greenhouse gas emissions and increasing yields from rice production in China over the next twenty years. J. Environ. Qual. 2006, 35, 1554–1565. [Google Scholar] [CrossRef] [PubMed]
- Mahmod, I.F.; Barakhah, S.S.; Osman, N.; Omar, O. Physiological response of local rice varieties to aerobic condition. Int. J. Agric. Biol. 2014, 16, 738–744. [Google Scholar]
- Raeisi, L.G.; Morid, S.; Delavar, M.; Srinivasan, R. Effect and side-effect assessment of different agricultural water saving measures in an integrated framework. Agric. Water Manag. 2019, 223, 105685. [Google Scholar] [CrossRef]
- Hassan, W.H.; Nile, B.K.; Mahdi, K.; Wesseling, J.; Ritsema, C. A Feasibility Assessment of Potential Artificial Recharge for Increasing Agricultural Areas in the Kerbala Desert in Iraq Using Numerical Groundwater Modeling. Water 2021, 13, 3167. [Google Scholar] [CrossRef]
- Ota, R.; Ohkubo, Y.; Yamashita, Y.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile CEPD-like 2 integrates shoot nitrogen status to systemically regulate nitrate uptake in Arabidopsis. Nat. Commun. 2020, 11, 641. Available online: https://www.nature.com/articles/s41467-020-14440-8 (accessed on 24 January 2023). [CrossRef]
- Kreszies, T.; Schreiber, L.; Ranathunge, K. Suberized transport barriers in Arabidopsis, barley and rice roots: From the model plant to crop species. J. Plant Physiol. 2018, 227, 75–83. [Google Scholar] [CrossRef]
- Ichsan, C.N.; Basyah, B.; Zakaria, S.; Efendia, E. Differences of water status and relationship with roots growth and yield of rice under water stress. Sys. Rev. Pharm. 2020, 11, 611–618. [Google Scholar] [CrossRef]
- Geng, D.; Chen, P.; Shen, X.; Zhang, Y.; Li, X.; Jiang, L.; Xie, Y.; Niu, C.; Zhang, J.; Huang, X.; et al. Mdmyb88 and Mdmyb124 enhance drought tolerance by modulating root vessels and cell walls in apple. Plant Physiol. 2018, 178, 1296–1309. [Google Scholar] [CrossRef]
- Hussain, M.; Latif, A.; Hassan, W.; Farooq, S.; Hussain, S.; Ahmad, S.; Nawaz, A. Maize hybrids with well-developed root system perform better under deficit supplemental irrigation. Soil Environ. 2019, 38, 203–213. [Google Scholar] [CrossRef]
- Hazman, M.Y.; Brown, K.M. Progressive drought alters architectural and anatomical traits of rice roots. Rice 2018, 11, 62. [Google Scholar] [CrossRef]
- Shi, Q.; Zeng, X.; Li, M.; Tan, X.; Xu, F. Effects of different water management practices on rice growth. In Water-Wise Rice Production; Bouman, B.A.M., Hengsdijk, H., Hardy, B., Bindraban, P.S., Tuong, T.P., Ladha, J.L., Eds.; International Rice Research Institute: Manilla, Phillipine, 2002; pp. 3–14. [Google Scholar]
- Mishra, A. Intermittent irrigation enhances morphological and physiolog-ical efficiency of rice plants. Agriculture 2012, 58, 121–130. [Google Scholar] [CrossRef]
- Fang, S.; Clark, R.T.; Zheng, Y.; Iyer-Pascuzzi, A.S.; Weitz, J.S.; Kochian, L.V.; Edelsbrunner, H.; Liao, H.; Benfey, P.N. Genotypic recognition and spatial responses by rice roots. Proc. Natl. Acad. Sci. USA 2013, 110, 2670–2675. [Google Scholar] [CrossRef]
- Zhu, J.; Ingram, P.A.; Benfey, P.N.; Elich, T. From lab to field, new approaches to phenotyping root system architecture. CurrOpin Plant Biol. 2011, 14, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Takehisa, H.; Sato, Y.; Igarashi, M.; Abiko, T.; Antonio, B.A.; Kamatsuki, K.; Minami, H.; Namiki, N.; Inukai, Y.; Nakazono, M.; et al. Genome-wide transcriptome dissection of the rice root system: Implications for developmental and physiological functions. Plant J. 2012, 69, 126–140. [Google Scholar] [CrossRef]
- Zhang, F.; Zhou, Y.; Zhang, M.; Luo, X.; Xie, J. Effects of drought stress on global gene expression profile in leaf and root samples of Dongxiang wild rice (Oryzarufipogon). Biosci. Rep. 2017, 37, BSR20160509. [Google Scholar] [CrossRef]
- Ramireddy, E.; Hosseini, S.A.; Eggert, K.; Gillandt, S.; Gnad, H.; Von Wirén, N.; Schmülling, T. Root engineering in barley: Increasing cytokinin degradation produces a larger root system, mineral enrichment in the shoot and improved drought tolerance. Plant Physiol. 2018, 177, 1078–1095. [Google Scholar] [CrossRef] [PubMed]
- Beyrouty, C.A.; Wells, B.R.; Norman, R.J.; Teo, Y.H.; Gbur, E.E. Distribution and dynamics of the rice root system. In Proceedings of the 4th JSRR Symposium (Theme: Root System Management That Leads to Maximize Rice Yields), Tokyo, Japan, 11–12 September 1997; Abe, J., Morita, S., Eds.; University of Tokyo: Tokyo, Japan, 1997. [Google Scholar]
- Bouman, B.A.M.; Toung, T.P. Field water management to save water and increase its productivity in irrigated lowland rice. Agric. Water Manag. 2001, 49, 11–30. [Google Scholar] [CrossRef]
- Azhiri-Sigari, T.; Yamauchi, A.; Kamoshita, A.; Wade, L.J. Genotypic variation in response of rainfed lowland rice to drought and rewatering. II. Root growth. Plant Prod. Sci. 2000, 3, 180–188. [Google Scholar] [CrossRef]
- Kamoshita, A.; Wade, L.J.; Yamauchi, A. Genotypic variation in response of rainfed lowland rice to drought and rewatering. III. Water extraction during the drought period. Plant Prod. Sci. 2000, 3, 189–196. [Google Scholar] [CrossRef]
- Kato, Y.; Okami, M. Root morphology, hydraulic conductivity and plant water relations of high-yielding rice grown under aerobic conditions. Ann. Bot. 2011, 108, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Das, D.K.; Jat, R.L. Influence of three soil-water regimes on root porosity and growth of four rice varieties. Agron. J. 1977, 69, 197–200. [Google Scholar] [CrossRef]
- Ravichandran, V.K.; Nayar, V.; Prakash, K.C. An Evaluation of the SRI on Increasing Yield, Water Productivity and Profitability; Experiences from TN-IAMWARM Project. Irrigat. Drainag. Sys. Eng. 2015, 4, 137. [Google Scholar] [CrossRef]
- Castillo, E.G.; Buresh, R.J.; Ingram, K.T. Lowland rice yield as affected by timing of water deficit and nitrogen fertilization. Agron. J. 1992, 84, 152–159. [Google Scholar] [CrossRef]
- Borrell, A.K.; Garside, A.L.; Fukai, S. Improving efficiency of water use for irrigated rice in a semi-arid tropical. Field Crops Res. 1997, 52, 231–248. [Google Scholar] [CrossRef]
- Lilley, J.M.; Fukai, S. Effects of timing and severity of water deficit on four diverse rice cultivars. III. Phenological development, crop growth and grain yield. Field Crop Res. 1994, 37, 225–234. [Google Scholar] [CrossRef]
- Subagyono, K.; Darias, A.; Sumaini, E.; Kurnia, U. Water Management on Lowland Rice in: Lowland Rice and Management Technology. J. Biosci. Med. 2004, 2, 68–74. [Google Scholar]
- Hayashi, S.; Kamoshita, A.; Yamagishi, J. Effect of planting density on grain yield and water productivity of rice (Oryza sativa L.) grown in flooded and non-flooded fields in Japan. Plant Prod. Sci. 2006, 9, 298–311. [Google Scholar] [CrossRef]
- Kamoshita, A.; Ishikawa, M.; Abe, J.; Imoto, H. Evaluation of Water-Saving Rice-Winter Crop Rotation System in a Suburb of Tokyo. Plant Prod. Sci. 2007, 10, 219–231. [Google Scholar] [CrossRef]
- UNDP and FAO (United Nations Development Program and Food and Agriculture Organization). Land Resources Appraisal of Bangladesh for Agricultural Development.Report 2. Agro-Ecological Region of Bangladesh; United Nations Development Program and Food and Agricultural Organization: Dhaka, Bangladesh, 1988; pp. 212–221. [Google Scholar]
- FRG. Fertilizer Recommendation Guide; Bangladesh Agricultural Research Council (BARC): Farmgate, Dhaka, 2018; p. 274.
- Pantuwan, G.; Fukai, S.; Cooper, M.; O’Toole, J.C.; Sarkarung, S. Root traits to increase drought resistance in rainfed lowland rice. In Breeding Strategies for Rainfed Lowland Rice in Drought-Prone Environments; Fukai, S.M., Cooper, J., Salisbury, Eds.; Australian Centre for International Agricultural Research: Canberra, Australia, 1997; pp. 170–179. [Google Scholar]
- Kato, Y.; Kamoshita, A.; Yamagishi, J. Growth of Three Rice Cultivars (Oryza sativa L.) under Upland Conditions with Different Levels of Water Supply. Plant Prod. Sci. 2006, 9, 435–445. [Google Scholar] [CrossRef]
- Bridgit, A.J.; Potty, N.N. Influence of root characters on rice productivity in iron soils of Kerala. Int. Rice Res. News 2002, 27, 45–46. [Google Scholar]
- Jensen, C.R.; Luxmoore, R.J.; Van Gundy, S.D.; Stolzy, L.H. Root air space measurements by a pycnometer method. Agron. J. 1969, 61, 474–475. [Google Scholar] [CrossRef]
- Evans, G.C. Quantitative Analysis of Growth; Blackwell Scientific Publication: Oxford, UK; London, UK, 1972. [Google Scholar]
- Hunt, R. Plant Growth Analysis; Edward Arnold: London, UK, 1978; pp. 26–38. [Google Scholar]
- Yoshida, S.; Foron, D.A.; Cock, J.H. Laboratory Manual for Physiological Studies of Rice; International Rice Research Institute: Los Baños, Philippines, 1971; p. 70. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation Statistical Computation: Vienna, Austria, 2013; Available online: http://www.R-project.org/ (accessed on 24 January 2023). [CrossRef]
- Suriyan, C.; Yoo, Y.S.; Supaibulneatana, K. Water deficit stress in the productive stage of four indica rice (Oryza sativa L.) genotypes. Pak. J. Bot. 2010, 42, 3387–3398. [Google Scholar]
- Aulakh, M.S.; Malhi, S.S. Interactions of nitrogen with other nutrients and water: Effect on crop yield and quality, nutrient use efficiency, carbon sequestration, and environmental pollution. Adv. Agron. 2005, 86, 341–409. [Google Scholar] [CrossRef]
- Wang, X.B.; Zhou, B.Y.; Sun, X.F.; Yue, Y.; Ma, W.; Zhao, M. Soil tillage management affects maize grain yield by regulating spatial distribution coordination of roots, soil moisture and nitrogen status. PLoS ONE 2015, 10, e0129231. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R.; Eizenga, G.; McCling, A.; Kovach, M.; Tyagi, W.; Ali, M.L.; Tung, C.W.; Reynold, A. Improving photosynthetic efficiency for greater yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef]
- Sakamoto, T.; Morinaka, Y.; Ohnish, T.; Sunohara, H.; Fujioka, S.; Ueguchi-Tanaka, M.; Mizutani, M.; Sakata, K.; Takatsuto, S.; Yoshida, S.; et al. Erect leaves caused by brassinosteroid deficiency increase biomass production and grain yield in rice. Nat. Biotechnol. 2006, 24, 105–109. [Google Scholar] [CrossRef]
- Abu El-Ezz, A.E. Evaluating of Some Rice Cultivars under Different Water Regimes and Tillage Systems. Ph.D. Thesis, Tanta University, Tanta, Egypt, 2014. [Google Scholar]
- Tadesse, T.; Dechassa, N.; Bayu, W.; Gebeyehu, S. Impact of rain water management on growth and yield of rainfed lowland rice. J. Agric. Res. 2013, 2, 108–114. [Google Scholar]
- Vaezi, B.; Bavei, V.; Shiran, B. Screening of barley genotypes for drought tolerance by agro-physiological traits in field condition. Afr. J. Agric. Res. 2010, 5, 881–892. [Google Scholar] [CrossRef]
- Sharma, A.P.; Singh, S.P. Genotypic variation in photosynthesis and yield components in rice. Indian. J. Plant Physiol. 1994, 37, 188–189. [Google Scholar]
- Alam, M.M.; Ali, M.H.; Hasanuzzaman, M.; Nahar, K.; Islam, M.R. Dry matter partitioning in hybrid and inbred rice varieties under variable doses of phosphorus. Int. J. Agric. Sustain. 2009, 1, 10–19. [Google Scholar]
- McCauley, G.N. Sprinkler vs flood irrigation in traditional rice production regions of southeast Texas. Agron. J. 1990, 82, 677–683. [Google Scholar] [CrossRef]
- Mishra, H.S.; Rathore, T.R.; Pant, R.C. Root growth, water potential and yield of irrigated rice. Irrig. Sci. 1997, 17, 69–75. [Google Scholar] [CrossRef]
- Sah, R.N.; Mikkelsen, D.S. Availability and utilization of fertilizer nitrogen by rice under alternate flooding II: Effects on growth and nitrogen use efficiency. Plant Soil 1983, 75, 227–234. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef]
- Mollah, M.S.I.; Paul, N.K. Growth attributes of barley (Hordeumvulgare L.) in relation to soil moisture regimes and NPK fertilizers. J. Biosci. 2008, 16, 19–24. [Google Scholar] [CrossRef]
- Moayedi, A.A.; Boyce, A.N.; Barakbah, S.S.; Kafi, M. Water deficit-induced changes on growth parameters and radiation use efficiency of promising durum wheat genotypes. J. Food Agric. Environ. 2011, 9, 563–565. Available online: http://eprints.um.edu.my/7779/ (accessed on 24 January 2023).
- Alam, M.Z.; Rahman, M.S.; Hossain, M.E.; Azad, M.A.K.; Khan, M.R.H. Response of irrigation frequencies and different doses of N fertilization on the growth and yield of wheat. Pak. J. Biol. Sci. 2003, 6, 732–734. [Google Scholar] [CrossRef]
- Sun, Y.F.; Liang, J.M.; Ye, J.; Zhu, W.Y. Cultivation of super-high yielding rice plants. China Rice 1999, 5, 38–39. [Google Scholar]
- Stevenson, C.A.; Laidlaw, A.S. The effect of moisture stress on stolon and adventitious root development in white clover (Trifoliumrepens L.). Plant Soil 1985, 85, 249. [Google Scholar] [CrossRef]
- Cruz, R.T.; O’Toole, M.; Dingkhun, M.; Yambao, E.B.; Thangaraj, M.; De Datta, S.K. Shoot and root responses to water deficits in rainfed lowland rice. Aust. J. Plant Physiol. 1986, 13, 567–575. [Google Scholar] [CrossRef]
- Fageria, N.K. Yield physiology rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- Ascha, F.; Dingkuhn, M.; Sow, A.; Audebert, A. Drought-induced changes in rooting patterns and assimilate partitioning between root and shoot in upland rice. Field Crops Res. 2005, 93, 223–236. [Google Scholar] [CrossRef]
- Kima, A.S.; Chung, W.G.; Wang, Y.M. Improving irrigated lowland rice water use efficiency under saturated soil culture for adoption in tropical climate conditions. Water Environ. J. 2014, 6, 2830–2846. [Google Scholar] [CrossRef]
- Armstrong, W.; Drew, M.C. Root Growth and Metabolism under Oxygen Deficiency. Plant Roots: The Hidden Half, 3rd ed.; Marcel Dekker: New York, NY, USA, 2002; pp. 729–761. [Google Scholar]
- Evans, D.E. Aerenchymaformation. New Phytol. 2003, 161, 35–49. [Google Scholar] [CrossRef]
- Suralta, R.R.; Inukai, Y.; Yamauchi, A. Genotypic variations in responses of lateral root development to transient moisture stresses in rice cultivars. Plant Protect Sci. 2008, 11, 324–335. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Zhang, F.; van der Zee, S.E.A.T.M.; Hoffland, E. Tolerance to zinc deficiency in rice correlates with zinc uptake and translocation. Plant Soil. 2005, 278, 253–261. [Google Scholar] [CrossRef]
- Kaysar, M.S.; Sarker, U.K.; Monira, S.; Hossain, M.A.; Haque, M.S.; Somaddar, U.; Saha, G.; Chaki, A.K.; Uddin, M.R. Dissecting the relationship between root morphological traits and yield attributes in diverse rice cultivars under subtropical condition. Life 2022, 12, 1519. [Google Scholar] [CrossRef]
- Kaysar, M.S.; Sarker, U.K.; Monira, S.; Hossain, M.A.; Somaddar, U.; Saha, G.; Hossain, S.S.F.; Mokarroma, N.; Chaki, A.K.; Bhuiya, M.S.U.; et al. Optimum Nitrogen Application Acclimatizes Root Morpho-Physiological Traits and Yield Potential in Rice under Subtropical Conditions. Life 2022, 12, 2051. [Google Scholar] [CrossRef]
- Rajesh, V.; Thanunathan, K. Effect of seedling age, number and spacing on yield and nutrient uptake of traditional Kambamchambarice. Madras Agric. J. 2003, 90, 47–49. [Google Scholar]
- Dass, A.; Chandra, S. Effect of different components of SRI on yield, quality, nutrient accumulation and economics of rice in tarai belt of northern India. Indian J. Agron. 2012, 57, 250–254. [Google Scholar]
- Dass, A.; Chandra, S. Irrigation, spacing and cultivar effects on net photosynthetic rate, dry matter partitioning and productivity of rice under SRI in Mollisols of northern India. Exp. Agric. 2013, 49, 504–523. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety (V) | PH (cm) | ET (no.) | PL (cm) | GP | TGW (g) | GY (g pot−1) | SY (g pot−1) | HI (%) |
|---|---|---|---|---|---|---|---|---|
| V1 | 75.83 b | 9.92 c | 19.64 b | 93.58 c | 21.17 b | 19.90 c | 20.15 c | 49.65 |
| V2 | 71.17 c | 10.50 b | 20.28 b | 96.58 b | 22.39 ab | 20.45 b | 20.72 b | 49.61 |
| V3 | 84.42 a | 11.58 a | 21.59 a | 100.67 a | 23.78 a | 21.31 a | 21.59 a | 49.61 |
| Irrigation (I) | ||||||||
| I0 | 92.33 a | 12.11 b | 18.42 c | 112.00 b | 22.56 b | 24.16 b | 24.30 b | 49.86 a |
| I2 | 93.67 a | 13.78 a | 23.67 a | 120.11 a | 24.25 a | 25.13 a | 25.28 a | 49.86 a |
| I3 | 70.11 b | 9.44 c | 22.16 b | 91.78 c | 22.64 b | 19.37 c | 19.76 c | 49.51 ab |
| I4 | 52.44 c | 7.33d | 17.75 c | 63.89 d | 20.33 c | 13.54 d | 13.94 d | 49.27 b |
| ANOVA | ||||||||
| V | ** | ** | ** | ** | ** | ** | ** | NS |
| I | ** | ** | ** | ** | ** | ** | ** | ** |
| CV (%) | 1.70 | 5.18 | 4.91 | 2.04 | 4.27 | 1.67 | 2.16 | 0.87 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaysar, M.S.; Sarker, U.K.; Monira, S.; Hossain, M.A.; Mokarroma, N.; Somaddar, U.; Saha, G.; Hossain, S.S.F.; Chaki, A.K.; Uddin, M.R. Water Stress Induced Changes in Root Traits and Yield of Irrigated Rice under Subtropical Condition. Water 2023, 15, 618. https://doi.org/10.3390/w15040618
Kaysar MS, Sarker UK, Monira S, Hossain MA, Mokarroma N, Somaddar U, Saha G, Hossain SSF, Chaki AK, Uddin MR. Water Stress Induced Changes in Root Traits and Yield of Irrigated Rice under Subtropical Condition. Water. 2023; 15(4):618. https://doi.org/10.3390/w15040618
Chicago/Turabian StyleKaysar, Md. Salahuddin, Uttam Kumer Sarker, Sirajam Monira, Md. Alamgir Hossain, Nadira Mokarroma, Uzzal Somaddar, Gopal Saha, S. S. Farhana Hossain, Apurbo Kumar Chaki, and Md. Romij Uddin. 2023. "Water Stress Induced Changes in Root Traits and Yield of Irrigated Rice under Subtropical Condition" Water 15, no. 4: 618. https://doi.org/10.3390/w15040618