
Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions

1
Biological Sciences Department, Faculty of Science, King Abdulaziz University, P.O. Box 80203, Jeddah 21589, Saudi Arabia
2
Botany Department, Faculty of Science, University of Mansoura, Mansoura 35516, Egypt
*
Authors to whom correspondence should be addressed.
Water 2023, 15(4), 664; https://doi.org/10.3390/w15040664
Submission received: 10 December 2022
/
Revised: 1 February 2023
/
Accepted: 6 February 2023
/
Published: 8 February 2023
(This article belongs to the Special Issue Smart Microalgal Biotechnology)
Abstract
:The growth response and biological activity of the cyanobacterium Oscillatoria sancta were investigated in starvation conditions. Oscillatoria sancta growth potential was examined on BG11 and Zarrouk’s media. Zarrouk’s medium supported the maximum growth of the test cyanobacterium. Zarrouk’s medium composition was modified by excluding CaCl2·2H2O, NaCl, EDTA (Na), micronutrients, and replacing sodium nitrate with urea. Using Zarrouk’s medium and three different concentrations of modified Zarrouk’s media (Treatments 1–3), the growth response of Oscillatoria sancta (MZ366482) was examined and compared. Zarrouk’s medium and modified Zarrouk’s medium at 12.5% nutrient concentration had non-significant differences in both the dry weight biomass and total protein of Oscillatoria sancta. Oscillatoria sancta crude biomass extracts grown on Zarrouk’s and modified Zarrouk’s media (T3) inhibited human breast cancer, pathogenic bacteria, and acetylcholinesterase activity. Oscillatoria sancta grown on T3 showed the most potency against MDA-MB-231 cells with an IC50 of 165.2 µg mL−1, antibacterial activity only against Bacillus cereus (17.2 mm) and Staphylococcus aureus (15.3 mm), and acetylcholinesterase inhibition activity by 60.7%. Thus, it is advisable to use the 12.5% nutrient concentration of modified Zarrouk’s medium as a reduced-cost medium for mass cultivation of Oscillatoria sancta with potential anticancer, antibacterial diseases, and anti-Alzheimer purposes.
1. Introduction
Microalgae are rich sources of secondary metabolites that are naturally synthesized and can be used in the pharmaceutical, cosmetic, and healthcare industries [1]. Depending on stress conditions, different suites of microalgal secondary metabolites with significant structural variations can be produced. As cyanobacterial strains directly relate to eukaryotic photosynthetic organisms, such as plants, they can serve as a model to investigate the stress response in this context [2,3].
Cyanobacteria can produce different suites of secondary metabolites in response to the different biotic and abiotic stresses to defend their cells against grazers and predators, as well as antioxidant, chemosensory, and photoprotection functions [4]. Under nutrient stress conditions, cyanobacteria undergo significant alterations and develop sophisticated coping mechanisms that allow them to endure stress using a “stand-by” energy metabolism [5]. Cyanobacteria synthesized poly-b-hydroxybutyrate (PHB) inclusions, cyanophycin, and phycobiliproteins under nitrogen, phosphorus, and sulfur starvation conditions, exhibiting anti-oxidant, anti-inflammatory, neuroprotective, hepatoprotective, and anticancer effects [6,7,8,9,10,11].
Furthermore, L-valine, L-tryptophan, and methionine were synthesized in response to carbon stress, and are efficient against Staphylococcus and Clostridium bacteria. However, phycobiliprotein production and antioxidant activity increased under nitrogen stress [12,13,14,15,16]. Anatoxins, flavodoxin, and iron chelators are also produced by cyanobacteria when there is a lack of iron, which increases their antimicrobial action [17,18].
Additionally, an extensive list of indole alkaloids’ secondary metabolites can be synthesized by cyanobacteria to maintain their survival under starvation conditions such as norharmane (9H-pyrido(3,4-b) indole), which, when excreted by Nodularia harveyana, exhibited high algicidal activity [19]. Nostocarboline from Nostoc 78–12A and anatoxin-a from Nostoc sp. could be used as an acetylcholinesterase inhibitor for the treatment of neuronal diseases like Alzheimer’s [20,21]. Furthermore, hapalindole synthesized from Hapalosiphon fontinalis exhibits antibacterial and anti-tuberculosis effects [22]. Phenolics, bromophenols, terpenoids, cyclic dep-sipeptides, lipoprotein, and isonitrile-containing indole alkaloids are produced by different strains of cyanobacteria and are efficient as antifungal [23].
They also produce several effective compounds against cancer progression by stimulating apoptotic death, such as synthadotin [24], cryptophycin, and curacin [25]. Some authors [26,27] reported using marine blue-green algae for anticancer compounds. Numerous researchers have investigated the bioactive components of cyanobacteria and found that they have pharmacological characteristics. However, more in vivo and in vitro investigations employing various animal models and clinical studies will be necessary to bring cyanobacteria and their active components into translational mode [28]
The present work investigates the effect of nutrient deficiency on the biochemical composition and activity of Oscillatoria sancta crude extract against MDA-MB-231 cell line, some pathogenic bacteria, and AChE inhibition to evaluate the possible activity against breast cancer, Alzheimer’s disease, and bacterial diseases.
2. Materials and Methods
2.1. The Isolate
The cyanobacterium used in this study was obtained from the culture collection of the phycology laboratory, Faculty of Science, Mansoura University, Egypt. Cultures were maintained in blue–green (BG11) nutrient medium at 28 ± 2 °C under continuous illumination of 50 µmol photons m−2 s−1 [29].
2.2. Molecular Identification
The test isolate’s genomic DNA was isolated using the Thermo Scientific GeneJET Genomic DNA Purification Kit (Thermo Scientific Sciences, Hilden, Germany). Polymerase chain reaction (PCR) amplification was performed for purified DNA slightly modified from that used by [30]. Specific primers targeting part of the 16S rRNA gene: (forward primer) CYA106F (5′ CGG ACG GGT GAG TAA CGC GTGT 3′) and (reverse primer) CYA781R (5′ GAC TAC TGG GGT ATC TAA TCC CAT 3′), were used. The PCR reaction was performed in a total volume of 25 μL reaction mixtures containing the following: 3 μL from forward and reverse primer (10 pmol/μL from each), dNTPs 2.5 μL, Taq DNA polymerase 0.2 μL (Fermentas, Hilden, Germany), 10× Taq buffer 5 μL, 8 μL nuclease-free water, and 1 μL template DNA (equivalent to 10 ng). The mixture was subjected to the following amplification conditions: an initial denaturalizing step of 4 min at 94 °C, 40 cycles of denaturation for 30 s at 94 °C, annealing for 30 s at 48 °C, and extension for 1 min at 72 °C, ending with a final extension step at 72 °C for 7 min. Amplified DNA fragments were estimated using a DNA ladder of 2 kb. The PCR products were electrophoresed on 2% (w/v) agarose gel in 0.5× TBE buffer at 90 mV for 3 h 30 min, using the 2 kb plus DNA Ladder (Gibco-BRL) as a molecular size marker. The target band was excised and purified with a DNA gel Extraction Kit (Fermentas, Hilden, Germany). The BLAST tool and MEGA ver. 5 software were used to perform multiple alignments and phylogenetic analyses on the gene sequences of Oscillatoria strains (16S rRNA) [31].
2.3. Impact of Various Nutrient Media on the Growth Potential of Oscillatoria sancta
The growth of Oscillatoria sancta was assessed using the BG11 medium [29] and Zarrouk’s medium [32] to compare the growth potential of Oscillatoria sancta in the two distinct nutritional media. In this research, we employed sterile cultures with a pH of 8 and maintained the temperature at 28 ± 2 °C for two weeks while the cells were exposed to 50 µmol photons m−2 s−1. The optical density of the inoculum at the start was 0.18 (OD682).
2.4. Modification of the Chemical Composition of Zarrouk’s Medium
The experimental results in this study indicated that Zarrouk’s medium supported the maximum biomass production of O. sancta. Thus, it was chosen for further growth experiments. Modified Zarrouk’s medium was devoid of CaCl2·2H2O, EDTA (Na), NaCl, and micronutrients. Sodium nitrate (NaNO3) was substituted by urea (CO (NH2)2) (0.8 g L−1) (Table 1). Three-grade nutrient concentrations (100%, 50%, and 12.5%) of modified Zarrouk’s medium representing T1, T2, and T3 were prepared for testing and comparing growth with standard Zarrouk’s medium (control). The initial inoculum was comparable to 0.05 g L−1 dry wt. The goal was to examine the growth potential of O. sancta under starvation conditions.
2.5. Growth Assessment
The growth of O. sancta was evaluated using a combination of gravimetric dry-weight biomass and measurements of the chlorophyll a concentration [33]. The established O. sancta culture volume was filtered through a dry pre-weighed GF/C filter to estimate the dry weight. To obtain an average dry weight, we dried O. sancta biomass filters for 12 h in an oven at 105 °C before cooling them in a desiccator and weighing them (g L−1).
2.6. Growth Rate
A formula was used to calculate the O. sancta cultures’ specific growth rates according to Anderson [34] using the following Equation (1):
where μ is the specific growth rate and X1 and X2 are the chlorophyll a concentration (mg L−1) at times t1 and t2 in this experiment, respectively.
The divisions per day (Dd−1) were calculated following Equation (2):
2.7. Biomass Harvesting
Known volumes of Oscillatoria sancta cultures were collected after 14 days of growth and centrifuged at 2688× g for 10 min. Glass-distilled water was used to thoroughly clean the pellets before they were placed in a freezer and kept at −20 °C overnight. After the algal pellets had been frozen, the freeze-dried biomass was weighed using a four-decimal point balance [35].
2.8. Biochemical Properties
Biochemical properties include many parameters, such as the determination of total protein, carbohydrate, and lipid contents. Protein concentration was determined using the Bradford methods [36] and updated by Stoscheck [37]. Carbohydrate concentration was determined using the Hedge and Hofreiter method [38]. Lipid extraction was performed according to the exhaustive Soxhlet process of Sadasivam and Manickam [39].
2.9. Crude Extraction with Methylene Chloride/Methanol (1:1) v/v
Soxhlet extractors were used to extract algal biomass from a sample of 5 g freeze-dried powder wrapped in filter paper and gently squeezed [39].
2.10. Biological Activities of Oscillatoria sancta Crude Extract
2.10.1. Antibacterial Screening Assay
The antibacterial activity of Oscillatoria sancta methylene chloride was investigated using agar well diffusion; methylene chloride/methanol crude extracts were used [40]. Staphylococcus aureus, Bacillus cereus, Pseudomonas aeruginosa, and Escherichia coli were collected from the bacteriology laboratory of the Faculty of Science at Mansoura University, Egypt. Then, 8 mm wells on nutrient agar plates were made using a sterile cork borer and filled with 100 µL of O. sancta crude extracts (10 mg mL−1). Methylene chloride was employed with methanol serving as a negative check. At 37 ± 2 °C, the diameters of clean zones were assessed for antibacterial activity after 24 h.
2.10.2. In Vitro Acetylcholinesterase Activity Test
The activity of acetylcholinesterase (AChE) in the presence of O. sancta crude extracts was determined according to the method proposed by Refaay [41]. The assay was performed in a 96-well microplate containing 200 μL assay mixture containing 0.75 mM of 5, 5_-dithio-bis-[2-nitrobenzoic acid] (DTNB) and electric-eel AChE (0.0004 µg mL−1) in 50 mM Tris-HCl (pH 8). Then, 1.5 mM of acetylthiocholine iodide (ATCI) was added to the reaction mixture to start the reaction. The developed yellow color of 5-thio-2-nitrobenzoate (TNB) was measured at 405 nm. Then, 5 μL (10 µg mL−1) of O. sancta crude extracts were pre-incubated with the enzyme for one minute before initiating the reaction. The AChE activity was also performed in the presence of methylene chloride/methanol as the negative control and donepezil (10 µg mL−1) as the positive control. The AChE inhibition percent was calculated according to the following equation:
where A0 is the absorbance in the presence of the solvent only and A1 reflects the absorbance when the tested extract was utilized instead.
2.10.3. Evaluation of Anticancer Activity
Cell Line
The American Type Culture Collection (ATCC) breast cancer cell line (MDA-MB-231) was grown on Dulbecco’s Modification of Eagle’s Medium (DMEM) supplemented with 10% fetal bovine serum, penicillin, and streptomycin, and then incubated at 37 °C in a 95% relative humidified environment with 5% CO2 for 30 min.
In Vitro MTT Assay
The cytotoxicity of crude extracts of Oscillatoria sancta was examined utilizing the Mosmann method [42]. MDA-MB-231 cells were plated in 96-well microplates containing different doses of extract (0.125, 0.25–0.5, 0.5, 1, and 3 mg mL−1) at a concentration of 3 × 103 cells/well (including 2% antibiotics and 2% FBS). MDA-MB-231 cells cultured in a medium were used to create a parallel monitor. During a 72 hr. incubation period at 37 °C, wells were filled with MTT (5 mg mL−1) solution and incubated for another 4 h at 37 °C until violet (formazon) crystals emerged. Dimethyl sulfoxide (DMSO) was used to dissolve 50 μL of Formulazan crystals at 37 °C for 30 min. Using an ELISA reader, we determined an absorbance of 570 nm. For each extract, the MDA-MB-231 cells’ growth rate inhibition % was calculated according to the following equation:
Chart Pad Prism 8 software was used to calculate the IC50 value (the concentration that affects 50% of cell death) using data on the cell growth inhibition rate.
2.11. GC-MS Analysis
Gas chromatography (Aglient 6890) equipped with an Aglient mass spectrometric detector was used to determine the chemical composition of O. sancta crude extract. An electron impact ionization mode detector was used to accomplish mass spectrophotometric scanning between 50 and 500 m/z. Here, 70 e.v. was the ionizing power. The electron multiplier’s EM voltage was kept constant at 1250 V using auto-tune. The instrument was tuned by hand with perfluorotributyl amine (PFTBA). The mass spectral database Wiley and Wiley NIST was used to identifying the peaks.
2.12. Statistical Analysis
One-way analysis of variance (ANOVA) was performed on the data at p ≤ 0.05 to determine the significance of the findings using SPSS Ver. 20.
3. Results
3.1. Phylogenetic Analysis and Placement of Oscillatoria sancta
The dendrogram of phylogenetic analysis (Figure 1) indicated that the Egyptian isolate Oscillatoria sancta HSDM1 falls into a clade together with Oscillatoria sancta strain VMRJHK012020 (accession No. MW364271), maintaining a 100% bootstrap value and in conjunction with the help of the local pairwise orientation (http://www.ebi.ac.uk/Tools/psa/emboss_water/nucleotide.html) (accessed on 20 May 2022) using the Smith–Waterman algorithm of the two sequences. The test findings demonstrate that the two sequences are 99% similar. The Oscillatoria sancta HSDM 1 sequencing product was placed in the GenBank database with the entry number MZ366482.
3.2. Growth Assessment of Oscillatoria sancta on Different Nutrient Media
Oscillatoria sancta growth on BG11 and Zarrouk’s nutritional media was assessed using chlorophyll a as a concentration variation. Growth curves are illustrated in Figure 2. O. sancta maintained the highest chlorophyll a content at the end of the sixth day on Zarrouk’s medium and at the end of day eight on the BG11 medium at 1554 ± 31.6 and 889 ± 27.9 mg L−1, respectively.
3.3. Growth Rates of the Test Cyanobacterium Oscillatoria sancta
To perform an accurate growth comparison of O. sancta on both BG11 and Zarrouk’s media, the specific growth rate (µ) and divisions per day (Dd−1) were calculated. The results are shown in Figure 3. Zarrouk’s medium exhibited a highly significant (p ≤ 0.05) increment in specific growth rate (0.452 ± 0.029) and divisions per day (0.652 ± 0.034) of O. sancta by 1.68-fold compared with BG11 medium.
3.4. Oscillatoria sancta Growth in Modified Zarrouk’s Medium at Varying Nutrient Concentrations
Very slight and almost non-meaningful (p ≤ 0.05) differences were observed in the dry weight (g L−1) biomass of different O. sancta cultures grown at different modified Zarrouk’s medium concentrations (T1 to T3) compared with control Zarrouk’s medium (Table 2). The highest dry-weight biomass, 0.552 ± 0.002 g L−1, was recorded on standard Zarrouk’s medium and 0.509 ± 0.022 g L−1 was recorded at T3 (12.5% modified Zarrouk’s medium). Accordingly, a 12.5% change in Zarrouk’s medium concentration was selected for further growth test experiments.
3.5. Biochemical Composition of Oscillatoria sancta Grown on Both 12.5% Modified Zarrouk’s Medium and Standard Zarrouk’s Medium
The total carbohydrate, total protein, and total lipid contents of O. sancta dry biomass grown on both T3 (12.5% modified Zarrouk’s medium) and standard Zarrouk’s medium (control) are illustrated in Figure 4. There was no significant difference (p > 0.05) between O. sancta cultures cultured on T3 (47 ± 2.3%) and Zarrouk’s medium (50.3 ± 2.5%) in terms of protein content.
Carbohydrate content was relatively high at T3 (26.2 ± 1.31%) compared with the control (15.4 ± 0.7%). Nutrient concentrations did not affect the lipid content. Wt.% of total lipids was 3 ± 0.14% and 2.8 ± 0.14% of biomass grown on T3 and Zarrouk’s medium (control), respectively.
3.6. Anticancer Activity of Oscillatoria sancta Methylene Chloride/Methanol Crude Extracts against MDA-MB-231 Cell Line
Cell viability in the MDA-MB-231 cell line was assessed using the MTT colorimetric test using crude methanol extracts of O. sancta cultures cultivated on T3 and Zarrouk’s media. The extracts promoted MDA-MB-231 cell line anticancer activity in a dose-dependent manner (Figure 5). Compared with the conventional Zarrouk’s medium (control, 195 ± 2.3 μg mL−1), the O. Sancta crude extract cultivated in T3 demonstrated the strongest anticancer activity, with an IC50 value of 165.2 ± 0.54 μg mL−1.
The O. sancta crude extracts exhibited antibacterial activity only against Gram + ve, B. cereus, and S. aureus (Table 3). With inhibition zones of 17.2 ± 0.44 mm and 15.3 ± 0.58 mm, O. sancta crude extracts prepared under nutrient scarcity (T3) exhibited the best antibacterial activity against Gram + ve, B. cereus, and S. aureus.
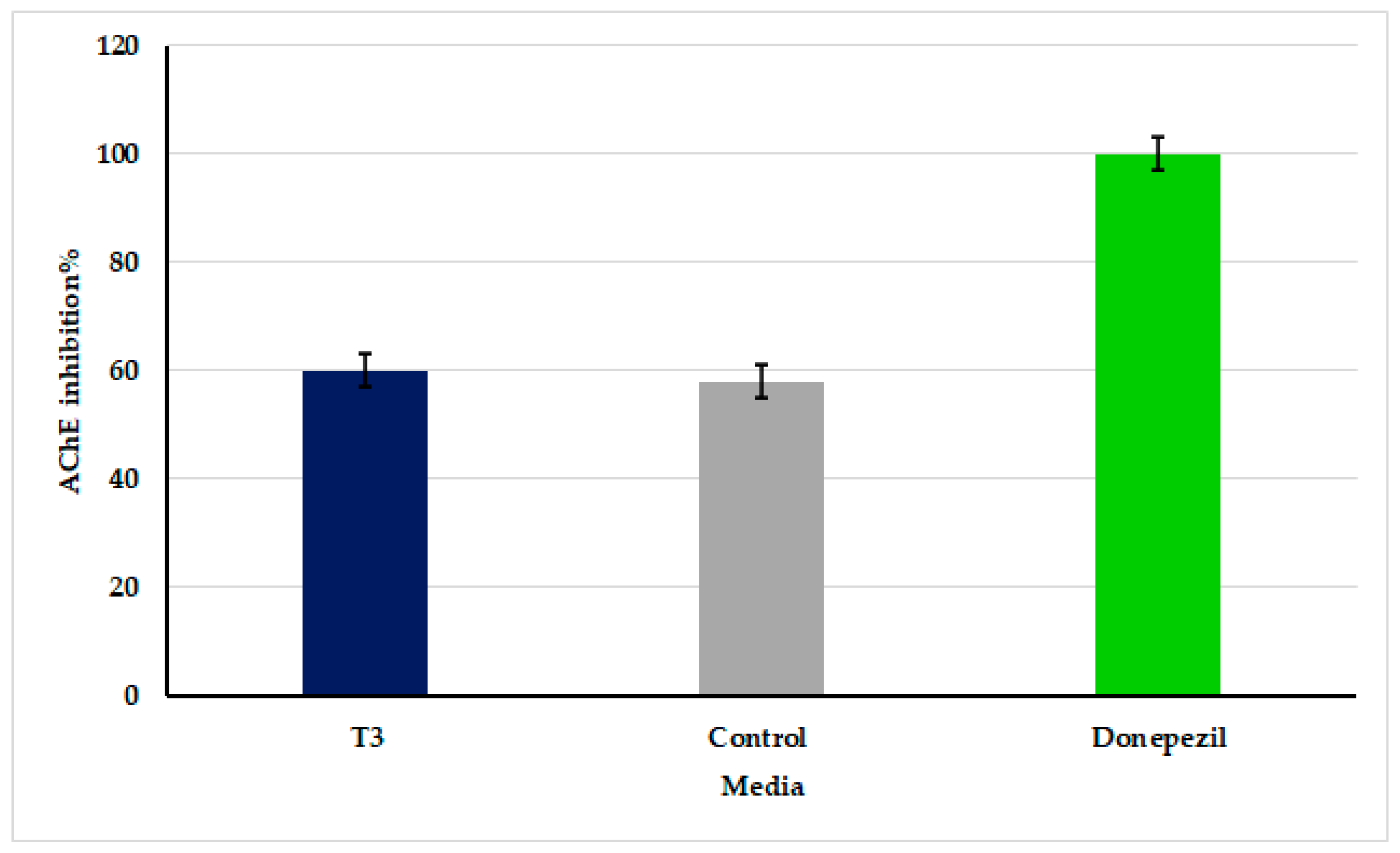
3.7. Inhibition of Acetylcholinesterase Activity by Oscillatoria sancta Crude Extracts
Inhibition% of AChE activity calculated of the tested extracts are presented in Figure 6, where donepezil was used as the positive control. Moderate significant (p ≤ 0.05) inhibition of AChE activity was shown in O. sancta crude extracts grown on both T3 and Zarrouk’s medium, by 60.7 ± 3.1% and 58 ± 2.1%, respectively, compared with donepezil, which was recorded as 100 ± 0.0%.
3.8. GC/MS
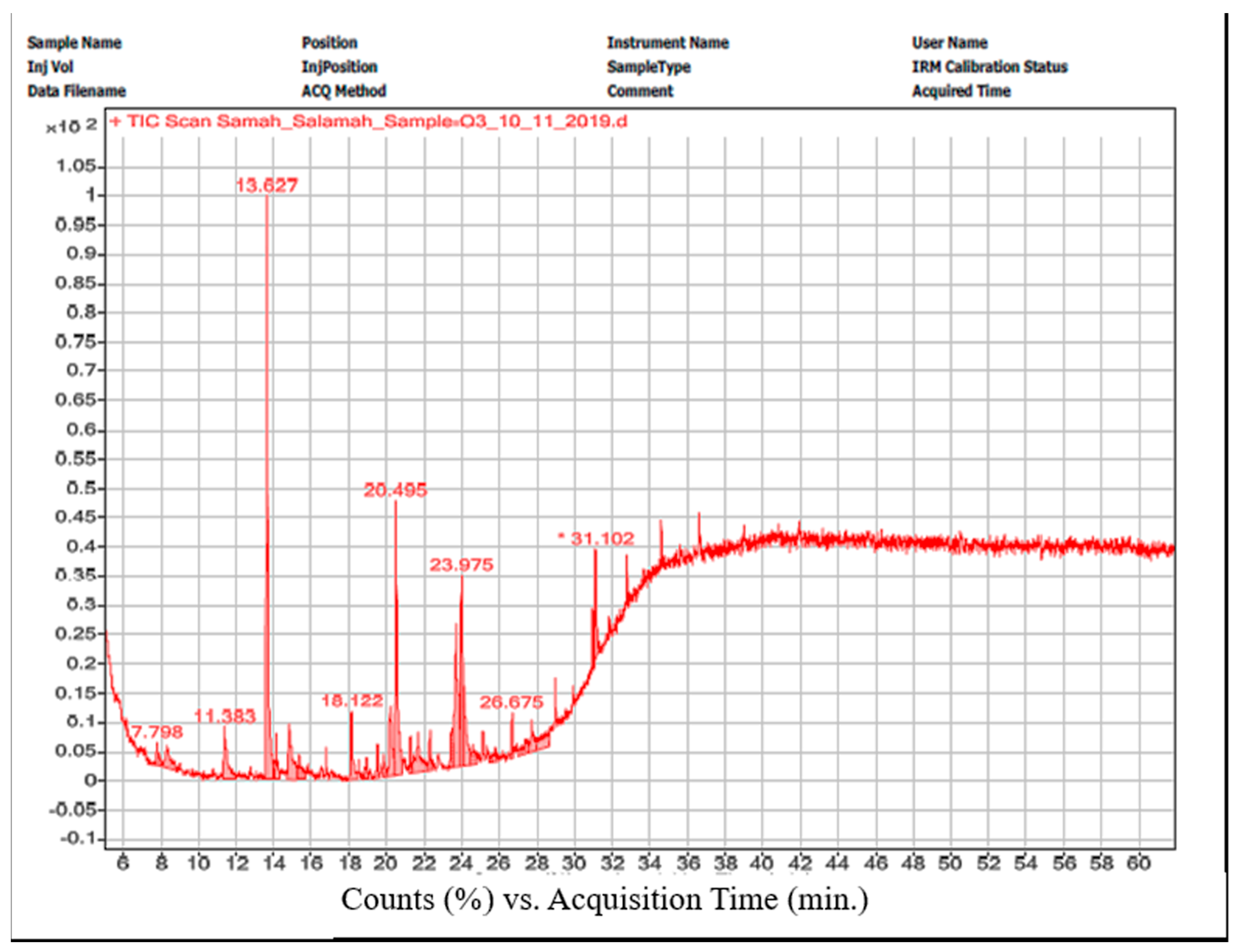
The gas chromatogram for O. sancta crude extract cultivated on T3 revealed ten peaks corresponding to ten compounds included (Figure 7). In Table 4, we recorded a complete list of chemical names, peak locations, retention times, and formulations. These substances comprise hydrocarbons, fatty acids, esters, and alkaloids.
4. Discussion
One of the major challenges influencing microalgae biomass production for high-value products or metabolites with applied purposes is the cost of nutrients. It is also possible for microalgae, particularly cyanobacteria, to develop high-value secondary compounds with a variety of medical applications in nutritional stress settings.
As seen in Figure 2 and Figure 3, Zarrouk’s medium exhibited the maximum biomass reproduction of O. sancta. Hong and others [43,44] have shown that the composition of nutritional media impacts both biomass production and the content and yield of a specific metabolic product in this situation. Owing to its increased growth rate, this cyanobacterium mainly depended on its culture medium’s composition, making it a good candidate for future growth studies.
Since, the possible economic biomass production of microalgae is the key issue for microalgae-based applied purposes, it was decided to test the potential of O. sancta to grow under nutrient deficiency using different nutrient concentrations of Zarrouk’s medium after modification by substituting sodium nitrate with a cheaper nitrogen source (urea). According to Schwartz [45], cyanobacteria prefer inorganic nitrogen, particularly nitrates, for growth, but some strains can also thrive on organic nitrogen.
The experimental results (Table 2) may indicate relatively low nutritional demands of O. sancta to grow as dense as in standard Zarrouk’s medium. They may highlight this cyanobacterium’s feasible and economical biomass production in large-scale growth experiments. It is important to note that blue-green algae can develop under different nutrient conditions with flexible metabolic capabilities that enable them to grow under conditions of nutrient scarcity while synthesizing a variety of bioactive molecules [46,47].
Biochemical compositions of biomass grown on 12.5% modified Zarrouk’s medium and standard Zarrouk’s media should be analyzed because the culture medium composition influences not only the growth and metabolic activities of microalgae, but also their biochemical composition [44,48].
The protein analysis results (Figure 4) indicated that the biomass of this cyanobacterium is rich in protein content. This finding may trigger further research to investigate this cyanobacterium protein’s composition and possible bioactivity. Spirulina, a type of cyanobacterium, is a high-protein food alternative to traditional plants like soybean, as it contains up to 70% of its dry-weight biomass as a protein [49].
Figure 4 reveals that the carbohydrate content of O. sancta is endorsed by some scientists [50,51]. They stated that minimal nitrate and phosphorus concentrations significantly increased the carbohydrate content in Synechococcus sp. and Arthrospira platensis biomass compared with the control (Figure 3).
These findings of lipid content (Figure 4) of O. sancta agree with the results stated by Dean and others [52,53]. It is relevant to highlight the almost comparable growth production, as well as protein, carbohydrate, and lipid contents, of O. sancta biomass grown in 12.5% modified Zarrouk’s medium concentration. Standard Zarrouk’s medium represents a concrete guideline to develop this cyanobacterium in 12.5% modified Zarrouk’s medium and to use the produced biomass for further analysis and bioactivity assays.
The significant anticancer activity results of O. sancta methylene chloride/methanol crude extract grown under nutrient deficiency against the MDA-MB-231 cell line (Figure 5) may emphasize the potential role of cyanobacterium for breast cancer therapy. The MDA-MB-231 cell line underwent a thorough literature study and was a very invasive and aggressive triple-negative breast cancer cell line. Some scientists studied the cytotoxic effect of some cyanobacterial isolates Chroococcus minutes and Anabaena oryzae, against MCF-7 cells [54,55]. The MDA-MB-231 is a highly invasive, metastatic, and poorly differentiated human breast cancer, whereas MCF-7 is an invasive and well-differentiated human breast cancer [44]. The anticancer potential of O. sancta extract may be attributed to the GC-MS results (Table 4) that showed the presence of some compounds known previously with their antioxidant and anticancer activity, such as oleic acid, linoleic acid phenol, 2,4-bis (1,1-dimethylethyl)-, 3′,8,8′-trimethoxy-3-piperidyl-2,2′-binaphthalene-1,1′,4,4′-tetrone, hexadecanoic acid methyl ester, and cholestan-3-ol, 2-methylene-, (3β,5α) [56,57,58,59,60].
There is currently a lack of understanding of how cyanobacteria chemicals in tumor cell lines cause cytotoxicity, although multiple investigations have shown that apoptosis is activated [61]. It has been documented in many articles that several cyanobacteria compounds can stimulate an arrest in the cell cycle phase, particularly the G2/M phase, by disrupting microtubules and actin proteins [62,63] involved in mitosis and, thereby, apoptosis [63,64]. Furthermore, cyanobacteria secondary metabolites induce mitochondrial dysfunction, oxidative stress, and DNA damage to cancerous cells [65,66]. Apoptosis can be induced by cyanobacteria’s ability to create various chemicals that boost the activity of caspase proteins, specifically caspase 1 and 3 [62,67,68]. Aside from this, they create chemicals that can start the phosphorylation of Bcl-2 protein and block human breast cancer cells’ anti-apoptotic response [69]. Changes in tumor cell membrane sodium channel kinetics in response to cyanobacteria chemical contact suggest its potential importance in inducing apoptosis [69,70,71,72].
The effectiveness of O. sancta crude extract against Gram + ve bacteria like Staphylococcus aureas and Bacilli cereus and Gram − ve bacteria like Pseudomonas alginolyticus and E. coli has been demonstrated in numerous studies [73,74,75]. The above result reveals that O. sancta, when grown in a nutrient-deficient media, shows the sharpest antibacterial activity against S. aureus and B. cereus [76]. Crude O. sancta extract has been connected to substances such as linoleic acid, hexadecanoic acid methyl ester, 9-decadienoic acid methyl ester, 2-bis(1,1-dimethylethyl), and dasycarpidan-1-methanol acetate, previously linked to antimicrobial action [77,78,79] (Table 4).
As shown in Figure 6, O. sancta extracts greatly reduced AChE activity, suggesting that they may be useful in treating dementias like Alzheimer’s disease and dementia-related diseases like Parkinson’s disease. As acetylcholinesterase inhibitors have been and will continue to be the primary treatment for Alzheimer’s disease symptoms, there is little dispute (AD). Three studies [80,81,82] have shown that a person’s genetic makeup can influence their risk of developing Alzheimer’s disease. Owing to phytochemicals such as dasycarpidan-1-methanol, acetate (ester), cholestan-3-ol, 2-methylene-, (3,5), and 3′,8,8′ trimethoxy-3-piperidyl-2′,2′,4,4′-tetrone that have been demonstrated in GC-MS data, O. sancta extract may have anticholinesterase action (i.e., a possible role for AD treatment) [41,73,83].
5. Conclusions
It can be concluded that modifying Zarrouk’s medium by excluding CaCl2·2H2O, EDTA (Na), NaCl, micronutrients and substituting sodium nitrate with urea might be promising in reducing the medium cost. The marked biomass production of Oscillatoria sancta on 12.5% concentration of modified Zarrouk’s medium indicates the possible economic mass cultivation of such algae under starvation conditions. The potency of O. sancta crude extract against the MDA-MB-231 cell line, pathogenic bacteria, and AChE activity showed the potential of O. sancta secondary metabolites as drug candidates for breast cancer, Alzheimer’s disease, and bacterial diseases. More in-depth in vivo studies may be required to ascertain the bioactivity results of O. sancta extract against these diseases.
Author Contributions
Conceptualization, H.E.T. and D.A.R.; methodology, D.A.R.; formal analysis, H.E.T. and D.A.R.; investigation, H.E.T. and D.A.R.; writing—original draft preparation, H.E.T. and D.A.R.; writing—review and editing, H.E.T. and D.A.R.; funding acquisition, H.E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by the Deanship of Scientific Research (DSR), King Abdulaziz University (Jeddah, Saudi Arabia), by grant No. G: 278-662-1443.
Data Availability Statement
All required data are available in the article.
Acknowledgments
The Deanship of Scientific Research (DSR) at King Abdulaziz University (KAU), Jeddah, Saudi Arabia, has funded this project under grant no (G: 278-662-1443).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Sijil, P.V.; Cherita, C.; Jethani, H.; Chauhan, V.S. Chapter 1. Microalgae as a renewable and sustainable source of high value metabolites. In Microalgae for Sustainable Products; The Royal Society of Chemistry: London, UK, 2022; pp. 1–26. [Google Scholar]
- Burnap, R.L. Systems and photosystems: Cellular limits of autotrophic productivity in cyanobacteria. Front. Bioeng. Biotechnol. 2015, 3, 1. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Hua, Q.; Shimizu, K. Metabolic flux analysis in Synechocystis using isotope distribution from 13c-labeled glucose. Metab. Eng. 2002, 4, 202–216. [Google Scholar] [CrossRef] [PubMed]
- Ambrosi, M.; Reinehr, C.; Bertolin, T.; Costa, J.A.; Colla, L. Propriedades de saúde de Spirulina spp. Rev. Ciências Farm. Básica Apl. 2008, 29, 109–117. [Google Scholar]
- Schwarz, R.; Forchhammer, K. Acclimation of unicellular cyanobacteria to macronutrient deficiency: Emergence of a complex network of cellular responses. Microbiology 2005, 151, 2503–2514. [Google Scholar] [CrossRef]
- Romay, C.; González, R.; Ledón, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef]
- Mogany, T.; Swalaha, F.M.; Kumari, S.; Bux, F. Elucidating the role of nutrients in c-phycocyanin production by the halophilic cyanobacterium euhalothece sp. J. Appl. Phycol. 2018, 30, 2259–2271. [Google Scholar] [CrossRef]
- Tan, L.T. Filamentous tropical marine cyanobacteria: A rich source of natural products for anticancer drug discovery. J. Appl. Phycol. 2010, 22, 659–676. [Google Scholar] [CrossRef]
- Ahmed, W.A.; El-Semary, N.A.; Abd El-Hameed, O.M.; El Tawill, G.; Ibrahim, D.M. Bioactivity and cytotoxic effect of cyanobacterial toxin against hepatocellular carcinoma. J. Cancer Sci. Ther. 2017, 9, 505–511. [Google Scholar]
- Pelizer, L.H.; Danesi, E.D.G.; de O Rangel, C.; Sassano, C.E.; Carvalho, J.C.M.; Sato, S.; Moraes, I.O. Influence of inoculum age and concentration in Spirulina platensis cultivation. J. Food Eng. 2003, 56, 371–375. [Google Scholar] [CrossRef]
- Peter, P.; Sarma, A.P.; Azeem ul Hasan, M.; Murthy, S.D.D.; Attitalla, I.H. Studies on the impact of nitrogen sarma starvation on the photosynthetic pigments through spectral properties of the cyanobacterium, Spirulina platensis: Identification of target phycobiliprotein under nitrogen chlorosis. Bot. Res. Int. 2010, 3, 30–34. [Google Scholar]
- Hai, T.; Hein, S.; Steinbuchel, A. Multiple evidence for widespread and general occurrence of type-iii pha synthases in cyanobacteria and molecular characterization of the pha synthases from two thermophilic cyanobacteria: Chlorogloeopsis fritschii pcc 6912 and synechococcus sp. Strain ma19. Microbiology 2001, 147, 3047–3060. [Google Scholar] [CrossRef]
- El-Kassas, H.Y.; El-Sheekh, M.M. Cytotoxic activity of biosynthesized gold nanoparticles with an extract of the red seaweed corallina officinalis on the mcf-7 human breast cancer cell line. Asian Pac. J. Cancer Prev. 2014, 15, 4311–4317. [Google Scholar] [CrossRef]
- El-Sheekh, M.M.; El-Kassas, H.Y. Application of biosynthesized silver nanoparticles against a cancer promoter cyanobacterium, microcystis aeruginosa. Asian Pac. J. Cancer Prev. 2014, 15, 6773–6779. [Google Scholar] [CrossRef] [Green Version]
- El-Sheekh, M.M.; El-Kassas, H.Y. Algal production of nano-silver and gold: Their antimicrobial and cytotoxic activities: A review. J. Genet. Eng. Biotechnol. 2016, 14, 299–310. [Google Scholar] [CrossRef]
- Gheda, S.; El-Sheekh, M.; Abou-Zeid, A. In vitro anticancer activity of polysaccharide extracted from the red alga Jania rubens against breast and colon cancer cell lines. Asian Pac. J. Trop. Med. 2018, 11, 583–589. [Google Scholar]
- Gademann, K.; Portmann, C. Secondary metabolites from cyanobacteria: Complex structures and powerful bioactivities. Curr. Org. Chem. 2008, 12, 326–341. [Google Scholar] [CrossRef]
- LewisOscar, F.; Nithya, C.; Alharbi, S.A.; Alharbi, N.S.; Thajuddin, N. Microfouling inhibition of human nosocomial pathogen Pseudomonas aeruginosa using marine cyanobacteria. Microb. Pathog. 2018, 114, 107–115. [Google Scholar] [CrossRef]
- Volk, R.-B. Screening of microalgal culture media for the presence of algicidal compounds and isolation and identification of two bioactive metabolites, excreted by the cyanobacteria Nostoc insulare and Nodularia harveyana. J. Appl. Phycol. 2005, 17, 339–347. [Google Scholar] [CrossRef]
- Zelík, P.; Lukešová, A.; Čejka, J.; Buděšínský, M.; Havlíček, V.; Čegan, A.; Kopecký, J. Nostotrebin 6, a bis(cyclopentenedione) with cholinesterase inhibitory activity isolated from nostoc sp. Str. Lukešová 27/97. J. Enzym. Inhib. Med. Chem. 2010, 25, 414–420. [Google Scholar] [CrossRef]
- Becher, P.G.; Baumann, H.I.; Gademann, K.; Jüttner, F. The cyanobacterial alkaloid nostocarboline: An inhibitor of acetylcholinesterase and trypsin. J. Appl. Phycol. 2009, 21, 103–110. [Google Scholar] [CrossRef]
- Dittmann, E.; Gugger, M.; Sivonen, K.; Fewer, D.P. Natural product biosynthetiAc diversity and comparative genomics of the cyanobacteria. Trends Microbiol. 2015, 23, 642–652. [Google Scholar] [CrossRef] [PubMed]
- Mo, S.; Krunic, A.; Chlipala, G.; Orjala, J. Antimicrobial ambiguine isonitriles from the cyanobacterium Fischerella ambigua. J. Nat. Prod. 2009, 72, 894–899. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.K.; Tiwari, S.P.; Rai, A.K.; Mohapatra, T.M. Cyanobacteria: An emerging source for drug discovery. J. Antibiot. 2011, 64, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Karan, T.; Aydin, A. Anticancer potential and cytotoxic effect of some freshwater cyanobacteria. Trop. J. Pharm. Res. 2018, 17, 2183–2188. [Google Scholar] [CrossRef]
- Sithranga Boopathy, N.; Kathiresan, K. Anticancer drugs from marine flora: An overview. J. Oncol. 2010, 2010, 214186. [Google Scholar] [CrossRef]
- Robles-Banuelos, B.; Duran-Riveroll, L.M.; Rangel-Lopez, E.; Perez-Lopez, H.I.; Gonzalez-Maya, L. Marine cyanobacteria as sources of lead anticancer compounds: A review of families of metabolites with cytotoxic, antiproliferative, and antineoplastic effects. Molecules 2022, 27, 4814. [Google Scholar] [CrossRef]
- Tiwari, A.K.; Tiwari, B.S. Cyanotherapeutics: An emerging field for future drug discovery. Appl. Phycol. 2020, 1, 44–57. [Google Scholar] [CrossRef]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [CrossRef]
- Smoker, J.A.; Susan, R.B. Rapid small-scale DNA isolation from filamentous cyanobacteria. FEMS Microbiol. Lett. 1988, 56, 119–122. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Zarrouk, C. Contribution a l’etude d’une Cyanophycee. Influence de Divers Facteurs Physiques et Chimiques sur la Croissance et la Photosynthese de Spirulina maxima. Ph.D. Thesis, Université De Paris, Paris, France, 1966. [Google Scholar]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association, American Water Works Association and Water Environment Federation: Washington, DC, USA, 2008. [Google Scholar]
- Andersen, R.A. Algal Culturing Techniques; Elsevier Academic Press: Birlington, MA, USA; San Diego, CA, USA; London, UK, 2005. [Google Scholar]
- Stein, J.R.; Hellebust, J.A.; Craigie, J. Handbook of Phycological Methods: Culture Methods and Growth Measurements; Cambridge University Press: New York, NY, USA, 1973; 448p. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Stoscheck, C.M. [6] Quantitation of protein. In Methods in Enzymology; Deutscher, M.P., Ed.; Academic Press: Cambridge, MA, USA, 1990; Volume 182, pp. 50–68. [Google Scholar]
- Hedge, J.E.; Hofreiter, B.T. Carbohydrate Chemistry; Whistler, R.L., Be Miller, J.N., Eds.; Academic Press: New York, NY, USA, 1962; Volume 17. [Google Scholar]
- Sadasivam, S.; Manickam, A. Biochemical Methods; New Age International (P) Limited: New Delhi, India, 1996; Volume 2, pp. 124–126. [Google Scholar]
- Perez, C. Antibiotic assay by agar-well diffusion method. Acta Biol. Med. Exp. 1990, 15, 113–115. [Google Scholar]
- Refaay, D.A.; Abdel-Hamid, M.I.; Alyamani, A.A.; Abdel Mougib, M.; Ahmed, D.M.; Negm, A.; Mowafy, A.M.; Ibrahim, A.A.; Mahmoud, R.M. Growth Optimization and Secondary Metabolites Evaluation of Anabaena variabilis for Acetylcholinesterase Inhibition Activity. Plants 2022, 11, 735. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Hong, S.-J.; Lee, C.G. Statistical optimization of culture media for production of phycobiliprotein by Synechocystis sp. Pcc 6701. Biotechnol. Bioprocess Eng. 2008, 13, 491–498. [Google Scholar] [CrossRef]
- Grobbelaar, J. Factors governing algal growth in photobioreactors: The “open” versus “closed” debate. J. Appl. Phycol. 2009, 21, 489–492. [Google Scholar] [CrossRef]
- Schwartz, W.; Fogg, G.E.; Stewart, W.D.P.; Fay, P.; Walsby, A.E. The blue-green algae. Vii, 459 s., 170 abb. Und 8 farbige abb., 33 tab., 1 farbtafel. London-new york 1973: Academic press, £ 8.50. Z. Allg. Mikrobiol. 1975, 15, 68. [Google Scholar] [CrossRef]
- Hu, Q.; Sommerfeld, M.; Jarvis, E.; Ghirardi, M.; Posewitz, M.; Seibert, M.; Darzins, A. Microalgal triacylglycerols as feedstocks for biofuel production: Perspectives and advances. Plant J. Cell Mol. Biol. 2008, 54, 621–639. [Google Scholar] [CrossRef]
- Ferrell, J.; Sarisky-Reed, V. National Algal Biofuels Technology Roadmap; EERE Publication and Product Library: Washington, DC, USA, 2010. [Google Scholar]
- Borowitzka, M. Culturing Microalgae in Outdoor Ponds; Academic Press: Boston, MA, USA, 2005; pp. 205–221. [Google Scholar]
- Mehla, S.; Kukade, S.; Kumar, P.; Rao, P.; Sriganesh, G.; Ravishankar, R. Fine tuning h-transfer and β-scission reactions in vgo fcc using metal promoted dual functional zsm-5. Fuel 2019, 242, 487–495. [Google Scholar] [CrossRef]
- Möllers, K.B.; Cannella, D.; Jørgensen, H.; Frigaard, N.-U. Cyanobacterial biomass as carbohydrate and nutrient feedstock for bioethanol production by yeast fermentation. Biotechnol. Biofuels 2014, 7, 64. [Google Scholar] [CrossRef]
- Babushkina, E.; Belokopytova, L.; Grachev, A.; Meko, D.; Vaganov, E.A. Variation of the hydrological regime of bele-shira closed basin in southern siberia and its reflection in the radial growth of Larix sibirica. Reg. Environ. Chang. 2017, 17, 1725–1737. [Google Scholar] [CrossRef]
- Dean, A.P.; Nicholson, J.M.; Sigee, D.C. Impact of phosphorus quota and growth phase on carbon allocation in Chlamydomonas reinhardtii: An ftir microspectroscopy study. Eur. J. Phycol. 2008, 43, 345–354. [Google Scholar] [CrossRef]
- Cade-Menun, B.J.; Paytan, A. Nutrient temperature and light stress alter phosphorus and carbon forms in culture-grown algae. Mar. Chem. 2010, 121, 27–36. [Google Scholar] [CrossRef]
- Felczykowska, A.; Pawlik, A.; Mazur-Marzec, H.; Toruńska-Sitarz, A.; Narajczyk, M.; Richert, M.; Węgrzyn, G.; Herman-Antosiewicz, A. Selective inhibition of cancer cells’ proliferation by compounds included in extracts from Baltic sea cyanobacteria. Toxicon 2015, 108, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Han, B.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Aurilides B and C, cancer cell toxins from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2006, 69, 572–575. [Google Scholar] [CrossRef]
- Yu, F.R.; Lian, X.Z.; Guo, H.Y.; McGuire, P.M.; Li, R.D.; Wang, R.; Yu, F.H. Isolation and characterization of methyl esters and derivatives from Euphorbia kansui (euphorbiaceae) and their inhibitory effects on the human sgc-7901 cells. J. Pharm. Pharm. Sci. 2005, 8, 528–535. [Google Scholar]
- Altameme, H.; Hameed, I.; Idan, S.; Hadi, M. Biochemical analysis of Origanum vulgare seeds by fourier-transform infrared (ft-ir) spectroscopy and gas chromatography-mass spectrometry (gc-ms). J. Pharmacogn. Phytother. Res. 2015, 7, 221–237. [Google Scholar]
- Ibraheam, I.A.; Hadi, M.Y.; Hameed, I.H. Analysis of bioactive compounds of methanolic leaves extract of mentha pulegium using gas chromatography-mass spectrometry (gc-ms) technique. Int. J. Pharm. Qual. Assur. 2017, 8, 174–182. [Google Scholar] [CrossRef]
- Nair, R.V.R.; Jayasree, D.V.; Biju, P.G.; Baby, S. Anti-inflammatory and anticancer activities of erythrodiol-3-acetate and 2,4-di-tert-butylphenol isolated from humboldtia unijuga. Nat. Prod. Res. 2020, 34, 2319–2322. [Google Scholar] [CrossRef]
- Zhao, F.; Wang, P.; Lucardi, R.; Su, Z.; Li, S. Natural sources and bioactivities of 2,4-di-tert-butylphenol and its analogs. Toxins 2020, 12, 35. [Google Scholar] [CrossRef]
- Costa, M.; Costa-Rodrigues, J.; Fernandes, M.H.; Barros, P.; Vasconcelos, V.; Martins, R. Marine cyanobacteria compounds with anticancer properties: A review on the implication of apoptosis. Mar. Drugs 2012, 10, 2181–2207. [Google Scholar] [CrossRef]
- Oftedal, L.; Myhren, L.; Jokela, J. The lipopeptide toxins anabaenolysin a and b target biological membranes in a cholesterol-dependent manner. Biochim. Biophys. Acta Biomembr. 2012, 1818, 3000–3009. [Google Scholar] [CrossRef]
- Catassi, A.; Cesario, A.; Arzani, D.; Menichini, P.; Alama, A.; Bruzzo, C.; Imperatori, A.; Rotolo, N.; Granone, P.; Russo, P. Characterization of apoptosis induced by marine natural products in non small cell lung cancer a549 cells. Cell. Mol. Life Sci. 2006, 63, 2377–2386. [Google Scholar] [CrossRef]
- Sato, M.; Sagawa, M.; Nakazato, T.; Ikeda, Y.; Kizaki, M. A natural peptide, dolastatin 15, induces g2/m cell cycle arrest and apoptosis of human multiple myeloma cells. Int. J. Oncol. 2007, 30, 1453–1459. [Google Scholar] [CrossRef]
- Sato, S.; Murata, A.; Orihara, T.; Shirakawa, T.; Suenaga, K.; Kigoshi, H.; Uesugi, M. Marine natural product aurilide activates the opa1-mediated apoptosis by binding to prohibitin. Chem. Biol. 2011, 18, 131–139. [Google Scholar] [CrossRef]
- Paradies, G.; Petrosillo, G.; Paradies, V.; Ruggiero, F. Mitochondrial dysfunction in brain aging: Role of oxidative stress and cardiolipin. Neurochem. Int. 2011, 58, 447–457. [Google Scholar] [CrossRef]
- Oftedal, L.; Selheim, F.; Wahlsten, M.; Sivonen, K.; Døskeland, S.; Herfindal, L. Marine benthic cyanobacteria contain apoptosis-inducing activity synergizing with daunorubicin to kill leukemia cells, but not cardiomyocytes. Mar. Drugs 2010, 8, 2659–2672. [Google Scholar] [CrossRef]
- Yonezawa, T.; Mase, N.; Sasaki, H.; Teruya, T.; Hasegawa, S.-I.; Cha, B.-Y.; Yagasaki, K.; Suenaga, K.; Nagai, K.; Woo, J.-T. Biselyngbyaside, isolated from marine cyanobacteria, inhibits osteoclastogenesis and induces apoptosis in mature osteoclasts. J. Cell. Biochem. 2012, 113, 440–448. [Google Scholar] [CrossRef]
- Pereira, A.; Cao, Z.; Murray, T.F.; Gerwick, W.H. Hoiamide a, a sodium channel activator of unusual architecture from a consortium of two papua new guinea cyanobacteria. Chem. Biol. 2009, 16, 893–906. [Google Scholar] [CrossRef]
- Choi, H.; Pereira, A.; Cao, Z.; Shuman, C.; Engene, N.; Byrum, T.; Matainaho, L.; Murray, T.; Mangoni, A.; Gerwick, W. The hoiamides, structurally intriguing neurotoxic lipopeptides from papua new guinea marine cyanobacteria. J. Nat. Prod. 2010, 73, 1411–1421. [Google Scholar] [CrossRef]
- Acuña, U.M.; Mo, S.; Zi, J.; Orjala, J.; de Blanco, E.J.C. Hapalindole h induces apoptosis as an inhibitor of nf-kb and affects the intrinsic mitochondrial pathway in pc-3 androgen-insensitive prostate cancer cells. Anticancer Res. 2018, 38, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Chaudhry, G.E.; Md Akim, A.; Sung, Y.Y.; Sifzizul, T.M.T. Cancer and apoptosis: The apoptotic activity of plant and marine natural products and their potential as targeted cancer therapeutics. Front. Pharmacol. 2022, 13, 842376. [Google Scholar] [CrossRef]
- Ramamurthy, V.; Raveendran, S.; Thirumeni, S.; Krishnaveni, S. Antimicrobial activity of heterocytic cyanobacteria. Int. J. Adv. Life Sci. 2012, 1, 32–39. [Google Scholar]
- Khairy, H.M.; El-Kassas, H.Y. Active substance from some blue green algal species used as antimicrobial agents. Afr. J. Biotechnol. 2010, 9, 2789–2800. [Google Scholar]
- El-Sheekh, M.; Osman, M.; Dyab, M.; Amer, M. Production and characterization of antimicrobial active substance from the cyanobacterium Nostoc muscorum. Environ. Toxicol. Pharmacol. 2006, 21, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Mian, P.; Heilmann, J.; Bürgi, H.R.; Sticher, O. Biological screening of terrestrial and freshwater cyanobacteria for antimicrobial activity, brine shrimp lethality, and cytotoxicity. Pharm. Biol. 2008, 41, 243–247. [Google Scholar] [CrossRef]
- Huang, C.B.; George, B.; Ebersole, J.L. Antimicrobial activity of n-6, n-7 and n-9 fatty acids and their esters for oral microorganisms. Arch. Oral Biol. 2010, 55, 555–560. [Google Scholar] [CrossRef]
- Amaral, A.C.; Gomes, L.A.; Silva, J.R.; Ferreira, J.L.; Ramos Ade, S.; Rosa Mdo, S.; Vermelho, A.B.; Rodrigues, I.A. Liposomal formulation of turmerone-rich hexane fractions from Curcuma longa enhances their antileishmanial activity. BioMed Res. Int. 2014, 2014, 694934. [Google Scholar] [CrossRef]
- Sundar, S.; Justin-Koil Pillai, Y. Phytochemical screening and gas chromatograph-mass spectrometer profiling in the leaves of Solanum incanum. Asian J. Pharm. Clin. Res. 2015, 8, 179–188. [Google Scholar]
- Mehta, M.; Adem, A.; Sabbagh, M. New acetylcholinesterase inhibitors for alzheimer’s disease. Int. J. Alzheimer’s Dis. 2012, 2012, 728983. [Google Scholar] [CrossRef] [Green Version]
- Konrath, E.L.; Neves, B.M.; Lunardi, P.S.; Passos, C.d.S.; Simões-Pires, A.; Ortega, M.G.; Gonçalves, C.A.; Cabrera, J.L.; Moreira, J.C.F.; Henriques, A.T. Investigation of the in vitro and ex vivo acetylcholinesterase and antioxidant activities of traditionally used lycopodium species from south america on alkaloid extracts. J. Ethnopharmacol. 2012, 139, 58–67. [Google Scholar] [CrossRef]
- Ferreira, A.; Proença, C.; Serralheiro, M.L.M.; Araújo, M.E.M. The in vitro screening for acetylcholinesterase inhibition and antioxidant activity of medicinal plants from portugal. J. Ethnopharmacol. 2006, 108, 31–37. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Kumar, V.; Mal, M.; Houghton, P.J. Acetylcholinesterase inhibitors from plants. Phytomedicine 2007, 14, 289–300. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree dendrogram based on O. sancta 16S rRNA gene sequence using the bootstrap test of the maximum-likelihood (ML) technique.
Figure 1.
Phylogenetic tree dendrogram based on O. sancta 16S rRNA gene sequence using the bootstrap test of the maximum-likelihood (ML) technique.

Figure 2.
Mean changes in chlorophyll concentration (mg L−1) of O. sancta grown on BG11 and Zarrouk’s media.
Figure 2.
Mean changes in chlorophyll concentration (mg L−1) of O. sancta grown on BG11 and Zarrouk’s media.

Figure 3.
Specific growth rate (µ) and divisions per day (Dd−1) of O. sancta grown on BG 11 and Zarrouk’s media. Values are the averages of three replicates ± standard deviation.
Figure 3.
Specific growth rate (µ) and divisions per day (Dd−1) of O. sancta grown on BG 11 and Zarrouk’s media. Values are the averages of three replicates ± standard deviation.

Figure 4.
Biochemical composition of O. sancta grown on Zarrouk’s medium (control) and 12.5% modified Zarrouk’s medium concentration (T3). Error bars show the SD for three measurements.
Figure 4.
Biochemical composition of O. sancta grown on Zarrouk’s medium (control) and 12.5% modified Zarrouk’s medium concentration (T3). Error bars show the SD for three measurements.

Figure 5.
IC50 of O. sancta crude extracts grown on standard Zarrouk’s medium (control) and modified Zarrouk’s medium concentration (T3).
Figure 5.
IC50 of O. sancta crude extracts grown on standard Zarrouk’s medium (control) and modified Zarrouk’s medium concentration (T3).

Figure 6.
The AChE inhibitory activity of O. sancta crude extracts grown on Zarrouk’s medium (control) and 12.5% modified Zarrouk’s medium concentration (T3).
Figure 6.
The AChE inhibitory activity of O. sancta crude extracts grown on Zarrouk’s medium (control) and 12.5% modified Zarrouk’s medium concentration (T3).

Figure 7.
GC chromatogram of O. sancta crude extracts grown on T3.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
The modified Zarrouk’s medium chemical composition (g L−1).
| Constituents | Zarrouk’s Medium (g L−1) * | Modified Zarrouk’s Medium (g L−1) | ||
|---|---|---|---|---|
| 100% (T1) | 50% (T2) | 12.5% (T3) | ||
| NaNO3 | 2.5 | - | - | - |
| Urea | - | 0.8 | 0.4 | 0.1 |
| CaCl2·2H2O | 0.04 | - | - | - |
| NaCl | 1.0 | - | - | - |
| FeSO4·7H2O | 0.01 | 0.01 | 0.005 | 0.00125 |
| EDTA (Na) | 0.08 | - | - | - |
| K2SO4 | 1.0 | 1.0 | 0.5 | 0.125 |
| MgSO4·7H2O | 0.2 | 0.2 | 0.1 | 0.025 |
| NaHCO3 | 16.8 | 16.8 | 8.4 | 2.1 |
| K2HPO4 | 0.5 | 0.5 | 0.25 | 0.0625 |
| Micronutrients (H3BO3, MnCl2·4H2O, ZnSO4·4H2O Na2MoO4, CuSO4·5H2O) | 1.0 mL | - | - | - |
Note(s): * standard Zarrouk’s medium (control).
Table 2.
Dry weight biomass (g L−1) of Oscillatoria sancta grown on different modified Zarrouk’s medium concentrations (100%, 50%, 12.5%).
Table 2.
Dry weight biomass (g L−1) of Oscillatoria sancta grown on different modified Zarrouk’s medium concentrations (100%, 50%, 12.5%).
| Treatments | Dry wt. (g L−1) |
|---|---|
| Control (standard Zarrouk’s medium) | 0.552 a ± 0.027 * |
| T1 | 0.539 ab ± 0.025 |
| T2 | 0.53 ab ± 0.022 |
| T3 | 0.509 ab ± 0.022 |
Note(s): * values shown are the averages of three replicates ± standard deviation. Different letters indicate significant differences at (p > 0.05).
Table 3.
The inhibition zone diameter (mm) measurement of different O. sancta crude extracts against pathogenic bacteria.
Table 3.
The inhibition zone diameter (mm) measurement of different O. sancta crude extracts against pathogenic bacteria.
| Bacteria | Diameter of Inhibition Zone (mm) | ||
|---|---|---|---|
| Zarrouk’s Medium (Control) | T3 a | Solvent b | |
| Pseudomonas aeruginosa | N | N | N c |
| Escherichia coli | N | N | N |
| Bacillus cereus | 16.3 ± 0.5 | 17.2 ± 0.44 | N |
| Staphylococcus aureus | 12.3 ± 0.7 | 15.3 ± 0.58 | N |
Note(s): a 12.5% modified Zarrouk’s medium concentrations, b methylene chloride/methanol in a ratio of 1:1 (v/v), c no inhibition.
Table 4.
The data of GC-MS analysis of methylene chloride/methanol crude extract of O. sancta grown on 12.5% modified Zarrouk’s medium concentration.
Table 4.
The data of GC-MS analysis of methylene chloride/methanol crude extract of O. sancta grown on 12.5% modified Zarrouk’s medium concentration.
| Peak No. | Compound | Chemical Group | Retention Time (min.) | Formula | Area% |
|---|---|---|---|---|---|
| 1 | Oleic acid, eicosyl ester | Oleic acid ester | 11.383 | C38H74O2 | 5.5 |
| 2 | Phenol, 2,4-bis(1,1-dimethylethyl)- | Phenol | 13.652 | C14H22O | 25 |
| 3 | Pentatriacont-17-ene | Alkene | 14.808 | C35H70 | 6.2 |
| 4 | 9-Hexadecenoic acid, 9-octadecenyl ester, (Z,Z)- | Ester | 18.134 | C34H64O2 | 3.8 |
| 5 | 7-Hexadecenoic acid, methyl ester, (Z)- | Methyl ester | 20.175 | C17H32O2 | 5.7 |
| 6 | Hexadecanoic acid, methyl ester | Methyl ester | 20.532 | C17H34O2 | 16 |
| 7 | Dasycarpidan-1-methanol, acetate (ester) | Indole alkaloid | 23.477 | C20H26N2O2 | 13.1 |
| 8 | 9,12-Octadecadienoyl chloride, (Z, Z)- | Linoleic acid chloride | 23.668 | C18H31ClO | 10.5 |
| 9 | Cholestan-3-ol, 2-methylene-, (3β,5α)- | Steroid | 23.975 | C28H48O | 7.1 |
| 10 | 3′,8,8′-Trimethoxy-3-piperidyl-2,2′-binaphthalene-1,1′,4,4′-tetrone | Terpene | 31.095 | C28H25NO7 | 7.15 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Touliabah, H.E.; Refaay, D.A. Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions. Water 2023, 15, 664. https://doi.org/10.3390/w15040664
AMA Style
Touliabah HE, Refaay DA. Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions. Water. 2023; 15(4):664. https://doi.org/10.3390/w15040664
Chicago/Turabian StyleTouliabah, Hussein E., and Dina A. Refaay. 2023. "Enhancement of Anticancer, Antibacterial, and Acetylcholinesterase Inhibition Activities from Oscillatoria sancta under Starvation Conditions" Water 15, no. 4: 664. https://doi.org/10.3390/w15040664
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.