
Silicon Nutrition in Plants under Water-Deficit Conditions: Overview and Prospects
 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
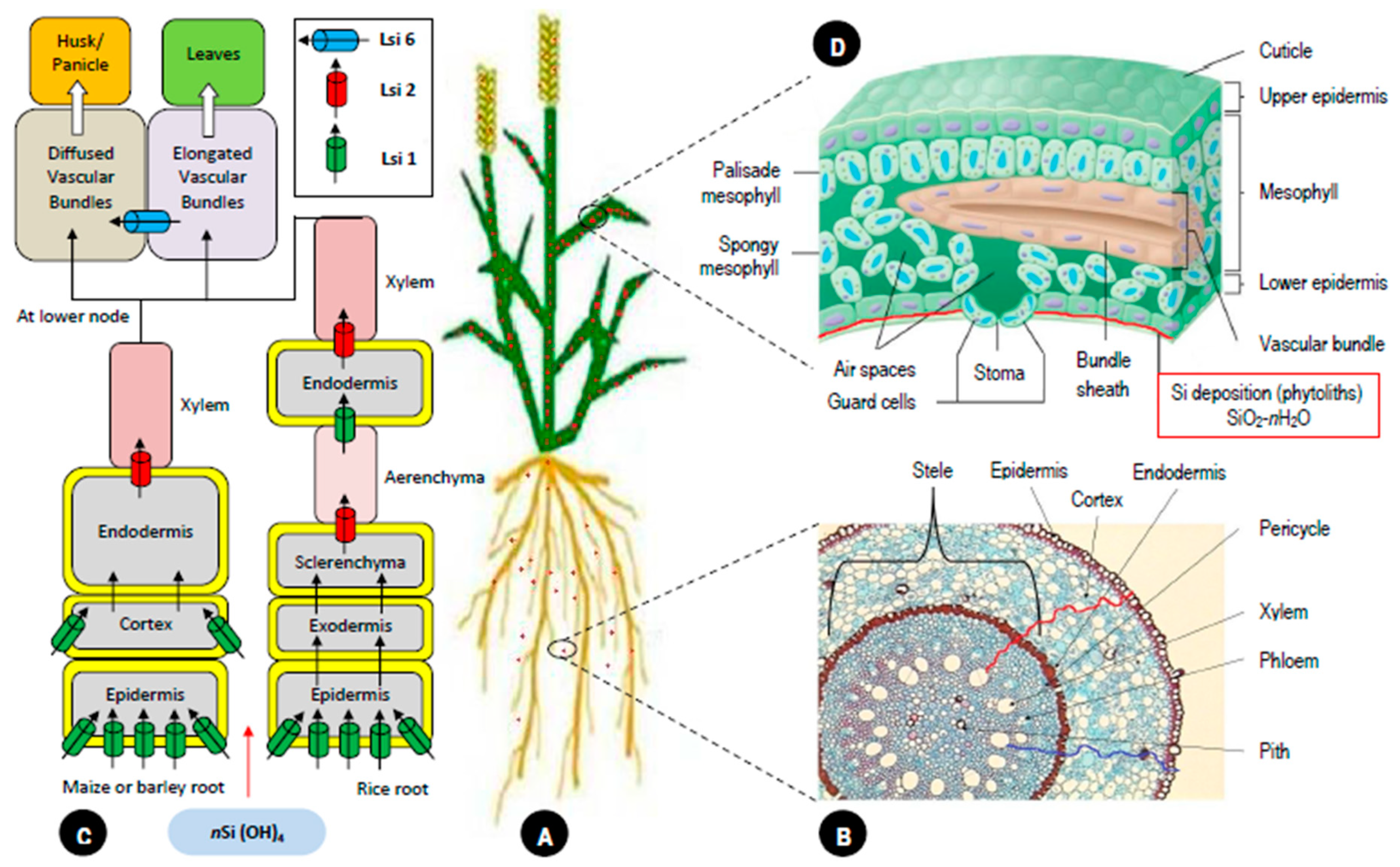
2. Silicon: Uptake, Translocation and Deposition in Plants
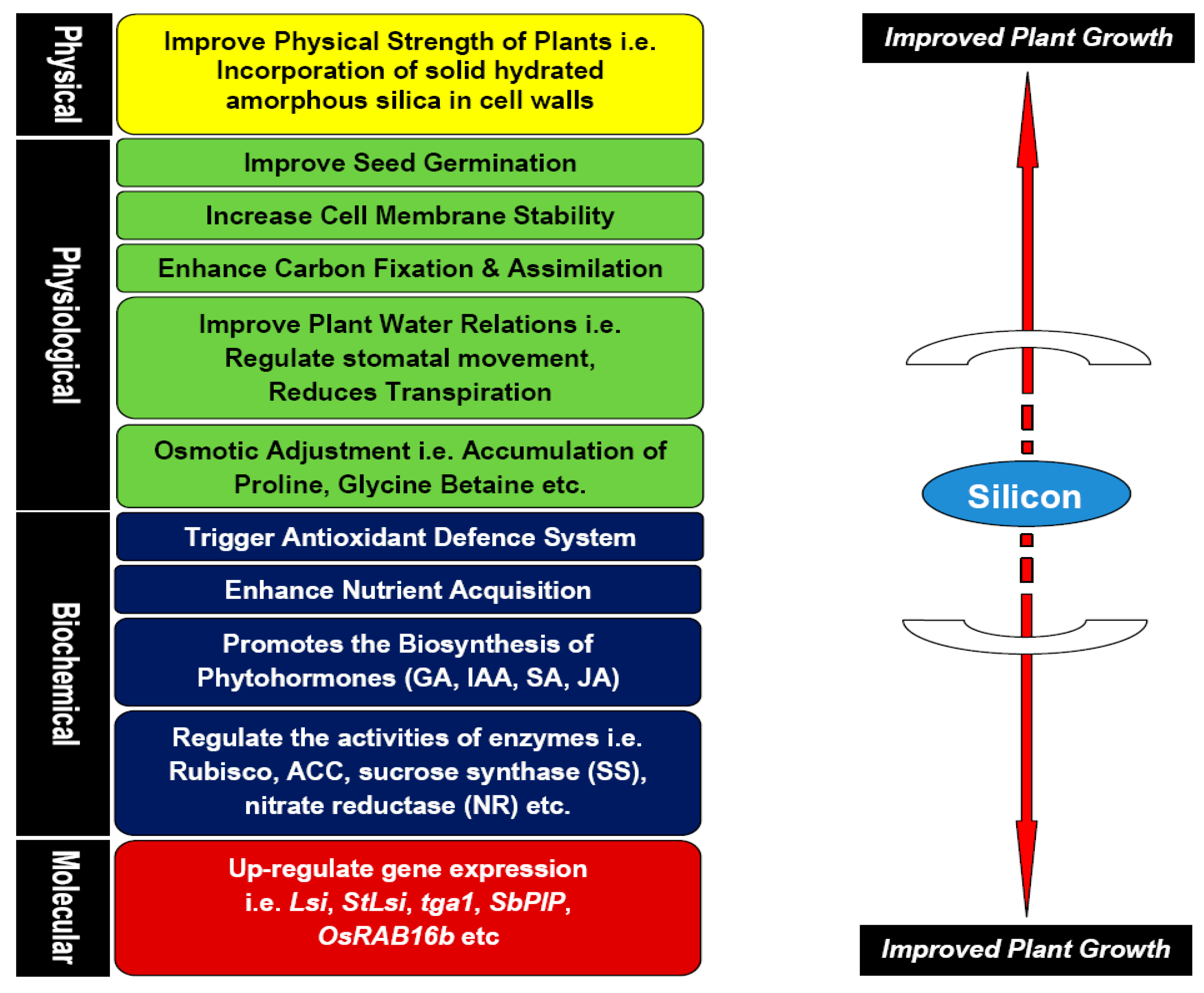
3. Role of Silicon in Plant Biology
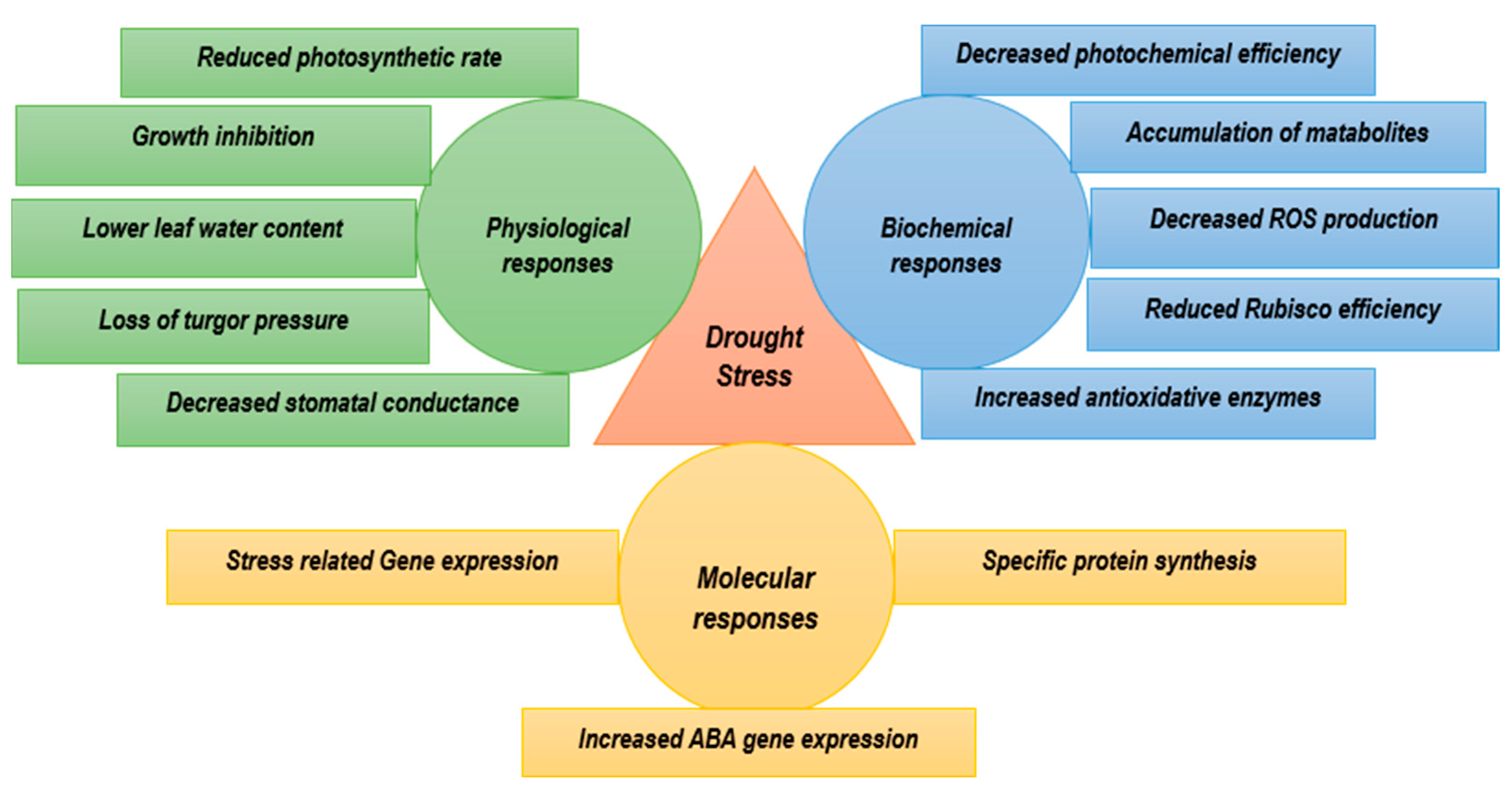
4. Silicon Nutrition and Plant Drought Tolerance
4.1. Seed Germination
4.2. Cell Membrane Stability (CMS)
4.3. Carbon Fixation and Assimilation
4.4. Plant–Water Relations
4.5. Osmotic Adjustment (OA)
4.6. Phytohormone Metabolism
4.7. Antioxidant Defense Mechanism
4.8. Acquisition of Mineral Nutrients
5. Molecular Basis of Si-Induced Drought Tolerance
6. Conclusions and Perspectives
- The potential of Si in mitigating drought stress is poorly understood at the molecular level; the molecular physiology of the Si-induced regulation of osmotic adjustment, water and nutrient uptake, the biosynthesis of stress proteins and phytohormones, including K-regulated aquaporin expression, need investigation.
- Some members of the poaceae family, including maize and wheat, also accumulate Si but in lesser amounts than rice. Understanding mechanisms for Si uptake in these plants needs further identification of transporters. Likely, transporters involved in Si efflux in non-accumulating species are required.
- Advancements in molecular biology and biotechnology such as CRISPAR/Cas and DNA recombinant technologies have enabled the transfer of desired genes or the restriction/knockout of undesired genes to be achieved, which is not possible with traditional plant breeding techniques. Therefore, gene transfer from rice to other crop plants, especially dicots, should be exploited to enhance plant resistance against drought stress by genetically manipulating the Si uptake ability of other plants.
- Optimizing the rates and timing for Si foliar and soil application may increase its agronomic application, especially in water-soluble fertilizers.
- The sole application of Si as a fertilizer may increase input costs in terms of fertilizer price and labor costs for application and may affect the direct income of the farm. However, the enrichment of essential fertilizers such as urea and phosphate fertilizers with Si in areas facing acute risk of abiotic stresses, particularly water deficit, may provide a sustainable solution for Si application to field crops.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Ma, D.; Sun, D.; Wang, C.; Qin, H.; Ding, H.; Li, Y.; Guo, T. Silicon application alleviates drought stress in wheat through transcriptional regulation of multiple antioxidant defense pathways. J. Plant Growth Regul. 2016, 35, 1–10. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Pessarakli, M. Effect of silicon on photosynthetic gas exchange, photosynthetic pigments, cell membrane stability and relative water content of different wheat cultivars under drought stress conditions. J. Plant Nutr. 2016, 39, 1001–1015. [Google Scholar] [CrossRef]
- Romm, J. The next dust bowl. Nature 2011, 478, 450–451. [Google Scholar] [CrossRef]
- The World Bank. World Development Indicators. Available online: https://datacatalog.worldbank.org/dataset/world-development-indicators (accessed on 15 April 2014).
- Noman, A.; Ali, S.; Naheed, F.; Ali, Q.; Farid, M.; Rizwan, M.; Irshad, M.K. Foliar application of ascorbate enhances the physiological and biochemical attributes of maize (Zea mays L.) cultivars under drought stress. Arch. Agron. Soil Sci. 2015, 61, 1659–1672. [Google Scholar] [CrossRef]
- Farooq, M.A.; Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant Drought Stress: Effects, Mechanisms and Management. In Sustainable Agriculture; Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C., Eds.; Springer Nature: Cham, Switzerland, 2009; pp. 153–188. [Google Scholar]
- Kim, W.; Iizumi, T.; Nishimori, M. Global patterns of crop production losses associated with droughts from 1983 to 2009. J. Appl. Meteorol. Climatol. 2019, 58, 1233–1244. [Google Scholar] [CrossRef]
- Xie, Z.; Song, R.; Shao, H.; Song, F.; Xu, H.; Lu, Y. Silicon improves maize photosynthesis in saline-alkaline soils. Sci. World J. Article 2015, 2015, 245072. [Google Scholar] [CrossRef]
- Sahebi, M.; Hanafi, M.M.; Siti Nor Akmar, A.; Rafii, M.Y.; Azizi, P.; Tengoua, F.F.; Nurul Mayzaitul Azwa, J.; Shabanimofrad, M. Importance of silicon and mechanisms of biosilica formation in plants. BioMed Res. Int. 2015, 2015, 396010. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Huynh, W.Q.; Kronzucker, H.J. The role of silicon in higher plants under salinity and drought stress. Front. Plant Sci. 2016, 18, 1072. [Google Scholar] [CrossRef] [PubMed]
- Garg, N.; Bhandari, P.; Kashyap, L.; Singh, S. Silicon Nutrition and Arbuscular Mycorrhizal Fungi. Metalloids in Plants: Advances and Future Prospects; Deshmukh, D.K., Ed.; Jhon Wiley & Sons, Inc.: Hoboken, NJ, USA, 2020; pp. 315–354. [Google Scholar]
- Ma, J.F.; Miyake, Y.; Takahashi, E. Silicon as a beneficial element for crop plants. In Silicon in Agriculture; Datnoff, L.E., Snyder, G.H., Korndörfer, G.H., Eds.; Elsevier Science: New York, NY, USA, 2001; pp. 17–39. [Google Scholar]
- Hodson, M.J.; White, P.J.; Mead, A.; Broadley, M.R. Phylogenetic variation in the silicon composition of plants. Ann. Bot. 2005, 96, 1027–1046. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Annu. Rev. Plant Biol. 1999, 50, 641. [Google Scholar] [CrossRef]
- Richmond, K.E.; Sussman, M. Got silicon? The non-essential beneficial plant nutrient. Curr. Opin. Plant Biol. 2003, 6, 268–272. [Google Scholar] [CrossRef]
- Guntzer, F.; Keller, C.; Poulton, P.R.; McGrath, S.P.; Meunier, J.D. Long-term removal of wheat straw decreases soil amorphous silica at Broadbalk, Rothamsted. Plant Soil 2012, 352, 173–184. [Google Scholar] [CrossRef]
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon pools and fluxes in soils and landscapes—A review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef]
- Chanchal Malhotra, C.; Kapoor, R.; Ganjewala, D. Alleviation of abiotic and biotic stresses in plants by silicon supplementation. Scientia 2016, 13, 59–73. [Google Scholar]
- Ma, J.F.; Yamaji, N. Silicon uptake and accumulation in higher plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Rains, D.W.; Epstein, E.; Zasoski, R.J.; Aslam, M. Active silicon uptake by wheat. Plant Soil 2006, 280, 223–228. [Google Scholar] [CrossRef]
- Liang, Y.; Si, J.; Römheld, V. Silicon uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef]
- Takahashi, E.; Ma, J.F.; Miyake, Y. The possibility of silicon as an essential element for higher plants. Comm. Agric. Food Chem. 1990, 2, 99–102. [Google Scholar]
- Mitani, N.; Chiba, Y.; Amaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef]
- Feng, J.; Yamaji, N.; Mitani-Ueno, N. Transport of silicon from roots to panicles in plants. Proc. Jpn. Acad. Ser. 2011, 87, 377–385. [Google Scholar]
- Ma, J.F.; Yamaji, N. Functions and transport of silicon in plants. Cell Mol. Life Sci. 2008, 65, 3049–3057. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Yamaji, N.; Mitatni, N.; Ma, J.F. A transporter regulating silicon distribution in rice shoots. Plant cell. 2008, 20, 1381–1389. [Google Scholar] [CrossRef]
- Ma, J.F.; Takahashi, E. Soil, Fertilizer, and Plant Silicon Research in Japan; Elsevier: Amsterdam, The Netherlands, 2002. [Google Scholar]
- Rao, G.B.; Susmitha, P. Silicon uptake, transportation and accumulation in Rice. J. Pharmacogn. Phytochem. 2017, 6, 290–293. [Google Scholar]
- Keller, C.; Rizwan, M.; Davidian, J.C.; Pokrovsky, O.S.; Bovet, N.; Chaurand, P.; Meunier, J.D. Effect of silicon on wheat seedlings (Triticum turgidum L.) grown in hydroponics and exposed to 0 to 30 µM Cu. Planta 2015, 24, 847–860. [Google Scholar] [CrossRef] [PubMed]
- Haynes, R.J. Significance and role of Si in crop production. Adv. Agron. 2017, 146, 83–166. [Google Scholar]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and future prospects on the action mechanisms in alleviating biotic and abiotic stresses in plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Vulavala, V.K.; Elbaum, R.; Yermiyahu, U.; Fogelman, E.; Kumar, A.; Ginzberg, I. Silicon fertilization of potato: Expression of putative transporters and tuber skin quality. Planta 2016, 243, 217–229. [Google Scholar] [CrossRef] [PubMed]
- Van Bockhaven, J.; Steppe, K.; Bauweraerts, I.; Kikuchi, S.; Asano, T.; Höfte, M.; De Vleesschauwer, D. Primary metabolism plays a central role in moulding silicon—Inducible brown spot resistance in rice. Mol. Plant Pathol. 2015, 16, 811–824. [Google Scholar] [CrossRef] [PubMed]
- Markovich, O.; Steiner, E.; Kouřil, Š.; Tarkowski, P.; Aharoni, A.; Elbaum, R. Silicon promotes cytokinin biosynthesis and delays senescence in Arabidopsis and Sorghum. Plant Cell Environ. 2017, 40, 1189–1196. [Google Scholar] [CrossRef] [PubMed]
- Currie, H.A.; Perry, C.C. Silica in plants: Biological, biochemical and chemical studies. Ann. Bot. 2007, 100, 1383–1389. [Google Scholar] [CrossRef]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxová, M.; Lux, A. Application of silicon enhanced drought tolerance in Sorghum bicolor. Physiol. Plant. 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Fleck, A.T.; Nye, T.; Repenning, C.; Stahl, F.; Zahn, M.; Schenk, M.K. Silicon enhances suberization and lignification in roots of rice (Oryza sativa). J. Exp. Bot. 2011, 62, 2001–2011. [Google Scholar] [CrossRef]
- Apel, K.; Hirt, H. Reactive oxygen species: Metabolism, oxidative stress, and signaling transduction. Annu. Rev. Plant Biol. 2004, 55, 373. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, Y.; Yao, H.; Wu, J.; Sun, H.; Gong, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Biochem. 2014, 78, 27–36. [Google Scholar] [CrossRef]
- Liu, H.; Shen, X.; Guo, Z. Effects of silicon addition on seed germination and seedling growth of alfalfa. Acta Pratacul. Sinica. 2011, 20, 155–160. [Google Scholar]
- Firoozeh, T.; Ahmad, M.; Shekoofeh, E. Effect of exogenous silicon on germination and seedling establishment in Borago officinalis L. J. Med. Plant Res. 2012, 6, 1896–1901. [Google Scholar]
- Zargar, S.M.; Agnihotri, A. Impact of silicon on various agro-morphological and physiological parameters in maize and revealing its role in enhancing water stress tolerance. Emir. J. Food. Agric. 2013, 25, 138–141. [Google Scholar]
- Hameed, A.; Sheikh, M.A.; Jamil, A.; Basra, S.M.A. Seed priming with sodium silicate enhances seed germination and seedling growth in wheat (Triticum aestivum L.) under water deficit stress induced by polyethylene glycol. Pak. J. Life Soc. Sci. 2013, 11, 19–24. [Google Scholar]
- Biju, S.; Fuentes, S.; Gupta, D. Silicon improves seed germination and alleviates drought stress in lentil crops by regulating osmolytes, hydrolytic enzymes and antioxidant defense system. Plant Physiol. Biochem. 2017, 119, 250–264. [Google Scholar] [CrossRef]
- Hacisalihoglu, G.; White, J. Determination of vigor differences in pepper seeds by using radicle area test. Soil Plant Sci. 2010, 60, 335–340. [Google Scholar] [CrossRef]
- Li, Q.; Ma, C.; Li, H.; Xiao, Y.; Liu, X. Effects of soil available silicon on growth, development and physiological functions of soybean. Chin. J. Appl. Environ. Biol. 2004, 15, 73–76. [Google Scholar]
- Wu, X.; Liu, Y.; Wang, B. Effect of silicon and decomposed liquids from asparagus bean stubs on the growth and antioxidant system in Vigna unguiculata W. ssp. sesquipedlis. Acta Agric. Shanghai 2008, 20, 49. [Google Scholar]
- Zhang, D.; Chen, L.; Li, D.; Lv, B.; Chen, Y.; Chen, J.; Liang, J. OsRACK1 is involved in abscisic acid-and H2O2-mediated signaling to regulate seed germination in rice (Oryza sativa L.). PLoS ONE 2014, 9, e97120. [Google Scholar] [CrossRef]
- Tripathy, J.N.; Zhang, J.; Robin, S.; Nguyen, T.T.; Nguyen, H.T. QTLs for cell-membrane stability mapped in rice (Oryza sativa L.) under drought stress. Theor. Appl. Genet. 2000, 100, 1197–1202. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Ashraf, M.; Arvin, M.J. Alleviation of field water stress in wheat cultivars by using silicon and salicylic acid applied separately or in combination. Crop Pasture Sci. 2019, 70, 36–43. [Google Scholar] [CrossRef]
- Yin, L.; Wang, S.; Liu, P.; Wang, W.; Cao, D.; Deng, X.; Zhang, S. Silicon-mediated changes in polyamine and 1-aminocyclopropane-1-carboxylic acid are involved in silicon-induced drought resistance in Sorghum bicolor L. Plant Physiol. Biochem. 2014, 80, 268–277. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, B.; Jiang, D.; Chen, G. Silicon improves photosynthetic performance by optimizing thylakoid membrane protein components in rice under drought stress. Environ. Exp. Bot. 2019, 158, 117–124. [Google Scholar] [CrossRef]
- Karmollachaab, A.; Bakhshandeh, A.; Gharineh, M.H.; Telavat, M.M.; Fathi, G. Effect of silicon application on physiological characteristics and grain yield of wheat under drought stress condition. Int. J. Agron. Plant Prod. 2013, 4, 30–37. [Google Scholar]
- Shen, X.; Zhou, Y.; Duan, L.; Li, Z.; Eneji, A.E.; Li, J. Silicon effects on photosynthesis and antioxidant parameters of soybean seedlings under drought and ultraviolet-B radiation. J. Plant Phys. 2010, 167, 1248–1252. [Google Scholar] [CrossRef] [PubMed]
- Amin, M.; Ahmad, R.; Ali, A.; Hussain, I.; Mahmood, R.; Aslam, M.; Lee, D.J. Influence of silicon fertilization on maize performance under limited water supply. Silicon 2018, 10, 177–183. [Google Scholar] [CrossRef]
- Faisal, S.; Callis, K.L.; Slot, M.; Kitajima, K. Transpiration-dependent passive silica accumulation in cucumber (Cucumis sativus) under varying soil silicon availability. Pak. J. Bot. 2012, 90, 1058–1064. [Google Scholar] [CrossRef]
- Naseer, M.A.; Hussain, S.; Nengyan, Z.; Ejaz, I.; Ahmad, S.; Farooq, M.; Xiaolong, R. Shading under drought stress during grain filling attenuates photosynthesis, grain yield and quality of winter wheat in the Loess Plateau of China. J. Agron. Crop Sci. 2022, 208, 255–263. [Google Scholar] [CrossRef]
- Wang, Z.; Li, G.; Sun, H.; Ma, L.; Guo, Y.; Zhao, Z.; Gao, H.; Mei, L. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol. Open 2018, 7, bio035279. [Google Scholar] [CrossRef]
- Sherin, G.; Aswathi, K.R.; Puthur, J.T. Photosynthetic functions in plants subjected to stresses are positively influenced by priming. Plant Stress 2022, 4, 100079. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Ming, D.F.; Pei, Z.F.; Naeem, M.S.; Gong, H.J.; Zhou, W.J. Silicon alleviates PEG--induced water--deficit stress in upland rice seedlings by enhancing osmotic adjustment. J. Agron. Crop Sci. 2012, 198, 14–26. [Google Scholar] [CrossRef]
- Ahmed, M.; Hassen, F.; Qadeer, U.; Aslam, M.A. Silicon application and drought tolerance mechanism of sorghum. Afr. J. Agric. Res. 2011, 6, 594–607. [Google Scholar]
- Pereira, T.S.; da Silva Lobato, A.K.; Tan, D.K.Y.; da Costa, D.V.; Uchoa, E.B.; do Nascimento Ferreira, R.; dos Santos Pereira, E.; Avila, F.W.; Marques, D.J.; Silva Guedes, E.M. Positive interference of silicon on water relations, nitrogen metabolism, and osmotic adjustment in two pepper (Capsicum annuum) cultivars under water deficit. Aust. J. Crop Sci. 2013, 7, 1064–1071. [Google Scholar]
- Abdalla, M.M. Beneficial effects of diatomite on the growth, the biochemical contents and polymorphic DNA in Lupinus albus plants grown under water stress. Agric. Biol. J. N. Am. 2011, 2, 207–220. [Google Scholar] [CrossRef]
- Zhang, W.J.; Zhang, X.J.; Lang, D.Y.; Li, M.; Liu, H.; Zhang, X.H. Silicon alleviates salt and drought stress of Glycyrrhiza uralensis plants by improving photosynthesis and water status. Biol. Plant. 2020, 64, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Parveen, N. Role of silicon in mitigating the adverse effects of salt stress on growth and photosynthetic attributes of two maize (Zea mays L.) cultivars grown hydroponically. Pak. J. Bot. 2010, 42, 1675–1684. [Google Scholar]
- Zhang, C.; Moutinho-Pereira, J.M.; Correia, C.; Coutinho, J.; Gonçalves, A.; Guedes, A.; Gomes-Laranjo, J. Foliar application of Sili-K® increases chestnut (Castanea spp.) growth and photosynthesis, simultaneously increasing susceptibility to water deficit. Plant Soil 2013, 365, 211–225. [Google Scholar] [CrossRef]
- Khan, W.U.D.; Aziz, T.; Maqsood, M.A.; Farooq, M.; Abdullah, Y.; Ramzani, P.M.A.; Bilal, H.M. Silicon nutrition mitigates salinity stress in maize by modulating ion accumulation, photosynthesis, and antioxidants. Photosynthetica 2018, 56, 1047–1057. [Google Scholar] [CrossRef]
- Gong, H.; Zhu, X.; Chen, K.; Wang, S.; Zhang, C. Silicon alleviates oxidative damage of wheat plants in pots under drought. Plant Sci. 2005, 169, 313–321. [Google Scholar] [CrossRef]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, Y. Silicon priming regulates morpho-physiological growth and oxidative metabolism in maize under drought stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef]
- Dehghanipoodeh, S.; Ghobadi, C.; Baninasab, B.; Gheysari, M.; Shiranibidabadi, S. Effect of silicon on growth and development of strawberry under water deficit conditions. Hortic. Plant J. 2018, 4, 226–232. [Google Scholar] [CrossRef]
- Li, S.; Chen, J.; Zuo, Q. Influences of optimizing fertilization on the growth and yield of rice variety Wandao68. J. Anhui. Agric. Sci. 2007, 35, 8571–8573. [Google Scholar]
- Das, S.; Pattanayak, S. Nanotechnological approaches in sustainable agriculture and plant disease management. Org. Agric. 2020, 13–30. [Google Scholar] [CrossRef]
- Raven, J.A. Cycling silicon: The role of accumulation in plants. New Phytol. 2003, 158, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Mitani, N.; Nagao, S.; Konishi, S.; Tamai, K.; Iwashita, T.; Yano, M. Characterization of the silicon uptake system and molecular mapping of the silicon transporter gene in rice. Plant Physiol. 2004, 136, 3284–3289. [Google Scholar] [CrossRef]
- Hattori, T.; Sonobe, K.; Inanaga, S.; An, P.; Tsuji, W.; Araki, H.; Eneji, A.E.; Morita, S. Short term stomatal responses to light intensity changes and osmotic stress in sorghum seedlings raised with and without silicon. Environ. Exp. Bot. 2007, 60, 177–182. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, L.; Higgs, D. Effect of silicon on plant growth and mineral nutrition of maize grown under water-stress conditions. J. Plant Nutr. 2006, 29, 1469–1480. [Google Scholar] [CrossRef]
- Ali, A.; Tahir, M.; Amin, M.; Basra, S.M.A.; Maqbool, M.; Lee, D. Si induced stress tolerance in wheat (Triticum aestivum L.) hydroponically grown under water deficit conditions. Bulg. J. Agric. Sci. 2013, 19, 952–958. [Google Scholar]
- Gong, H.; Chen, K. The regulatory role of silicon on water relations, photosynthetic gas exchange, and carboxylation activities of wheat leaves in field drought conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Amin, M.; Ahmad, R.; Basra, S.; Murtaza, G. Silicon induced improvement in morpho-physiological traits of maize (Zea Mays L.) under water deficit. Pak. J. Agric. Sci. 2014, 51, 187–196. [Google Scholar]
- Ahmed, M.; Hassan, F.; Asif, M. Amelioration of drought in sorghum (Sorghum bicolor L.) by silicon. Commun. Soil Sci. Plant Anal. 2014, 45, 470–486. [Google Scholar] [CrossRef]
- Kurdali, F.; Mohammad, A.C.; Ahmad, M. Growth and nitrogen fixation in silicon and/or potassium fed chickpeas grown under drought and well watered conditions. J. Stress Physiol. Biochem. 2013, 9, 385–406. [Google Scholar]
- Ma, C.C.; Li, Q.F.; Gao, Y.B.; Xin, T.R. Effects of silicon application on drought resistance of cucumber plants. Soil Sci. Plant Nutr. 2004, 50, 623–632. [Google Scholar] [CrossRef]
- Gunes, A.; Pilbeam, D.J.; Inal, A.; Coban, S. Influence of silicon on sunflower cultivars under drought stress, I: Growth, antioxidant mechanisms, and lipid peroxidation. Comm. Soil Sci. Plant Anal. 2008, 39, 1885–1903. [Google Scholar] [CrossRef]
- Gunes, A.; Kadioglu, Y.K.; Pilbeam, D.J.; Inal, A.; Coban, S.; Aksu, A. Influence of silicon on sunflower cultivars under drought stress, II: Essential and nonessential element uptake determined by polarized energy dispersive X-ray fluorescence. Commun. Soil Sci. Plant Anal. 2008, 39, 1904–1927. [Google Scholar] [CrossRef]
- Gong, H.J.; Chen, K.M.; Chen, G.C.; Wang, S.M.; Zhang, C.L. Effects of silicon on growth of wheat under drought. J. Plant Nutr. 2003, 26, 1055–1063. [Google Scholar] [CrossRef]
- Lux, A.; Luxová, M.; Abe, J.; Tanimoto, E.; Hattori, T.; Inanaga, S. The dynamics of silicon deposition in the sorghum root endodermis. New Phytol. 2003, 158, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Putra, E.T.S.; Zakaria, W.; Abdullah, N.A.P.; Saleh, G.B. Stomatal morphology, conductance and transpiration of Musa sp. cv. Rastali in relation to magnesium, boron and silicon availability. Am. J. Plant Physiol. 2012, 7, 84–96. [Google Scholar] [CrossRef]
- Saud, S.; Li, X.; Chen, Y.; Zhang, L.; Fahad, S.; Hussain, S.; Sadiq, A.; Chen, Y. Silicon application increases drought tolerance of Kentucky bluegrass by improving plant water relations and morphophysiological functions. Sci. World J. 2014, 2014, 368694. [Google Scholar] [CrossRef]
- Liang, Y.; Sun, W.; Zhu, Y.G.; Christie, P. Mechanisms of silicon-mediated alleviation of abiotic stresses in higher plants: A review. Environ. Poll. 2007, 147, 422–428. [Google Scholar] [CrossRef] [Green Version]
- Mandlik, R.; Thakral, V.; Raturi, G.; Shinde, S.; Nikolić, M.; Tripathi, D.K.; Sonah, H.; Deshmukh, R. Significance of silicon uptake, transport, and deposition in plants. J. Exp. Bot. 2020, 71, 6703–6718. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S.P. Response of to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Qadeer, U.; Ahmed, Z.I.; Hassan, F.U. Improvement of wheat (Triticum aestivum) drought tolerance by seed priming with silicon. Arch. Agron. Soil Sci. 2016, 62, 299–315. [Google Scholar] [CrossRef]
- Crusciol, C.A.; Pulz, A.L.; Lemos, L.B.; Soratto, R.P.; Lima, G.P. Effects of silicon and drought stress on tuber yield and leaf biochemical characteristics in potato. Crop Sci. 2009, 49, 949–954. [Google Scholar] [CrossRef]
- Maghsoudi, K.; Emam, Y.; Ashraf, M.; Pessarakli, M.; Arvin, M.J. Silicon application positively alters pollen grain area, osmoregulation and antioxidant enzyme activities in wheat plants under water deficit conditions. J. Plant Nutr. 2019, 42, 2121–2132. [Google Scholar] [CrossRef]
- Farouk, S.; Omar, M.M. Sweet basil growth, physiological and ultrastructural modification, and oxidative defense system under water deficit and silicon forms treatment. J. Plant Growth Regul. 2020, 39, 1307–1331. [Google Scholar] [CrossRef]
- Sonobe, K.; Hattori, T.; An, P.; Tsuji, W.; Eneji, A.E.; Kobayashi, S.; Kawamura, Y.; Tanaka, K.; Inanaga, S. Effect of silicon application on sorghum root responses to water stress. J. Plant Nutr. 2010, 34, 71–82. [Google Scholar] [CrossRef]
- Ahmad, S.T.; Haddad, R. Study of silicon effects on antioxidant enzyme activities and osmotic adjustment of wheat under drought stress. Czech J. Genet. Plant Breed. 2011, 47, 17–27. [Google Scholar] [CrossRef]
- Hamayun, M.; Sohn, E.Y.; Khan, S.A.; Shinwari, Z.K.; Khan, A.L.; Lee, I.J. Silicon alleviates the adverse effects of salinity and drought stress on growth and endogenous plant growth hormones of soybean (Glycine max L.). Pak. J. Bot. 2010, 42, 1713–1722. [Google Scholar]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Poll. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Lee, J.W.; Kim, Y.C.; Yun, H.K.; Seo, T.C.; Park, K.W. Influence of silicate application on the sucrose synthetic enzyme activity of tomato in perlite media culture. In Proceedings of the XXVI International Horticultural Congress: Protected Cultivation 2002: In Search of Structures, Systems and Plant Materials for 633, Toronto, ON, Canada, 11–17 August 2002; pp. 259–262. [Google Scholar]
- Tripathi, D.K.; Bashri, G.; Shweta, S.; Ahmad, P.; Singh, V. Efficacy of silicon against aluminum toxicity in plants: An overview. Silicon Plants 2017, 1, 355–366. [Google Scholar]
- Ahmad, P.; Sarwat, M.; Sharma, S. Reactive oxygen species, antioxidants and signaling in plants. J. Plant Biol. 2008, 51, 167–173. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Biol. 2000, 51, 463–499. [Google Scholar] [CrossRef] [PubMed]
- Gunes, A.; Pilbeam, D.J.; Inal, A.; Bagci, E.G.; Coban, S. Influence of silicon on antioxidant mechanisms and lipid peroxidation in chickpea (Cicer arietinum L.) cultivars under drought stress. J. Plant Interact. 2007, 2, 105–113. [Google Scholar] [CrossRef]
- Gong, H.J.; Chen, K.M.; Zhao, Z.G.; Chen, G.C.; Zhou, W.J. Effects of silicon on defense of wheat against oxidative stress under drought at different developmental stages. Biol. Plant. 2008, 52, 592–596. [Google Scholar] [CrossRef]
- Ahmad, M.; El-Saeid, M.H.; Akram, M.A.; Ahmad, H.R.; Haroon, H.; Hussain, A. Silicon fertilization–a tool to boost up drought tolerance in wheat (Triticum aestivum L.) crop for better yield. J. Plant Nutr. 2016, 39, 1283–1291. [Google Scholar] [CrossRef]
- Liu, H.; Guo, Z. Effects of supplementary silicon on nitrogen, phosphorus and potassium contents in the shoots of Medicago sativa plants and in the soil under different soil moisture conditions. Chin. J. Appl. Environ. Biol. 2011, 17, 809–813. [Google Scholar]
- Emam, M.M.; Khattab, H.E.; Helal, N.M.; Deraz, A.E. Effect of selenium and silicon on yield quality of rice plant grown under drought stress. Aust. J. Crop Sci. 2014, 8, 596–605. [Google Scholar]
- Alsaeedi, A.; El-Ramady, H.; Alshaal, T.; El-Garawany, M.; Elhawat, N.; Al-Otaibi, A. Silica nanoparticles boost growth and productivity of cucumber under water deficit and salinity stresses by balancing nutrients uptake. Plant Physiol. Biochem. 2019, 139, 1–10. [Google Scholar] [CrossRef]
- Liang, Y.C.; Qirong, S.; Zhenguo, S. Effect of silicon on enzyme activity and sodium, potassium and calcium concentration in barley under salt stress. J. Plant Soil. 1999, 209, 217–224. [Google Scholar] [CrossRef]
- Liu, P.; Yin, L.; Wang, S.; Zhang, M.; Deng, X.; Zhang, S.; Tanaka, K. Enhanced root hydraulic conductance by aquaporin regulation accounts for silicon alleviated salt-induced osmotic stress in Sorghum bicolor L. Environ. Exp. Bot. 2015, 111, 42–51. [Google Scholar] [CrossRef]
- Chen, D.; Cao, B.; Wang, S.; Liu, P.; Deng, X.; Yin, L.; Zhang, S. Silicon moderated the K deficiency by improving the plant-water status in sorghum. Sci. Rep. 2016, 6, 22882. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.X.; Xu, X.B.; Hu, Y.H.; Han, W.H.; Yin, J.L.; Li, H.L.; Gong, H.J. Silicon improves salt tolerance by increasing root water uptake in Cucumis sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Wang, S.; Yin, L.; Deng, X. How does silicon mediate plant water uptake and loss under water deficiency? Front. Plant Sci. 2018, 9, 281. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Guo, J.; Duan, Y.; Zhang, T.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi1, a silicon transporter gene in Cucumis sativus. Physiol. Plant. 2017, 159, 201–214. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Duan, Y.; Qi, X.; Zhang, L.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi2, a cucumber silicon efflux transporter gene. Ann. Bot. 2018, 122, 641–648. [Google Scholar] [CrossRef] [PubMed]
- Krysan, P.J.; Young, J.C.; TAx, F.; Sussman, M.R. Identification of transferred DNA insertions within Arabidopsis genes involved in signal transduction and ion transport. Proc. Natl. Acad. Sci. USA 1996, 93, 8145–8150. [Google Scholar] [CrossRef] [PubMed]
- Piperno, D.R.; Holst, I.; Wessel-Beaver, L.; Andres, T.C. Evidence for the control of phytolith formation in Cucurbita fruits by the hard rind (Hr) genetic locus: Archaeological and ecological implications. Proc. Natl. Acad. Sci. USA 2002, 99, 10923–10928. [Google Scholar] [CrossRef]
- Dorweiler, J.E.; Doebley, J. Developmental analysis of teosinte glume architecture1: A key locus in the evolution of maize (Poaceae). Am. J. Bot. 1997, 84, 1313–1322. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.C.; Keeling-Tucker, T. Biosilicification: The role of the organic matrix in structure control. J. Biol. Inorg. Chem. 2000, 5, 537–550. [Google Scholar] [CrossRef] [PubMed]
- Fawe, A.; Menzies, J.G.; Chérif, M.; Bélanger, R.R. Silicon and disease resistance in dicotyledons. Plant Biol. 2001, 8, 159–169. [Google Scholar]
- Mitani, N.; Ma, J.F. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef]
- Liu, P.; Yin, L.; Deng, X.; Wang, S.; Tanaka, K.; Zhang, S. Aquaporin-mediated increase in root hydraulic conductance is involved in silicon-induced improved root water uptake under osmotic stress in Sorghum bicolor L. J. Exp. Bot. 2014, 65, 4747–4756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khattab, H.I.; Emam, M.A.; Emam, M.M.; Helal, N.M.; Mohamed, M.R. Effect of selenium and silicon on transcription factors NAC5 and DREB2A involved in drought-responsive gene expression in rice. Biol. Plant. 2014, 58, 265–273. [Google Scholar] [CrossRef]
- Al-Mayahi, A.M.W. Effect of silicon (Si) application on Phoenix dactylifera L. growth under drought stress induced by polyethylene glycol (PEG) in vitro. Am. J. Plant Sci. 2016, 7, 1711–1728. [Google Scholar] [CrossRef]
- Bukhari, M.A.; Ahmad, Z.; Ashraf, M.Y.; Afzal, M.; Nawaz, F.; Nafees, M.; Jatoi, W.N.; Malghani, N.A.; Shah, A.N.; Manan, A. Silicon mitigates drought stress in wheat (Triticum aestivum L.) through improving photosynthetic pigments, biochemical and yield characters. Silicon 2021, 13, 4757–4772. [Google Scholar] [CrossRef]
- Othmani, A.; Ayed, S.; Bezzin, O.; Farooq, M.; Ayed-Slama, O.; Slim-Amara, H.; Ben Younes, M. Effect of silicon supply methods on durum wheat (Triticum durum Desf.) response to drought stress. Silicon 2021, 13, 3047–3057. [Google Scholar] [CrossRef]
- Sattar, A.; Cheema, M.A.; Sher, A.; Abbas, T.; Irfan, M.; Ijaz, M.; Hussain, S.; Ali, Q. Foliage applied silicon alleviates the combined effects of salinity and drought stress on wheat seedlings. Int. J. Agric. Biol. 2018, 20, 2537–2543. [Google Scholar]
- Ahmad, M.; Akram, Z.; Munir, M.; Rauf, M. Physio-morphic response of wheat genotypes under rainfed conditions. Pak. J. Bot. 2006, 38, 1697–1702. [Google Scholar]
- Bukhari, M.A.; Ashraf, M.Y.; Ahmad, R.; Waraich, E.A.; Hameed, M. Improving drought tolerance potential in wheat (Triticum aestivum L.) through exogenous silicon supply. Pak. J. Bot. 2015, 47, 1641–1648. [Google Scholar]
- Gharineh, M.H.; Karmollachaab, A. Effect of silicon on physiological characteristics wheat growth under Water-Deficit Stress Induced by PEG. Int. J. Agron. Plant Prod. 2013, 4, 1543–1548. [Google Scholar]
- Abd El-Mageed, T.A.; Shaaban, A.; Abd El-Mageed, S.A.; Semida, W.M.; Rady, M.O.A. Silicon defensive role in maize (Zea mays L.) against drought stress and metals-contaminated irrigation water. Silicon 2020, 13, 2165–2176. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon improves water use efficiency in maize plants. J. Plant Nutr. 2005, 27, 1457–1470. [Google Scholar] [CrossRef]
- Cao, B.L.; Ma, Q.; Xu, K. Silicon restrains drought-induced ROS accumulation by promoting energy dissipation in leaves of tomato. Protoplasma 2020, 257, 537–547. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, S.H.I.; Gong, H.J.; Zhao, H.L.; Li, H.L.; Hu, Y.H.; Wang, Y.C. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integr. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef]
- Cao, B.L.; Wang, L.; Gao, S.; Xia, J.; Xu, K. Silicon-mediated changes in radial hydraulic conductivity and cell wall stability are involved in silicon-induced drought resistance in tomato. Protoplasma 2017, 254, 2295–2304. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, J.; Gong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, M.A.; Merwad, A.R.M.; Elnaka, E.A. Rice (Oryza Sativa L.) tolerance to drought can be improved by silicon application. Commun. Soil Sci. Plant Anal. 2018, 49, 945–957. [Google Scholar] [CrossRef]
- Ullah, H.; Luc, P.D.; Gautam, A.; Datta, A. Growth, yield and silicon uptake of rice (Oryza sativa) as influenced by dose and timing of silicon application under water-deficit stress. Arch. Agron. Soil Sci. 2018, 64, 318–330. [Google Scholar] [CrossRef]
- Nolla, A.; de Faria, R.J.; Korndoerfer, G.H.; da Silva, T.B. Effect of silicon on drought tolerance of upland rice. J. Food Agric. Environ. 2012, 10, 269–272. [Google Scholar]
- Avila, R.G.; Magalhães, P.C.; da Silva, E.M.; Gomes Júnior, C.C.; de Paula Lana, U.G.; de Alvarenga, A.A.; de Souza, T.C. Silicon supplementation improves tolerance to water deficiency in sorghum plants by increasing root system growth and improving photosynthesis. Silicon 2020, 12, 2545–2554. [Google Scholar] [CrossRef]
- Verma, K.K.; Song, X.P.; Zeng, Y.; Li, D.M.; Guo, D.J.; Rajput, V.D.; Chen, G.L.; Barakhov, A.; Minkina, T.M.; Li, Y.R. Characteristics of leaf stomata and their relationship with photosynthesis in Saccharum officinarum under drought and silicon application. ACS Omega 2020, 5, 24145–24153. [Google Scholar] [CrossRef] [PubMed]
- de Camargo, M.S.; Bezerra, B.K.L.; Holanda, L.A.; Oliveira, A.L.; Vitti, A.C.; Silva, M.A. Silicon fertilization improves physiological responses in sugarcane cultivars grown under water deficit. J. Soil Sci. Plant Nutr. 2019, 19, 81–91. [Google Scholar] [CrossRef]
- Asgharipour, M.R.; Mosapour, H.A. Foliar application silicon enhances drought tolerance in fennel. J. Anim. Plant Sci. 2016, 26, 1056–1062. [Google Scholar]
- Mosapour, Y.H.; Asgharipour, M.R. Effects of drought stress and its interaction with silicon on stimulates the antioxidant system and lipid peroxidation in fennel (Foeniculum vulgar). J. Plant Proc. Func. 2016, 5, 71–84. [Google Scholar]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Khan, M.I.R.; Fujita, M. Silicon-mediated regulation of antioxidant defense and glyoxalase systems confers drought stress tolerance in Brassica napus L. S. Afr. J. Bot. 2018, 115, 50–57. [Google Scholar] [CrossRef]
- Hamid, N.; Naz, B.; Rehman, A. Effect of exogenous application of silicon with drought stress on protein and carbohydrate contents of edible beans (Vigna radiate & Vigna unguiculata). J. Chem. Soc. Pak. 2012, 2, 99–105. [Google Scholar]
- Hajiboland, R.; Cheraghvareh, L.; Poschenrieder, C. Improvement of drought tolerance in Tobacco (Nicotiana rustica L.) plants by Silicon. J. Plant Nutr. 2017, 40, 1661–1676. [Google Scholar] [CrossRef]
- Desoky, E.S.M.; Mansour, E.; Yasin, M.A.; El Sobky, E.S.E.; Rady, M.M. Improvement of drought tolerance in five different cultivars of Vicia faba with foliar application of ascorbic acid or silicon. Span. J. Agric. Res. 2020, 18, e0802. [Google Scholar] [CrossRef]
- Moradtalab, N.; Hajiboland, R.; Aliasgharzad, N.; Hartmann, T.E.; Neumann, G. Silicon and the association with an arbuscular-mycorrhizal fungus (Rhizophagus clarus) mitigate the adverse effects of drought stress on strawberry. Agronomy 2019, 9, 41. [Google Scholar] [CrossRef]
- Alam, A.; Hariyanto, B.; Ullah, H.; Salin, K.R.; Datta, A. Effects of silicon on growth, yield and fruit quality of cantaloupe under drought stress. Silicon 2021, 13, 3153–3162. [Google Scholar] [CrossRef]
- Habibi, G.; Hajiboland, R. Alleviation of drought stress by silicon supplementation in pistachio (Pistacia vera L.) plants. Folia Hortic. 2013, 25, 21–29. [Google Scholar] [CrossRef]
- Helaly, M.N.; El-Hoseiny, H.; El-Sheery, N.I.; Rastogi, A.; Kalaji, H.M. Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Biochem. 2017, 118, 31–44. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crop | Drought Level | Source and Rate | Method of Application | Experimental Conditions | Major Findings | Reference |
|---|---|---|---|---|---|---|
| Phoenix dactylifera L. | PEG-8000 (15%) | 3.6 mM Si Ca2SiO4 | Nutrient medium | In vitro (MS media) | Improved physiological (Chlorophyll contents) and biochemical attributes, and antioxidant activity | [132] |
| Triticum aestivum L. | 60% of FC | 4 mM Si (K2SiO4) | Foliar spray at anthesis | Pot | Improved biochemical attributes and accumulation of nutrients | [133] |
| −0.3 MPa | 150 mg L−1 Si (Na2SiO3. H2O) | Nutrient medium | Hydroponics | Higher chlorophyll content, relative water content, and shoot and root length | [134] | |
| 100 and 40% of FC | 6 mM Si (Na2Si3O7) | Foliar application | Field | Enhanced activity of key antioxidant enzymes, accumulation of osmolytes, and reduction in H2O2 and MDA levels | [54] | |
| 40% WHC | 50 mM Si | Foliar application | Pot | Significant increase in photosynthetic pigments, photosynthesis rate, stomatal conductance, leaf turgor pressure, and enzymatic antioxidants | [135] | |
| Irrigation frequencies (2, 3 and 4) | 0–12 kg ha−1 (K2SiO3) | Soil | Field | Improved growth and yield; higher tissue K+ ion contents | [136] | |
| 60% of FC | 1 mM Na2SiO3.5H2O solution per pot | Fertigation | Pot | Higher activity of antioxidant enzymes such as APX, POD and CAT | [137] | |
| PEG-6000 (20%) | 1.0 mM Si (K2SiO4) | Soil | Hydroponics | Increased total soluble protein contents and SOD activity, and reduced electrolyte leakage | [138] | |
| 75, 50 and 25% of FC | 30 kg Si ha−1 | Soil | Field | Reduced electrolyte leakage and increased SOD activity, and grain K+ and Si concentrations | [57] | |
| Withholding irrigation | 0.28 g Si kg−1 soil | Soil | Field | Improved tissue water status and leaf stomatal conductance; increased phosphoenol pyruvate carboxylase (PEPCase) activity | [83] | |
| Withholding water for 12 days in 26-day-old seedlings | 7.14 mmol Si kg−1 soil | Soil | Pot | Greater leaf water ratio (LWR) and lower specific leaf area (SLA) | [90] | |
| Zea mays L. | Deficit irrigation (DI20% = 80% of full irrigation) | 4 mM Si (Na2SiO3) | Foliar application | Field | Enhanced physiological performance, water use efficiency, and productivity of maize under water deficit | [139] |
| 60% of FC | 0, 4 and 6 mM Si (Na2SiO3) | Seed priming | Pot | Increased shoot and root length, and photosynthetic pigments; enhanced activity of antioxidant enzymes (SOD, POD and CAT) and reduced MDA and H2O2 contents | [74] | |
| 100 and 60% of FC | 150 mg Ca2SiO4 kg−1 soil | Foliar | Pot | Improved growth, dry matter, plant–water relations and gas exchange attributes | [84] | |
| Naturally rain-fed conditions | 700 kg Ca2SiO4 ha−1 | Soil | Field | Increased kernel number per ear; improved relative water content and stomatal density, and reduced canopy temperature | [46] | |
| 50% of FC | 1–2 mM Si (Na2SiO3) | Soil | Pot | Improved physiological attributes, i.e., chlorophyll and relative water contents; higher tissue Ca and K contents | [81] | |
| PEG-6000 | 2 mmol L−1 (H4SiO4) | Nutrient medium | Hydroponics | Higher WUE; lower transpiration rate | [140] | |
| Solanum lycopersicum L. | 1.0% PEG | 0.6 mM Si (Na2SiO3) | Nutrient medium | Hydroponics | Improved energy dissipation in mitochondria and chloroplasts via Si-mediated alternative oxidase and malate/oxaloacetate shuttle, and reduced ROS accumulation | [141] |
| (PEG-6000) 10% (w/v) | 2.5 mmol L−1 Si (K2SiO3) | Nutrient medium | Hydroponics | Promoted photosynthesis by modulating some photosynthesis-related genes and regulating the photochemical process | [142] | |
| 1.0% PEG | 0.6 mM Si (Na2SiO3) | Nutrient solution | Greenhouse | Improved hydraulic conductivity in radial direction, which enhanced water uptake of tomato roots; high solute accumulation, such as proline, soluble sugar and soluble protein; enhanced antioxidant activity (SOD and CAT) and reduced O2− production rate, and H2O2 and MDA contents | [143] | |
| (PEG-6000) 10% (w/v) | 2.5 mmol L−1 Si (K2SiO3) | Nutrient medium | Hydroponics | Enhanced root hydraulic conductance and water uptake, and decreased membrane oxidative damage | [144] | |
| PEG-6000 (10%) | 0.5 mM Si | Nutrient medium | Hydroponics | Increased seed germination and antioxidant enzymes activity, i.e., SOD, CAT, APX and POD | [43] | |
| Oryza Sativa L. | 70, 80, 90, 100 and 120% of the soil saturation point | 0, 2.1, 4.2, 6.3 and 8.4 mg Si per pot. (K-Silicate) | Foliar spray | pot | Increased plant height, rice straw, root yield and grain yield; increased concentrations of N, P, K and Si in straw | [145] |
| 100, 75 and 50% of FC | 0, 75, 150, 300 and 600 kg Si ha−1 | Soil | Pot | Increased grain yield (34–45%) and highest number of panicles per plant | [146] | |
| 80, 70 and 60% of FC | 0, 200, 400 and 600 kg Si ha−1. (Calcium. Silicate) | Soil | Field | Increased grain yield and plant height, and reduced number of stalks | [147] | |
| Sorghum bicolor L. | −138 KPa | 2 mM Si (K2SiO3) | Soil | Pot | Increased growth of the root system, and improved photosynthesis | [148] |
| PEG-6000 (−4.0, −6.0, −8.0 and −10.0 MPa) | 3.0 mmol Si L−1) (K2SiO3) | Solution application | Pot | Increased leaf water potential, leaf area index, SPAD chlorophyll, net assimilation and relative growth rate | [85] | |
| 40 mm of irrigation | 200 mL L−1 kg−1 soil | Soil | Pot | Improved plant–water relations and higher chlorophyll contents | [66] | |
| Saccharum officinarum L. | 100–95, 55–50 and 35–30% of FC | 0, 100, 300 and 500 mg L−1 Si (CaO·SiO2) | Fertigation | Pot | Increased plant growth, photosynthetic efficiency and biomass/yield; promoted better adaptation of stomata to drought | [149] |
| 55% of FC | 0 and 600 kg Si ha−1 | Soil | Field | Increased relative water content, dry weight of leaves, carotenoids content, leaf water potential (Ψw) and SPAD, and decreased electrolyte leakage in leaves | [150] | |
| Glycine max L. | −0.5 MPa PEG-6000 (20%) | 1.70 mM Si | Nutrient medium | Hydroponics | Improved photosynthesis and antioxidant activity (i.e., CAT, POD, SOD and H2O2); increased proline and relative water contents | [58] |
| PEG-6000 | 100–200 mg Si L−1 | Nutrient medium | Hydroponics | Improved growth and hormonal homeostasis | [103] | |
| Foeniculum vulgar L. | 90, 75 and 60% of FC | 0, 2.5, 5, 7.5 and 10 mM Si | Foliar application | Field | Increased concentrations of proline, soluble sugars, Na+ and K+ in seeds; improved LAI, seed yield and essential oil percentage | [151] |
| 40, 70 and 100% of FC | 0, 2.5, 5, 7.5 and 10 mM | Foliar | Field | Enhanced ascorbate peroxidase activity and increased osmotic adjustment | [152] | |
| Brassica napus L. | 10 and 20% PEG | Si (1 mM SiO2) | Hyponex solution | Semi-hydroponic | Significantly improved antioxidant enzymes, AsA-GSH pool, glyoxalase systems and proline | [153] |
| Vigna radiata L. | 100, 75, 50 and 25% soil moisture | 20, 40 and 60 ppm MgSiO3 | Fertigation | Pot | Increased total carbohydrate and protein contents | [154] |
| Nicotiana Rustica L. | 100, 60 and 30% of FC | 2 mM Si (Na2SiO3) | Solution application | Pots having Perlite | Increased soluble proteins, free α-amino acids and proline concentrations; enhanced activity of antioxidative enzymes and decreased H2O2 concentration | [155] |
| Vicia faba L. | 4000, 3000 and 2000 m3 water ha−1 | 2.0 mM Si (K2SiO3) | Foliar application | Field | Improved growth, yields and water use efficiency by raising antioxidant activities and suppressing lipid peroxidation and electrolyte leakage | [156] |
| Fragaria x ananasa var. Paros | 90, 75 and 35% WHC | 3 mmol Si L−1 (Na2SiO3) | Nutrient solution | Hydroponics | Enhanced plant biomass production by increasing photosynthesis rate, water content and use efficiency, antioxidant enzyme defense and nutritional status | [157] |
| Cucumis melo L. | 100, 75 and 50% of FC | 0, 100, 200 and 400 kg Si ha−1 (H4SiO4) | Soil | Pot | Higher yield, flesh thickness and total soluble solids content | [158] |
| Poa pratensis L. | Withholding irrigation for 20 days | 0, 200, 400 and 800 mg Si L−1 (Na2SiO3⋅9H2O) | Nutrient solution | Glasshouse (Vermiculite + loam soil) | Increased net photosynthesis, leaf water contents, leaf green color and turf quality | [93] |
| Ocimum basilicum L. | 45 and 65% of FC | 250 mg L−1 Si | Soil | Pot | Improved essential oil yield, antioxidant system and osmoregulation, and maintained organelle ultrastructure | [100] |
| Pistacia vera L. | 70% of FC | 2.73 mmol Si kg−1 soil (Na2SiO3) | Soil | Pot | Enhanced photochemical efficiency and photosynthetic gas exchange; improved maximum quantum yield of PSII, stomatal conductance and antioxidant defense capacity | [159] |
| Mangifera indica L. | −0.77-bar water potential | 1.5 mM Si (K2SiO3) | Fertigation through drip irrigation | Field | Increased IAA, GA and CK, and decreased ABA levels; improved antioxidative enzyme activity and fruit quality | [160] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Irfan, M.; Maqsood, M.A.; Rehman, H.u.; Mahboob, W.; Sarwar, N.; Hafeez, O.B.A.; Hussain, S.; Ercisli, S.; Akhtar, M.; Aziz, T. Silicon Nutrition in Plants under Water-Deficit Conditions: Overview and Prospects. Water 2023, 15, 739. https://doi.org/10.3390/w15040739
Irfan M, Maqsood MA, Rehman Hu, Mahboob W, Sarwar N, Hafeez OBA, Hussain S, Ercisli S, Akhtar M, Aziz T. Silicon Nutrition in Plants under Water-Deficit Conditions: Overview and Prospects. Water. 2023; 15(4):739. https://doi.org/10.3390/w15040739
Chicago/Turabian StyleIrfan, Muhammad, Muhammad Aamer Maqsood, Hafeez ur Rehman, Wajid Mahboob, Nadeem Sarwar, Osama Bin Abdul Hafeez, Saddam Hussain, Sezai Ercisli, Muhammad Akhtar, and Tariq Aziz. 2023. "Silicon Nutrition in Plants under Water-Deficit Conditions: Overview and Prospects" Water 15, no. 4: 739. https://doi.org/10.3390/w15040739