
Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1
by
Shuaiguo Li
1,
Kejian Tian
1,
Qing Qiu
1,
Yue Yu
1,
Han Li
1,
Menghan Chang
1,
Xuejian Sun
1,
Jinming Gu
1,
Fenglin Zhang
1,
Yibing Wang
1 and
Hongliang Huo
1,2,3,* 1
School of Environment, Northeast Normal University, Changchun 130024, China
2
Jilin Province Laboratory of Water Pollution Treatment and Resource Engineering, Changchun 130117, China
3
Northeast China Low Carbon Water Pollution Treatment and Green Development Engineering Research Center, Changchun 130117, China
*
Author to whom correspondence should be addressed.
Water 2023, 15(4), 830; https://doi.org/10.3390/w15040830
Submission received: 13 January 2023
/
Revised: 15 February 2023
/
Accepted: 15 February 2023
/
Published: 20 February 2023
(This article belongs to the Special Issue Functional Microorganisms in Wastewater Treatment)
Abstract
:As a widespread pollutant, bisphenol A (BPA) has created a serious threat to ecosystem and human health. Therefore, expanding the available microbial resources used to screen highly efficient BPA-degrading bacteria with BPA as the sole carbon source is very important for the removal of this pollutant from the environment. In this study, the BPA degradation rate of Pseudomonas sp. P1 to 30 mg/L was 96.89% within 120 h. Whole genome sequencing showed that the genome of strain P1 was composed of a single circular chromosome with a full length of 6.17 Mb, which contained 5636 predicted coding genes. Comparative genomic analysis showed that strain P1 contained 210 functional genes related to BPA degradation. It was confirmed that BPA degradation genes ferredoxin (bisdA), P450 (bisdB), CotA and Lac in strain P1 were highly expressed under the induction of BPA. Combined with the identification of metabolites, the route of BPA degradation by Pseudomonas was proposed. A new metabolite, 4-vinylphenol, was detected for the first time in pathway Ⅰ. In pathway Ⅱ, BPA is directly oxidized to phenol and 4-isopropenyl phenol in the presence of laccase, which is rarely reported in the process of bacterial degradation of BPA. This study confirmed that strain P1 had good tolerance to various environmental factors at the gene level and enriched the degradation mechanism of BPA.
1. Introduction
Bisphenol A (BPA) is one of many chemicals with important commercial value and high output worldwide [1,2]. Because of its cross-linking properties, BPA is an effective plasticizer that is widely used in the manufacture of polycarbonate plastics and epoxy resins, which are further used in the production of packaging materials including reusable tableware, storage containers, and health care equipment [3]. In addition, during the pandemic caused by COVID-19, a large amount of plastic was used as protective equipment, resulting in a large amount of plastic waste, the consequences of which have yet to be resolved [4,5]. Nanoplastics (NPs) are formed when plastic fragments are subjected to ultraviolet radiation, natural weathering, and biodegradation [6]. The consumption of large volumes of BPA has led to an accumulation of BPA in the environment. Many environmental investigations have shown that BPA is commonly found in surface water, groundwater, wastewater from sewage treatment plants, and landfill leachate. The concentrations of BPA in groundwater and surface drinking water sources in North America and Europe are <0.0001–6.4 μg/L and <0.0002–5.1 μg/L, respectively, and the BPA levels in drinking water and surface drinking water sources in Asia are in the range of <0.0007–4.23 μg/L [7]. The concentrations of BPA detected around farms, surface water, and sewage treatment plants in South Africa reached 1.14 μg/L, 0–181.28 μg/L, and Nd-0.02 μg/L, respectively [8,9,10], and BPA with a concentration of 0.18–0.4 μg/L was also detected in surface water and groundwater in Nigeria [11]. The average concentration of BPA in leachate samples from landfills ranged from 0.70 to 2.72 mg/L, and even reached 12 mg/L in some places [12,13,14]. At the same time, BPA is an endocrine-disrupting chemical that can have harmful effects on organisms by interfering with the thyroid hormone and estrogen and androgen receptors [15,16]. Therefore, even small levels of BPA exposure may have negative effects on organisms, such as damage to the reproductive and nervous systems, lead-to-food web contamination, and drug resistance in pathogens [17] and immune systems, and may even lead to a variety of cancers [18,19]. An effective and reasonable method used to eliminate BPA in the environment urgently needs to be developed for the protection of the environment and human health.
The high chemical stability and low water solubility of BPA make it difficult for BPA to be degraded naturally, and it is usually difficult to completely degrade and remove BPA from the environment using traditional technology [20]. In recent years, a series of physical and chemical methods including physical adsorption, membrane filtration, photocatalysis, and advanced oxidation have been developed [21,22,23,24]. Although these methods have been shown to effectively remove BPA, these methods usually require a large financial cost. In addition, the resulting degradation by-products may be more toxic than BPA itself. Compared with physical and chemical degradation methods, a biological method has its unique advantages in removing pollutants from water [25]. Biodegradation is considered to be a more convenient, economical, and safer method creating the least interference to the environment [26]. Therefore, many studies have been devoted to the removal of BPA by microorganisms. Bacteria such as Picocystis sp., Ochrobactrum sp., and Bacillus and other sphingosine-degrading bacteria can degrade BPA [27,28,29,30]. However, the benzene ring structure of BPA makes it difficult to be metabolized by microorganisms. Therefore, most screened individual types of bacteria usually need to be provided with additional carbon or nitrogen sources, such as yeast or glucose, to improve their degradation of BPA [30,31]. However, in this process, the complexity of degradation will increase along with additional operation and maintenance costs, so a good strain of bacterial resources for the degradation of BPA is still lacking. Therefore, in this experiment, BPA was used as the only carbon source to screen bacteria that could efficiently degrade BPA and study their degradation behavior in water.
Microorganisms can often degrade pollutants using enzymatic reactions regulated by functional genes. In the process of the microbial degradation of BPA, microorganisms will employ a series of metabolic regulation mechanisms to resist BPA stress while further degrading it [32]. Screening the most important genes involved in this process and determining the mechanism of microbial cell degradation of BPA will help researchers to understand the most important factors involved in the degradation process [33]. At present, some achievements have been made in the analysis of the molecular mechanism involved in the biodegradation of BPA, but studies and analysis are needed. Some experiments have studied the effects of individual factors such as temperature, the acidity of a solution, the state of bacteria, or bacterial metabolites on degradation [29,34,35,36]. Some studies have analyzed the removal and degradation pathways of BPA used by microorganisms [35,37]. The analysis of bacterial degradation behavior has mostly focused on the analysis of degradation effects or pathways. Although the BPA degradation pathways have been widely studied, the understanding of the key genes and metabolic mechanisms of BPA degradation is still limited. Therefore, the study of a microbial genome is an important step that can be used to advance our genome knowledge, especially in the BPA-related defense and degradation mechanism of microorganisms. This will promote the design of precise bioremediation technologies for polluted environments.
BPA is one of the most common endocrine disrupters in the environment so it is necessary to screen BPA-degrading bacteria and study their genome. The main goal of this study is to isolate and identify BPA-degrading bacteria with high efficiency, and analyze the genes involved in BPA degradation and toxicity resistance, so as to fully understand the genomic characteristics and degradation mechanism of BPA-degrading bacteria. Strain P1 was screened and identified by detecting the degradation efficiency of BPA, and its degradation products were analyzed. The expression of genes encoded by enzymes involved in BPA degradation was verified by qPCR and qPCR to determine the degradation performance of strain P1 under the influence of different environmental factors. On the basis of functional annotation, the defense mechanism of bacteria against toxic pollutants was analyzed and expounded. The purpose of this study is to further enrich the BPA-degrading bacteria bank, improve the BPA degradation and resistance mechanism of strains, and lay a theoretical foundation for microbial remediation of BPA-polluted environments.
2. Materials and Methods
2.1. Experimental Chemicals and Samples
Sediment samples of landfill leachate were collected from a landfill (N: 43°, E: 125°) in Changchun City, Jilin Province, China; the samples were enriched, separated, and purified to obtain bacteria.
In this study, BPA (Product No. B108653) was purchased from Aladdin Chemistry (Shanghai, China); meanwhile, ethyl acetate (Product No. 34858), methanol (Product No. 1.06035), and acetonitrile (Product No. 1.00029) were purchased from Sigma-Aldrich (https://www.sigmaaldrich.cn/CN/zh, (accessed on 2 October 2022)) (Shanghai, China); a bacterial genomic DNA extraction kit was purchased from Beijing Tiangen Biochemical Technology Co., Ltd. (Beijing, China); and an agarose gel electrophoresis recovery kit (QIAquick Gel Extraction Kit (50)) was purchased from Qiagen (Shanghai, China).
2.2. Isolation and Identification of Strains
In this study, BPA was used as the only carbon and energy source for long-term culture. Inorganic salt medium (MSM) containing BPA as the sole carbon source was used for the isolation of BPA-degrading bacteria. One gram of sediment sample was put into MSM medium containing 100 mg/L BPA and cultured at 30 °C and 120 rpm. When BPA degradation was detected, 3 mL of the culture was transferred to MSM containing 100 mg/L BPA and cultured under the same conditions. This process was repeated 10 times in a row to ensure that bacteria that could tolerate BPA stress were screened. The MSM medium contained 4.26 g/L NaHPO4, 2.65 g/L KH2PO4, 0.2 g/L MgSO4·7H2O, 1.5 g/L (NH4)2SO4, and 0.02 g/L CaCl2, pH 7. A total of 1 mL of a trace element solution was added to the MSM, which contained 0.024 g/L NiCl2·6H2O, 0.19 g/L CoCl2·6H2O, 0.002 g/L CuCl2·2H2O, 0.061 g/L MnSO4·H2O, 0.024 g/L Na2MoO4·2H2O, 0.07 g/L ZnCl2, and 0.006 g/L H3BO3.
After the purified single colony was cultured in LB medium for 24 h, the morphology of the strain was observed by scanning under a electron microscope (FEl XL-30, OSU, Columbus, OH, USA), as described by Qiu et al. [38]. The physiological and biochemical characteristics of the strain were analyzed by API 20E G- Bacillus identification kit (Art.No. CN2010010). The 16S rDNA sequence of the strain was then amplified by PCR. The BLAST functional module was used to compare the gene sequence with the 16S rDNA sequence in GenBank database. In addition, MEGA 5.2.1 software was used to analyze the phylogenetic tree according to the Neighbor-Joining method. Each strain has been preserved in the typical Culture Preservation Center of China using preservation no. CCTCCAB2021476 and the preservation address of Bayi Luojia Mountain, Wuchang District, Wuhan City, Hubei Province. The sequence of Pseudomonas sp. P1 gene was submitted to the GenBank database of the National Biotechnology Information Center (NCBI) with the accession number CP0084714.
2.3. Degradation of BPA by Strain P1
In this study, we intended to use single-factor analysis to study these factors affecting the degradation ability of each strain, including temperature, pH, initial concentration of pollutants, inoculum size, and so on. For example, 30 mg/L BPA was added to MSM medium as the only carbon source, and the single-factor change was set up according to Table 1; three repetitions were used in all treatment groups. 100 mL of culture medium was autoclaved in 250 mL conical flasks at 121 °C for 30 min and, after the culture medium cooled, the strain was inoculated into 100 mL aseptic MSM, in which 30 mg/L BPA was the only carbon and energy source. The strains were cultured in triplicate for 120 h. 200 μL. Samples were taken at 6 h, 12 h, 24 h, 48 h, 72 h, 96 h, and 120 h, respectively, from the cultures and cell densities were measured at OD 600 nm using a microplate reader. Samples were taken at the same time point, procreated by equal volume extraction, and then the remaining BPA concentration in the culture medium was measured by high performance liquid chromatography (Shimadzu LC-10AVP, Shimadzu Co., Kyoto, Japan). The chromatographic column was a Zorbax Eclipse Plus C18 column (150 × 4.6 mm, 3.5 mm). The detection conditions were as follows: detection wavelength, 281 nm; acetonitrile: water ratio = 60:40; injection volume, 20 μL; flow rate, 0.6 mL/min; detection temperature, 30 °C. The BPA retention time was 2.89 min. The detection method of BPA degradation products is as described in our previous research [39].
2.4. Genome Sequencing and Functional Annotation
The strain was inoculated in LB liquid medium at 30 °C and 120 rpm for 24 h, and centrifuged at 4000 rpm for 10 min. The supernatant was removed, the cells were re-suspended with a 0.2 M phosphate buffer (pH 7.0), centrifuged again, and the sediment was collected. Genomic DNA was extracted by a bacterial genomic DNA extraction kit, and DNA concentration was measured by NanoDrop ND-1000 ultramicro-UV spectrophotometer. The genome of strain P1 was sequenced by the third generation Nanopore sequencing method. The offline data of Nanopore sequencing were filtered through Guppy v3.2.6 software to filter the reads of low quality and short segments. The filtered subreads was assembled by Canu v1.5 software, the assembly results were corrected by the third generation subreads through the Racon v3.4.3 software, the cyclization and the starting site were adjusted by the Circlator v1.5.5 software, and the second-generation data were further corrected by the Pilon v1.22 software to obtain a more accurate genome for follow-up analysis.
Prodigal v2.6.3 software was used to predict the coding genes, tRNAscan-SE v2.0 software was used to predict the tRNA in the genome, and IslandPath-DIMOB v0.2 software was used to predict the gene island in the bacterial genome. The Kyoto Encyclopedia of Genes and Genomes (KEGG) database was selected to annotate the entire genome, and the annotation results were obtained. The genomic sequence of strain P1 is preserved in GenBank, login no. PRJNA769798.
2.5. Amino Acid Sequence Homology Analysis
The online software Clustal W (https://www.genome.jp/tools-bin/clustalw, (accessed on 2 October 2022)) was used for multiple sequence alignment, the online software SWISS-MODEL (https://swissmodel.expasy.org/, (accessed on 2 October 2022)) was used to predict the protein secondary structure, and ESPript was used for sequence staining. The phylogenetic tree was constructed by MEGA5.1, and the accession number of the protein sequence can be found in this paper.
2.6. PCR, RT-PCR, and qPCR Amplification of Target Genes
The main genes related to bisphenol A degradation were detected by PCR to verify the existence of BPA degradation enzymes related coding genes in strain P1. Total RNA was extracted by TRIzol Kit (TRIzolTM MaxTM Bacterial RNA Isolation Kit, article number: 16096-020, Life Technologies, Carlsbad, CA, USA). The concentration and purity of the extracted RNA were determined by Nanodrop 2000, RNA integrity was determined by agarose gel electrophoresis, and RIN value was determined by Agilent 2100. After the RNA samples were qualified, rRNA was removed from the total RNA samples. Reagents of Shanghai Shenggong Bioengineering Co. (Shanghai, China) were used for amplification; the specific primers used are shown in Table S1. The PCR methods were as mentioned earlier [40]. In order to analyze the expression of induced genes, each strain was inoculated in MSM with 30 mg/L BPA as the sole carbon source and glucose as the sole carbon source in the control group. After being cultured at 30 °C and 120 rpm for 48 h, reverse transcription and qPCR were performed using our previously described method [41]. In addition, qPCR was used to analyze the expression level of related genes during the degradation of BPA by strain P1, and samples were taken at 48 h. Gene expression was quantified relative to the expression levels of the recA reference gene in each sample using the 2−ΔΔCt quantification method.
3. Results and Discussion
3.1. Screening and Genome Sequencing of BPA-Degrading Bacteria
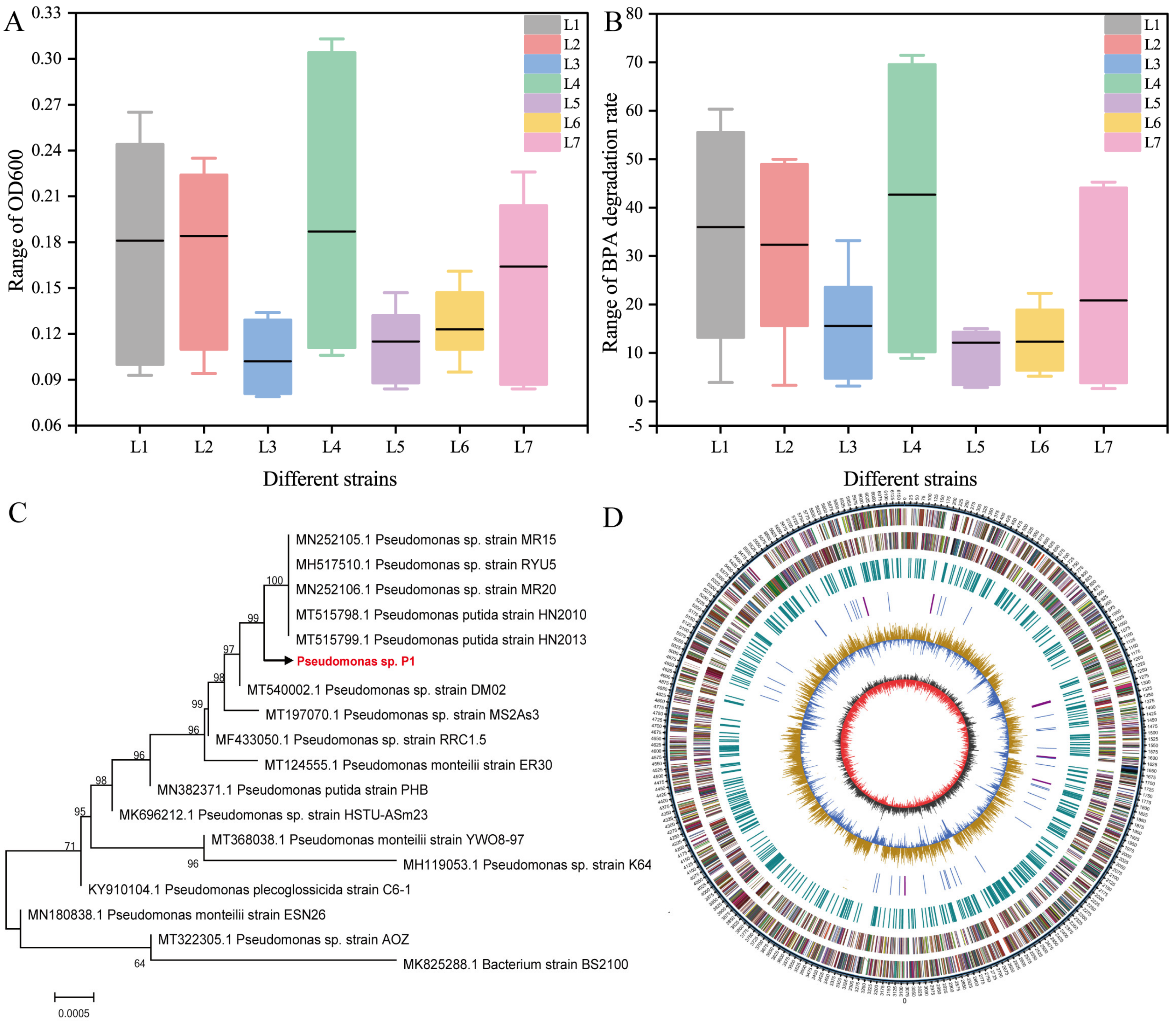
Degradation tests for BPA were carried out on 23 bacterial strains isolated from leachate sediment. Among them, seven strains had BPA degradation activity. The seven bacterial strains were randomly numbered as L1–L7, and strain L4 grew best after aerobic and oscillatory culture for 120 h (Figure 1A). High performance liquid chromatography analysis of the culture supernatant showed that the seven bacteria could degrade BPA to varying degrees; the BPA removal rates ranged from 14.06% to 71.44% after 120 h of culture, of which strain L4 had the best degradation effect (Figure 1B). Therefore, strain L4 was selected as the target strain for further identification and study. An electron micrograph of this strain showed that the cells were short and rod-shaped (Figure S1A). The strain formed round, glossy, light yellow, or white colonies (Figure S1B). The physiological and biochemical characteristics of the strain showed that it was gram-negative (Table S2). Based on the 16S rDNA sequence alignment (Figure 1C), physiological and biochemical detection along with bacterial morphological identification, the strain was identified as Pseudomonas and named Pseudomonas sp. P1. Studies have shown that Pseudomonas is one of the dominant bacteria in BPA degradation [42]. Pseudomonas putida strain YC-AE1 isolated from soil can degrade 80–100% BPA(0.5–12 mg/L) within 15 h; it can also degrade 60-100% BPA (50–900 mg/L) within 72 h [43]. Meanwhile, Pseudomonas sp. LM-1 isolated from activated sludge can also completely degrade BPA (14–24 mg/L) [44]. In addition, Pseudomonas palleroniana GBPI_508 [45] and Pseudomonas aeruginosa PAb1 [46] can grow in the medium containing 10 to 270 mg/L and 5 to 35 mM BPA, respectively. The results of genome sequencing showed that the genome of the strain was composed of a single circular chromosome (Figure 1D). The content of the genome sequence with a total length of 6,172,654 bp, G+C was 62.43%, and a total of 5636 coding genes was annotated. The total length of the coding gene accounted for 87.98% of the total genome length, and the average gene length was 963 bp. The specific genetic information is shown in Table S3.
3.2. Screening of BPA Degradation Genes in Pseudomonas sp. P1
The functional annotation of the gene was obtained by bioinformatics analysis of the sequencing results of the entire genome of strain P1 and compared the gene sequence of the strain with the annotation results of the KEGG database. Based on referring to the annotation results of the KEGG database, we manually reviewed all the annotation results and counted the genes that may be involved in the process of BPA degradation (Table S4). The metabolic pathways of many different types of compounds are involved in strain P1, such as citrate cycle, benzoate, geraniol, xylene, polycyclic aromatic hydrocarbons, and so on. Figure S2 lists some of the metabolic pathways annotated. Therefore, it can be inferred that a large number of unknown genes are involved in BPA degradation in strain P1, and these functional genes need to be mined through further analysis. Meanwhile, this information indicates that the strain P1 exhibits good metabolic pluripotency in BPA degradation with good application potential and complex molecular mechanisms.
In the process of biodegradation of BPA, many types of enzymes play a role in this degradation pathway. Oxidase often plays a very important role in the aerobic degradation of benzene-containing compounds. The Cytochrome P450 (CYP450) monooxygenase system encoded by bisdA and bisdB is a system needed for BPA hydroxylation. These two genes are involved in the first step of hydroxylation during BPA degradation [47]. It has been reported that two key intermediates exist in the BPA degradation products of the CYP450 monooxygenase system, specifically 1,2-bis (4-hydroxyphenyl)-2-propanol and 2,2-bis (4-hydroxyphenyl)-1-propanol. These two key intermediates both converged into 3,4-Dihydroxybenzoate (3,4-DHB) and then went into the tricarboxylic cycle through two divergent pathways via β-Carboxy-cis, cis-muconate (β-CM) or 4-Carboxy-2-hydroxymuconate semialdehyde (4-CHS) [48]. Bacterial CYP450 catalyzes epoxidation, dealkylation sulfoxidation, and hydroxylation of a variety of xenobiotics, including fipronil and dimethyl phthalate, working in coordination with specific electron transport systems [49,50]. Catechol 1,2-dioxygenase (catA), 4,5-DOPA dioxygenase extradiol (LigB), and acetyl-CoA acyltransferase (acetyl-coA) encode aromatic ring-opening dioxygenase, which is related to the aromatic ring-opening of BPA. Some key genes encode enzymes such as monooxygenase (bisdB, bisdA, Lac, and CotA), hydroxylase (3-demethoxyubiquinol 3-hydroxylase (coq7) and phenylalanine-4-hydroxylase (phhA)), and CYP450, which hydroxylate aromatic compounds or hydrocarbon bonds [51,52]. The genes encoded by the above enzymes were annotated in strain P1. Therefore, strain P1 contains genes (bisdB, bisdA, Lac, CotA, catA, and ligB) necessary for the biodegradation of BPA, which are considered to be the key genes for the biodegradation and detoxification of BPA. In order to fully understand the BPA degradation pathway of strain P1 and screen more potential functional genes, we compared the genomic sequences of strain P1 with other BPA-degrading bacteria. Among them, 10 genes have high sequence homology. The specific information is shown in Table 2.
CYP450 is usually involved in the metabolism of a variety of exogenous and endogenous compounds as a terminal oxidase in the multicomponent electron transfer chain. At present, many reports have confirmed that CYP450 plays an important role in the degradation of BPA. In this study, two genes, specifically GE002964 and GE002994, encoding CYP450s were found to have the highest expression during the degradation of BPA. These two CYP450 genes had 27.15% and 25.21% sequence identity with Sphingobium sp. YC-JY1 CYP450 studied by Jia Yang et al., and 27.68% and 24.86% sequence identity with Sphingomonas sp. Strain AO1 CYP450 monooxygenase, which was studied by Sasaki et al. (Figure 2B). The high sequence similarity with the above two known BPA-degrading bacteria indicates that the two CYP450s in strain P1 may have similar mechanisms and play a similar role in the degradation of BPA.
The amino acid sequence of CYP450 in strain P1 was compared with that in strain Sphingobium sp. YC-JY1 (QHW04736.1), Sphingomonas bisphenolicum. AO1 (BAG15884.1), Bacillus sp. GZB (OMQ03826.1), and Bacillus thuringiensis strain GIMCC1.817 (ART94430.1). The results showed that there were 17 α helices and 9 β sheets in the amino acid sequence of CYP450 (Figure 2A). Sequence alignment shows that a heme binding region also exists. All CYP450s contain strictly conserved cysteine residues, which are located at position 334 of the common sequence FGXGXHXCXG region. Heme is anchored to the active site of CYP450 [53]. Two P450 genes in strain P1 may be similarly involved in the biodegradation of BPA. CYP450 can catalyze the initial hydroxylation of BPA methyl and quaternary carbon, resulting in two different products. Two CYP450 genes in strain P1 may be similarly involved in the biodegradation of BPA. Therefore, the enzymes encoded by these genes may show application potential in alleviating BPA pollution.
3.3. Expression of the BPA Degradation Gene in Pseudomonas sp. P1
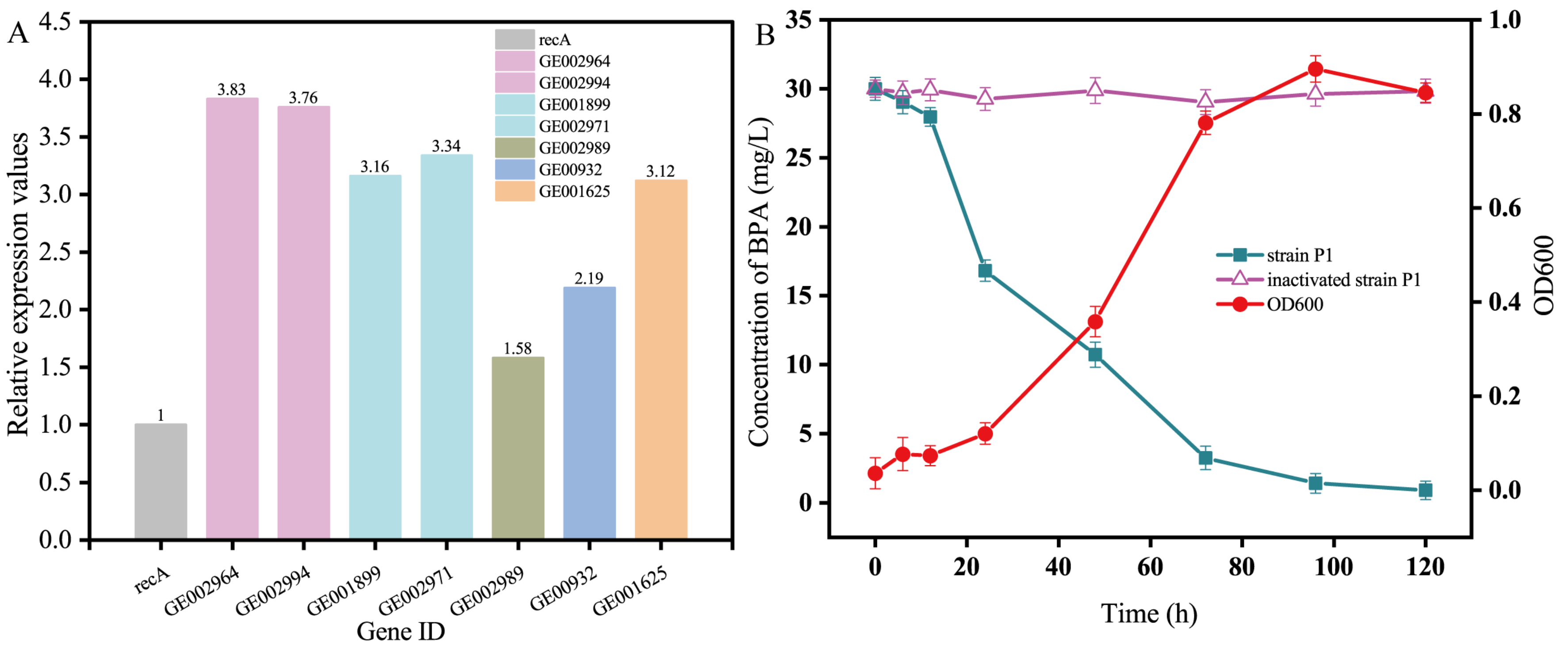
The results of PCR amplification (Figure S3A) proved the existence of the BPA-degrading enzyme-coding gene screened in strain P1. The results of agarose gel electrophoresis of RT-PCR products showed that under culture conditions with BPA as the sole carbon source, genes encoding CYP450, ferredoxin, hydrolase, laccase (Lac), aldo/keto reductase, and copper oxidase (CotA) were expressed (Figure S3B). The RT-PCR results of the control group (glucose culture) showed that 10 BPA-degrading enzyme-coding genes were not transcribed, indicating that the above-mentioned BPA-degrading enzymes were not constitutively expressed, but specifically expressed under the induction of a BPA substrate. The expression of the above genes was determined by fluorescence quantitative PCR using the recA gene as an internal reference gene. The results showed that (Figure 3A) the relative expression of the nine genes was basically consistent with the results of RT-PCR, which proved that the expression of the above genes was true and reliable. From the number and quantity of enzyme expression, we can infer the reason why strain P1 can efficiently degrade BPA. Among those reasons, CYP450 has the highest expression level in qPCR, and many times has been reported as a key enzyme in microbial catabolism of BPA and has its own conserved domain. Therefore, we can conclude that CYP450 is one of the key enzymes in the degradation of BPA by strain P1. The experimental results of qPCR showed that there were not only corresponding BPA degradation genes in the genome of strain P1, but also some genes were highly expressed in the medium with BPA as the sole carbon source. Strain P1 degrades BPA through the enzymes produced by the expression of BPA degradation genes; thus, carrying outgrowth and metabolism.
In order to verify the ability of strain P1 to degrade BPA, this strain was cultured with different temperatures, pH, inoculation amounts of bacterial liquid, and different initial concentrations of BPA. Strain P1 could degrade BPA at 20–40 °C (Figure S4A) with an optimal growth temperature of 30 °C and a degradation rate of 95.1%. The best pH for the growth of strain P1 was pH 7 (Figure S4B). The low membrane permeability of neutral MSM is suitable for maintaining bacterial cell activity, which may enhance BPA degradation [54]. According to Figure S4C, when the dosage of strain P1 was 3%, the degradation rate of BPA peaked. Therefore, a dosage of 3% was selected in the follow-up experiment. Under the conditions of pH 7 and temperature 30 °C, the degradation efficiency of strain P1 at different BPA concentrations was measured (Figure S4D). When the concentration of BPA was 30 mg/L, the highest degradation rate of strain P1 was 95.6%. When the concentration of BPA was from 50 mg/L to 100 mg/L, the degradation ability of strain P1 was significantly inhibited. These findings inferred that BPA is toxic to this strain of bacteria and can inhibit bacterial growth and cell viability. However, the degradation rate of BPA by strain P1 reached 96.89% (Figure 3B). Researchers have isolated and studied several BPA-degrading bacteria (Table S5). Most of the isolated BPA-degrading bacteria showed degradation activity at low concentration, and it took several days to degrade. In the concentration of 30 mg/L, strain P1 used BPA as the only carbon and energy source, and the degradation rate of BPA was more than 96% within 120 h. Obviously, strain P1 is an excellent BPA degrader, which can effectively degrade high concentration of BPA. To sum up, strain P1 has good degradation ability in a variety of different environmental conditions, and can tolerate a wide range of culture temperatures, pH, and BPA initial concentrations. At the same time, strain P1 can effectively degrade BPA with BPA as the only carbon source.
Although many types of bacteria can degrade BPA, only a few have a strong ability to biodegrade BPA [1]. Under optimal conditions, the ability of strain P1 to degrade BPA was tested further. In the control groups that were sterilized, nearly no BPA loss was observed (Figure 3B). Thus, the main removal route of BPA was biodegradation. Strain P1 degraded almost all BPA within 120 h, and the biomass of the strain increased with an increase in the degradation rate (Figure 3B). The degradation rate and biomass of strain P1 increased slowly from 0 to 24 h, which was the adaptive stage. After 24 h of culture, strain P1 entered a logarithmic growth period (24–72 h). At this time, the growth and metabolism of strain P1 require the bacteria to consume a considerable amount of nutrients while the substrate concentration gradually decreased, and the degradation rate of BPA reached 89.2%. During this period, the biomass increased rapidly, and the culture gradually changed from clear to turbid with yellowing. During the period of 72–120 h, the degradation rate of BPA reached 96.89%. Pseudomonas is a common strain in activated sludge, which has a good function of nitrogen and phosphorus removal. Therefore, Pseudomonas is widely used as a degrading bacteria and is highly recognized [55]. It can be judged from the above results that strain P1 can effectively make use of the degradation of BPA to meet the needs of its growth and metabolism; the final degradation rate exceeded 90%.
3.4. The Defense Mechanism of Pseudomonas sp. P1 against BPA
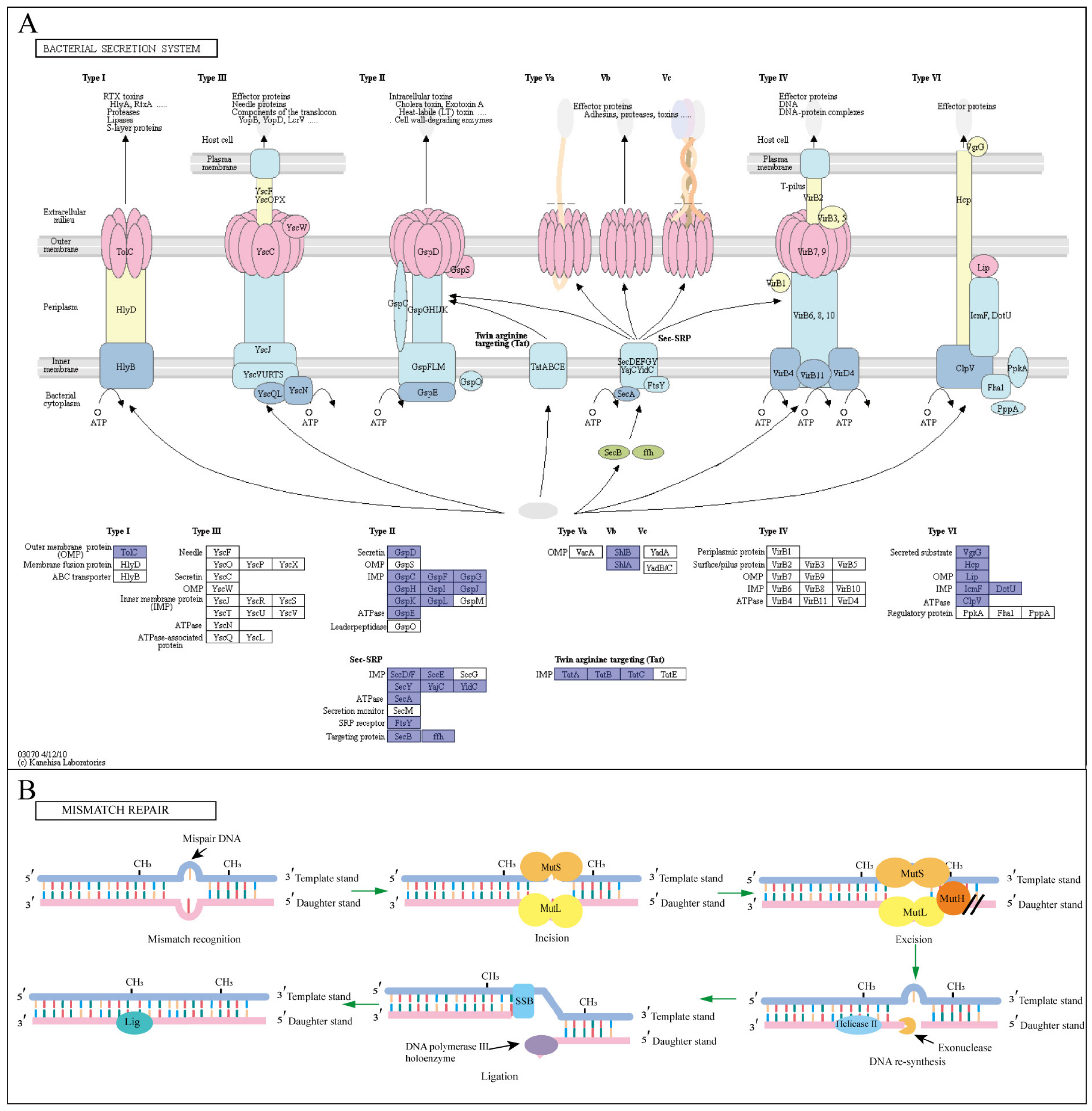
The metabolites secreted by bacteria and the oxidative stress response of bacteria to BPA are important indicators of the bacterial response to BPA degradation. The different secretion systems of bacteria require corresponding gene expression proteins (signal, transmembrane, and secretory proteins) to perform the corresponding transport functions [56]. The secretory pathway of strain P1 showed that this strain had a large proportion of Sec-SRP, Tat, type II, and Type VI pathway genes (Figure 4A; Table S6). Strain P1 is a gram-negative bacterial species with inner and outer membrane structure and intermediate periplasmic space. The Sec-SRP pathway mainly transports substances to the periplasm and extracellular areas. The Type II pathway is mainly used to secrete various hydrolases and toxins. The VI pathway transports secreted substances to the outer membrane of bacteria [57]. The Tat pathway of strain P1 transports secretions to the periplasmic space. The Tat system consists of four membrane proteins: TatA, TatB, TatC, and TatE. The first three proteins are encoded by a manipulator (TatABC). The TatE protein is an inactivated form of TatA and may have originated from a gene doubling event of TatA. It is currently believed that TatBC first recognizes the signal sequence of the secreted protein, and then TatA binds to TatBC and transports the protein using proton movement force (PMF) for energy supply. When the transit is complete, the signal sequence is excised by the lead peptidase and the protein is released into the periplasm for the next transit step [58]. Substrates bound to the Tat secretion pathway are often involved in redox reactions [59]. It was also shown that when BPA was in contact with bacteria, BPA exerted oxidative stress on bacteria, and bacteria mainly secreted antioxidants and oxidoreductases to degrade BPA through the Tat pathway, while the Sec-SRP and Type VI pathways mainly secreted extracellular functional proteins for BPA degradation [60]. Therefore, the above analysis shows that these four secretory systems play an important role in the resistance of metabolites secreted by strain P1 to BPA toxicity.
Several studies have shown that BPA exposure is related to oxidative damage to DNA [61,62]. Mismatch repair plays a unique and significant regulatory role in bacteria, helping them to maintain genomic stability. In the process of repair, MutS homodimers recognize mismatched bases on the sub-chain and bind to mutant DNA (Figure 4B). In addition, MutH binds to the semi-methylation site of the sub-chain DNA and becomes active only when it comes into contact with the MutL homodimer, which binds to the MutS-DNA complex and acts as a medium between MutS and MutH, activating the latter protein [63]. The key active components of mismatch repair systems include MutL, MutH, and MutS, which are very important for detecting mismatches and guiding the repair machine. At the same time, studies have shown that CYP450 is the key gene that affects mismatch repair so as to protect cells from BPA-induced toxicity [64]. Functional annotation of the mismatch repair system gene of strain P1 showed that this strain had a complete mismatch repair system as well as related pathway genes (Table S6).
Strain P1 has a complete antioxidant enzyme secretion pathway and a complete mismatch repair system gene (Table S6). It can be inferred that strain P1 has good resistance to BPA toxicity, which is a key step for the bacteria to further metabolize BPA.
![Water 15 00830 g004]()
Figure 4.
Graphic illustration of the defense mechanism of the strain P1 (A) bacterial secretory system [65]. The two-step secretory pathway includes Sec and Tat pathways, and secretions can pass through the plasma membrane. The Type II pathway is mainly used to secrete various hydrolases and toxins, and the one-step secretion pathway includes a Type VI pathway, which is mainly responsible for transporting secreted substances to the outer membrane of bacteria; (B) mismatch repair system, the proteins involved in the figure are annotated in the genome of strain P1.
Figure 4.
Graphic illustration of the defense mechanism of the strain P1 (A) bacterial secretory system [65]. The two-step secretory pathway includes Sec and Tat pathways, and secretions can pass through the plasma membrane. The Type II pathway is mainly used to secrete various hydrolases and toxins, and the one-step secretion pathway includes a Type VI pathway, which is mainly responsible for transporting secreted substances to the outer membrane of bacteria; (B) mismatch repair system, the proteins involved in the figure are annotated in the genome of strain P1.

3.5. Pathway of BPA Degradation by Pseudomonas sp. P1
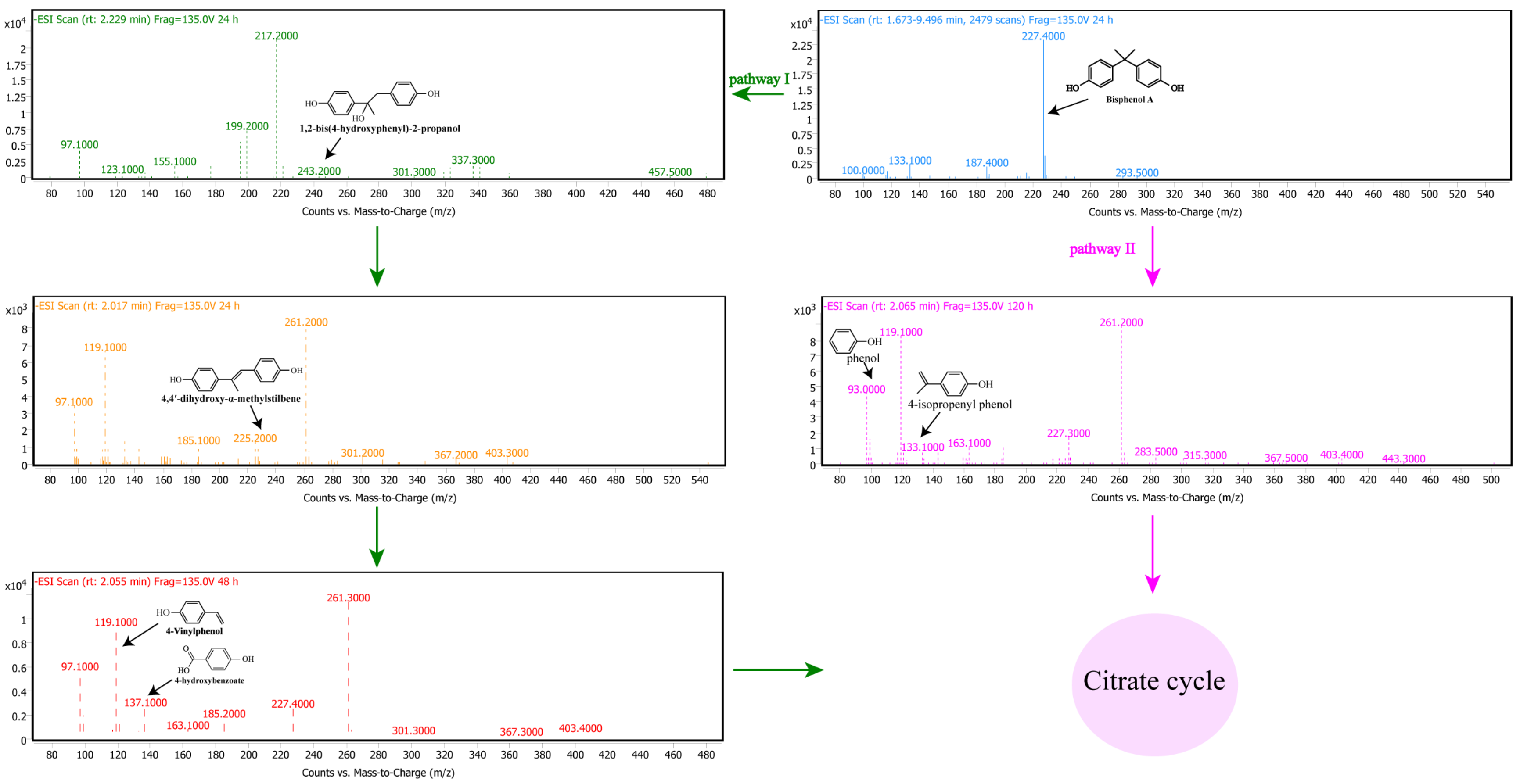
Based on the above results and analysis, Pseudomonas sp. P1 can effectively degrade BPA. Based on the functional annotation of the genome, it is hypothesized that strain P1 first secretes antioxidants, such as SOD/CAT/GSH, through the secretory pathway to maintain its survival. Simultaneous mismatch repair also occurs to protect cells from BPA-induced toxicity. The P1 genome of strain P1 includes a large number of protein-coding genes, which play an important role in predicting the BPA degradation pathway, including a carbohydrate active enzyme (cazyme), short chain dehydrogenase, oxidase, and hydrolase. Then, according to the expression results of BPA degradation genes in strain P1 and the detection of BPA degradation intermediates in strain P1 (Table S7), the inferred BPA degradation pathway is shown in Figure 5.
In pathway I, through the action of the enzyme complex CYP450, ferredoxin, and ferredoxin reductase on the quaternary carbon atom of BPA, the BPA undergoes hydroxylation to produce 1,2-bis (4-hydroxyphenyl)-2-propanol. It is a main intermediate product in the process of BPA degradation [37]; almost the whole process of BPA degradation was detected. It is further dehydrated to produce 4,4’-dihydroxy-alpha-methylstilbene. The oxidative cleavage of 4,4’-dihydroxy-alpha-methylstilbene by copper oxidase produced two intermediates, 4-hydroxybenzoate and 4-Vinylphenol. 4-Vinylphenol is detected for the first time during BPA biodegradation.
In pathway II, BPA is directly converted to phenol and 4-isoproenyl phenol. A previous study found that laccase in Trametes villosa produces phenol and 4-isopropylphenol during BPA degradation [66]. When immobilized laccase degrades BPA on silica nanoparticles, three main intermediates, phenol, 4-isopropyl phenol, and 4-hydroxyisopropyl phenol, can be produced [67]. It had been hypothesized that laccase or ligninolytic enzymes could aid in the polymerization of BPA, followed by formation of dimers and oligomers through coupling reactions between C-C or C-O groups of the phenolic residues, as well as further cleavage on the oligomers-produced phenol, and 4-isopropenylphenol [68]. Based on the detected intermediates and the laccase gene in the genome, it can be inferred that the BPA degradation of strain P1 may be completed through pathway II.
4. Conclusions
In this study, a strain of Pseudomonas sp. P1, which can effectively degrade BPA, was obtained. The highest degradation rate was 96.89% when the temperature was 30 °C, the pH value was 7, the content of BPA was 30 mg/L, and the amount of inoculation was 3 mL. The genome of strain P1 contains 5636 genes that encode proteins. Through comparative genomic analysis, all the essential genes for BPA biodegradation in strain P1 were screened, including 72 functional genes involved in the mechanism of BPA stress and 138 functional genes that may be involved in BPA degradation. Seven genes such as cytochrome P450, ferredoxin, ferredoxin reductase complex, and laccase were expressed under BPA induction. Strain P1 has strong environmental adaptability to an environmental temperature tolerance range of 25–40 °C, pH value of 5–8, and BPA concentration of 15–100 mg/L. Six intermediates were detected in the process of BPA degradation, among which 4-vinylphenol was found for the first time in the process of BPA degradation. On this basis, the biodegradation pathway was proposed. The results show that strain P1 has rich functional genes and good environmental adaptability and can be used for bioremediation of all kinds of BPA-contaminated environment.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15040830/s1, Table S1: PCR specific primer sequence; Table S2: Physiological and biochemical characteristics of strain P1; Table S3: Genomic characteristics of Pseudomonas sp. P1; Table S4: Description of genes obtained from genome of P1 relevant to BPA degradation; Table S5: Comparison of BPA degradation rates by various microorganisms; Table S6: Information about functional genes resistant to BPA toxicity; Table S7: LC-MS data of BPA degradation intermediates; Figure S1: Scanning electron microscope (A) and morphological observation (B) of strain P1; Figure S2: The complete TCA cycle (A) and Benzoate degradation (B) were present in the genome of strain P1. The data are based on RAST analysis. Enzyme committee numbers were obtained from the KEGG database; Figure S3: (A) PCR results of BPA degradation-related genes. (B) RT-PCR agarose gel electrophoresis of BPA degradation-related genes under different conditions; Figure S4: Degradation of BPA by strain P1 (A) different temperature, (B) different pH, (C) different inoculum size, (D) different initial concentration.
Author Contributions
Conceptualization, S.L. and K.T.; methodology, Q.Q.; software, Y.Y. and H.L.; validation, M.C. and X.S.; investigation, J.G., F.Z., Y.W.; writing—original draft preparation, investigation, validation, writing—review and editing, visualization, S.L.; supervision, project administration, funding acquisition, H.H. All authors have read and agreed to the published version of the manuscript.
Funding
This study was funded by National Natural Science Foundation of China (No. 51978132).
Data Availability Statement
Data will be made available on request.
Acknowledgments
We are grateful to Lu Nan from the Experiment Center of the College of Environment of Northeast Normal University for her help.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Michalowicz, J. Bisphenol A—Sources, toxicity and biotransformation. Environ. Toxicol. Phar. 2014, 37, 738–758. [Google Scholar] [CrossRef] [PubMed]
- Cantonwine, D.E.; Hauser, R.; Meeker, J.D. Bisphenol A and Human Reproductive Health. Obstet. Gynecol. 2013, 8, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Wazir, U.; Mokbel, K. Bisphenol A: A Concise Review of Literature and a Discussion of Health and Regulatory Implications. In Vivo 2019, 33, 1421–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleme, J.J.; Fan, Y.V.; Tan, R.R.; Jiang, P. Minimising the present and future plastic waste, energy and environmental footprints related to COVID-19. Renew. Sust. Energ. Rev. 2020, 127, 109883. [Google Scholar] [CrossRef]
- Kalina, M.; Tilley, E. “This is our next problem”: Cleaning up from the COVID-19 response. Waste Manag. 2020, 108, 202–205. [Google Scholar] [CrossRef]
- Haldar, S.; Muralidaran, Y.; Miguez, D.; Mulla, S.I.; Mishra, P. Eco-toxicity of nano-plastics and its implication on human metabolism: Current and future perspective. Sci. Total Environ. 2023, 861, 160571. [Google Scholar] [CrossRef]
- Arnold, S.M.; Clark, K.E.; Staples, C.A.; Klecka, G.M.; Dimond, S.S.; Caspers, N.; Hentges, S.G. Relevance of drinking water as a source of human exposure to bisphenol A. J. Expo. Sci. Env. Epidemiol. 2013, 23, 137–144. [Google Scholar] [CrossRef]
- Olatunji, O.S.; Fatoki, O.S.; Opeolu, B.O.; Ximba, B.J.; Chitongo, R. Determination of selected steroid hormones in some surface water around animal farms in Cape Town using HPLC-DAD. Environ. Monit. Assess. 2017, 189, 363. [Google Scholar] [CrossRef]
- Wanda, E.M.M.; Mamba, B.B.; Msagati, T.A.M. Hydrochemical modelling of water quality in terms of emerging micropollutants in Mpumalanga, Gauteng and North West Provinces. Phys. Chem. Earth 2017, 100, 143–157. [Google Scholar] [CrossRef]
- Wanda, E.M.M.; Nyoni, H.; Mamba, B.B.; Msagati, T.A.M. Occurrence of Emerging Micropollutants in Water Systems in Gauteng, Mpumalanga, and North West Provinces, South Africa. Int. J. Env. Pub. Health 2017, 14, 79. [Google Scholar] [CrossRef] [Green Version]
- Maduka Ignatius, C.; Ezeonu Francis, E.; Neboh Emeka, E.; Shu Elvis, N.; Ikekpeazu Ebele, J. BPA and Environmental Estrogen in Potable Water Sources in Enugu Municipality, South-East, Nigeria. Bull. Environ. Contam. Toxicol. 2010, 85, 534–537. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhao, J.L.; Zhang, C.Y.; Rao, W.L.; Liang, G.H.; Zhang, H.; Liu, Y.H.; Guan, Y.F.; Zhang, H.Y.; Ying, G.G. Profile and removal of bisphenol analogues in hospital wastewater, landfill leachate, and municipal wastewater in South China. Sci. Total Environ. 2021, 790, 148269. [Google Scholar] [CrossRef] [PubMed]
- Andujar, N.; Galvez-Ontiveros, Y.; Zafra-Gomez, A.; Rodrigo, L.; Jesus Alvarez-Cubero, M.; Aguilera, M.; Monteagudo, C.; Rivas, A. Bisphenol A Analogues in Food and Their Hormonal and Obesogenic Effects: A Review. Nutrients 2019, 11, 2136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narevski, A.C.; Novakovic, M.I.; Petrovic, M.Z.; Mihajlovic, I.J.; Maodus, N.B.; Vujic, G.V. Occurrence of bisphenol A and microplastics in landfill leachate: Lessons from South East Europe. Environ. Sci. Pollut. Res. 2021, 28, 42196–42203. [Google Scholar] [CrossRef] [PubMed]
- Greca, S.-C.d.A.; Kyrou, I.; Pink, R.; Randeva, H.; Grammatopoulos, D.; Silva, E.; Karteris, E. Involvement of the Endocrine-Disrupting Chemical Bisphenol A (BPA) in Human Placentation. J. Clin. Med. 2020, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Ejaredar, M.; Lee, Y.; Roberts, D.J.; Sauve, R.; Dewey, D. Bisphenol A exposure and children’s behavior: A systematic review. J. Expo. Sci. Environ. Epidemiol. 2017, 27, 175–183. [Google Scholar] [CrossRef]
- Ashfaq, M.; Khan, K.N.; Rehman, M.S.U.; Mustafa, G.; Nazar, M.F.; Sun, Q.; Iqbal, J.; Mulla, S.I.; Yu, C.P. Ecological risk assessment of pharmaceuticals in the receiving environment of pharmaceutical wastewater in Pakistan. Ecotoxicol. Environ. Saf. 2017, 136, 31–39. [Google Scholar] [CrossRef]
- Mokra, K.; Wozniak, K.; Bukowska, B.; Sicinska, P.; Michalowicz, J. Low-concentration exposure to BPA, BPF and BPAF induces oxidative DNA bases lesions in human peripheral blood mononuclear cells. Chemosphere 2018, 201, 119–126. [Google Scholar] [CrossRef]
- Naveira, C.; Rodrigues, N.; Santos, F.S.; Santos, L.N.; Neves, R.A.F. Acute toxicity of Bisphenol A (BPA) to tropical marine and estuarine species from different trophic groups. Environ. Pollut. 2021, 268, 115911. [Google Scholar] [CrossRef]
- Leech, D.M.; Snyder, M.T.; Wetzel, R.G. Natural organic matter and sunlight accelerate the degradation of 17 beta-estradiol in water. Sci. Total Environ. 2009, 407, 2087–2092. [Google Scholar] [CrossRef]
- Sun, Z.; Zhao, L.; Liu, C.; Zhen, Y.; Ma, J. Fast adsorption of BPA with high capacity based on pi-pi electron donoracceptor and hydrophobicity mechanism using an in-situ sp(2) C dominant N-doped carbon. Chem. Eng. J. 2020, 381, 122510. [Google Scholar] [CrossRef]
- Hou, Z.; Wen, Z.; Wang, D.; Wang, J.; Francois-Xavier, C.P.; Wintgens, T. Bipolar jet electrospinning bi-functional nanofibrous membrane for simultaneous and sequential filtration of Cd2+ and BPA from water: Competition and synergistic effect. Chem. Eng. J. 2018, 332, 118–130. [Google Scholar] [CrossRef]
- Ma, R.; Zhang, S.; Liu, X.; Sun, M.; Cao, J.; Wang, J.; Wang, S.; Wen, T.; Wang, X. Oxygen defects-induced charge transfer in Bi7O9I3 for enhancing oxygen activation and visible-light degradation of BPA. Chemosphere 2022, 286, 131783. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.; Franke, M.; Stelter, M.; Braeutigam, P. Degradation of endocrine disruptor bisphenol A by ultrasound-assisted electrochemical oxidation in water. Ultrason. Sonochemistry 2017, 39, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Bharagava, R.N.; Kishor, R.; Purchase, D.; Ferreira, L.; Bilal, M. Environmental and Health Hazards of Textile Industry Wastewater Pollutants and Its Treatment Approaches; Springer: Cham, Switzerland, 2021; pp. 1–24. [Google Scholar] [CrossRef]
- Peng, Y.-H.; Chen, Y.J.; Chang, Y.J.; Shih, Y.H. Biodegradation of bisphenol A with diverse microorganisms from river sediment. J. Hazard. Mater. 2015, 286, 285–290. [Google Scholar] [CrossRef]
- Ben Ouada, S.; Ben Ali, R.; Leboulanger, C.; Ben Ouada, H.; Sayadi, S. Effect of Bisphenol A on the extremophilic microalgal strain Picocystis sp. (Chlorophyta) and its high BPA removal ability. Ecotoxicol. Environ. Saf. 2018, 158, 1–8. [Google Scholar] [CrossRef]
- Li, G.; Xiong, J.; Wong, P.K.; An, T. Enhancing tetrabromobisphenol A biodegradation in river sediment microcosms and understanding the corresponding microbial community. Environ. Pollut. 2016, 208, 796–802. [Google Scholar] [CrossRef]
- Das, R.; Li, G.; Mai, B.; An, T. Spore cells from BPA degrading bacteria Bacillus sp. GZB displaying high laccase activity and stability for BPA degradation. Sci. Total Environ. 2018, 640, 798–806. [Google Scholar] [CrossRef]
- Sakai, K.; Yamanaka, H.; Moriyoshi, K.; Ohmoto, T.; Ohe, T. Biodegradation of bisphenol A and related compounds by Sphingomonas sp. strain BP-7 isolated from seawater. Biosci. Biotech. Biochem. 2007, 71, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Cydzik-Kwiatkowska, A.; Zielinska, M.; Bernat, K.; Bulkowska, K.; Wojnowska-Baryla, I. Insights into mechanisms of bisphenol A biodegradation in aerobic granular sludge. Bioresour. Technol. 2020, 315, 123806. [Google Scholar] [CrossRef]
- Yu, Y.; Yin, H.; Peng, H.; Lu, G.; Dang, Z. Proteomic mechanism of decabromodiphenyl ether (BDE-209) biodegradation by Microbacterium Y2 and its potential in remediation of BDE-209 contaminated water-sediment system. J. Hazard. Mater. 2020, 387, 121708. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Wang, Y.; Mao, Y.; Zhang, T. Partnership of Arthrobacter and Pimelobacter in Aerobic Degradation of Sulfadiazine Revealed by Metagenomics Analysis and Isolation. Environ. Sci. Technol. 2018, 52, 2963–2972. [Google Scholar] [CrossRef] [PubMed]
- Yue, W.; Yin, C.F.; Sun, L.; Zhang, J.; Xu, Y.; Zhou, N.Y. Biodegradation of bisphenol-A polycarbonate plastic by Pseudoxanthomonas sp. strain NyZ600. J. Hazard. Mater. 2021, 416, 125775. [Google Scholar] [CrossRef]
- Kyrila, G.; Katsoulas, A.; Schoretsaniti, V.; Rigopoulos, A.; Rizou, E.; Doulgeridou, S.; Sarli, V.; Samanidou, V.; Touraki, M. Bisphenol A removal and degradation pathways in microorganisms with probiotic properties. J. Hazard. Mater. 2021, 413, 125363. [Google Scholar] [CrossRef] [PubMed]
- Shobnam, N.; Sun, Y.; Mahmood, M.; Loffler, F.E.; Im, J. Biologically mediated abiotic degradation (BMAD) of bisphenol A by manganese-oxidizing bacteria. J. Hazard. Mater. 2021, 417, 125987. [Google Scholar] [CrossRef]
- Zhang, W.; Yin, K.; Chen, L. Bacteria-mediated bisphenol A degradation. Appl. Microbiol. Biotechnol. 2013, 97, 5681–5689. [Google Scholar] [CrossRef]
- Qiu, Q.; Wang, P.; Kang, H.; Wang, Y.; Tian, K.; Huo, H. Genomic Analysis of a New Estrogen-Degrading Bacterial Strain, Acinetobacter sp. DSSKY-A-001. Int. J. Genomics. 2019, 2019, 2804134. [Google Scholar] [CrossRef] [Green Version]
- Tian, K.; Yu, Y.; Qiu, Q.; Sun, X.; Meng, F.; Bi, Y.; Gu, J.; Wang, Y.; Zhang, F.; Huo, H. Mechanisms of BPA Degradation and Toxicity Resistance in Rhodococcus equi. Microorganisms 2023, 11, 67. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Y.; Xu, X.; Tian, K.; Zhou, D.; Meng, F.; Zhang, H.; Huo, H. Genomics analysis of the steroid estrogen-degrading bacterium Serratia nematodiphila DH-S01. Biotechnol. Biotechnol. Equip. 2020, 34, 430–440. [Google Scholar] [CrossRef]
- Tian, K.; Meng, F.; Meng, Q.; Gao, Y.; Zhang, L.; Wang, L.; Wang, Y.; Li, X.; Huo, H. The Analysis of Estrogen-Degrading and Functional Metabolism Genes inRhodococcus equi DSSKP-R-001. Int. J. Genom. 2020, 2020, 9369182. [Google Scholar] [CrossRef]
- De Morais Farias, J.; Krepsky, N. Data for: Bacterial degradation of bisphenol analogues: An overview. Environ. Sci. Pollut. 2022, 29, 76543–76564. [Google Scholar] [CrossRef] [PubMed]
- Eltoukhy, A.; Jia, Y.; Nahurira, R.; Abo-Kadoum, M.A.; Khokhar, I.; Wang, J.; Yan, Y. Biodegradation of endocrine disruptor Bisphenol A by Pseudomonas putida strain YC-AE1 isolated from polluted soil, Guangdong, China. BMC Microbiol. 2020, 20, 11. [Google Scholar] [CrossRef] [Green Version]
- Gu, C.; Liang, J.; Liu, M.; Rui, J.; Shi, J.; Yu, Y.; Zhang, X. Aerobic degradation of bisphenol A by Pseudomonas sp. LM-1: Characteristic and pathway. Biodegradation 2023, 34, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Thathola, P.; Agnihotri, V.; Pandey, A.; Upadhyay, S.K. Biodegradation of bisphenol A using psychrotolerant bacterial strain Pseudomonas palleroniana GBPI_508. Arch. Microbiol. 2022, 204, 272. [Google Scholar] [CrossRef]
- Vijayalakshmi, V.; Senthilkumar, P.; Mophin-Kani, K.; Sivamani, S.; Sivarajasekar, N.; Vasantharaj, S. Bio-degradation of Bisphenol A by Pseudomonas aeruginosa PAb1 isolated from effluent of thermal paper industry: Kinetic modeling and process optimization. J. Radiat. Res. Appl. Sci. 2018, 11, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Akahira, A.; Oshiman, K.I.; Tsuchido, T.; Matsumura, Y. Purification of cytochrome P450 and ferredoxin, involved in bisphenol A degradation, from Sphingomonas sp. strain AO1. Appl. Environ. Microb. 2005, 71, 8024–8030. [Google Scholar] [CrossRef] [Green Version]
- Tong, T.; Li, R.; Chai, M.; Wang, Q.; Yang, Y.; Xie, S. Metagenomic analysis of microbial communities continuously exposed to Bisphenol A in mangrove rhizosphere and non-rhizosphere soils. Sci. Total Environ. 2021, 792, 148486. [Google Scholar] [CrossRef]
- Mandal, K.; Singh, B.; Jariyal, M.; Gupta, V.K. Microbial degradation of fipronil by Bacillus thuringiensis. Ecotoxicol. Environ. Saf. 2013, 93, 87–92. [Google Scholar] [CrossRef]
- Surhio, M.A.; Talpur, F.N.; Nizamani, S.M.; Amin, F.; Bong, C.W.; Lee, C.W.; Ashraf, M.A.; Shah, M.R. Complete degradation of dimethyl phthalate by biochemical cooperation of the Bacillus thuringiensis strain isolated from cotton field soil. Rsc. Adv. 2014, 4, 55960–55966. [Google Scholar] [CrossRef]
- Bathelt, C.M.; Ridder, L.; Mulholland, A.J.; Harvey, J.N. Aromatic hydroxylation by cytochrome P450: Model calculations of mechanism and substituent effects. J. Am. Chem. Soc. 2003, 125, 15004–15005. [Google Scholar] [CrossRef] [PubMed]
- Rittle, J.; Green, M.T. Cytochrome P450 Compound I: Capture, Characterization, and C-H Bond Activation Kinetics. Science 2010, 330, 933–937. [Google Scholar] [CrossRef] [Green Version]
- Degtyarenko, K.N. Structural domains of P450-containing monooxygenase systems. Protein Eng. Des. Sel. 1995, 8, 737–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, J.; Zhao, H.; Yin, H.; Peng, H.; Tang, L.; Gao, J.; Ma, Y. Triphenyltin biodegradation and intracellular material release by Brevibacillus brevis. Chemosphere 2014, 105, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Bi, X.; Peng, Y.; Bai, M. Research advances of the phosphorus-accumulating organisms of Candidatus Accumulibacter, Dechloromonas and Tetrasphaera: Metabolic mechanisms, applications and influencing factors. Chemosphere 2022, 307, 135675. [Google Scholar] [CrossRef]
- Hui, X.; Chen, Z.; Zhang, J.; Lu, M.; Cai, X.; Deng, Y.; Hu, Y.; Wang, Y. Computational prediction of secreted proteins in gram-negative bacteria. Comput. Struct. Biotechnol. J. 2021, 19, 1806–1828. [Google Scholar] [CrossRef] [PubMed]
- Kudva, R.; Denks, K.; Kuhn, P.; Vogt, A.; Mueller, M.; Koch, H.G. Protein translocation across the inner membrane of Gram-negative bacteria: The Sec and Tat dependent protein transport pathways. Res. Microbiol. 2013, 164, 505–534. [Google Scholar] [CrossRef] [PubMed]
- Palmer, T.; Sargent, F.; Berks, B.C. The Tat Protein Export Pathway. EcoSal Plus 2010, 4. [Google Scholar] [CrossRef]
- Prajapati, B.; Bernal-Cabas, M.; Lopez-Alvarez, M.; Schaffer, M.; Bartel, J.; Rath, H.; Steil, L.; Becher, D.; Volker, U.; Mader, U.; et al. Double trouble: Bacillus depends on a functional Tat machinery to avoid severe oxidative stress and starvation upon entry into a NaCl-depleted environment. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118914. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Xie, Y.; Hou, J.; Zhou, X.; Chen, J.; Yao, C.; Zhang, Y.; Li, Y. Biodegradation of bisphenol A by alginate immobilized Phanerochaete chrysosporium beads: Continuous cyclic treatment and degradation pathway analysis. Biochem. Eng. J. 2022, 177, 108212. [Google Scholar] [CrossRef]
- Watkins, D.J.; Ferguson, K.K.; Del Toro, L.V.A.; Alshawabkeh, A.N.; Cordero, J.F.; Meeker, J.D. Associations between urinary phenol and paraben concentrations and markers of oxidative stress and inflammation among pregnant women in Puerto Rico. Int. J. Hyg. Environ. Health 2015, 218, 212–219. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.; Lu, S.; Dai, Y.; Rui, C.; Wang, Y.; Zhou, Y.; Li, Y.; Pang, Q.; Fan, R. Higher dermal exposure of cashiers to BPA and its association with DNA oxidative damage. Environ. Int. 2017, 98, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, P. Molecular mechanisms of DNA mismatch repair. DNA Repair. 2001, 486, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yu, H.; Qin, H.; Long, Y.; Ye, J.; Qu, Y. Bisphenol A degradation pathway and associated metabolic networks in Escherichia coli harboring the gene encoding CYP450. J. Hazard. Mater. 2020, 388, 121737. [Google Scholar] [CrossRef] [PubMed]
- Hou, S.; Yang, P. BPA biodegradation driven by isolated strain SQ-2 and its metabolism mechanism elucidation. Biochem. Eng. J. 2022, 185, 108540. [Google Scholar] [CrossRef]
- Fukuda, T.; Uchida, H.; Takashima, Y.; Uwajima, T.; Kawabata, T.; Suzuki, M. Degradation of bisphenol a by purified laccase from Trametes villosa. Biochem. Biophys. Res. Commun. 2001, 284, 704–706. [Google Scholar] [CrossRef]
- Galliker, P.; Hommes, G.; Schlosser, D.; Corvini, P.F.X.; Shahgaldian, P. Laccase-modified silica nanoparticles efficiently catalyze the transformation of phenolic compounds. J. Colloid Interface Sci. 2010, 349, 98–105. [Google Scholar] [CrossRef]
- Huang, Q.G.; Weber, W.J. Transformation and removal of bisphenol A from aqueous phase via peroxidase-mediated oxidative coupling reactions: Efficacy, products, and pathways. Environ. Sci. Technol. 2005, 39, 6029–6036. [Google Scholar] [CrossRef]
Figure 1.
Screening and identification of bisphenol A-(BPA)-degrading bacteria. For seven bacterial strains (L1–L7): (A) Bacterial growth in MSM medium containing 100 mg/L BPA, and (B) BPA degradation rates; different strains are expressed in different colors in the figure and corresponding to the abscissa. (C) 16S rDNA sequence alignment used for morphological identification of bacterial strains; and (D) results of genome sequencing showing that the genome of strain Pseudomonas sp. P1 was composed of a singular circular chromosome. Different colors represent the distribution of different functional genes.
Figure 1.
Screening and identification of bisphenol A-(BPA)-degrading bacteria. For seven bacterial strains (L1–L7): (A) Bacterial growth in MSM medium containing 100 mg/L BPA, and (B) BPA degradation rates; different strains are expressed in different colors in the figure and corresponding to the abscissa. (C) 16S rDNA sequence alignment used for morphological identification of bacterial strains; and (D) results of genome sequencing showing that the genome of strain Pseudomonas sp. P1 was composed of a singular circular chromosome. Different colors represent the distribution of different functional genes.

Figure 2.
(A) Multiple sequence alignments for the Cytochrome P450 (GE002964, GE002994) compared to other homologs. (B) Phylogenetic reconstruction of Cytochrome P450 (CYP450) from strain P1 and other CYP450s.
Figure 2.
(A) Multiple sequence alignments for the Cytochrome P450 (GE002964, GE002994) compared to other homologs. (B) Phylogenetic reconstruction of Cytochrome P450 (CYP450) from strain P1 and other CYP450s.

Figure 3.
(A) Expression levels of genes encoding BPA-degrading enzymes: (GE002964, GE002994) Cytochrome P450; (GE001899, GE002971) Ferredoxin; (GE002989) Hydrolase; (GE000932) Copper oxidase; (GE001625) Laccase; (B) degradation capacity of strain P1 under optimal conditions; OD600 value of the bacterial solution.
Figure 3.
(A) Expression levels of genes encoding BPA-degrading enzymes: (GE002964, GE002994) Cytochrome P450; (GE001899, GE002971) Ferredoxin; (GE002989) Hydrolase; (GE000932) Copper oxidase; (GE001625) Laccase; (B) degradation capacity of strain P1 under optimal conditions; OD600 value of the bacterial solution.

Figure 5.
Two pathways for Pseudomonas sp. P1 to degrade BPA. The green arrows represent BPA conversion through pathway I, and the pink arrows represent BPA conversion through pathway II. Finally, both pathways enter the tricarboxylic acid cycle for complete metabolism.
Figure 5.
Two pathways for Pseudomonas sp. P1 to degrade BPA. The green arrows represent BPA conversion through pathway I, and the pink arrows represent BPA conversion through pathway II. Finally, both pathways enter the tricarboxylic acid cycle for complete metabolism.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Conditions of single-factor analysis.
| Culture Temperature (°C) | Initial pH | Inoculum Size (%, v/v) | Initial Concentration of BPA (mg/L) |
|---|---|---|---|
| 20, 25, 30, 35, 40 | 5, 6, 7, 8 | 1, 3, 5, 7, 9 | 15, 30, 50, 70, 100 |
Table 2.
Coding genes of key enzymes involved in Bisphenol A degradation in Pseudomonas sp. P1.
| Gene ID | Gene Name | Enzyme | Expression Level | Homologous Protein (Sequence Identity) |
|---|---|---|---|---|
| GE000741 | P-hydroxybenzoate hydroxylase | - | Hydroxylase a (39.39%) | |
| GE003776 | CotA | Copper oxidase | - | CotAb (24.32%) |
| GE000932 | Copper oxidase | 2.19 | CotAb (22.9%) | |
| GE001625 | Lac | Multi-copper polyphenol oxidoreductase laccase | 3.12 | Lacc (26.89) |
| GE001899 | bisdA | Ferredoxin | 3.16 | bisdAd (30%) |
| GE002971 | Ferredoxin | 3.34 | bisdAd (31.37%) | |
| GE002964 | P450 | 3.83 | bisdBe (27.86%) | |
| GE002994 | Cytochrome P450 | 3.76 | bisdBe (24.86%) | |
| GE002989 | ProJ | Hydrolase | 1.58 | proJf (27.05%) |
| GE003785 | Aldo/keto reductase | - | reductase g (32.01%) |
Note(s): a hydroxylase from Sphingobium sp. BiD32, NCBI version No. CCW19852.1; b CotA from B. pumilus A56, NCBI version No. JQ035528.1; c Lac from Bacillus subtilis ATCC 19217, NCBI version No. AIW37322.1; d bisdA from Sphingomonas bisphenolicum strain AO1, NCBI version No. BAG15883.1; e bisdB from Sphingomonas bisphenolicum strain AO1, NCBI version No. BAG15884.1; f ProJ from Sphingobium sp. BiD32, NCBI version No. CCW18563.1; g reductase from Sphingobium sp. BiD32, NCBI version No. CCW18590.1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, S.; Tian, K.; Qiu, Q.; Yu, Y.; Li, H.; Chang, M.; Sun, X.; Gu, J.; Zhang, F.; Wang, Y.; et al. Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1. Water 2023, 15, 830. https://doi.org/10.3390/w15040830
AMA Style
Li S, Tian K, Qiu Q, Yu Y, Li H, Chang M, Sun X, Gu J, Zhang F, Wang Y, et al. Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1. Water. 2023; 15(4):830. https://doi.org/10.3390/w15040830
Chicago/Turabian StyleLi, Shuaiguo, Kejian Tian, Qing Qiu, Yue Yu, Han Li, Menghan Chang, Xuejian Sun, Jinming Gu, Fenglin Zhang, Yibing Wang, and et al. 2023. "Study on Genomics of the Bisphenol A-Degrading Bacterium Pseudomonas sp. P1" Water 15, no. 4: 830. https://doi.org/10.3390/w15040830
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.