
Characterizing Free-Living and Particle-Attached Bacterial Communities of a Shallow Lake on the Inner Mongolia-Xinjiang Plateau, China
Abstract
:1. Introduction
2. Materials and Methods
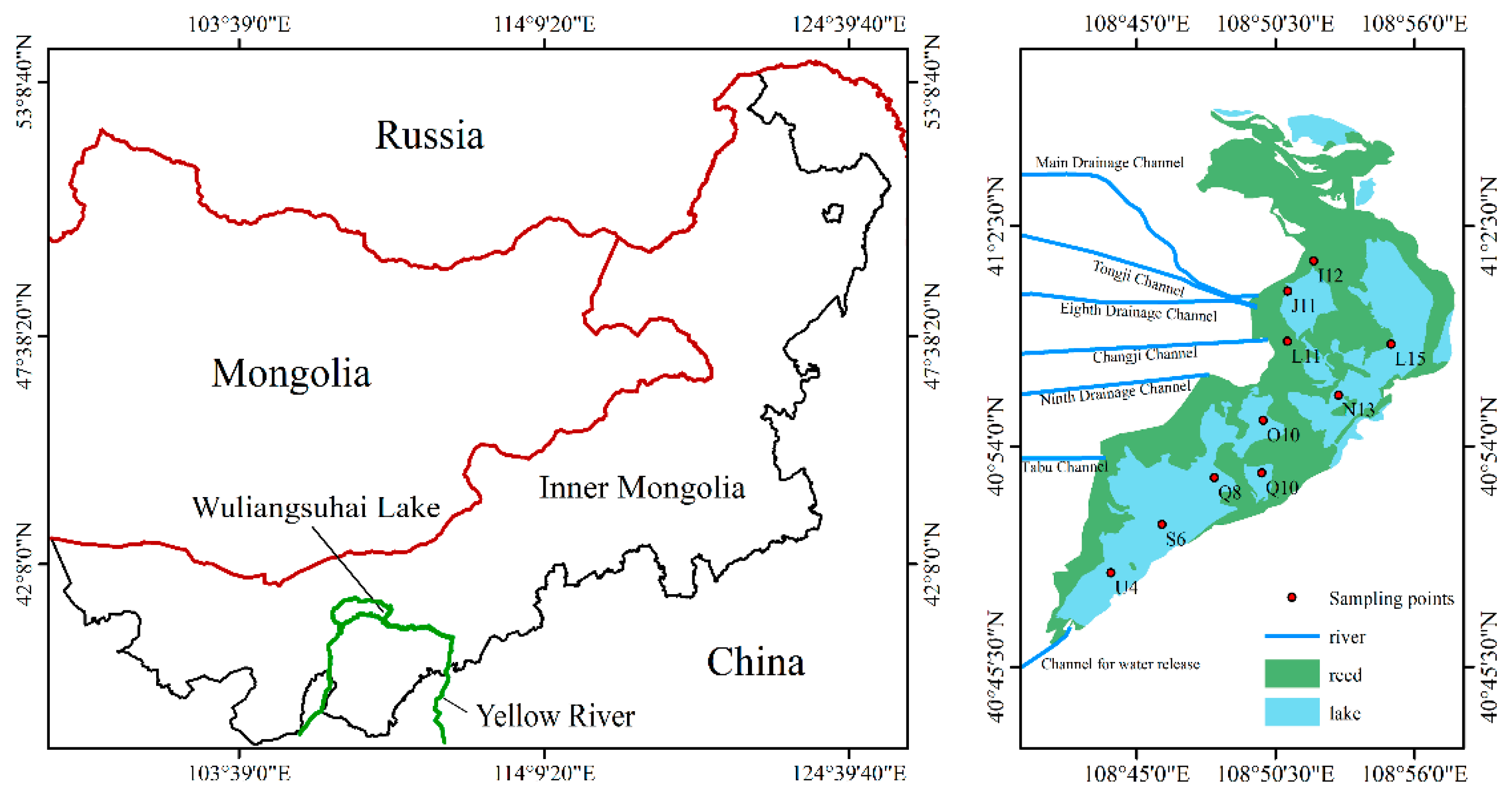
2.1. Study Area and Sample Collection
2.2. Determination of Physical and Chemical Indicators of Water Bodies
2.3. DNA Extraction and PCR Amplification
2.4. Data Analysis
3. Results
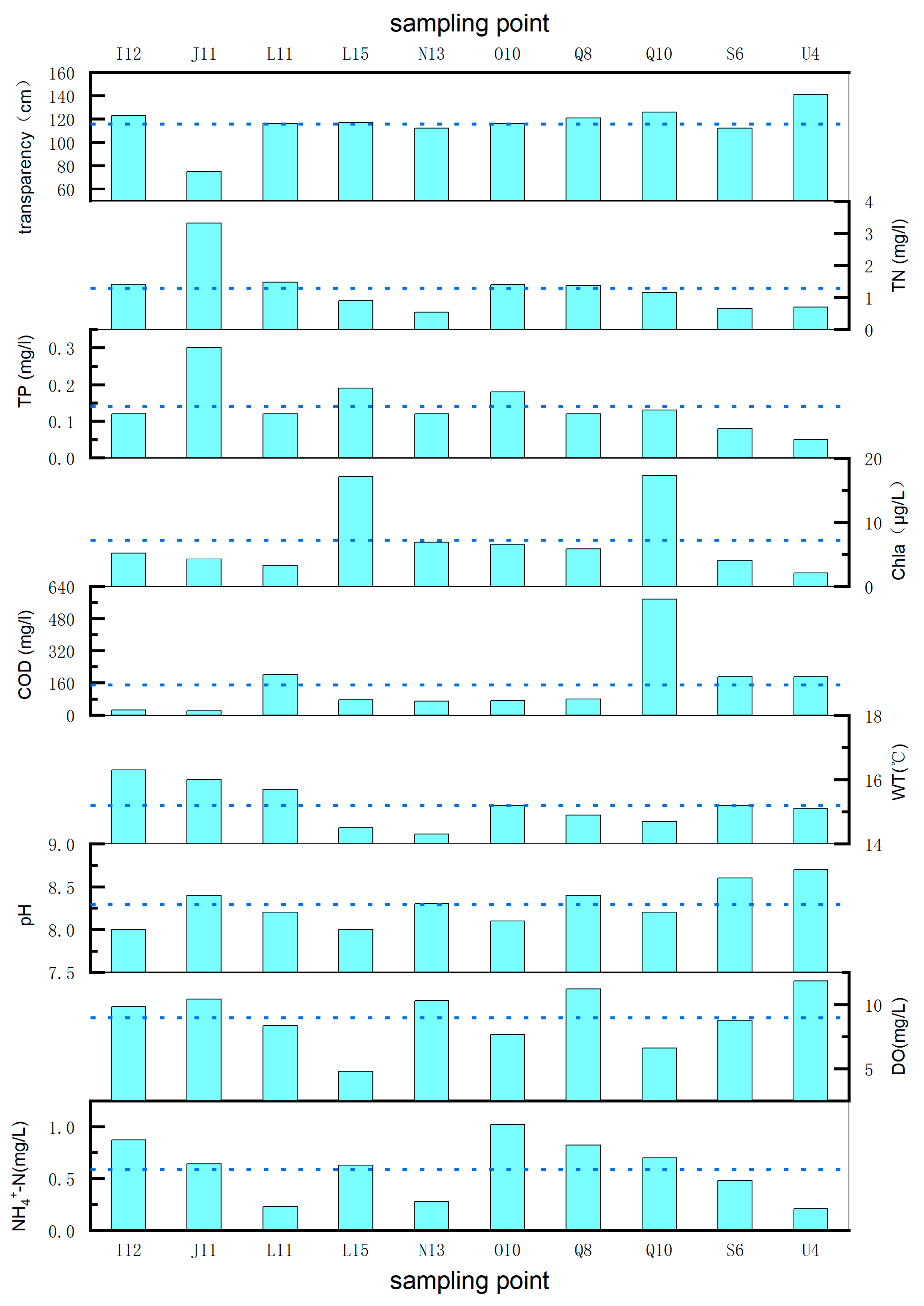
3.1. Physical and Chemical Properties of the Water in Wuliangsuhai Lake
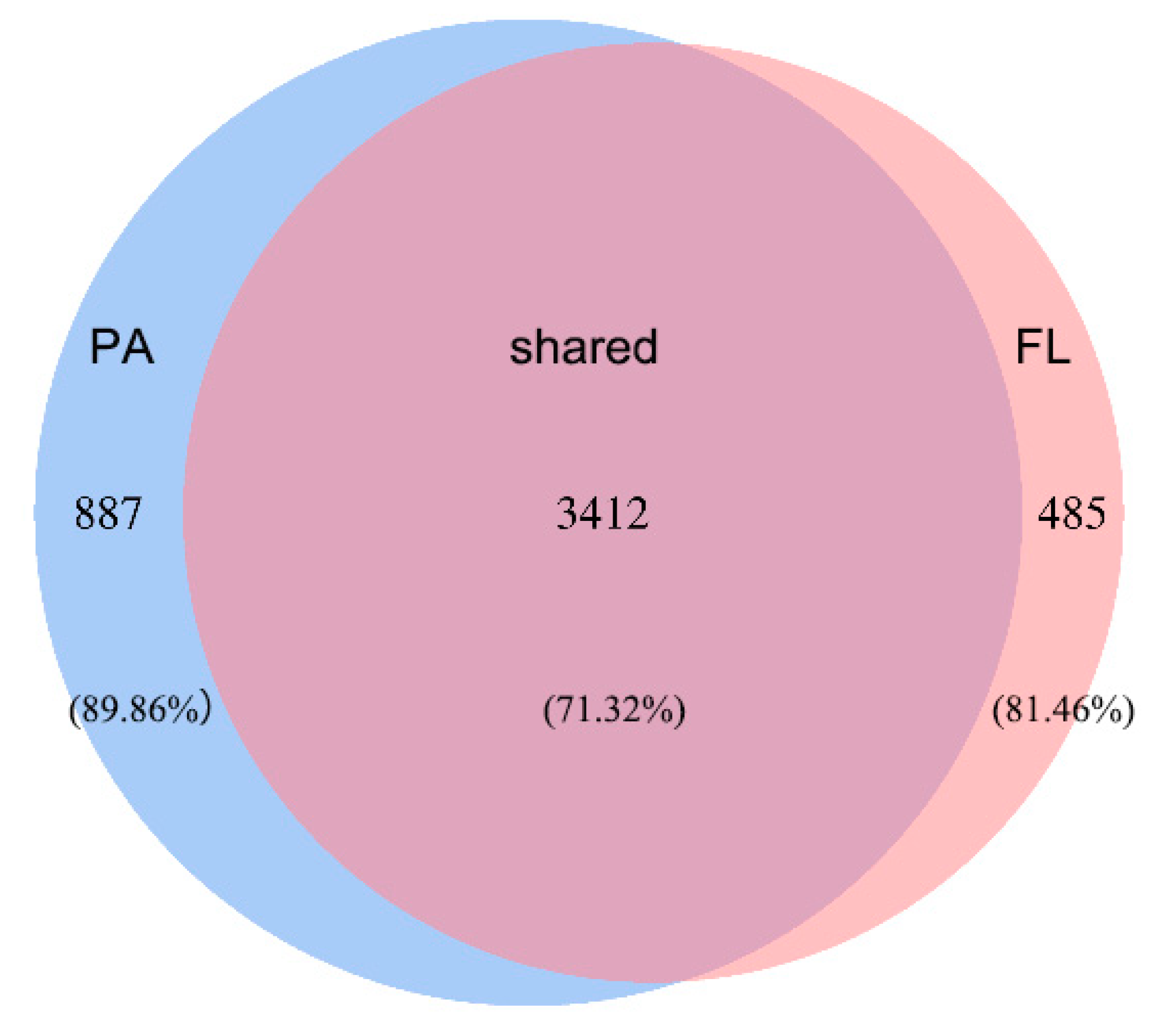
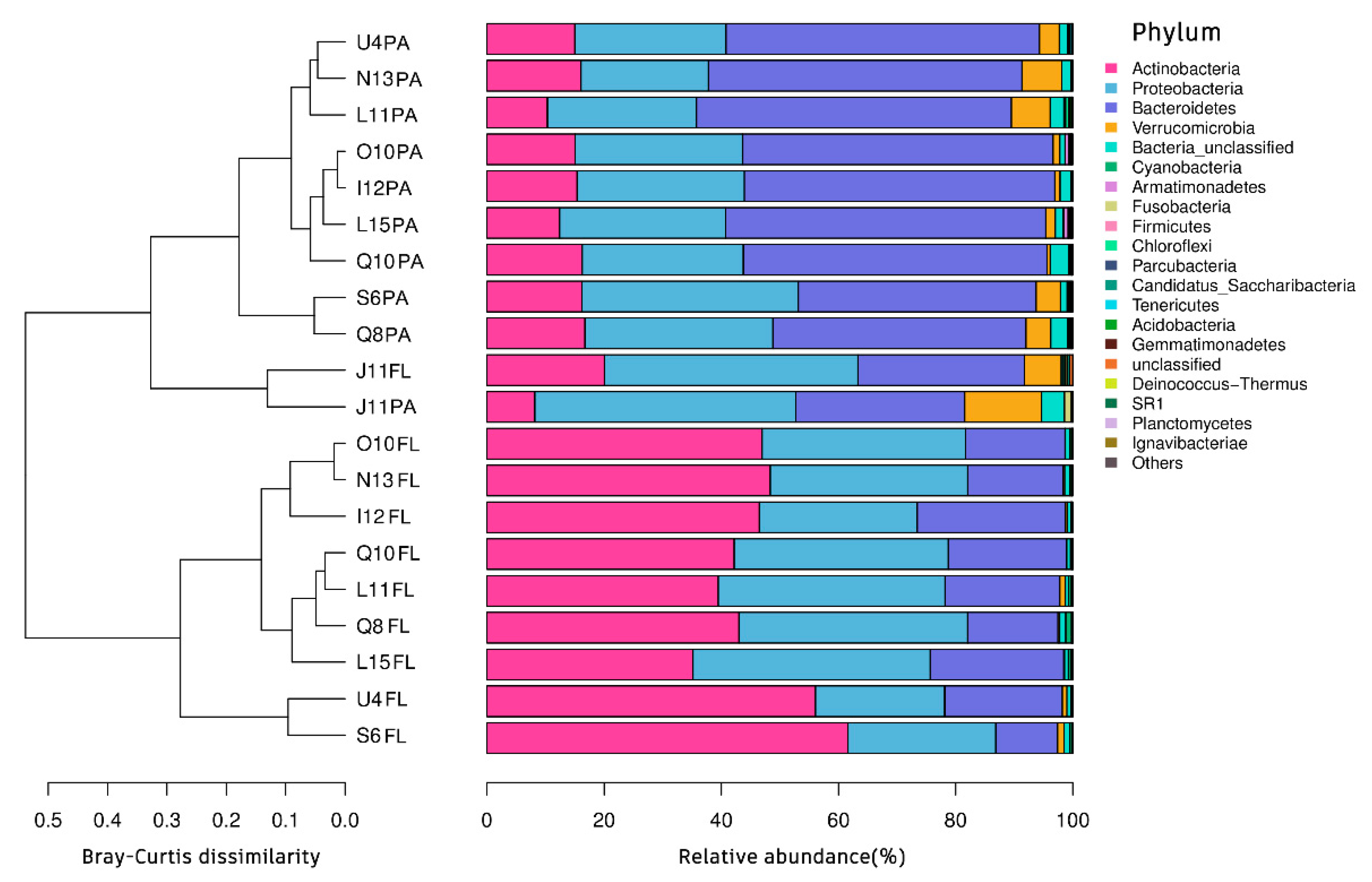
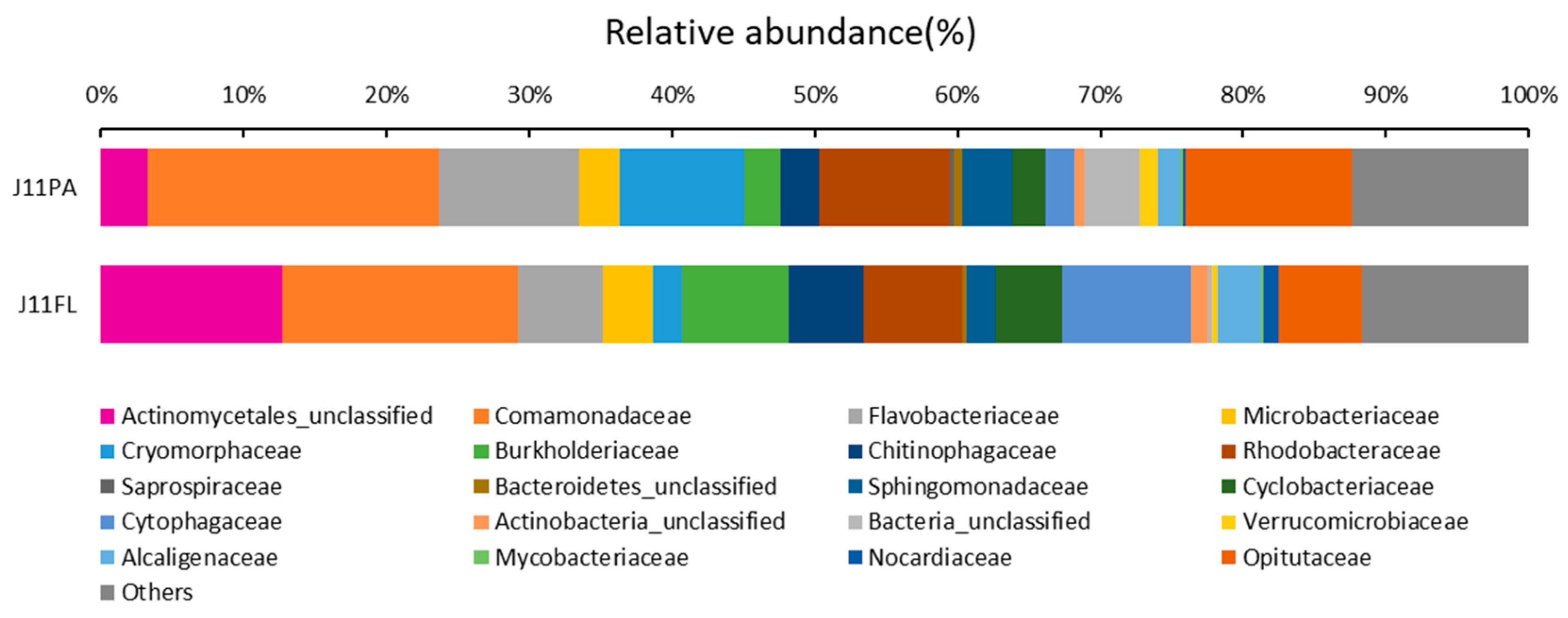
3.2. OTU Clustering and Bacterial Community Structure
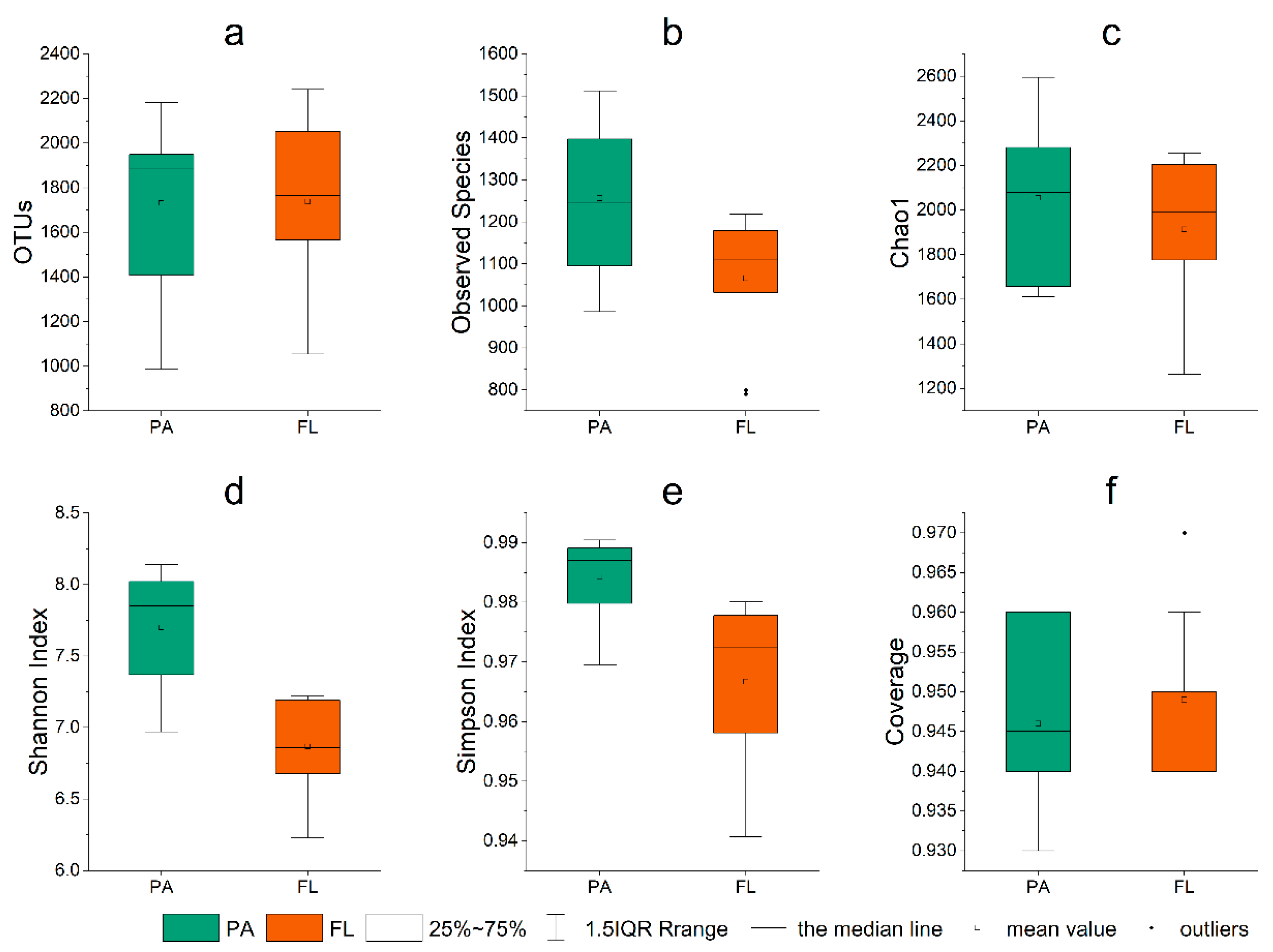
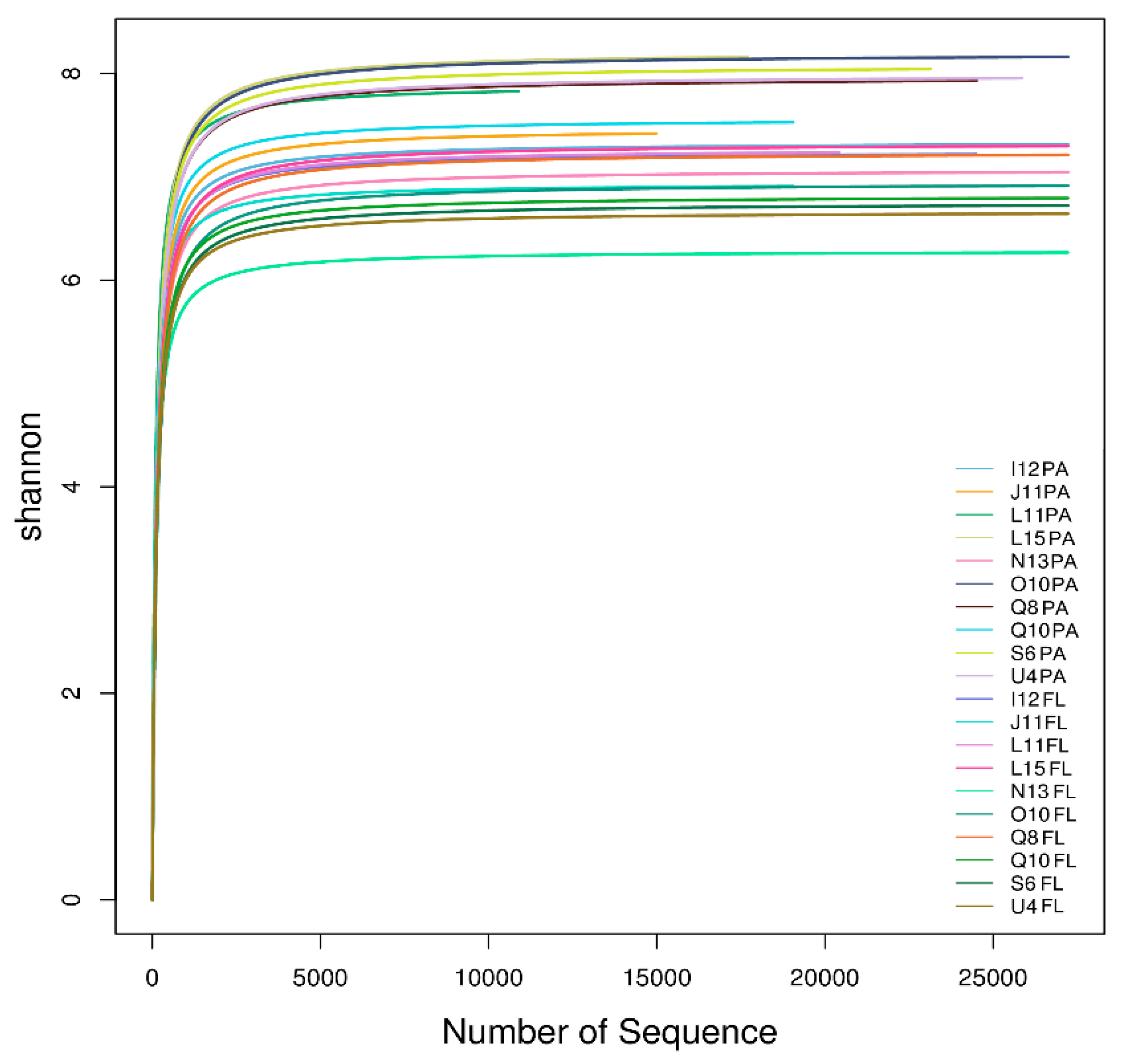
3.3. Alpha Diversity
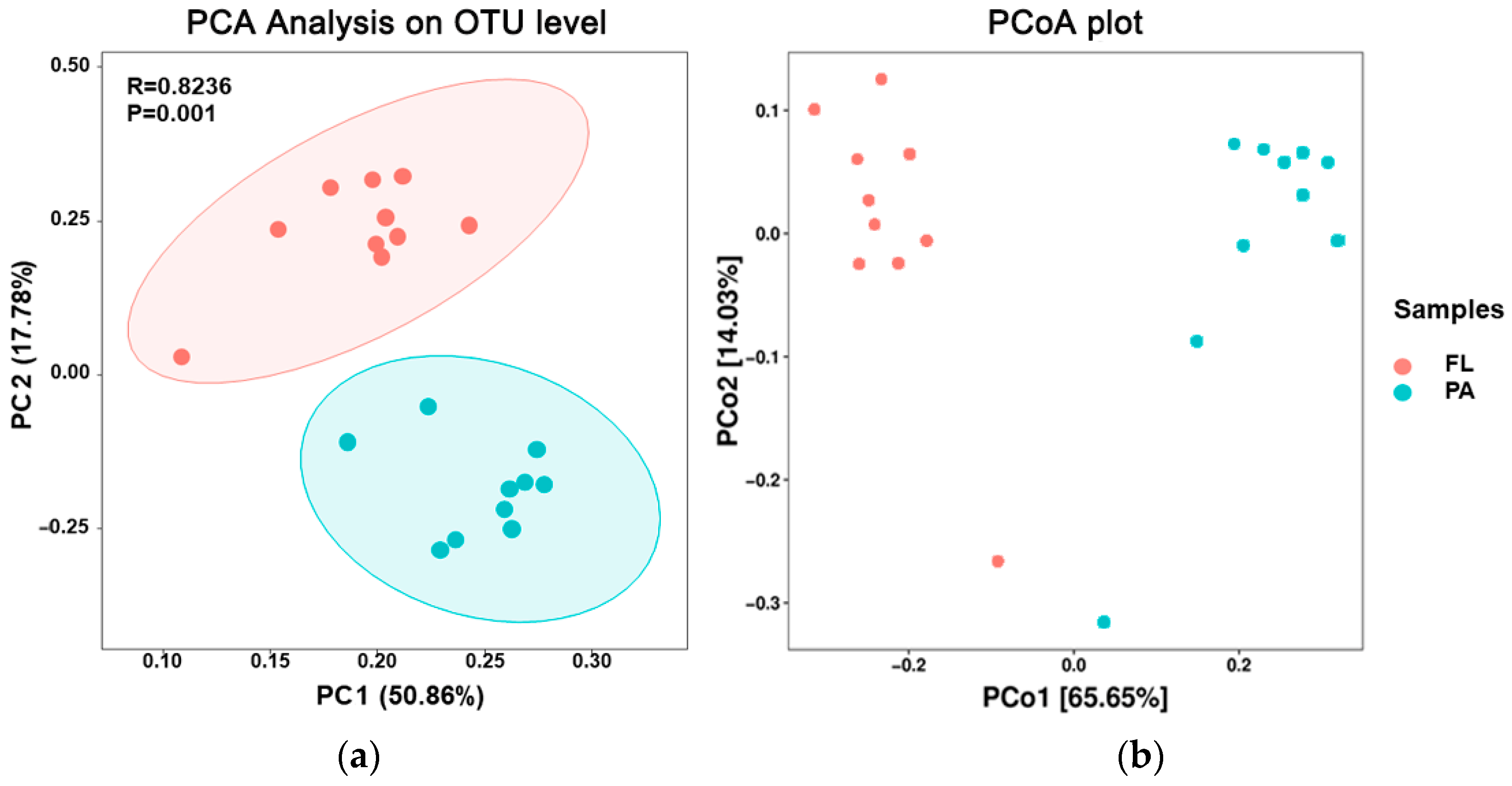
3.4. Beta Diversity Analysis
3.5. Correlation Analysis of Bacteria and Environmental Factors
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Points | Transparency | TN | TP | Chla | COD | WT | pH | DO | -N |
|---|---|---|---|---|---|---|---|---|---|
| cm | mg/L | mg/L | μg/L | mg/L | °C | mg/L | mg/L | ||
| I12 | 123 | 1.4 | 0.12 | 5.16 | 24 | 16.3 | 8 | 9.84 | 0.87 |
| J11 | 75 | 3.31 | 0.3 | 4.26 | 20 | 16 | 8.4 | 10.42 | 0.64 |
| L11 | 116 | 1.47 | 0.12 | 3.28 | 200 | 15.7 | 8.2 | 8.34 | 0.23 |
| L15 | 117 | 0.89 | 0.19 | 17.08 | 76 | 14.5 | 8 | 4.78 | 0.63 |
| N13 | 112 | 0.54 | 0.12 | 6.88 | 68 | 14.3 | 8.3 | 10.3 | 0.28 |
| O10 | 116 | 1.39 | 0.18 | 6.57 | 72 | 15.2 | 8.1 | 7.67 | 1.02 |
| Q8 | 121 | 1.37 | 0.12 | 5.86 | 80 | 14.9 | 8.4 | 11.2 | 0.82 |
| Q10 | 126 | 1.16 | 0.13 | 17.26 | 576 | 14.7 | 8.2 | 6.62 | 0.7 |
| S6 | 112 | 0.66 | 0.08 | 4.06 | 188 | 15.2 | 8.6 | 8.78 | 0.48 |
| U4 | 141 | 0.7 | 0.05 | 2.12 | 188 | 15.1 | 8.7 | 11.84 | 0.21 |
| Family | J11PA | J11FL |
|---|---|---|
| Actinomycetales_unclassified | 3.29 | 12.67 |
| Comamonadaceae | 20.37 | 16.59 |
| Flavobacteriaceae | 9.88 | 5.87 |
| Microbacteriaceae | 2.80 | 3.55 |
| Cryomorphaceae | 8.68 | 2.03 |
| Burkholderiaceae | 2.62 | 7.44 |
| Chitinophagaceae | 2.64 | 5.28 |
| Rhodobacteraceae | 9.14 | 6.93 |
| Saprospiraceae | 0.33 | 0.00 |
| Bacteroidetes_unclassified | 0.58 | 0.22 |
| Sphingomonadaceae | 3.46 | 2.09 |
| Cyclobacteriaceae | 2.39 | 4.68 |
| Cytophagaceae | 2.06 | 9.01 |
| Actinobacteria_unclassified | 0.67 | 1.14 |
| Bacteria_unclassified | 3.89 | 0.35 |
| Verrucomicrobiaceae | 1.20 | 0.34 |
| Alcaligenaceae | 1.60 | 3.01 |
| Mycobacteriaceae | 0.13 | 0.25 |
| Nocardiaceae | 0.22 | 1.05 |
| Opitutaceae | 11.69 | 5.85 |
| Others | 12.35 | 11.64 |
| Sampling Points | Observed_Species | Chao1 | Shannon | Simpson | Gcoverage |
|---|---|---|---|---|---|
| I12PA | 1138.00 | 1935.36 | 7.24 | 0.98 | 0.95 |
| J11PA | 1095.00 | 1657.13 | 7.37 | 0.98 | 0.96 |
| L11PA | 986.00 | 1610.96 | 7.84 | 0.99 | 0.96 |
| L15PA | 1512.00 | 2595.17 | 8.11 | 0.99 | 0.93 |
| N13PA | 1088.00 | 1944.61 | 6.97 | 0.97 | 0.95 |
| O10PA | 1512.00 | 2397.88 | 8.14 | 0.99 | 0.93 |
| Q8PA | 1355.00 | 2265.85 | 7.86 | 0.99 | 0.94 |
| Q10PA | 1095.00 | 1633.90 | 7.52 | 0.99 | 0.96 |
| S6PA | 1397.00 | 2280.19 | 8.02 | 0.99 | 0.94 |
| U4PA | 1384.00 | 2214.41 | 7.92 | 0.99 | 0.94 |
| I12FL | 1126.00 | 2032.90 | 7.19 | 0.98 | 0.95 |
| J11FL | 799.00 | 1265.22 | 6.88 | 0.98 | 0.97 |
| L11FL | 1179.00 | 2243.21 | 7.21 | 0.98 | 0.94 |
| L15FL | 1218.00 | 2204.15 | 7.22 | 0.98 | 0.94 |
| N13FL | 790.00 | 1518.01 | 6.23 | 0.96 | 0.96 |
| O10FL | 1163.00 | 2256.77 | 6.84 | 0.96 | 0.94 |
| Q8FL | 1189.00 | 2092.37 | 7.12 | 0.98 | 0.94 |
| Q10FL | 1070.00 | 1802.10 | 6.75 | 0.97 | 0.95 |
| S6FL | 1092.00 | 1775.32 | 6.68 | 0.94 | 0.95 |
| U4FL | 1032.00 | 1947.00 | 6.56 | 0.95 | 0.95 |

References
- Yadav, A.N.; Yadav, N.; Kour, D.; Kumar, A.; Yadav, K.; Kumar, A.; Rastegari, A.A.; Sachan, S.G.; Singh, B.; Chauhan, V.S.; et al. Chapter 1—Bacterial community composition in lakes. In Freshwater Microbiology; Bandh, S.A., Shafi, S., Shameem, N., Eds.; Academic Press: London, UK, 2019; pp. 1–71. ISBN 978-0-12-817495-1. [Google Scholar]
- Hahn, M.W. The microbial diversity of inland waters. Curr. Opin. Biotechnol. 2006, 17, 256–261. [Google Scholar] [CrossRef]
- Paver, S.F.; Hayek, K.R.; Gano, K.A.; Fagen, J.R.; Brown, C.T.; Davis-Richardson, A.G.; Crabb, D.B.; Rosario-Passapera, R.; Giongo, A.; Triplett, E.W. Interactions between specific phytoplankton and bacteria affect lake bacterial community succession. Environ. Microbiol. 2013, 15, 2489–2504. [Google Scholar] [CrossRef] [PubMed]
- Urvoy, M.; Gourmelon, M.; Serghine, J.; Rabiller, E.; L’Helguen, S.; Labry, C. Free-living and particle-attached bacterial community composition, assembly processes and determinants across spatiotemporal scales in a macrotidal temperate estuary. Sci. Rep. 2022, 12, 13897. [Google Scholar] [CrossRef]
- Jiao, C.; Zhao, D.; Zeng, J.; Guo, L.; Yu, Z. Disentangling the seasonal co-occurrence patterns and ecological stochasticity of planktonic and benthic bacterial communities within multiple lakes. Sci. Total Environ. 2020, 740, 140010. [Google Scholar] [CrossRef]
- Xu, H.; Zhao, D.; Huang, R.; Cao, X.; Zeng, J.; Yu, Z.; Hooker, K.V.; Hambright, K.D.; Wu, Q.L. Contrasting Network Features between Free-Living and Particle-Attached Bacterial Communities in Taihu Lake. Microb. Ecol. 2018, 76, 303–313. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Chen, C.; Wang, J.; Xu, T. Characterizing free-living and particle-attached bacterial communities of a canyon river reservoir on the Yungui Plateau, China. Front. Microbiol. 2022, 13, 986637. [Google Scholar] [CrossRef] [PubMed]
- Bier, R.L.; Voss, K.A.; Bernhardt, E.S. Bacterial community responses to a gradient of alkaline mountaintop mine drainage in Central Appalachian streams. ISME J. 2015, 9, 1378–1390. [Google Scholar] [CrossRef] [Green Version]
- Garcia, S.L.; Salka, I.; Grossart, H.-P.; Warnecke, F. Depth-discrete profiles of bacterial communities reveal pronounced spatio-temporal dynamics related to lake stratification. Environ. Microbiol. Rep. 2013, 5, 549–555. [Google Scholar] [CrossRef]
- Ying, G.; Liu, F.; Sun, M.; Jiang, X.; Geng, J.; Teng, J.; Xie, W.; Zhang, H.; Chen, X. Community Structure and Influencing Factors of Bacterioplankton in Spring in Zhushan Bay, Lake Taihu. Environ. Sci. 2018, 39, 1151–1158. [Google Scholar] [CrossRef]
- Guo, D.; Liang, J.; Chen, W.; Wang, J.; Ji, B.; Luo, S. Bacterial Community Analysis of Two Neighboring Freshwater Lakes Originating from One Lake. Pol. J. Environ. Stud. 2021, 30, 111–117. [Google Scholar] [CrossRef]
- Fan, T.; Fang, W.; Zhao, Y.; Lu, A.; Wang, S.; Wang, X.; Xu, L.; Wei, X.; Zhang, L. Spatial Variations of Aquatic Bacterial Community Structure and Co-Occurrence Patterns in a Coal Mining Subsidence Lake. Diversity 2022, 14, 674. [Google Scholar] [CrossRef]
- Xie, Y.; Sheng, Y.; Li, D.; He, F.; Du, J.; Jiang, L.; Luo, C.; Li, G.; Zhang, D.; Du, J. Change of the structure and assembly of bacterial and photosynthetic communities by the ecological engineering practices in Dianchi Lake. Environ. Pollut. 2022, 315, 120386. [Google Scholar] [CrossRef] [PubMed]
- Yi, Y.; Lin, C.; Wang, W.; Song, J. Habitat and seasonal variations in bacterial community structure and diversity in sediments of a Shallow lake. Ecol. Indic. 2021, 120, 106959. [Google Scholar] [CrossRef]
- Xing, P.; Tao, Y.; Jeppesen, E.; Wu, Q.L. Comparing microbial composition and diversity in freshwater lakes between Greenland and the Tibetan Plateau. Limnol. Oceanogr. 2021, 66, S142–S156. [Google Scholar] [CrossRef]
- Liu, K.; Liu, Y.; Han, B.-P.; Xu, B.; Zhu, L.; Ju, J.; Jiao, N.; Xiong, J. Bacterial community changes in a glacial-fed Tibetan lake are correlated with glacial melting. Sci. Total Environ. 2019, 651, 2059–2067. [Google Scholar] [CrossRef]
- Fan, Y.; Hu, N.; Ding, S.; Liang, G.; Lu, X. Progress in terrestrial ecosystem services and biodiversity. Acta Ecol. Sin. 2016, 36, 4583–4593. [Google Scholar] [CrossRef]
- Zhang, H.; Yang, L.; Li, Y.; Wang, C.; Zhang, W.; Wang, L.; Niu, L. Pollution gradients shape the co-occurrence networks and interactions of sedimentary bacterial communities in Taihu Lake, a shallow eutrophic lake. J. Environ. Manag. 2022, 305, 114380. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Li, W.; Zhang, B.; Yao, G.; Shi, X. Characteristics of Bacterial Community Structure in Wuliangsu Lake During an Irrigation Interval in Hetao Plain. Environ. Sci. 2022, 43, 1424–1433. [Google Scholar] [CrossRef]
- Shen, Z.; Shang, Z.; Wang, F.; Liang, Y.; Zou, Y.; Liu, F. Bacterial diversity in surface sediments of collapsed lakes in Huaibei, China. Sci. Rep. 2022, 12, 15784. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Zhao, L.; Li, Y.; Xie, S.; Liu, Y. Distribution of sediment bacterial and archaeal communities in plateau freshwater lakes. Appl. Microbiol. Biotechnol. 2015, 99, 3291–3302. [Google Scholar] [CrossRef] [PubMed]
- Zhen, S.; Guijuan, X.; Yuqing, Z.; Bobing, Y.; Keqiang, S.; Guang, G.; Xiangming, T. Similar assembly mechanisms but distinct co-occurrence patterns of free-living vs. particle-attached bacterial communities across different habitats and seasons in shallow, eutrophic Lake Taihu. Environ. Pollut. 2022, 314, 120305. [Google Scholar] [CrossRef]
- Zhang, W.; Wan, W.; Lin, H.; Pan, X.; Lin, L.; Yang, Y. Nitrogen rather than phosphorus driving the biogeographic patterns of abundant bacterial taxa in a eutrophic plateau lake. Sci. Total Environ. 2021, 806, 150947. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Ramanathan, V.; Victor, D.G. Global warming will happen faster than we think. Nature 2018, 564, 30–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Yang, J.; Yang, F.; Yang, W.; Li, W.; Li, X. Distribution Characteristics of Microplastics in Ice Sheets and Its Response to Salinity and Chlorophyll a in the Lake Wuliangsuhai. Environ. Sci. 2021, 42, 673–680. [Google Scholar] [CrossRef]
- Sun, H.; Yu, R.; Liu, X.; Cao, Z.; Li, X.; Zhang, Z.; Wang, J.; Zhuang, S.; Ge, Z.; Zhang, L.; et al. Drivers of spatial and seasonal variations of CO2 and CH4 fluxes at the sediment water interface in a shallow eutrophic lake. Water Res. 2022, 222, 118916. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lu, X.; Yu, R.; Yang, J.; Liu, X.; Cao, Z.; Zhang, Z.; Li, M.; Geng, Y. Eutrophication decreased CO2 but increased CH4 emissions from lake: A case study of a shallow Lake Ulansuhai. Water Res. 2021, 201, 117363. [Google Scholar] [CrossRef]
- Quan, D.; Shi, X.; Zhao, S.; Zhang, S.; Liu, J. Eutrophication of Lake Ulansuhai in 2006–2017 and its main impact factors. J. Lake Sci. 2019, 31, 1259–1267. [Google Scholar] [CrossRef] [Green Version]
- Yang, F.; Li, C.; Shi, X.; Zhao, S.; Hao, Y. Impact of seasonal ice structure characteristics on ice cover impurity distributions in Lake Ulansuhai. J. Lake Sci. 2016, 28, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Roulet, N.T.; Strachan, I.B.; Tremblay, A. Modeling surface energy fluxes and thermal dynamics of a seasonally ice-covered hydroelectric reservoir. Sci. Total Environ. 2016, 550, 793–805. [Google Scholar] [CrossRef]
- Zhai, J.; Shi, X.; Liu, Y.; Zhao Sh, e.; Bao, W.Z.; Li, G. Change law of water temperature and dissolved oxygen concentration of Wuliangsu Sea in icebound period. Arid. Zone Res. 2021, 38, 629–639. [Google Scholar] [CrossRef]
- Müller, M.; Sekoulov, I. Waste Water Reuse by Freeze Concentration with a Falling Film Reactor. Water Sci. Technol. 1992, 26, 1475–1482. [Google Scholar] [CrossRef]
- Hao, R.; Shi, X.; Liu, Y.; Zhang, F. Spatial distribution and influencing factors of microplastics in water of Ulansuhai. China Environ. Sci. 2022, 42, 3316–3324. [Google Scholar] [CrossRef]
- Dickerson, T.L.; Williams, H.N. Functional Diversity of Bacterioplankton in Three North Florida Freshwater Lakes over an Annual Cycle. Microb. Ecol. 2014, 67, 34–44. [Google Scholar] [CrossRef]
- Yang, C.; Wang, Q.; Simon, P.N.; Liu, J.; Liu, L.; Dai, X.; Zhang, X.; Kuang, J.; Igarashi, Y.; Pan, X.; et al. Distinct Network Interactions in Particle-Associated and Free-Living Bacterial Communities during a Microcystis aeruginosa Bloom in a Plateau Lake. Front. Microbiol. 2017, 8, 1202. [Google Scholar] [CrossRef] [Green Version]
- Kanukollu, S.; Wemheuer, B.; Herber, J.; Billerbeck, S.; Lucas, J.; Daniel, R.; Simon, M.; Cypionka, H.; Engelen, B. Distinct compositions of free-living, particle-associated and benthic communities of the Roseobacter group in the North Sea. FEMS Microbiol. Ecol. 2016, 92, fiv145. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.C.; Tu, Q.Y. The Standard Methods for Observation and Analysis in Lake Eutrophication, 2nd ed.; Chinese Environmental Science Press: Beijing, China, 1990. [Google Scholar]
- Zeng, Q.; An, S. Identifying the Biogeographic Patterns of Rare and Abundant Bacterial Communities Using Different Primer Sets on the Loess Plateau. Microorganisms 2021, 9, 139. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M.; et al. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. Isme J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Delgado-Baquerizo, M.; Shi, Y.; Liu, X.; Yang, Y.; Chu, H. Co-existing water and sediment bacteria are driven by contrasting environmental factors across glacier-fed aquatic systems. Water Res. 2021, 198, 117139. [Google Scholar] [CrossRef]
- Jiao, C.; Zhao, D.; Huang, R.; Cao, X.; Zeng, J.; Lin, Y.; Zhao, W. Abundant and Rare Bacterioplankton in Freshwater Lakes Subjected to Different Levels of Tourism Disturbances. Water 2018, 10, 1075. [Google Scholar] [CrossRef] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon 1972, 21, 213–251. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao, A. Nonparametric Estimation of the Number of Classes in a Population. Scand. J. Stat. 1984, 11, 265–270. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Labs Tech. J. 1948, 27, 623–656. [Google Scholar] [CrossRef]
- Fu, P.; Bai, L.; Cai, Z.; Li, R.; Yung, K.K.L. Fine particulate matter aggravates intestinal and brain injury and affects bacterial community structure of intestine and feces in Alzheimer’s disease transgenic mice. Ecotoxicol. Environ. Saf. 2020, 192, 110325. [Google Scholar] [CrossRef]
- Lever, J.; Krzywinski, M.; Altman, N. Principal component analysis. Nat. Methods 2017, 14, 641–642. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Ibekwe, A.M.; Ma, J.; Murinda, S.E. Bacterial community composition and structure in an Urban River impacted by different pollutant sources. Sci. Total Environ. 2016, 566, 1176–1185. [Google Scholar] [CrossRef] [Green Version]
- Ter Braak, C.J.F. CANOCO—A FORTRAN Program for Canonical Community Ordination by Partial Detrended Canonical Correspondence Analysis, Principal Components Analysis and Redundancy Analysis; MLV: Wageningen, The Netherlands, 1988; (Technical report/Ministerie van Landbouw en Visserij, Groep Landbouwwiskunde; LWA-88-02). [Google Scholar]
- Ding, N.; Yang, Y.; Wan, N.; Xu, A.; Ge, J.; Song, Z. Seasonal Variation and Influencing Factors of Bacterial Communities in Storage Reservoirs. Environ. Sci. 2022, 1–14. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, X.; Liu, Y.; Xie, S.; Liu, Y. Bacterioplankton communities in a high-altitude freshwater wetland. Ann. Microbiol. 2014, 64, 1405–1411. [Google Scholar] [CrossRef]
- Yao, T.; Yan, H.; Liao, X.; Wang, Z.; Liu, C.; Wang, Z.; Chen, Q. Characteristics of nitrogen transformation and microbial community structure in the diversion river: A case study of the Wangyu River. Acta Sci. Circumstantiae 2022, 42, 195–204. [Google Scholar] [CrossRef]
- Shao, K.; Yao, X.; Wu, Z.; Jiang, X.; Hu, Y.; Tang, X.; Xu, Q.; Gao, G. The bacterial community composition and its environmental drivers in the rivers around eutrophic Chaohu Lake, China. Bmc Microbiol. 2021, 21, 179. [Google Scholar] [CrossRef]
- Sun, L.; Wang, J.; Wu, Y.; Gao, T.; Liu, C. Community Structure and Function of Epiphytic Bacteria Associated With Myriophyllum spicatum in Baiyangdian Lake, China. Front. Microbiol. 2021, 12, 705509. [Google Scholar] [CrossRef]
- Kolmonen, E.; Haukka, K.; Rantala-Ylinen, A.; Rajaniemi-Wacklin, P.; Lepisto, L.; Sivonen, K. Bacterioplankton community composition in 67 Finnish lakes differs according to trophic status. Aquat. Microb. Ecol. 2011, 62, 241–250. [Google Scholar] [CrossRef]
- Ma, J.; Shi, R.; Han, R.; Ji, M.; Xu, X.; Wang, G. Community structure of epiphytic bacteria on Potamogeton pectinatus and the surrounding bacterioplankton in Hongze Lake. Mar. Freshw. Res. 2021, 72, 997–1003. [Google Scholar] [CrossRef]
- Zhong, M.; Capo, E.; Zhang, H.; Hu, H.; Wang, Z.; Tian, W.; Huang, T.; Bertilsson, S. Homogenisation of water and sediment bacterial communities in a shallow lake (lake Balihe, China). Freshw. Biol. 2023, 68, 155–171. [Google Scholar] [CrossRef]
- Richardson, T.L.; Gibson, C.E.; Heaney, S.I. Temperature, growth and seasonal succession of phytoplankton in Lake Baikal, Siberia. Freshw. Biol. 2008, 44, 431–440. [Google Scholar] [CrossRef]
- Allgaier, M.; Brueckner, S.; Jaspers, E.; Grossart, H.-P. Intra- and inter-lake variability of free-living and particle-associated Actinobacteria communities. Environ. Microbiol. 2007, 9, 2728–2741. [Google Scholar] [CrossRef]
- Sebastián, F.; Valentina, M.; Patricia, A.; Michael, S. Bioremediation of petroleum hydrocarbons: Catabolic genes, microbial communities, and applications. Appl. Microbiol. Biotechnol. 2014, 98, 4781–4794. [Google Scholar] [CrossRef]
- Wan, T.; He, M.; Ren, J.-H.; Yan, X.; Cheng, W. Environmental Response and Ecological Function Prediction of Aquatic Bacterial Communities in the Weihe River Basin. Environ. Sci. 2019, 40, 3588–3595. [Google Scholar] [CrossRef]
- Parveen, B.; Mary, I.; Vellet, A.; Ravet, V.; Debroas, D. Temporal dynamics and phylogenetic diversity of free-living and particle-associated Verrucomicrobia communities in relation to environmental variables in a mesotrophic lake. FEMS Microbiol. Ecol. 2013, 83, 189–201. [Google Scholar] [CrossRef] [Green Version]
- Chiang, E.; Schmidt, M.L.; Berry, M.A.; Biddanda, B.A.; Burtner, A.; Johengen, T.H.; Palladino, D.; Denef, V.J. Verrucomicrobia are prevalent in north-temperate freshwater lakes and display class-level preferences between lake habitats. PLoS ONE 2018, 13, e0195112. [Google Scholar] [CrossRef] [Green Version]
- Hedlund, B.P.; Gosink, J.J.; Staley, J.T. Verrucomicrobia div. nov., a new division of the Bacteria containing three new species of Prosthecobacter. Antonie Leeuwenhoek 1997, 72, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Chin, K.J.; Hahn, D.; Hengstmann, U.; Liesack, W.; Janssen, P.H. Characterization and identification of numerically abundant culturable bacteria from the anoxic bulk soil of rice paddy microcosms. Appl. Environ. Microbiol. 1999, 65, 5042–5049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bergmann, G.T.; Bates, S.T.; Eilers, K.G.; Lauber, C.L.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. The under-recognized dominance of Verrucomicrobia in soil bacterial communities. Soil Biol. Biochem. 2011, 43, 1450–1455. [Google Scholar] [CrossRef] [Green Version]
- Tran, P.; Ramachandran, A.; Khawasik, O.; Beisner, B.E.; Rautio, M.; Huot, Y.; Walsh, D.A. Microbial life under ice: Metagenome diversity and in situ activity of Verrucomicrobia in seasonally ice-covered Lakes. Environ. Microbiol. 2018, 20, 2568–2584. [Google Scholar] [CrossRef] [Green Version]
- Yu, S.; Li, S.; Tang, Y. Succession of bacterial community along with the removal of heavy crude oil pollutants by multiple biostimulation treatments in the Yellow River Delta, China. Acta Sci. Circumstantiae 2011, 23, 1533–1543. [Google Scholar] [CrossRef]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients? NUTRIENTS 2020, 12, 1474. [Google Scholar] [CrossRef]
- Nshimyimana, J.P.; Freedman, A.; Shanahan, P.; Chua, L.; Thompson, J.R. Variation of Bacterial Communities with Water Quality in an Urban Tropical Catchment. Environ. Sci. Technol. 2017, 51, 5591–5601. [Google Scholar] [CrossRef]
- Du, R.; Li, J.; Zhao, J. Bacterial diversity in littoral wetland of Wuliangsuhai Lake. Acta Microbiol. Sin. 2014, 54, 1116–1128. [Google Scholar] [CrossRef]
- Liu, J.; Yi, N.; Wang, S.; Lu, L.; Huang, X. Impact of plant species on spatial distribution of metabolic potential and functional diversity of microbial communities in a constructed wetland treating aquaculture wastewater. Ecol. Eng. 2016, 94, 564–573. [Google Scholar] [CrossRef]
- Jin, Z.; Tu, C.; Wang, S.; Chen, J.; Lu, c.; Huang, W. Phosphorus adsorption characteristics and loss risk in sediments of lake day. Environ. Sci. 2022, 43, 1976–1987. [Google Scholar] [CrossRef]








| Index | PA | FL | ||
|---|---|---|---|---|
| Rho | p-Value | Rho | p-Value | |
| Transparency | 0.490 | 0.057 | 0.582 | 0.016 |
| TN | 0.509 | 0.030 | 0.589 | 0.008 |
| TP | 0.328 | 0.137 | 0.404 | 0.071 |
| Chla | −0.152 | 0.760 | −0.073 | 0.589 |
| COD | 0.267 | 0.134 | 0.104 | 0.267 |
| WT | 0.395 | 0.036 | 0.507 | 0.007 |
| pH | 0.134 | 0.210 | 0.168 | 0.164 |
| DO | −0.370 | 0.983 | −0.301 | 0.970 |
| -N | −0.114 | 0.753 | −0.104 | 0.717 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Shi, X.; Zhao, S.; Sun, B.; Liu, Y.; Li, W.; Yu, H.; Tian, Z.; Guo, X.; Shi, Y.; et al. Characterizing Free-Living and Particle-Attached Bacterial Communities of a Shallow Lake on the Inner Mongolia-Xinjiang Plateau, China. Water 2023, 15, 836. https://doi.org/10.3390/w15050836
Wang Y, Shi X, Zhao S, Sun B, Liu Y, Li W, Yu H, Tian Z, Guo X, Shi Y, et al. Characterizing Free-Living and Particle-Attached Bacterial Communities of a Shallow Lake on the Inner Mongolia-Xinjiang Plateau, China. Water. 2023; 15(5):836. https://doi.org/10.3390/w15050836
Chicago/Turabian StyleWang, Yanjun, Xiaohong Shi, Shengnan Zhao, Biao Sun, Yu Liu, Wenbao Li, Haifeng Yu, Zhiqiang Tian, Xin Guo, Yujiao Shi, and et al. 2023. "Characterizing Free-Living and Particle-Attached Bacterial Communities of a Shallow Lake on the Inner Mongolia-Xinjiang Plateau, China" Water 15, no. 5: 836. https://doi.org/10.3390/w15050836