
Niche Analysis of the Main Fish in the Lhasa River Basin
by
 ,
,
He Gao
1,
Haiping Liu
2,
Qiming Wang
3,
Fei Liu
4,
Junting Li
1,
Yuting Duan
1,
Luowu Zeren
1,
Suxing Fu
1,
Jingsen Zhang
1,
Yan Zhou
1,
Yan Li
1 and
Chaowei Zhou
1,* 1
College of Fisheries, Southwest University, Chongqing 402460, China
2
Integrative Science Center of Germplasm Creation in Western China (Chongqing) Science City & Southwest University, Chongqing 402460, China
3
Agriculture and Rural Affairs Department of Tibet Autonomous Region, Lhasa 850000, China
4
Institute of Fisheries Science, Tibet Academy of Agricultural and Animal Husbandry Sciences, Lhasa 851418, China
*
Author to whom correspondence should be addressed.
Water 2023, 15(5), 860; https://doi.org/10.3390/w15050860
Submission received: 17 January 2023
/
Revised: 11 February 2023
/
Accepted: 12 February 2023
/
Published: 23 February 2023
(This article belongs to the Special Issue Ecology of Freshwater Fishes)
Abstract
:The Lhasa River is one of the five tributaries of the Yarlung Zangbo River. It is the main industrial, agricultural and animal husbandry area in Tibet. It plays an important ecological security barrier role in regulating regional climate and maintaining biodiversity. In order to understand the resource utilization ability and relationship of fish in Lhasa River, and provide basic data for the protection and sustainable utilization of fish diversity in Lhasa River, the fishery resources in September 2019 (autumn), June 2020 (summer), November 2020 (winter) and March 2021 (spring) were investigated for the first time. The main fish in the catch were determined by the Index of Relative Importance (IRI). Shannon index and Pianka’s index were used to analyze the niche breadth and overlap values of the main fish caught in Lhasa River Basin from three dimensions of time, space and space–time. The results show that a total of 1399 fish of 3 orders and 4 families were collected. Among them, there is one dominant species, Schizopygopsis younghusbandi, and five common species, Schizothorax waltoni, Schizothorax oconnori, Ptychobarbus dipogon, Triplophysa stenura and Carassius auratus. The spatial–temporal niche breadth ranges from 1.395 to 3.628. They are Schizopygopsis younghusbandi, Carassius auratus, Ptychobarbus dipogon, Schizothorax oconnori, Triplophysa stenura and Schizothorax waltoni in a declining order. The temporal niche breadth is generally small, indicating that the survival activities of the main fish are greatly affected by seasonal changes. The fluctuation range of spatial–temporal overlap value is 0–0.535, and only 20% of the total logarithm is meaningful. The species pairs with temporal niche overlap values exceeding 0.300 reached 80%, while the spatial niche overlap value is generally low. The results show that the spatial and temporal distributions of the main fish in Lhasa River were quite different, and the spatial and temporal niche overlap value of the relationship between species is obviously affected by the change in spatial distribution. This demonstrates that the competition relationship and intensity of the main fish are quite different in different spatial positions.
1. Introduction
Ecological niches, as an important theory in ecology, have a wide range of applications in the study of interspecific relationships, community structure and succession, biodiversity and population evolution [1]. Most of the existing studies are focused on niche in terrestrial ecosystems, such as plants [2,3,4,5], insects [6,7,8] and woodlands [9]. Additionally, only a few studies look at niche in an aquatic ecosystem, most of which focus on macrobenthos to consider and evaluate the effects of disturbance and pollution on aquatic resources [10,11,12]. Niche breadth can reflect the ability of species to use environmental resources, and niche overlap can reflect the competitive relationship and intensity between species [1,13]. Studying the niche in ecosystems is conducive to exploring the temporal and spatial characteristics of biological communities, and is of great significance for exploring the adaptability of biological communities to the environment and the dynamic monitoring of the number and species of organisms in the environment. With the introduction of ecosystem-based management strategies [14,15], niche studies have provided a reference for fisheries resource management as a very important method of explaining and contemplating fishery resources.
The Lhasa River is located on the Qinghai–Tibet Plateau, the third pole of the world. It is the largest tributary of the Yarlung Zangbo River. The Yarlung Zangbo basin is vast, with an average elevation of more than 4000 m and a vertical drop of about 1800 m [16,17,18]. It is precisely because of the complexity of the geographical location, water flow and climate change in the basin that the fish composition of the Lhasa River is unique and diverse, and the fish grow slowly, have a long sexual maturity cycle, and have low fecundity. Once the fish community is damaged, it is difficult to recover [19,20]. However, with the rapid development of economy in Tibet, human activities have intensified, and overfishing has led to a sharp decrease in the number of native fish. Multiple and repeated introductions have allowed exotic fish to invade and occupy local rivers [21,22]. The construction of hydropower and water conservancy projects has destroyed the original habitat environments of native fish and provided a broader and more suitable living habitat for exotic fish [23]. Now, the fish in the Lhasa River Basin have become smaller, younger, and have been invaded by alien species. The fish resources have been seriously damaged and need to be protected [24,25]. However, there are few reports on the Lhasa River, resulting in a lack of understanding of the existing fish composition and distribution in the Lhasa River and the utilization of resources. In addition, there is a lack of targeted management and protection measures in the conservation of fish resources in the Lhasa River Basin. Understanding the ability of fish to use resources and the environment, and the competition and coexistence among species may provide insights into the conservation of local fish resources and the development of more targeted measures.
In order to explore the ability of the main fish in Lhasa River to utilize environmental resources, and examine the competition and coexistence relationship among species, this study conducted fishery resource surveys in September 2019 (Autumn), June 2020 (Summer), November 2020 (Winter) and March 2021 (Spring). The Index of Relative Importance (IRI) was used to identify the main fish in the catch, and the Shannon index and Pianka’s index were used to analyze the ecological niche breadth and overlap values of the main fish, firstly, in the Lhasa River basin in three dimensions: time, space and space–time. The aim is to carry out research on the spatio-temporal ecological position of major fish species in the Lhasa and analyze the use of spatio-temporal resources by each major fish species, providing information for the conservation and dynamic monitoring of fish diversity in the Lhasa River basin, as well as scientific research support for the conservation management and sustainable development of fishery resources.
2. Materials and Methods
2.1. Data Sources
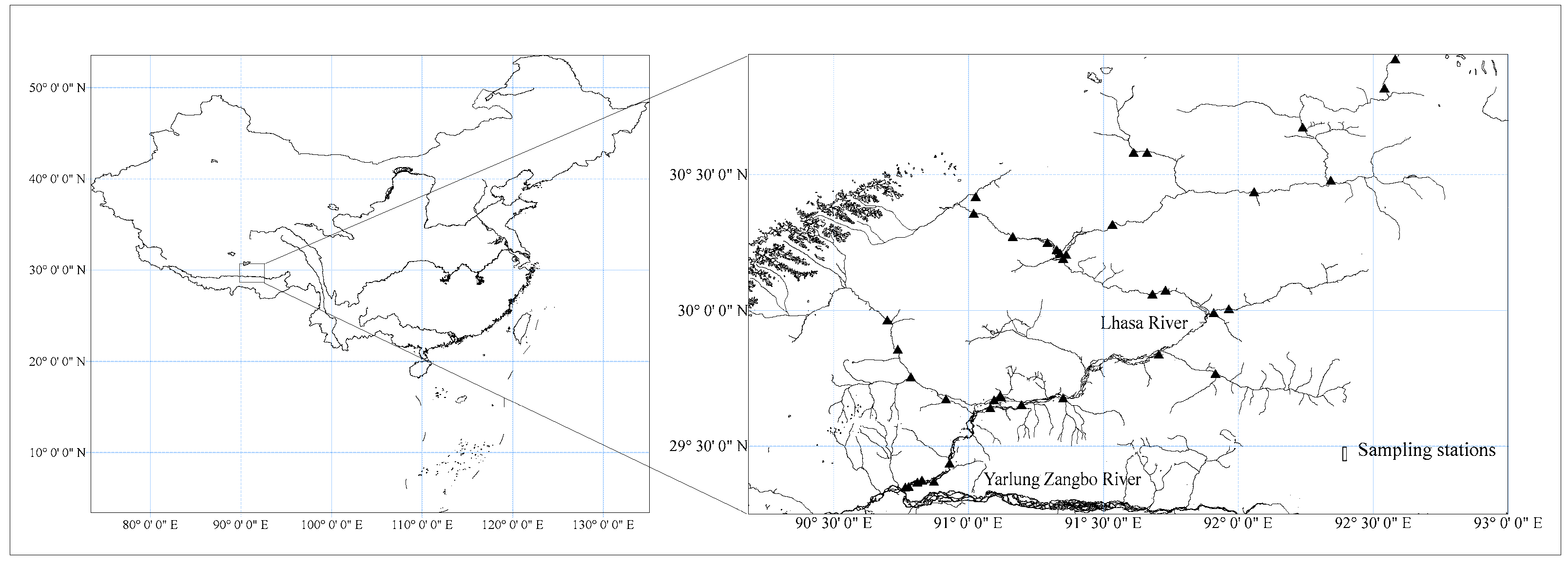
Field work was approved and supervised by the Agriculture and Rural Affairs Department of Tibet Autonomous Region. Data used in this study were derived from four fishery resource surveys (September 2019 (Autumn), June 2020 (Summer), November 2020 (Winter) and March 2021 (Spring)). According to the habitat characteristics of Lhasa River and previous survey data (Supplementary materials), a total of 39 sampling sites were set up (Figure 1), including the mainstream of Lhasa River, 7 tributaries and 2 wetlands. In order to make the catches representative, three-layer gill nets (for pelagic fish) and cages (for demersal fish) were used to catch fish in the Lhasa River Basin (gill nets: length × height: 20 m × 1.2 m, mesh size: 2–8 cm; cage size: length × width × height: 10 m × 0.3 m × 0.5 m), and the placement time was 12 h. The fish resources were investigated in strict accordance with the Inland Waters Fishery Natural Resources Survey Manual [26]. The catches captured at each site were quickly anesthetized, packed into sample bags, numbered and recorded. Classification and identification were carried out according to the literature, such as Chinese Fish System Search [27], Tibetan Plateau Fish [28]. The body length and weight of each fish were measured and recorded with a vernier caliper (0.1 mm) and an electronic balance (0.1 g).
2.2. Research Methods
The dominant species in the fish community were measured using the formula of the IRI [29]:
where N is the proportion of the number of individuals of a certain fish to the total number of fish surveyed, W is the proportion of the weight of a certain fish to the total weight of fish, and F is the proportion of the number of stations of a certain fish in the survey to the total number of stations. The division of dominant species and common species of fish was based on the standard of Cungen Yu et al. [30]. The IRI of dominant species is greater than 1000, and the IRI of common species ranges from 100 to 1000. According to the research of Zhenhai Yu [31] and He Xiongbo [32], the dominant species and common species were regarded as the main species of the community.
Calculation of species niche breadth using Shannon index [33]:
Calculation of species niche overlap using Pianka’s index [34]:
where is niche breadth; is the number of stations; is the proportion of individual number of species in resource state to the total number of species; is the proportion of individual number of species in resource state to the total number of species; the value range is [0, ]. According to the niche breadth, fish can be divided into three types: narrow niche species (0 ≤ < 1.00), medium niche species (1.0 ≤ < 2.0) and wide niche species (2.0 ≤ ). represents the niche overlap value, and the value range is [0, 1]. The larger the value is, the greater the degree of niche overlap is. > 0.3 is meaningful for overlap, and > 0.6 is significant overlap [35].
May [36] proposed that the eigenvalues of multidimensional niches can be expressed by calculating the product of the eigenvalues of different dimensions:
2.3. Data Processing
Surfer 11.0 was used to draw the survey station map of Lhasa River; the IRI was calculated with Microsoft Excel 2021; the niche breadth and niche overlap were calculated using the ‘spaa’ package of R 4.1.0; the similarity matrix of species niche breadth was constructed by Primer v7. Cluster analysis was performed using Bray–Curtis similarity coefficient.
3. Results
3.1. Composition of Main Fish in the Lhasa River Basin and Its Subsidiary Wetlands
Relevant research records show that there are 15 species of native fish in the Lhasa River Basin, mainly including 8 species of Schizothoracinae, 6 species of Triplophysa and 1 species of Sisoridae. The captured alien species, as recorded, are Carassius auratus(C. auratus), Cyprinus carpio, Pseudorasbora parva and other 14 species; there are also two indeterminate species [24,25]. In this study, a total of 1399 fish were captured. They belonged to three orders: Cypriniformes, Siluriformes, Perciformes; five families: Cyprinidae, Cobitidae, Odontobutidae, Siluridae, Sisoridae; and a total of twenty-five species. According to the calculated value of the IRI of all fishes caught, it is concluded that there are six main fish species in the catch of Lhasa River Basin (Table 1), among which the dominant species is Schizopygopsis younghusbandi (S. younghusbandi), and the common species are Ptychobarbus dipogon (P. dipogon), Schizothorax waltoni (Schizothorax waltoni), Schizothorax oconnori (S. oconnori), Triplophysa stenura (T. stenura), C. auratus.
3.2. Niche Breadth of the Main Fish Species
The niche breadth of the main fish species in the Lhasa River Basin fluctuates in the range of 0.780–1.248 in the temporal dimension. Among which, three species have a medium niche. They are C. auratus (1.248), S. younghusbandi (1.091), S. waltoni (1.053) and the rest are narrow niche species (Table 1).
The spatial niche breadth ranges from 1.324 to 3.326, among which three are the species with broad niche. The largest one is S. younghusbandi (3.326), followed by P. dipogon (2.230) and T. stenura (2.188). The remaining three species have a spatial niche width greater than 1 and less than 2. Hence, they are the species with medium niche.
The spatio-temporal niche breadth fluctuates in the range of 1.395–3.628, among which the fish with the largest niche breadth is S. younghusbandi (3.628), and the remaining five species were species with medium niche (1 ≤ niche breadth < 2). Among them, the species with the smallest niche breadth is S. waltoni (1.395), while the spatio-temporal niche breadth of S. oconnori, P. dipogon and C. auratus reaches more than 1.9.
In terms of the three niche indexes, the time niche breadth of C. auratus is the largest, and that of T. stenura is the smallest. The species with the largest spatial and spatio-temporal niche breadth is S. younghusbandi and the one with smallest value is S. waltoni.
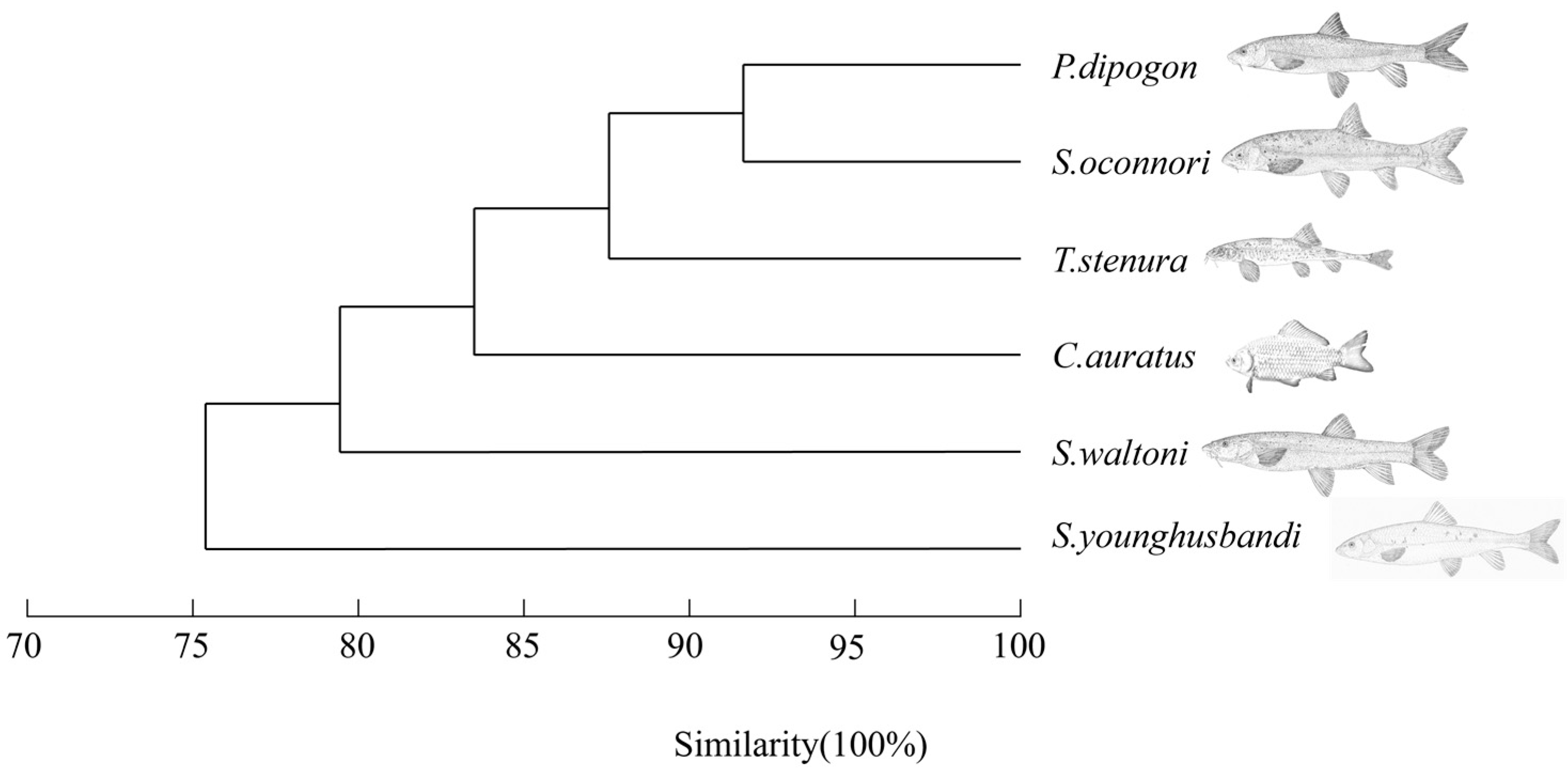
The cluster analysis of temporal, spatial and spatio-temporal niche indices shows that the main fish species in Lhasa River basin are clustered into five groups (Figure 2). Group 1 consists of P. dipogon and S. oconnori; group 2 consists of T. stenura; group 3 consists of C. auratus; group 4 consists of S. waltoni; and group 5 consists of S. younghusbandi. In general, the species in group 1 are similar in temporal, spatial and spatio-temporal dimensions. They were clustered into one group with group 2, then group 3 and group 4, and finally group 5 with the highest niche breadth. It can be seen that the closer the niche species, the higher the similarity.
3.3. Niche Overlap of Main Fish Species
The niche overlap values of the main fish species in the Lhasa River Basin fluctuate in the range of 0.107–0.978 in the temporal dimension (Table 2). There are 12 pairs of species whose niche overlap values reach a significant level (i.e., greater than 0.300), accounting for 80% of the total number of species pairs. Among them, nine pairs of species have an overlap value greater than 0.600, accounting for 60% of the total number of species pairs. The species pair with highest overlap value is S. younghusbandi–S. oconnori, reaching 0.978, followed by S. oconnori–P. dipogon (0.958), S. younghusbandi–P. dipogon (0.957). On the whole, their species pairs achieve meaningful overlap. There are three species pairs with an overlap value less than 0.300, accounting for 20% of the total species pairs. The species with the smallest overlap value is P. dipogon–T. stenura.
The spatial niche overlap values between various pairs range from 0 to 0.559. There are three species pairs with overlap values greater than 0.300, accounting for 20% of the total species pairs. No species pairs have overlap values greater than 0.600. In general, the proportion of species pairs with meaningful overlap was small. The species pair with the highest overlap value is S. younghusbandi–P. dipogon (0.559); the species pairs with the smallest overlap values (0) are composed of P. dipogon with other two species.
The spatio-temporal niche overlap values of the main fish caught in the Lhasa River Basin (Table 3) ranges from 0 to 0.535. Among them, there are three species pairs that achieve meaningful overlap, accounting for 20% of the total species pairs, and there is no significant overlap. There are 12 species pairs with insignificant overlap values (overlap value < 0.3), accounting for 80% of the total species pairs. Among them, there are three species pairs with the smallest overlap values. They are S. oconnori–C. auratus (0.003), P. dipogon–T. stenura (0.001) and P. dipogon–C. auratus (0).
4. Discussion
4.1. Niche Breadth Analysis of Fish Community in Lhasa River Basin and Its Subsidiary Wetlands
Niche breadth is an indicator of the diversity of resources utilized by the population, and its size represents the environmental adaptability and resource utilization ability of the population [1]. The size of the temporal niche breadth value reflects the length of the time when the species appeared. When the temporal niche breadth value is large, this indicates that the species are evenly distributed in the time series. On the contrary, if the niche breadth value is small, its distribution in the time series is fluctuating, and there are obvious low and high peaks. The time niche breadth of C. auratus, S. younghusbandi and S. waltoni is large, which indicates that they have a long activity cycle in the Lhasa River basin. S. younghusbandi and S. waltoni, as important native fish in Tibet, are mainly found in the middle and upper reaches of the Yarlung Zangbo River in Tibet. They are endemic to China and are suitable for growing and developing in areas with high altitudes and wide temperature fluctuations [37]. The surveyed basin is widely spread, and covers the whole Lhasa River basin. There are S. younghusbandi in four seasons and S. waltoni in three seasons, which is related to their adaptability to altitude and temperature changes. It also shows that the characteristics of water environment resources of Lhasa River are more suitable for the growth and development of S. younghusbandi and S. waltoni. Among them, C. auratus, as an invasive species, likes a hydrostatic environment and is mainly found in wetlands and the lower reaches of the Lhasa River. Because of its strong adaptability, high tolerance to various dissolved oxygen and water temperature environments, and diversity of spawning grounds, it can be caught all year round [38,39]. Other species appear in a short period of time, only in some seasons surveyed, and the temporal niche is small. It may be related to the living habits of the population, such as feeding, migration and spawning. Because water temperature is one of the important factors affecting the change in fish behavior [40], the temperature change during the four seasons of a year has a direct impact on fish migration, location transfer of fishing ground, cluster time and length. The niche breadth of fish in the temporal dimension in the living basin may be related to the feeding, migration, spawning and other living habits of the population. In this study, S. younghusbandi and C. auratus appear in four seasons, but the temporal niche breadth value of S. younghusbandi is relatively small, indicating that the distribution of S. younghusbandi is uneven in time series, and there is a phenomenon of seasonal biomass surge. This phenomenon may be related to the fact that S. younghusbandi overwinters in the Yarlung Zangbo River and migrates to the Lhasa River for reproduction in spring [24,41,42,43].
Spatial niche breadth can reflect the distribution range and uniformity of species in the spatial dimension [31]. In this study, the species with largest spatial niche breadth is S. younghusbandi, indicating that its spatial distribution in the Lhasa River basin is the most extensive. The niche breadth of S. waltoni is the smallest, indicating that it exists in some areas of the Lhasa River Basin, and its ability to use space resources is lower than other main fish. If you want to protect both, you should carry out different protection strategies. For example, the protection of S. waltoni should focus on regional areas.
The two-dimensional niche of space and time can reflect the utilization of temporal and spatial resources and the competitive coexistence among different species [44]. Among the main fish species collected in this study, the niche breadth values in spatial latitude are quite different (mean ± standard deviation: 2.109 ± 0.692), but there is no significant difference in temporal latitude (mean ± standard deviation: 1.003 ± 0.165). Therefore, the difference in the utilization of space resources has a great influence on the utilization level and interspecific competition among fishes for space-time resources. This reflects the impact of spatial variation on fish biomass in the surveyed waters. This can also provide us with an idea for protecting these fish, and can be biased towards regional protection of different fish, especially the protection of the main living environment.
Based on the clustering results, the main catches in the Lhasa River Basin were clustered into five groups. The temporal niche breadth of the three fishes in group 1 and group 2 is small, but the spatial niche breadth is large, the spatial distribution is wide, and the seasonal distribution uniformity is low, which is greatly affected by seasonal changes. Group 3 and Group 4 fish have the middle ecological niche species, and have strong utilizing ability in time and space. The fifth group of fish includes the fish with the largest niche breadth in space–time and space among all fish. These fish have a strong ability to utilize time and space resources.
4.2. Analysis of Overlapping Temporal, Spatial and Spatio-Temporal Niches
Niche overlap reflects the similarity of resource utilization among different species and the potential competitive relationship. In the temporal dimension, the overlap value between most fish reaches a meaningful level, which reflects that the main fish caught in the Lhasa River Basin in this survey have a high degree of synchronization in the utilization of Lhasa River resources in time. The Lhasa River is a branch river in the middle reaches of the Yarlung Zangbo River. It is an important place for fish spawning, reproduction and fattening [45]. The seasonal changes of its main caught fish are obvious. Many species migrate to the Lhasa River for spawning, reproduction and fattening in spring and summer when the water temperature is warm. In winter, when the water gets cold, some species swim to the Yarlung Zangbo River [41,43]. Among them, the overlap value between the species pairs of S. younghusbandi–S. oconnori, S. oconnori–P. dipogon and S. younghusbandi–P. dipogon reaches more than 0.9. However, S. oconnori and P. dipogon do not appear in winter, which may be related to their overwintering in the Yarlung Zangbo River [41,43,46]. What the data of the main fish species investigated in this study show are consistent with the fact that the catches of most fish species in summer and autumn are higher than those in winter and spring, resulting in generally high temporal niche overlaps of the main fish species. S. younghusbandi appears in spring, summer, autumn and winter. Only in autumn and winter, the catch was less, which was opposite to that of T. stenura; therefore, the competition for time resources between species pairs is not strong, while the coexistence relationship between species is strong. However, S. waltoni, S. oconnori, and P. dipogon all appear in spring, summer and autumn, so the time niche overlap between species pairs is higher, and the proportion of simultaneous occurrence is large, which has potential competition for time resources. This also shows that temporal niche overlap can reflect the similarity of species life histories in ecological communities.
In the spatial dimension, the niche overlap value of the main fish species in the Lhasa River is generally low, and only 20% of the overlap value exceeds 0.300, indicating that the interspecific competition is weak. The main reasons primarily lie in the large area of this survey, the small number of sampling points, and the inability to obtain sufficient data. What is more, the large altitude difference in the Lhasa River Basin, and the more complicated waters are contributing to this. Different fishes require different geographic environments for survival, and their ability to compete for spatial resources is strongly related to their feeding habits and living habits. The results show that the spatial niche overlap value of C. auratus with other native fish does not reach a meaningful level, revealing that C. auratus does not have a high degree of synchronization in the use of space resources. However, this does not mean that crucian carp will not affect native fish. According to the results of this survey and previous studies, C. auratus mostly exists in the lower reaches of the Lhasa River and affiliated wetlands, and prefers a still water environment. In this area, it is highly competitive with native fish [37,39]. For a specific region, the analysis of and study on the competition or coexistence relationship among the three species pairs may have different results.
If there are species pairs with niche overlap in multiple dimensions, there will be a competitive relationship, and the multidimensional niche overlap value can be used as an index of competition degree. Compared with the single overlap index, the spatio-temporal dimensional niche overlap index takes into account the similarity of fish distribution in time and space dimensions, so it can better reflect the utilization or degree of competition for resources among fish species in the Lhasa River [1]. In this study, the species pairs with significant spatial and temporal niche overlap values are S. younghusbandi–P. dipogon, S. younghusbandi–S. oconnori, S. waltoni–S. oconnori. The overlap value of S. younghusbandi–P. dipogon reaches 0.54, indicating that the two species have the same utilization trend of temporal and spatial resources, and there is a certain interspecific competition or coexistence relationship. The temporal and spatial overlap values of P. dipogon–T. stenura, P. dipogon–C. auratus and S. oconnori–C. auratus are 0, 0.001 and 0.003, respectively. This shows that the demand for the sharing of space–time resources is not high, and the potential competitive relationship is not strong. Among them, the spatial and temporal niche overlap between S. younghusbandi, S. oconnori, C. auratus and other species, or species pairs composed of other species, is extremely high or extremely low. This is because the spatial variation of these types is more obvious, and the two may or may not appear at the same time in a certain area, resulting in this overlapping extreme value.
4.3. Implications of Niche Analysis on Fishery Resource Conservation
Dynamic monitoring of the species niche has far-reaching implications for species conservation, ecological stability and sustainable development of fishery resources. The results of this study show that the main fish in the survey area are S. younghusbandi, P. dipogon, S. waltoni, S. oconnori, T. stenura and C. auratus. According to the information reflected by the niche, the niche breadth of the main fish is greatly affected by space, indicating that the geographical areas of different fish are different. Some fish can be protected from their main habitats, ‘spawning grounds, feeding grounds, and overwintering grounds’. Additionally, the temporal, spatial and spatio-temporal niche breadth values of crucian carp are relatively high. As an invasive species, it competes for resources with native fish from common species such as T. stenura and S. waltoni, to dominant species, S. younghusbandi. Based on the sampling information and previous studies [47], alien species are mainly distributed in the downstream area. So, protecting these native fish, especially in the downstream area, and reducing the invasion of alien species, may directly or indirectly increase the amount of resources and catch production.
This paper analyzes the temporal, spatial and spatio-temporal niche breadth and overlap of the main fish in the Lhasa River, and discusses the ability of the main fish to utilize environmental resources and interspecific competition for the first time. Multi-dimensional niche analysis can explain the utilization of temporal and spatial resources by species in an ecosystem during a certain period of time, and the competitive relationship and potential among species, thus providing a reference for the management, conservation and sustainable development of fishery resources. Continuing the ecological niche survey, increasing the number of sampling sites in space, expanding the dimension of time, and conducting an inter-annual survey can render a more in-depth and more accurate understanding of the temporal and spatial niche changes of the main fish in the Lhasa River. It provides a certain perspective for a more detailed understanding of the changes in the ecosystem of the Lhasa River Basin, and provides ideas for formulating fishery resource management strategies.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/w15050860/s1.
Author Contributions
Software, L.Z., Y.L. and J.Z.; validation, C.Z. and H.L.; investigation, Q.W., Y.Z. and F.L.; data curation, H.L. and C.Z.; writing—original draft preparation, H.G. and Y.D.; writing—review and editing, H.G., J.L., S.F., H.L. and C.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (32160864).
Institutional Review Board Statement
Field work was approved and supervised by the Agriculture and Rural Affairs Department of Tibet Autonomous Region.
Data Availability Statement
Numerical results reported in this paper may be shared by the interested parties if requested. Please contact the author.
Acknowledgments
The authors would like to gratefully thank the agency (Agriculture and Rural Department of Tibet Autonomous Region, Lhasa) for supporting the sampling.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Lin, K.M.; Guo, Y.H. The Research Advances on Niche Theory and Its Application. J. Fujian Coll. For. 2001, 21, 283–287. [Google Scholar]
- Zhang, J.Y.; Zhao, H.L.; Zhang, T.H.; Zhao, X.Y. Niche Dynamics of Main Populations of Plants Communities in the Restoring Succession Process in Horqin Sandy Land. Acta Ecol. Sin. 2003, 12, 2741–2746. [Google Scholar]
- Zheng, Y.R. Main Woody Species Niche of Plant Community in Daqinggou. Acta Phytoecol. Sin. 1999, 23, 475–479. [Google Scholar]
- Sun, H.B.; Gan, X.; Zhao, Q.; Li, Y.F.; Zhang, W.; Liu, X.J.; Liu, M.Y.; Zhang, J. Niche Characteristics of Dominant Species in Forest Community of Tiantoushan Nature Reserve in Shenzhen. For. Environ. Sci. 2022, 38, 75–86. [Google Scholar]
- Li, Y.F.; Liu, M.Y.; Gan, X.H.; Huang, F.F.; Zhang, W.Q. Niche Characteristics of the Dominant Species of the Heritiera Littoralis Community in Baguang Wetland Park. Ecol. Environ. Sci. 2020, 29, 2171–2178. [Google Scholar]
- Kang, L.; Chen, Y.L. Teophic Niche of Grasshoppers Within Steppe Ecosystem in Inner Mongolia. Acta Entomol. Sin. 1994, 37, 178–189. [Google Scholar]
- Huang, B.H.; Zou, Y.D.; Bi, S.D.; Li, H.K.; Zhu, Q.L. Characteristics, Dynamics and Niche of Insect Community in Plum Orchard. Chin. J. Appl. Ecol. 2005, 16, 307–312. [Google Scholar]
- Wang, F.; Ju, R.T.; Li, Y.Z. Niche Concept and Its Application in Insect Ecology. Chin. J. Ecol. 2006, 25, 1280–1284. [Google Scholar]
- Sun, Z.W.; Zhao, S.D. Interspecific Association and Correlation Oflime-Broad Leaved Korean Pine Forest on the Northern Slope of Changbai Mountain. Inst. Appl. Ecol. 1996, 7, 1–5. [Google Scholar]
- Ge, B.M.; Bao, Y.X.; Zheng, X.; Cheng, H.Y. The Structure of the Macrobenthic Community and Niche Analysis at a Tidal Flat of Linkun Island. Acta Ecol. Sin. 2005, 25, 3037–3043. [Google Scholar]
- Peng, S.Y.; Li, X.Z.; Wang, H.F.; Zhang, B.L. Niche Analysis of Dominant Species of Macrozoobenthic Community in the Southern Yellow Sea in Spring. Acta Ecol. Sin. 2015, 35, 1917–1928. [Google Scholar]
- Hu, Z.Y.; Bao, Y.X.; Ge, B.M.; Li, H.H. Macrobenthic Community and Niche Analysis of Creeks during Autumn in Diked Tidal Flat. Curr. Zool. 2006, 52, 800–809. [Google Scholar]
- Li, D.Z.; Shi, Q.; Zang, R.G.; Wang, X.P.; Sheng, L.J.; Zhu, Z.L.; Wang, C.A. Models for Niche Breadth and Niche Overlap of Species or Populations. Sci. Silvae Sin. 2006, 42, 95–103. [Google Scholar]
- Ren, H.; Wu, J.G.; Peng, S.L.; Zhao, L.Z. Concept of Ecosystem Management and Its Essential Elements. Chin. J. Appl. Ecol. 2000, 11, 455–458. [Google Scholar]
- Cal, Q.H.; Tang, T.; Liu, J.K. Several Research Hotspots in River Ecology. Chin. J. Appl. Ecol. 2003, 14, 1573–1577. [Google Scholar]
- Zhang, P.; Liu, Y.; An, R.Z.; Qiao, N.Q.; Da, Z.; Ba, S. Spatio-Temporal Niche of Dominant Protozoa Species in the Midstream and Downstream of Lhasa River, Tibet, China. Sci. Silvae Sin. 2022, 58, 78–88. [Google Scholar]
- Qiao, N.Q.; Zhang, P.; Da, Z.; An, R.Z.; Ba, S. Distribution of Functional Groups of Protozoa and Assessment of Water Environment Quality in the Middle and Lower Reaches of the Lhasa River. Wetl. Sci. 2021, 19, 726–742. [Google Scholar]
- Da, Z.; Zhang, P.; An, R.Z.; Pan, C.M.; Qiao, N.Q.; Ba, S. Species Distribution Patterns and Driving Factors of Protozoan Community in the Middle and Lower Reaches of Lhasa River. J. Hydroecology 2022, 4, 1–21. [Google Scholar] [CrossRef]
- Chen, Y.F.; Cao, W.X. Zoology of China, Teleost Fishes: Cyprinids: Part II; Science Press: Beijing, China, 2000. [Google Scholar]
- Tibet Autonomous Region Fisheries Bureau. Tibetan Fish and Their Resources; China Agriculture Press: Beijing, China, 1995. [Google Scholar]
- Li, C.; Fan, L. Influence of Religious Release on Fish Assemblages in the Lhasa River Basin, Tibet, China. J. Plateau Agric. 2018, 2, 462–469. [Google Scholar] [CrossRef]
- Fan, L.; Liu, H. Non-Native Fishes: Distribution and Assemblage Structure in the Lhasa River Basin, Tibet, China. Acta Hydrobiol. Sin. 2016, 40, 958–967. [Google Scholar]
- Ding, H.; Zhang, Z.; Xie, C. Effects of Fish Invasion on Aquatic Ecosystem of the Yarlung Zangbo River and the Prevention and Control Strategies. Chin. J. Ecol. 2022, 41, 2440–2448. [Google Scholar] [CrossRef]
- Chen, F.; Chen, Y.F. Investigation and Protection Strategies of Fishes of Lhsas River. Acta Hydrobiol. Sin. 2010, 34, 278–285. [Google Scholar] [CrossRef]
- Luo, S.; Dan, Z.; Bu, D. Current Status of Fish Resource in Lhasa River and Countermeasure of Its Utilization. J. Tibet Univ. 2011, 26, 7–10. [Google Scholar]
- Zhang, J.M.; He, Z.H. Handbook for the Survey of Natural Resources of Inland Water Fisheries; Agricultural Press: Beijing, China, 1991. [Google Scholar]
- Cheng, Q.T.; Zheng, B.H. Chinese Fish System Search; Science Press: Beijing, China, 1987. [Google Scholar]
- Wu, Y.F.; Wu, C.Z. Fish of the Tibetan Plateau; Sichuan Science and Technology Press: Chengdu, China, 1992. [Google Scholar]
- Pianka, E.R. Ecology of the Agamid Lizard Amphibolurus Isolepis in Western Australia. Copeia 1971, 3, 527–536. [Google Scholar] [CrossRef]
- Liu, K.; Yu, C.G.; Zheng, J.; Xu, Y.J.; Jiang, X.; Yu, N.J.; Zhang, P.Y.; Jiang, Q.L.; Niu, W.Z. Analysis of Functional Groups Characteristics and Niche of Major Fish Species in the Coastal Waters of Zhoushan Islands in Spring and Autumn. J. Zhejiang Univ. 2021, 48, 592–605. [Google Scholar]
- Yu, Z.H.; Jin, X.S.; Li, X.S. Analysis of Ecological Niche for Major Fish Species in the Central and Southern Yellow Sea. Prog. Fish. Sci. 2010, 31, 1–8. [Google Scholar]
- He, X.; Li, J.; Shen, C.; Shi, Y.; Feng, C.; Guo, J.H.; Zhao, C.X.; Yan, Y.R.; Kang, B. The Breadth and Overlap of Ecological Niche of Major Fish Species in the Minjiang River Estuary, China. Chin. J. Appl. Ecol. 2018, 29, 3085–3092. [Google Scholar]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar] [CrossRef] [Green Version]
- Pianka, E.R. The Structure of Lizard Communities. Annu. Rev. Ecol. Syst. 1973, 4, 53–74. [Google Scholar] [CrossRef] [Green Version]
- Charles, K. Ecological Methodology; HarperCollins Publishers: New York, NY, USA, 1984. [Google Scholar] [CrossRef]
- May, R.M. Some Notes on Estimating the Competition Matrix, a. Ecology 1975, 56, 737–741. [Google Scholar] [CrossRef]
- He, L.Q.; Wamg, W.L.; Zeng, B.H.; Yang, R.B.; Liu, H.P.; Zeng, X.L. The Study on the Temperature Tolerance of Diferent Sizes of Schizothorax Waltoni. Acta Hydrobiol. Sin. 2020, 44, 1230–1238. [Google Scholar]
- Chen, P.H. An Ecological Study of Carassius Auratus in Lake Liang-Tze. Acta Hydrobiol. Sin. 1959, 4, 411–419. [Google Scholar]
- Zhang, X.A.; Sui, X.Y.; Lü, Z.; Chen, Y.F. A Prediction of the Global Habitat of Two Invasive Fishes (Pseudorasbora Parva and Carassius Auratus) from East Asia Using Maxent. Biodivers. Sci. 2014, 22, 182–188. [Google Scholar]
- Lv, W.; Yuan, M. The Literature Review of Temperature Change Effect on Fish Behavior. J. Shanghai Ocean Univ. 2017, 26, 828–835. [Google Scholar]
- Ma, W.Z.; An, R.D.; Li, M.; Li, J. Habitat Hydrodynamic Characteristics and Behavior Forcasting for Migratory Fish Downstream of Dams. J. Beijing Norm. Univ. 2021, 57, 433–440. [Google Scholar]
- Duan, Y. Study on the Biology and Population Dynamics of Schizopygopsis Younghusbandi; Huazhong Agricultural University: Wuhan, China, 2015. [Google Scholar]
- Li, L.; Wu, S.; Wang, N. Community Struture of Fishery Resources From the Sangri to Jiacha Reach of Middle Yarlung Zangbo River of Tibet, China. Chin. J. Fish. 2021, 34, 40–45. [Google Scholar]
- Li, R.; Li, S.C.; Tian, R.J. Structure and Niche of Spider Community in Vegetable Field. Acta Arachnol. Sin. 2007, 16, 116–120. [Google Scholar]
- Lv, Y.; Hao, S. Investigation and Analysis on Plankton and Fish Resources in the Source Area of Lhasa River, Tibet, China. Oceanol. Et Limnol. Sin. 2016, 47, 407–413. [Google Scholar]
- Ma, B. Study on the Biology and Population Dynamics of Schizothorax o’connori. Ph.D. Thesis, Huazhong Agricultural University, Wuhan, China, 2011. [Google Scholar]
- Chen, X.; Jin, T.T.; Su, H.D.; Lin, J.Q.; Wang, D.B.; Liu, G.H.; Zhang, J.J. Construction and Application of Health Assessment Index System for Lhasa River. Acta Ecol. Sin. 2019, 39, 799–809. [Google Scholar]
Figure 1.
Distribution of sampling stations.

Figure 2.
Cluster analysis of niche breadth of main fish caught.


{kind=link}
{kind=link}
Table 1.
Niche breadth, IRI and Station number of major species in the Lhasa River.
| Species | Temporal Niche Breadth | Spatial Niche Breadth | Spatio-Temporal Niche Breadth | IRI | Station Number of Occurrence |
|---|---|---|---|---|---|
| Schizopygopsis younghusbandi (S. younghusbandi) | 1.091 | 3.326 | 3.628 | 1.2271 | 38 |
| Schizothorax waltoni (S. waltoni) | 1.053 | 1.324 | 1.395 | 0.0152 | 8 |
| Schizothorax oconnori (S. oconnori) | 0.962 | 1.992 | 1.916 | 0.0203 | 9 |
| Ptychobarbus dipogon (P. dipogon) | 0.885 | 2.230 | 1.974 | 0.0378 | 12 |
| Triplophysa tibetana | 1.034 | 1.890 | 1.954 | 0.0025 | 8 |
| Triplophysa orientalis | 0.517 | 1.155 | 0.597 | 0.0019 | 3 |
| Triplophysa stenura (T. stenura) | 0.780 | 2.188 | 1.707 | 0.0160 | 13 |
| Hypseleotris swinhonis | 0.606 | 1.085 | 0.657 | 0.0009 | 3 |
| Carassius auratus (C. auratus) | 1.248 | 1.594 | 1.989 | 0.0131 | 3 |
| Cyprinus carpio | 0.243 | 1.182 | 0.287 | 0.0047 | 3 |
| Pseudorasbora parva | 0.637 | 1.574 | 1.002 | 0.0090 | 6 |
Table 2.
Temporal and spatial niche overlap of main species in the Lhasa River.
| Species | S. younghusbandi | S. waltoni | S. oconnori | P. dipogon | T. stenura | C. auratus |
|---|---|---|---|---|---|---|
| S. younghusbandi | 0.137 | 0.368 | 0.559 | 0.202 | 0.039 | |
| S. waltoni | 0.704 | 0.508 | 0.035 | 0.043 | 0.055 | |
| S. oconnori | 0.978 | 0.700 | 0.000 | 0.240 | 0.006 | |
| P. dipogon | 0.957 | 0.769 | 0.958 | 0.006 | 0.000 | |
| T. stenura | 0.189 | 0.640 | 0.114 | 0.107 | 0.182 | |
| C. auratus | 0.589 | 0.712 | 0.461 | 0.473 | 0.844 |
Values under the main diagonal are temporal niche overlap parameter; values above the main diagonal are spatial niche overlap parameter.
Table 3.
Spatio-temporal niche overlap of main species in the Lhasa River.
| S. younghusbandi | S. waltoni | S. oconnori | P. dipogon | T. stenura | |
|---|---|---|---|---|---|
| S. waltoni | 0.096 | ||||
| S. oconnori | 0.360 | 0.356 | |||
| P. dipogon | 0.535 | 0.027 | 0.023 | ||
| T. stenura | 0.038 | 0.028 | 0.027 | 0.001 | |
| C. auratus | 0.023 | 0.039 | 0.003 | 0.000 | 0.153 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gao, H.; Liu, H.; Wang, Q.; Liu, F.; Li, J.; Duan, Y.; Zeren, L.; Fu, S.; Zhang, J.; Zhou, Y.; et al. Niche Analysis of the Main Fish in the Lhasa River Basin. Water 2023, 15, 860. https://doi.org/10.3390/w15050860
AMA Style
Gao H, Liu H, Wang Q, Liu F, Li J, Duan Y, Zeren L, Fu S, Zhang J, Zhou Y, et al. Niche Analysis of the Main Fish in the Lhasa River Basin. Water. 2023; 15(5):860. https://doi.org/10.3390/w15050860
Chicago/Turabian StyleGao, He, Haiping Liu, Qiming Wang, Fei Liu, Junting Li, Yuting Duan, Luowu Zeren, Suxing Fu, Jingsen Zhang, Yan Zhou, and et al. 2023. "Niche Analysis of the Main Fish in the Lhasa River Basin" Water 15, no. 5: 860. https://doi.org/10.3390/w15050860
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.