
Atmospheric Exchange of Carbon Dioxide and Water Vapor above a Tropical Sandy Coastal Plain
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
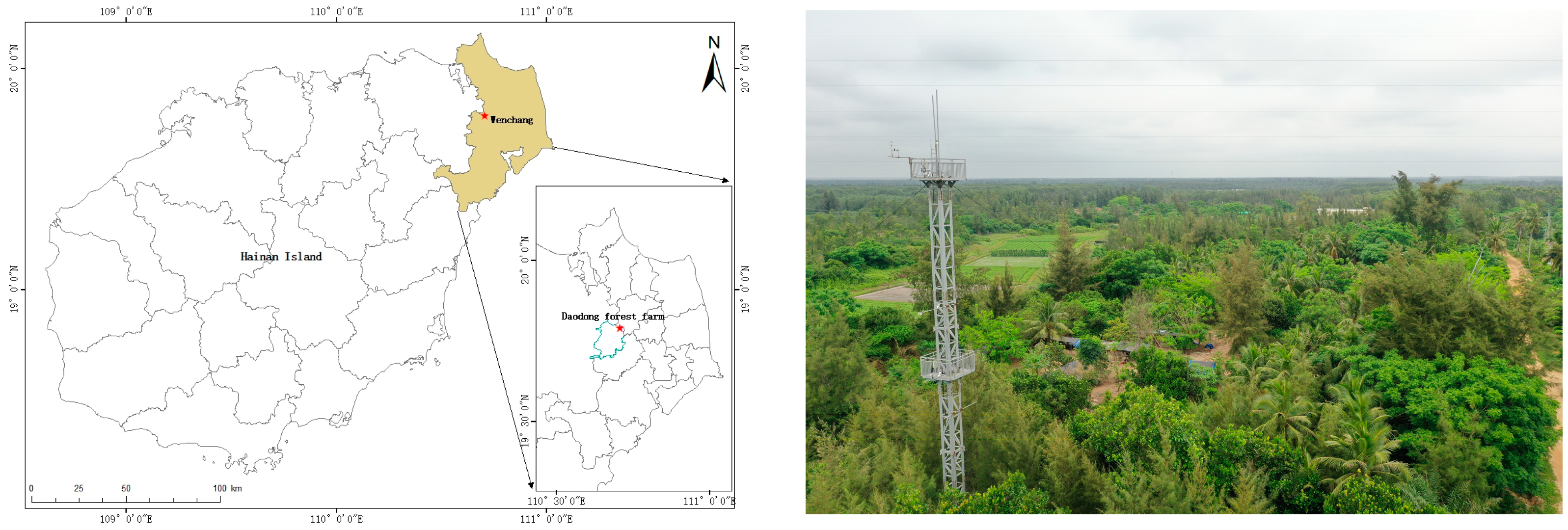
2.1. Site Description
2.2. Instrumentation and Observations
2.3. Eddy Flux Calculation
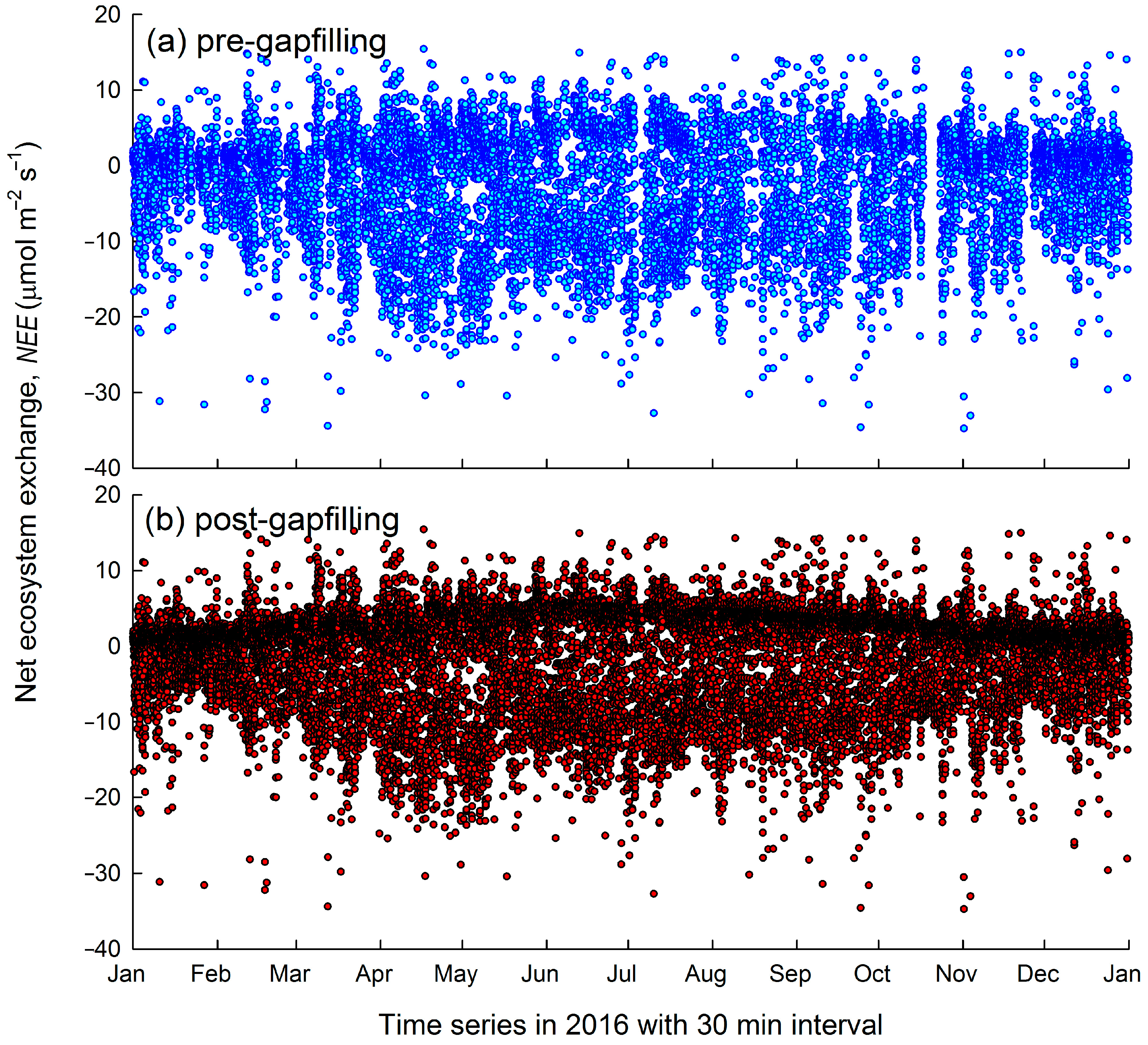
2.4. Processing of Eddy Flux Data
2.5. Calculations and Statistics
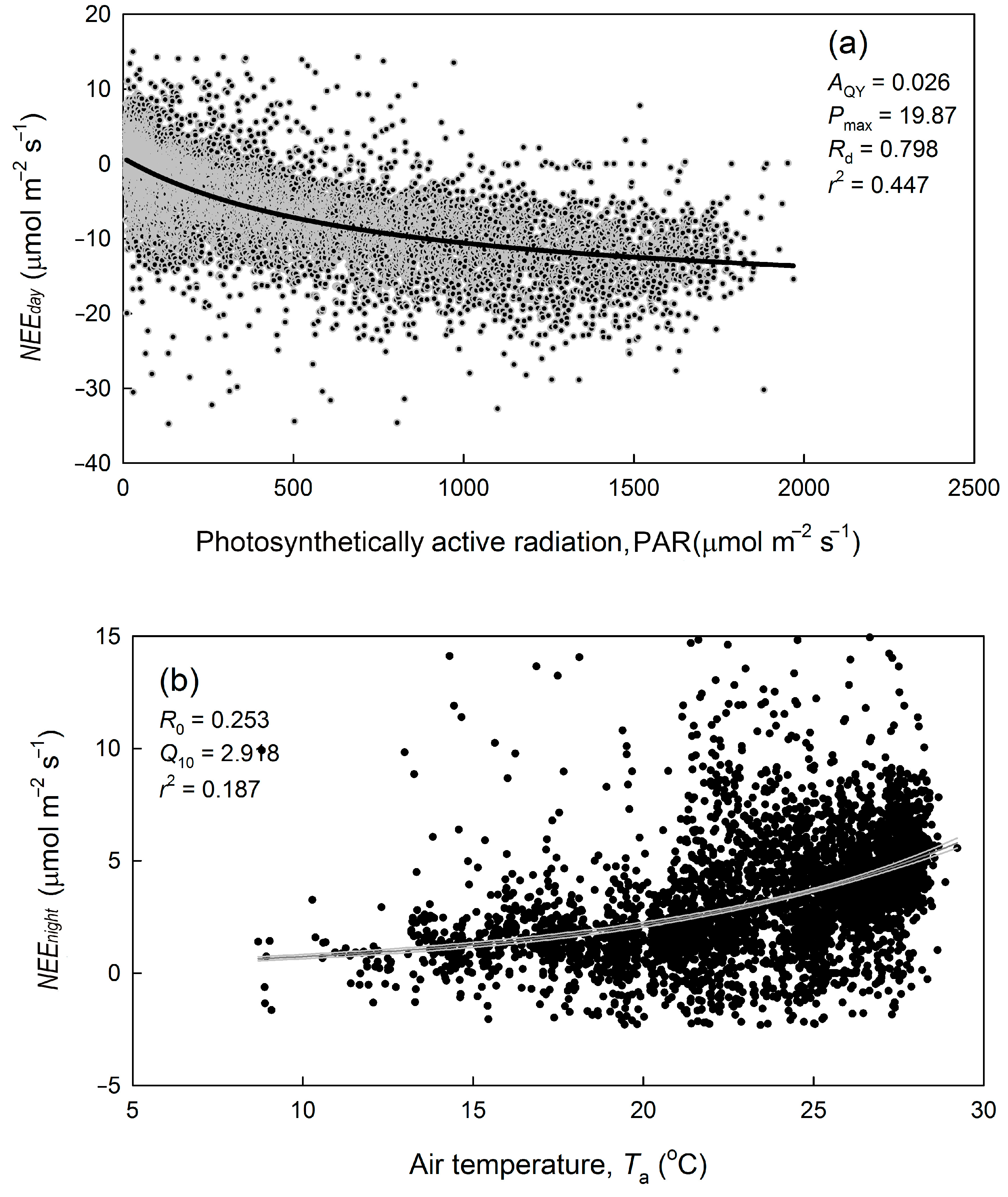
2.6. Net Photosynthetic Rate
3. Results
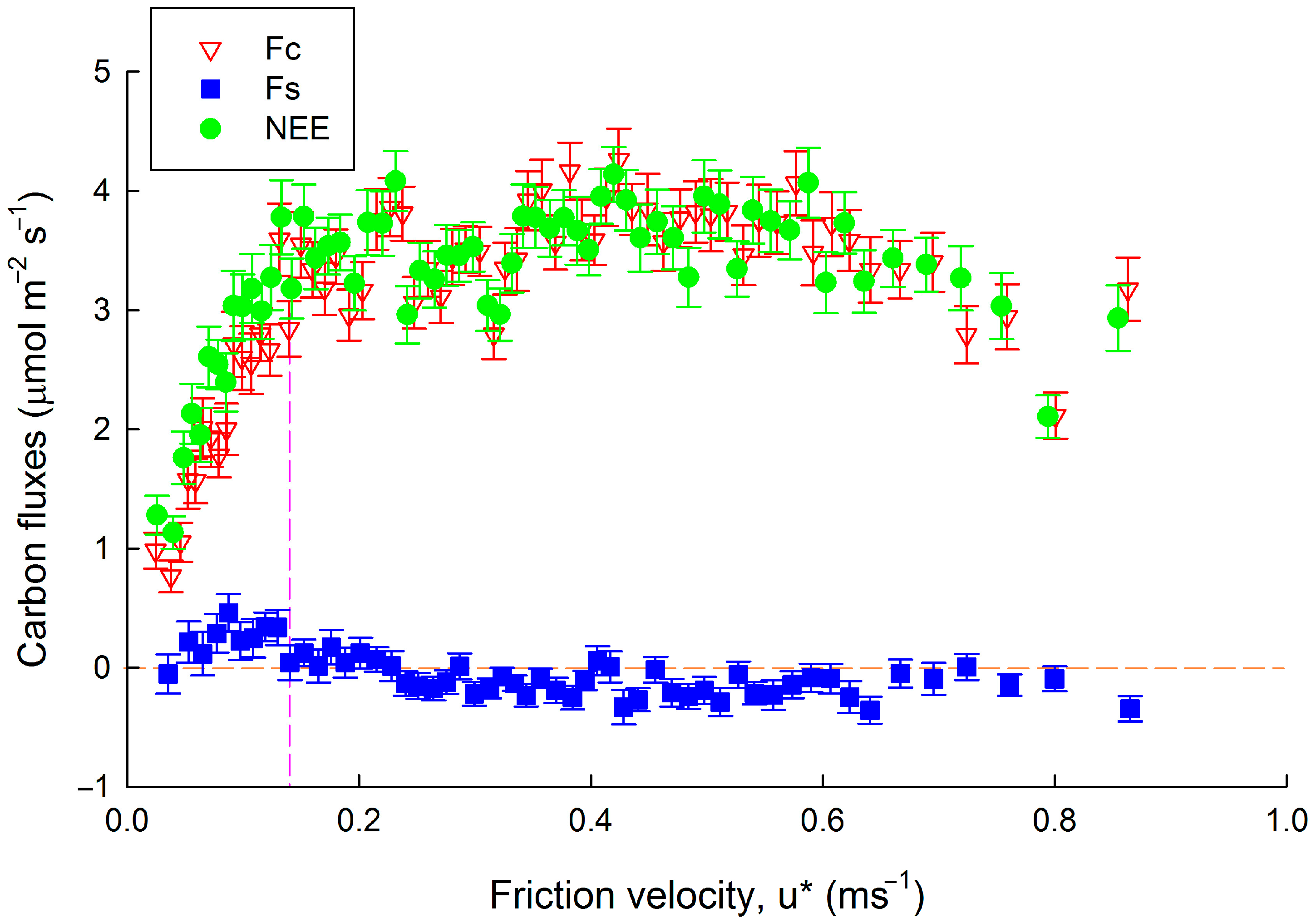
3.1. Nighttime Flux Underestimation
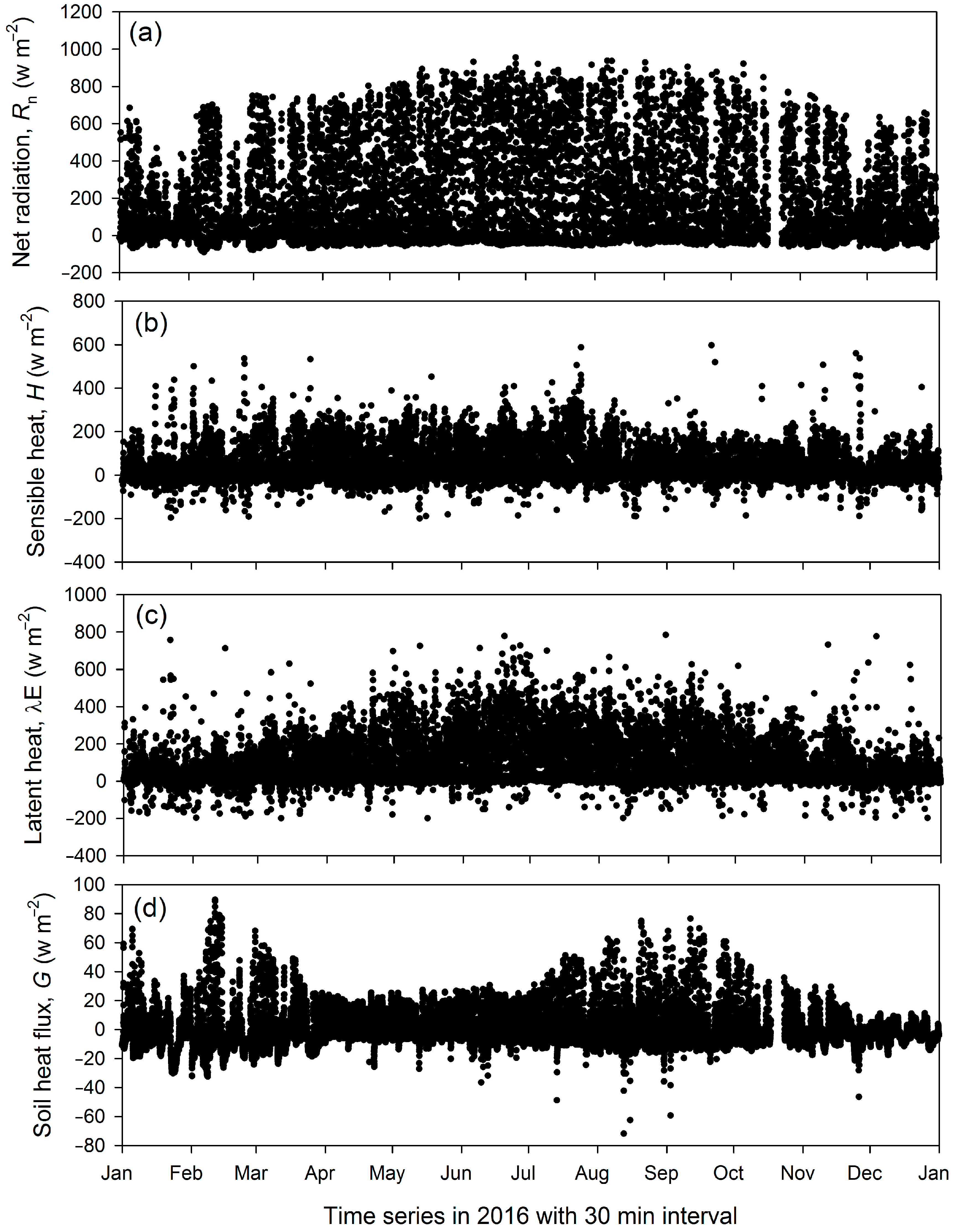
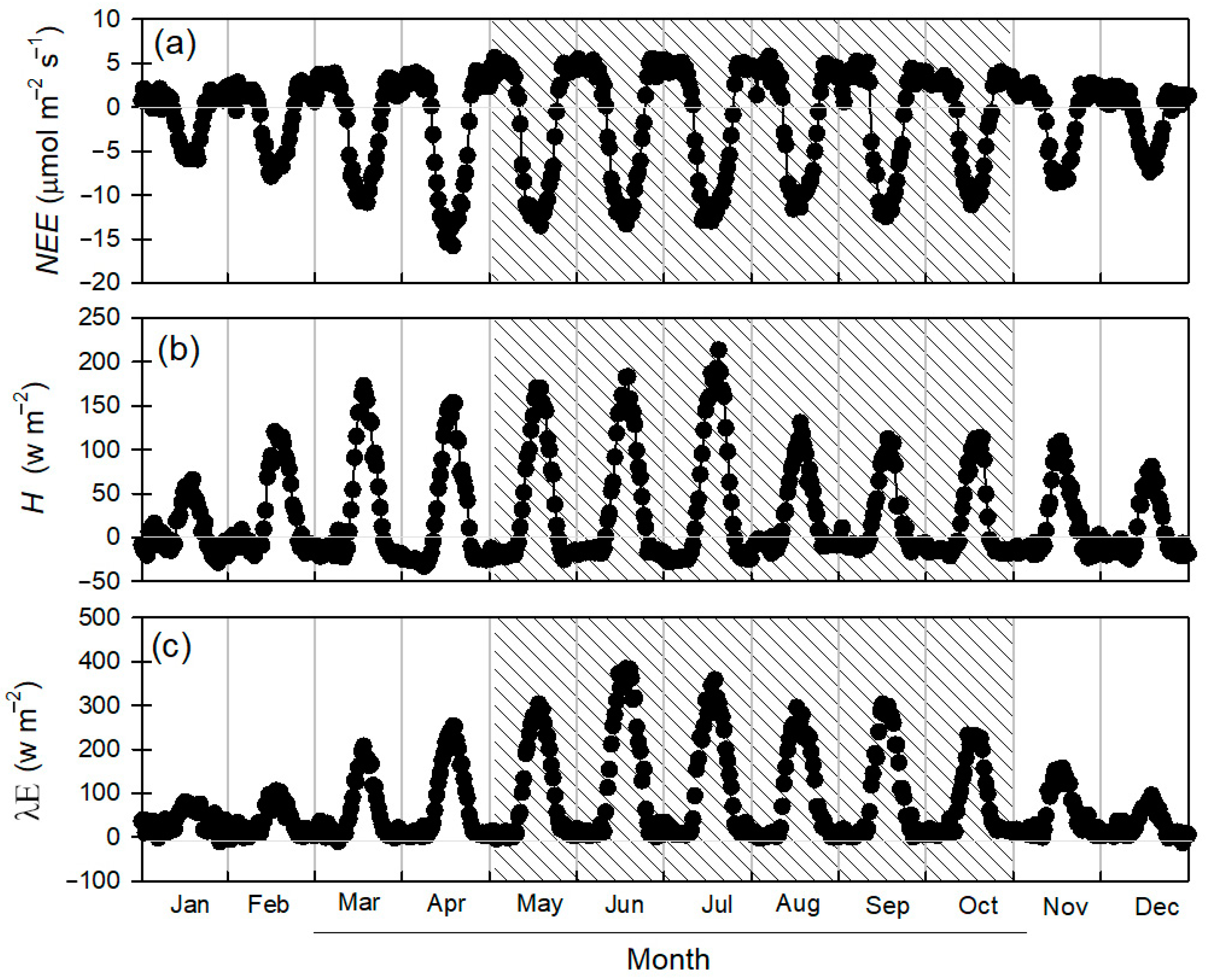
3.2. Energy Fluxes
3.3. Carbon Dioxide Flux and Its Environmental Response
3.4. Annual Sum of Carbon Dioxide and Water Vapor Flux
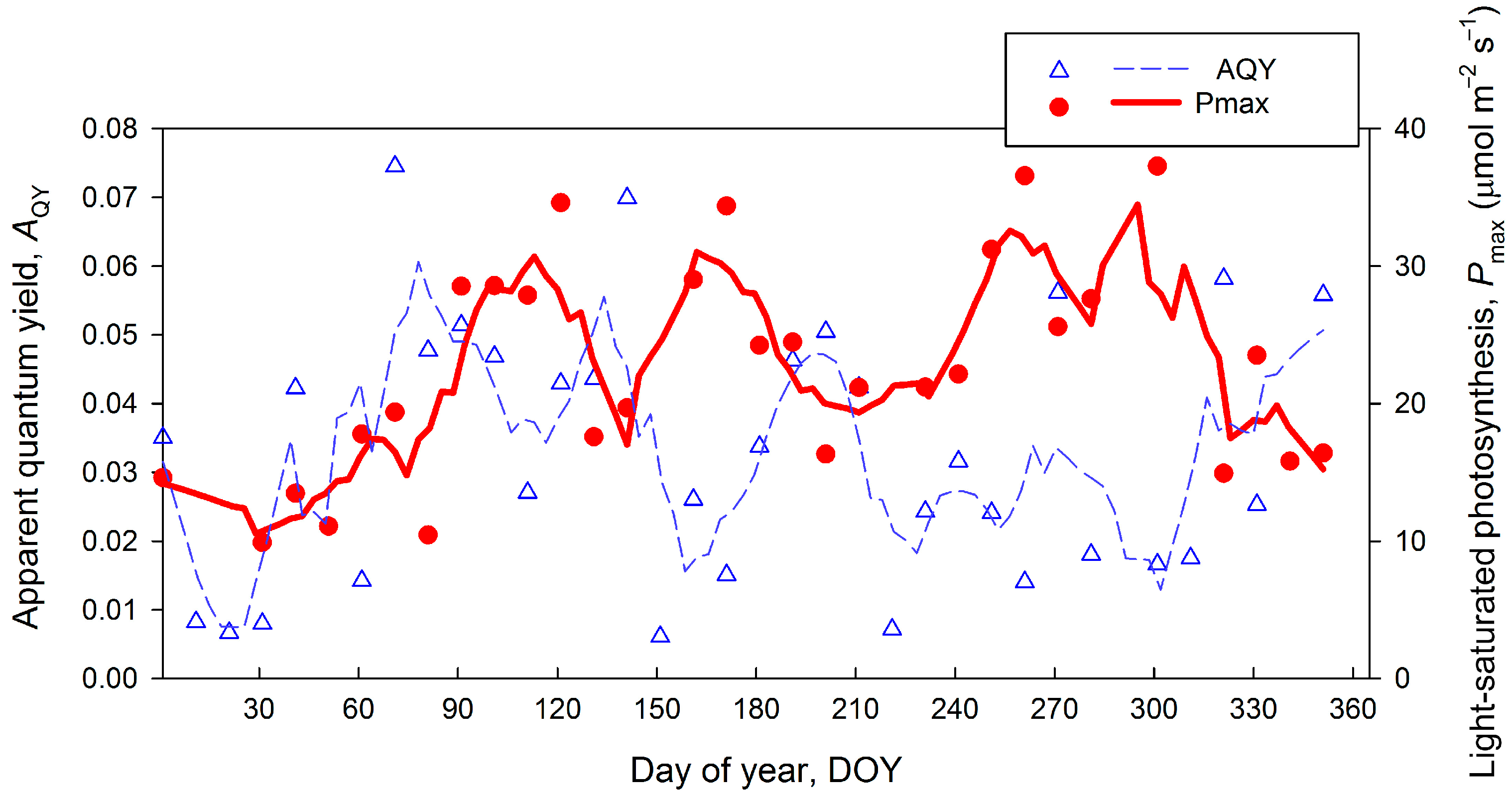
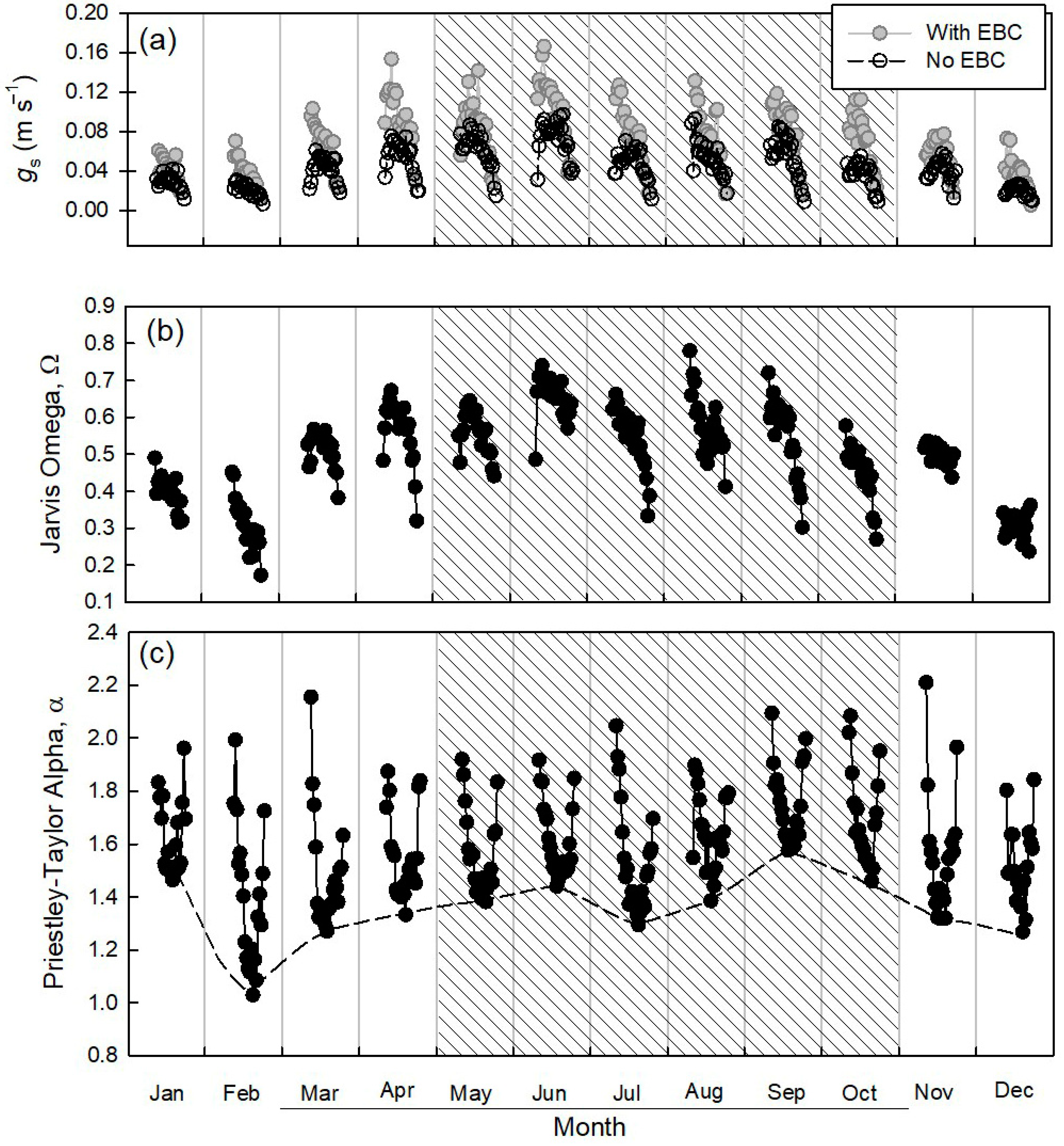
3.5. Variation of the Eddy Flux and Related Bulk Ecosystem Parameters
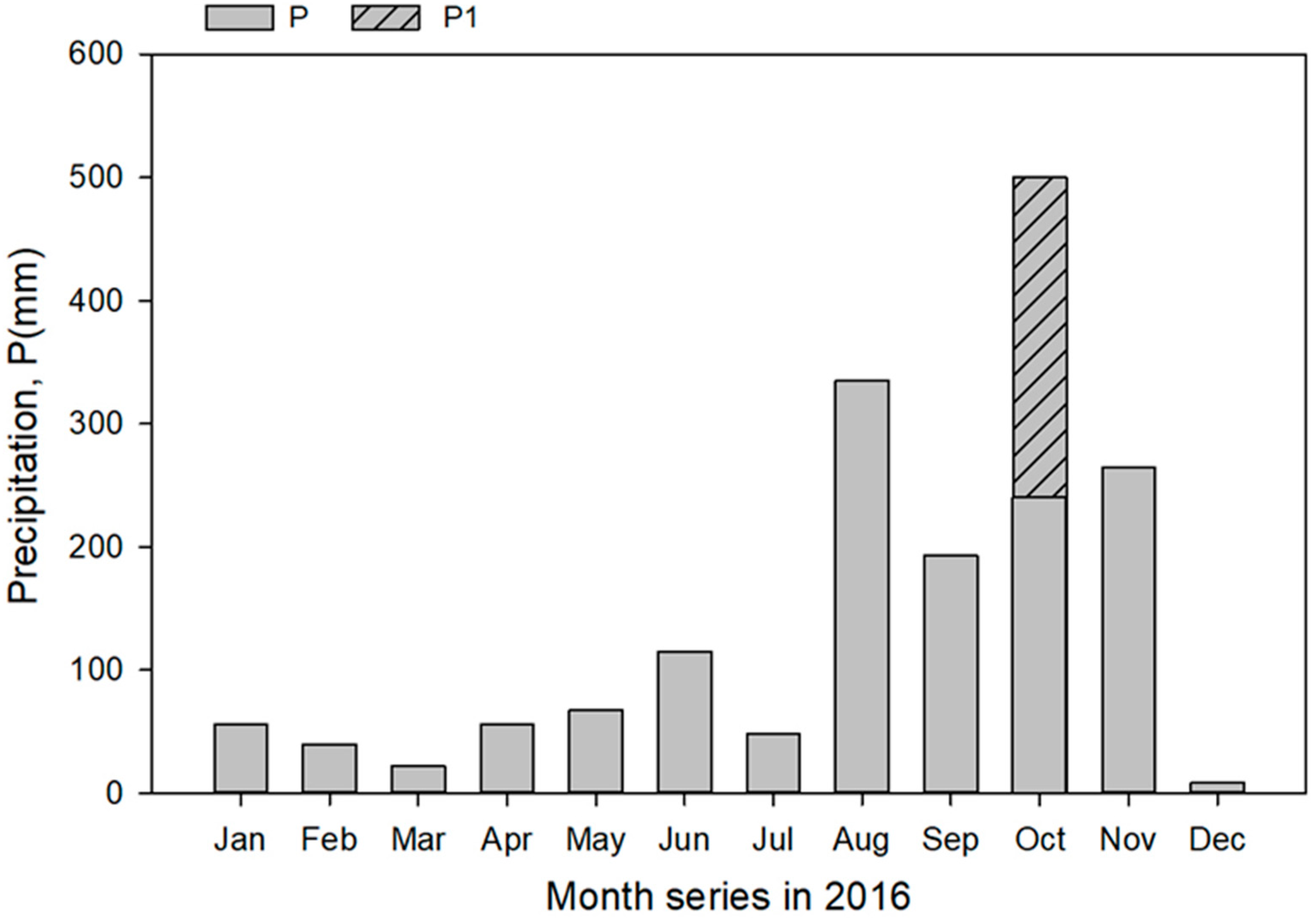
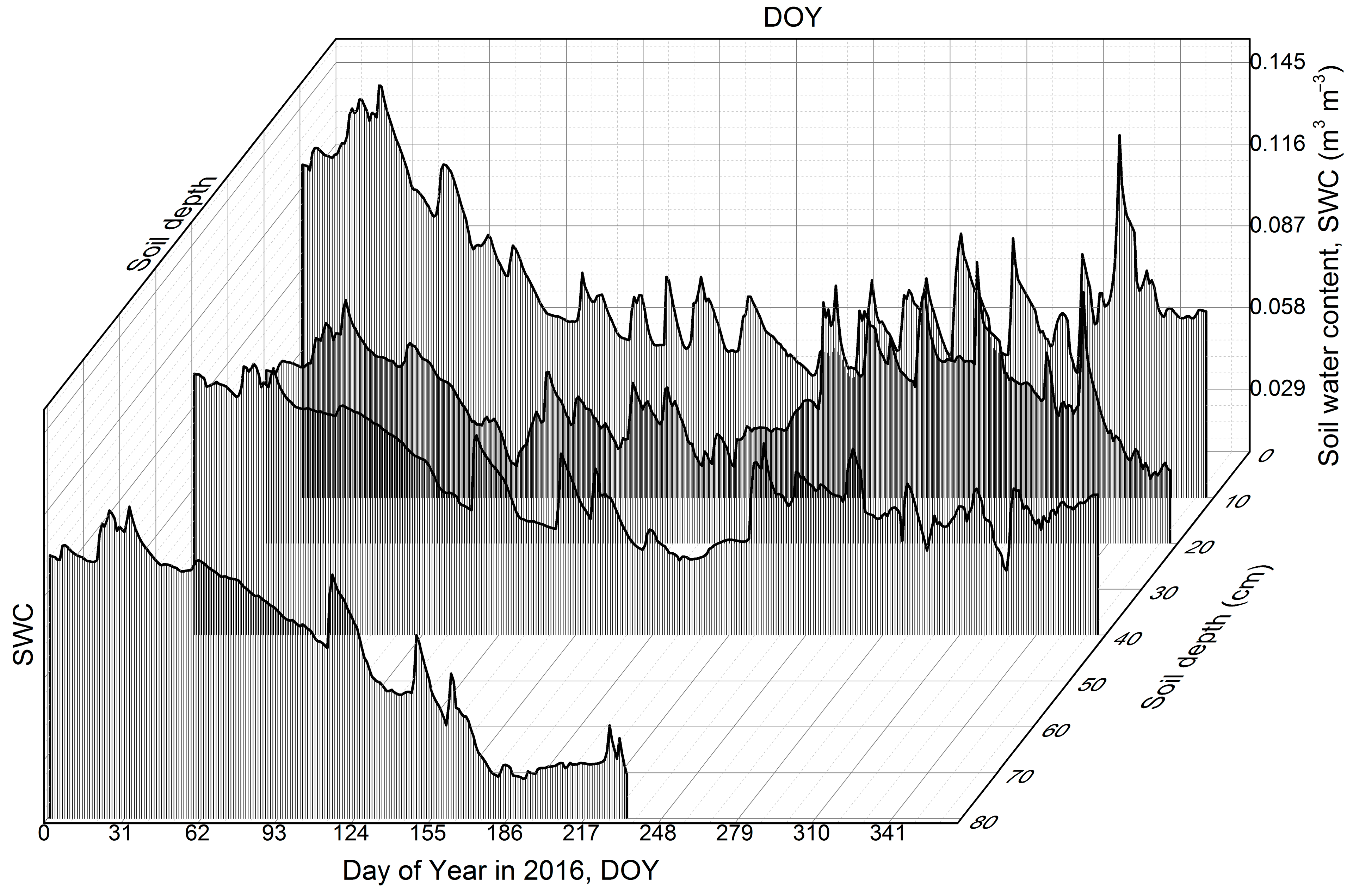
3.6. Sandy Soil Water Content Dynamics
4. Discussion
4.1. Uncertainties of Eddy Covariance Observations at This Study Site
4.2. Physiological Ecosystem Parameters

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest | AQY | Pmax | gs | α | Reference |
|---|---|---|---|---|---|
| Daodong forest farm | 0.026 | 19.87 | 0.015 | 1.477 | |
| Scots pine | 0.027 | 14.5 | [49] | ||
| Mixed oak | 0.033 | 24.54 | [49] | ||
| Boreal forest | 0.040 | 13.1 | 1.22 | [49,55] | |
| Tropical forest | 0.034 | 29.7 | 0.8 | [49,53,54] | |
| Mean global vegetation values | 0.018 | [52] |
4.3. Can the Studied Ecosystem Act as a Persistent Carbon Sink?
4.4. Unexpected Seasonal Soil Water Content Pattern and Its Possible Explanation
4.5. Possible Explanation of the Strongest Carbon Assimilation in the Late Dry Season Shortly before Rainfall Starts and Its Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kondo, M.; Saitoh, T.M.; Sato, H.; Ichii, K. Comprehensive synthesis of spatial variability in carbon flux across monsoon Asian for-ests. Agric. For. Meteorol. 2017, 232, 623–634. [Google Scholar] [CrossRef] [Green Version]
- Hirata, R.; Saigusa, N.; Yamamoto, S.; Ohtani, Y.; Ide, R.; Asanuma, J.; Gamo, M.; Hirano, T.; Kondo, H.; Kosugi, Y.; et al. Spatial distribution of carbon balance in forest ecosystems across East Asia. Agri-Cult. For. Meteorol. 2008, 148, 761–775. [Google Scholar] [CrossRef]
- Jin, Y.; Liu, Y.; Liu, J.; Zhang, X. Energy Balance Closure Problem over a Tropical Seasonal Rainforest in Xishuangbanna, Southwest China: Role of Latent Heat Flux. Water 2022, 14, 395. [Google Scholar] [CrossRef]
- Dennis, B. Measuring fluxes of trace gases and energy between ecosystems and the atmosphere—The state and future of the eddy covariance method. Glob. Chang. Biol. 2015, 20, 3600–3609. [Google Scholar]
- Zeri, M.; Sá, L.; Manzi, A.; Araújo, A.; Aguiar, R.; von Randow, C.; Sampaio, G.; Cardoso, F.; Nobre, C. Variability of carbon and water fluxes following climate extremes over a tropical forest insouthwestern Amazonia. PLoS ONE 2014, 9, e88130. [Google Scholar] [CrossRef] [Green Version]
- Yamanoi, K.; Mizoguchi, Y.; Utsugi, H. Effects of a windthrow disturbance on the carbon balance of a broadleaf deciduous forest in Hokkaido, Japan. Biogeosci. Discuss. 2015, 12, 10425–10468. [Google Scholar] [CrossRef] [Green Version]
- Baldocchi, D.; Chu, H.; Reichstein, M. Inter-annual variability of net and gross ecosystem carbon fluxes: A review. Agricul-Tural For. Meteorol. 2017, 249, 520–538. [Google Scholar] [CrossRef] [Green Version]
- Dunn, A.L.; Barford, C.C.; Wofsy, S.C.; Goulden, M.L.; Daube, B.C. A long-term record of carbon exchange in a boreal black spruce forest: Means, responses to in terannual variability, and decadal trends. Glob. Chang. Biol. 2007, 13, 577–590. [Google Scholar] [CrossRef] [Green Version]
- Ilvesniemi, H.; Levula, J.; Ojansuu, R.; Kolari, P.; Kulmala, L.; Pumpanen, J.; Launiainen, S.; Vesala, T.; Nikinmaa, E. Long-term measurements of the carbon balance of a boreal Scots pine dominated forest ecosystem. Boreal Environ. Res. 2009, 14, 731–753. [Google Scholar]
- Richardson, A.D.; Hollinger, D.Y.; Aber, J.D.; Ollinger, S.V.; Braswell, B.H. Environmental variation is directly responsible for short- but not long-term variation in forest-atmosphere carbon exchange. Glob. Chang. Biol. 2007, 13, 788–803. [Google Scholar] [CrossRef]
- Soloway, A.D.; Amiro, B.D.; Dunn, A.L.; Wofsy, S.C. Carbon neutral or a sink? Uncertainty caused by gap-filling long-term flux measurements for an old-growth boreal black spruce forest. Agric. For. Meteorol. 2017, 233, 110–121. [Google Scholar] [CrossRef]
- Ueyama, M.; Iwata, H.; Harazono, Y. Autumn warming reduces the CO2 sink of a black spruce forest in interior Alaska based on a nine-year eddy covariance mea surement. Glob. Chang. Biol. 2014, 20, 1161–1173. [Google Scholar] [CrossRef] [PubMed]
- Amiro, B.D.; Barr, A.G.; Barr, J.G.; Black, T.A.; Bracho, R.; Brown, M.; Chen, J.; Clark, K.L.; Davis, K.J.; Desai, A.R.; et al. Ecosystem carbon dioxide fluxes after disturbance in forests of North America. J. Geophys. Res. 2010, 115. [Google Scholar] [CrossRef] [Green Version]
- Goulden, M.L.; Mcmillan, A.M.S.; Winston, G.C.; Rocha, A.V.; Manies, K.L.; Harden, J.W.; Bond-Lamberty, B.P. Patterns of NPP, GPP, respiration, and NEP during boreal forest succession. Glob. Chang. Biol. 2011, 17, 855–871. [Google Scholar] [CrossRef] [Green Version]
- Jarvis, P.G.; Leverenz, J. Productivity of temperate, deciduous and evergreen forests. In Encyclopedia of Plant Physiology; Eal, O.L., Ed.; Springer: Berlin/Heidelberg, Germany, 1983. [Google Scholar]
- Bracho, R.; Starr, G.; Gholz, H.L.; Martin, T.A.; Cropper, W.P.; Loescher, H.W. Controls on carbon dynamics by ecosystem structure and climate for southeastern U. S:slash pine plantations. Ecol. Monogr. 2012, 82, 101–128. [Google Scholar] [CrossRef]
- Dore, S.; Montes-Helu, M.; Hart, S.C.; Hungate, B.A.; Koch, G.W.; Moon, J.B.; Finkral, A.J.; Kolb, T.E. Recovery of ponderosa pine ecosystem carbon and water fluxes from thinning and stand-replacing fire. Glob. Chang. Biol. 2012, 18, 3171–3185. [Google Scholar] [CrossRef]
- Froelich, N.; Croft, H.; Chen, J.M.; Gonsamo, A.; Staebler, R.M. Trends of carbon fluxes and climate over a mixed temperate-boreal transition forest in southern Ontario, Canada. Agric. For. Meteorol. 2015, 211, 72–84. [Google Scholar] [CrossRef]
- Granier, A.; Bréda, N.; Longdoz, B.; Gross, P.; Ngao, J. Ten years of fluxes and stand growth in a young beech forest at Hesse, North-eastern France. Ann. For. Sci. 2008, 65, 704. [Google Scholar] [CrossRef] [Green Version]
- Herbst, M.; Mund, M.; Tamrakar, R.; Knohl, A. Differences in carbon uptake and water use between a managed and an unmanaged beech forest in central Germany. For. Ecol. Manag. 2015, 355, 101–108. [Google Scholar] [CrossRef]
- Novick, K.; Oishi, C.; Ward, E.; Siqueira, M.; Juang, J.-Y.; Stoy, P. On the difference in the net ecosystem exchange of CO2 between deciduous and evergreen forests in the southeastern United States. Glob. Chang. Biol. 2015, 21, 827–842. [Google Scholar] [CrossRef] [Green Version]
- Pilegaard, K.; Ibrom, A.; Courtney, M.S.; Hummelshøj, P.; Jensen, N.O. Increasing net CO2 uptake by a Danish beech forest during the period from 1996 to 2009. Agric. For. Meteorol. 2011, 151, 934–946. [Google Scholar] [CrossRef]
- Baumgartner, A. Meteorological approach to the exchange of CO2 between the atmosphere and vegetation, particularly foreststands. Photosynthetica 1969, 3, 127–149. [Google Scholar]
- Denmead, O.T. Comparative micrometeorology of a wheat field and a forest of Pinus radiata. Agric. Meteorol. 1969, 6, 357–371. [Google Scholar] [CrossRef]
- Speckman, H.; Frank, J.; Bradford, J.; Miles, B.; Massman, W.; Parton, W.; Ryan, M. Forest ecosystem respiration estimated from eddy covariance and chambermeasurements under high turbulence and substantial tree mortality from bark beetles. Glob. Chang. Biol. 2015, 21, 708–721. [Google Scholar] [CrossRef]
- Shan, J. Study on the spermatophyte flora of the coastal sandy region of hainan island. Guangdong For. Ence Technol. 2008, 24, 37–40. [Google Scholar]
- Herrmann, M.; Najjar, R.G.; Kemp, W.M.; Alexander, R.B.; Boyer, E.W.; Cai, W.-J.; Griffith, P.C.; Kroeger, K.D.; McCallister, S.L.; Smith, R.A. Net ecosystem production and organic carbon balance of US East Coast estu-aries: A synthesis approach. Glob. Biogeochem. Cycles 2015, 29, 96–111. [Google Scholar] [CrossRef]
- Xue, Y.; Yang, Z.Y.; Su, S.F.; Wang, X.Y.; Lin, Z.P.; Zhao, J.F.; Tan, Z.H. A preliminary report on mass and energy flux over a coastal vegetation in Hainan Island. Trop. For. 2016, 44, 48–52, (In Chinese with English abstract). [Google Scholar]
- Xue, Y.; Yang, Z.Y.; Chen, Y.Q.; Wang, X.Y.; Su, S.F.; Lin, Z.P.; Lin, R.W.; Xue, Y.W. Characteristics of soil organic carbon and litter of typical forest types in tropical coastal zone. Chin. J. Trop. Crops 2016, 37, 2083–2088, (In Chinese with English abstract). [Google Scholar]
- Verma, S.B. Micrometeorological methods for measuring surface fluxes of mass and energy. Remote Sens. Rev. 1990, 5, 99–115. [Google Scholar] [CrossRef]
- Wilczak, J.M.; Oncley, S.P.; Stage, S.A. Sonic anemometer tilt correction algorithms. Bound. Layer Meteorol. 2001, 99, 127–150. [Google Scholar] [CrossRef]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurements for density effect due to heat and water vapor transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Kljun, N.; Calanca, P.; Rotach, M.W.; Schmid, H.P. A simple parameterization for flux footprint predictions. Bound. Layer Meteorol. 2004, 112, 503–523. [Google Scholar] [CrossRef]
- Moncrieff, J.B.; Massheder, J.M.; de Bruin, H.; Ebers, J.; Friborg, T.; Heusinkveld, B.; Kabat, P.; Scott, S.; Soegaard, H.; Verhoef, A. A system to measure surface fluxes of momentum, sensible heat, water vapor and carbon dioxide. J. Hydrol. 1997, 188–189, 589–611. [Google Scholar] [CrossRef]
- Vickers, D.; Mahrt, L. Quality control and flux sampling problems for tower and aircraft data. J. Atmos. Ocean. Technol. 1997, 14, 512–526. [Google Scholar] [CrossRef]
- Saleska, S.R.; Miller, S.D.; Matross, D.M.; Goulden, M.L.; Wofsy, S.C.; da Rocha, H.; de Camargo, P.B.; Crill, P.M.; Daube, B.C.; Freitas, C.L.; et al. Carbon in Amazon forests: Unexpected seasonal fluxes and disturbance-induced losses. Science 2003, 302, 1554–1557. [Google Scholar] [CrossRef] [Green Version]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Aubinet, M.; Bernhofer, C.; Burba, G.; Ceulemans, R.; Clement, R.; Dolman, H.; et al. Gap filling strategies for long term energy flux data sets. Agric. For. Meteorol. 2001, 107, 71–77. [Google Scholar] [CrossRef] [Green Version]
- Penman, H.L. Natural evaporation from open water, bare soil and grass. Proc. R. Soc. A Math. Phys. Eng. Sci. 1948, 193, 120–145. [Google Scholar]
- Verma, S.B. Aerodynamic resistances to transfers of heat, mass and momentum. In Estimation of Areal Evapotranspiration; Black, T.A., Spittlehouse, D.L., Novak, M.D., Price, D.T., Eds.; IAHS Publication: Vancouver, BC, Canada, 1989; pp. 13–20. [Google Scholar]
- Paulson, C.A. The mathematical representation of wind speed and temperature profiles in the unstable atmospheric surface layer. J. Appl. Meteorol. 1970, 9, 857–861. [Google Scholar] [CrossRef]
- Falge, E.; Baldocchi, D.; Olson, R.; Anthoni, P.; Tu, K. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric. For. Meteorol. 2001, 107, 43–69. [Google Scholar] [CrossRef] [Green Version]
- Hoff, J.H.; Lehfeldt, R.A. Lectures on Theoretical and Physical Chemistry. Nature 1900, 62, 245. [Google Scholar]
- Jarvis, P.G.; McNaughton, K.G. Stomatal control of transpiration: Scaling up from leaf to region. Adv. Ecol. Res. 1986, 15, 1–49. [Google Scholar]
- Priestley, C.B.H.; Taylor, R.J. On the assessment of surface heat flux and evaporation using large scale parameters. Mon. Weather. Rev. 1972, 100, 81–92. [Google Scholar] [CrossRef]
- McGloin, R.; Šigut, L.; Havránková, K.; Dušek, J.; Pavelka, M.; Sedlák, P. Energy balance closure at a variety of ecosystems in Central Europe with contrasting topographies. Agric. For. Meteorol. 2018, 248, 418–431. [Google Scholar] [CrossRef]
- Foken, T. The energy balance closure problem: An overview. Ecol. Appl. 2008, 18, 1351–1367. [Google Scholar] [CrossRef]
- Oncley, S.P.; Foken, T.; Vogt, R.; Kohsiek, W.; Debruin, H.A.R.; Bernhofer, C.; Christen, A.; van Gorsel, E.; Grantz, D.; Feigenwinter, C.; et al. The Energy Balance Experiment EBEX-2000. Part I: Overview and energy balance. Bound. Layer Meteorol. 2007, 123, 1–28. [Google Scholar] [CrossRef]
- Leuning, R.; van Gorsel, E.; Massman, W.J.; Isaac, P.R. Reflections on the surface energy imbalance problem. Agric. For. Meteorol. 2012, 156, 65–74. [Google Scholar] [CrossRef]
- Wilson, K.; Goldstein, A.; Falge, E.; Aubinet, M.; Baldocchi, D.; Berbigier, P.; Bernhofer, C.; Ceulemans, R.; Dolman, H.; Field, C.; et al. Energy balance closure at FLUXNET sites. Agric. For. Meteorol. 2002, 113, 223–243. [Google Scholar] [CrossRef] [Green Version]
- Franssen, H.J.H.; Stockli, R.; Lehner, I.; Rotenberg, E.; Seneviratne, S.I. Energy balance closure of eddy-covariance data: A multisite analysis for European FLUXNET stations. Agric. For. Meteorol. 2010, 150, 1553–1567. [Google Scholar] [CrossRef]
- Zhang, L.-M.; Yu, G.-R.; Sun, X.-M.; Wen, X.-F.; Ren, C.-Y.; Fu, Y.-L.; Li, Q.-K.; Li, Z.-Q.; Liu, Y.-F.; Guan, D.-X.; et al. Seasonal variations of ecosystem apparent quantum yield (α) and maximum photosynthesis rate (Pmax) of different forest ecosystems in China. Agric. For. Meteorol. 2006, 137, 176–187. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Thomson, A. Temperature-associated increases in the global soil respiration record. Nature 2010, 464, 579–582. [Google Scholar] [CrossRef]
- Mahecha, M.; Reichstein, M.; Carvalhais, N.; Lasslop, G.; Lange, H.; Seneviratne, S.; Vargas, R.; Ammann, C.; Arain, A.; Cescatti, A.; et al. Global convergence in the temperature sensitivity of respiration at ecosystem level. Science 2010, 329, 838–840. [Google Scholar] [CrossRef] [Green Version]
- Kelliher, F.M.; Leuning, R.; Raupach, M.R.; Schulze, E.D. Maximum conductances for evaporation from global vegetation types. Agric. For. Meteorol. 1995, 73, 1–16. [Google Scholar] [CrossRef]
- Grace, J.; Lloyd, J.; McIntyre, J.; Miranda, A.; Meir, P.; Miranda, H.; Moncrieff, J.; Massheder, J.; Wright, I.; Gash, J. Fluxes of carbon dioxide and water over an undisturbed tropical forest in south-west Amazonia. Glob. Chang. Biol. 1995, 1, 1–12. [Google Scholar] [CrossRef]
- Kumagai, T.; Saitoh, T.M.; Sato, Y.; Morooka, T.; Manfroi, O.J.; Kuraji, K.; Suzuki, M. Transpiration, canopy conductance and the decoupling coefficient of a lowland mixed dipterocarp forest in Sarawak, Borneo: Dry spell effect. J. Hydrol. 2004, 287, 237–251. [Google Scholar] [CrossRef]
- Pereira, A.R. The priestley–taylor parameter and the decoupling factor for estimating reference evapotranspiration. Agric. For. Meteorol. 2004, 125, 305–313. [Google Scholar] [CrossRef]
- Blanken, P.D.; Black, T.A.; Yang, P.C.; Neumann, H.H.; Nesic, Z.; Staebler, R.; den Hartog, G.; Novak, M.D.; Lee, X. Energy balance and canopy conductance of a boreal aspen forest: Partitioning overstory and understory components. J. Geophys. Res. Atmos. 1997, 102, 28915–28927. [Google Scholar] [CrossRef] [Green Version]
- Monteith, J.L. Accommodation between transpiring vegetation and the convective boundary layer. J. Hydrol. 1995, 116, 251–263. [Google Scholar] [CrossRef]
- Yang, H.; Li, Y.-D.; Ren, H.; Luo, T.-S.; Chen, R.-L.; Liu, W.-J.; Chen, D.-X.; Xu, H.; Zhou, Z.; Lin, M.-X.; et al. Soil organic carbon density and influencing factors in tropical virgin forests of Hainan Island, China. Chin. J. Plant Ecol. 2016, 40, 292–303. [Google Scholar]
- Takada, M.; Yamada, T.; Kadir, W.R.; Okuda, T. Spatial and temporal variation in soil respiration in a Southeast Asian tropical rainforest. Agric. For. Meteorol. 2007, 147, 35–47. [Google Scholar]
- Xue, Y.; Yang, Z.; Wang, X.; Lin, Z.; Li, D.; Su, S. Tree biomass allocation and its model Additivity for Casuarina equisetifolia in a tropical forest of Hainan Island, China. PLoS ONE 2016, 11, e0151858. [Google Scholar] [CrossRef] [Green Version]
- Malhi, Y.; Aragao, L.E.O.C.; Metcalfe, D.B.; Paiva, R.; Quesada, C.A.; Almeida, S.; Anderson, L.; Brando, P.; Chambers, J.Q.; da Costa, A.C.L.; et al. Comprehensive assessment of carbon productivity, allocation and storage in three Amazonian forests. Glob. Chang. Biol. 2009, 15, 1255–1274. [Google Scholar] [CrossRef]
- da Rocha, H.R.; Manzi, A.O.; Cabral, O.M.; Miller, S.D.; Goulden, M.L.; Saleska, S.R.; Coupe, N.R.; Wofsy, S.C.; Borma, L.S.; Artaxo, P.; et al. Patterns of water and heat flux across a biome gradient from tropical forest to savanna in Brazil. J. Geophys. Res. Biogeosci. 2009, 114. [Google Scholar] [CrossRef]
- Wu, J.; Albert, L.P.; Lopes, A.P.; Restrepo-Coupe, N.; Hayek, M.; Wiedemann, K.T.; Guan, K.; Stark, S.C.; Christoffersen, B.; Prohaska, N.; et al. Leaf development and demography explain photosynthesis seasonality in Amazon evergreen forests. Science 2016, 351, 972–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boggs, J.L.; Sun, G.; Domec, J.-C.; McNulty, S.G. Variability of tree transpiration across three zones in a southeastern US Piedmont watershed. Hydrol. Process. 2021, 35, e14389. [Google Scholar] [CrossRef]











| Instruments | Model | Manufacturer | Height or Depth |
|---|---|---|---|
| Sonic anemometer | WindMaster | Gill Instruments, Lymington, UK | 25 m |
| Infrared gas analyzer | Li-7500A | Li-Cor Inc., Lincoln, USA | 25 m |
| Net radiometer | NR01 | Hukseflux, Delft, Netherlands | 12 m |
| Photosynthetically active radiation | Li-190SL | Li-Cor Inc., Lincoln, USA | 24, 18, 13, 3 m |
| Wind cup | 03001 | RM Young, Traverse City, USA | 24, 18, 13, 3 m |
| Humidity and temperature probe | HMP-60 | Vaisala, Vantaa, Finland | 24, 18, 13, 3 m |
| Rain gauge | TE-525 | Texas Electronics, Dallas, USA | 24, 2 m |
| Soil heat flux | HFP01 | Hukseflux, Delft, Netherlands | −10 cm |
| Soil temperature | TM-L10 | Dynamax Inc., Houston, USA | −10, −20, −40, −80 cm |
| Soil water content | EC-5 | Decagon, Pullman, USA | −10, −20, −40, −80 cm |
| Flux-calculating module | SmartFlux | Li-Cor Inc., Lincoln, Nebraska USA | 24 m |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, J.-T.; Xue, Y.; Zhao, J.-F.; Yang, Z.-Y.; Su, S.-F.; Wang, X.-Y.; Lin, Z.-P.; Wang, G.-Z.; Yang, L.-Y.; Zhang, X. Atmospheric Exchange of Carbon Dioxide and Water Vapor above a Tropical Sandy Coastal Plain. Water 2023, 15, 877. https://doi.org/10.3390/w15050877
Jia J-T, Xue Y, Zhao J-F, Yang Z-Y, Su S-F, Wang X-Y, Lin Z-P, Wang G-Z, Yang L-Y, Zhang X. Atmospheric Exchange of Carbon Dioxide and Water Vapor above a Tropical Sandy Coastal Plain. Water. 2023; 15(5):877. https://doi.org/10.3390/w15050877
Chicago/Turabian StyleJia, Jun-Ting, Yang Xue, Jun-Fu Zhao, Zhong-Yang Yang, Shao-Feng Su, Xiao-Yan Wang, Zhi-Pan Lin, Guan-Ze Wang, Lian-Yan Yang, and Xiang Zhang. 2023. "Atmospheric Exchange of Carbon Dioxide and Water Vapor above a Tropical Sandy Coastal Plain" Water 15, no. 5: 877. https://doi.org/10.3390/w15050877