
Source Water Apportionment Using Stable Isotopes for Typical Riparian Plants along the Manas River in Xinjiang, Northwest China
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
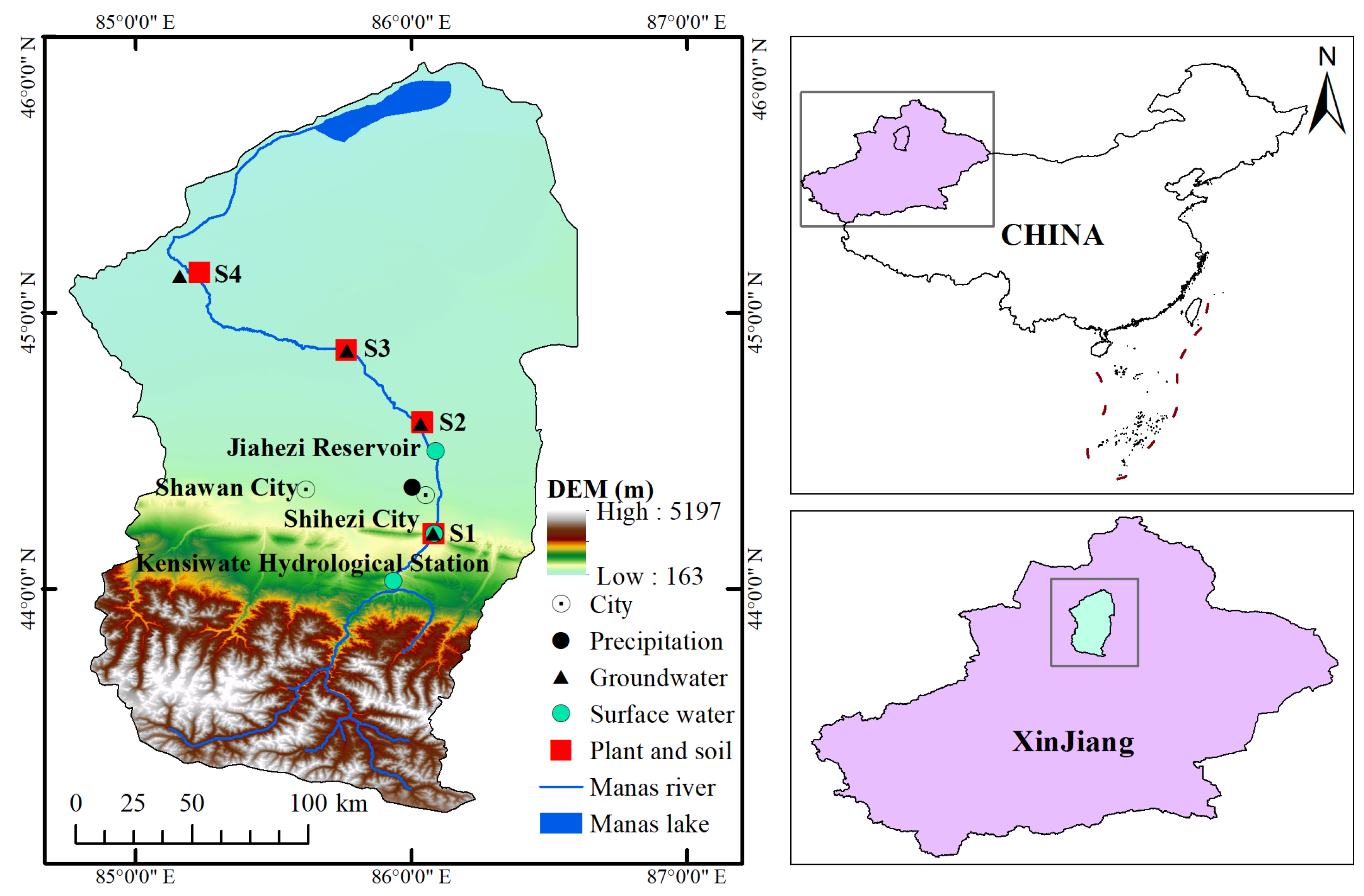
2.1. Study Area
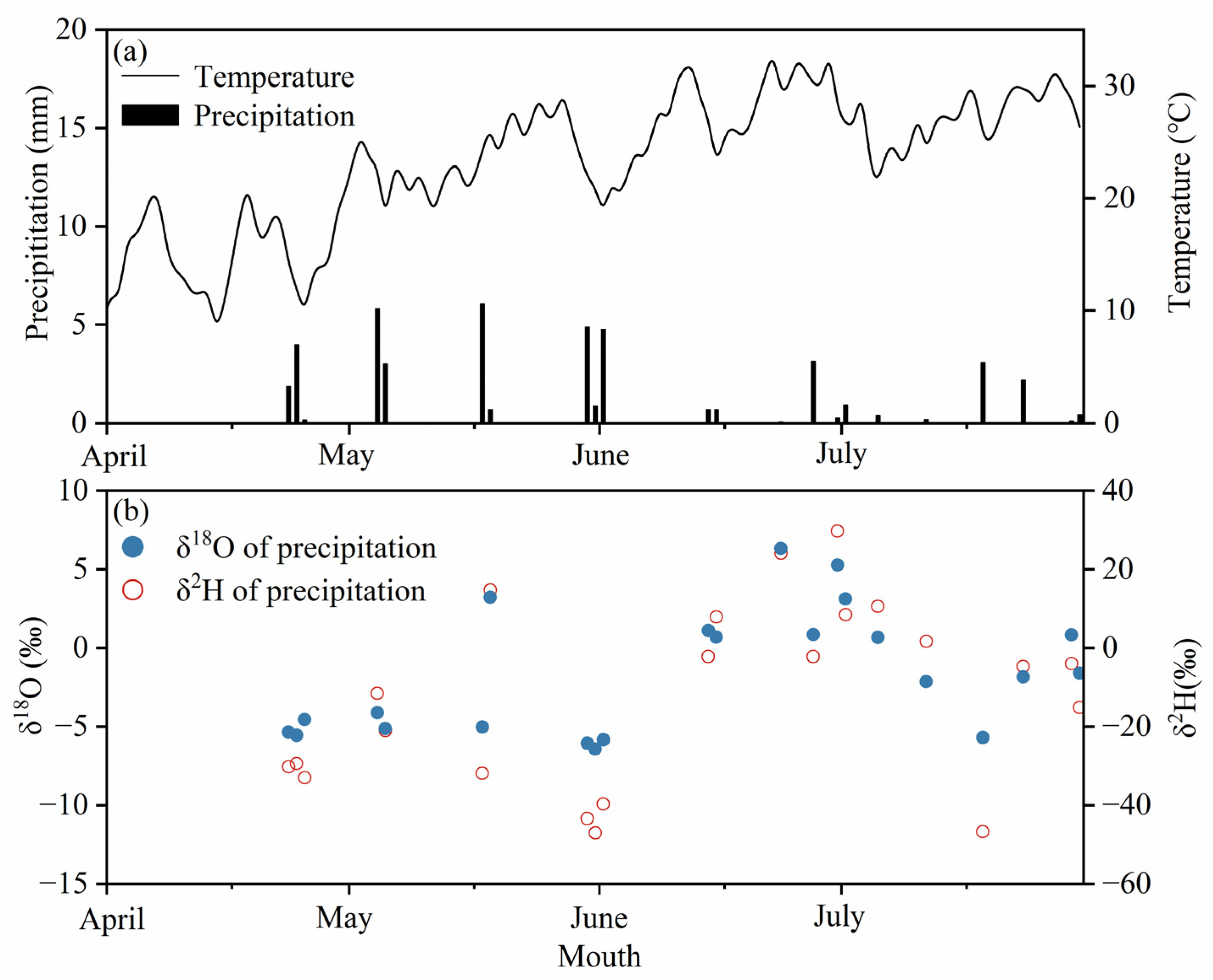
2.2. Sample Collection and Analysis
2.2.1. Sample Collection
2.2.2. Isotopic Analysis
2.2.3. MixSIAR Model
2.2.4. Proportional Similarity Index
3. Results and Discussion
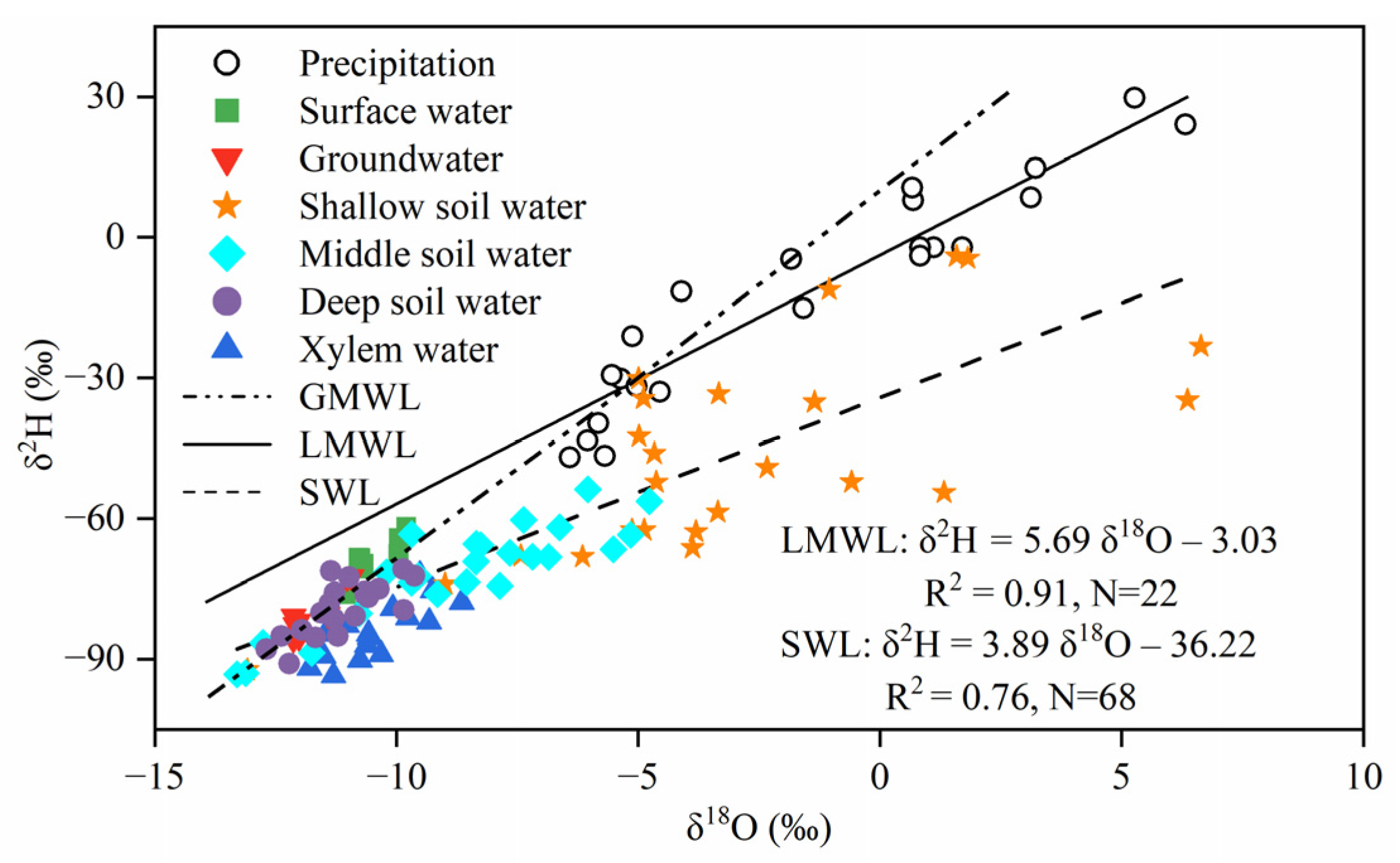
3.1. Stable Isotope Compositions of Different Water Bodies
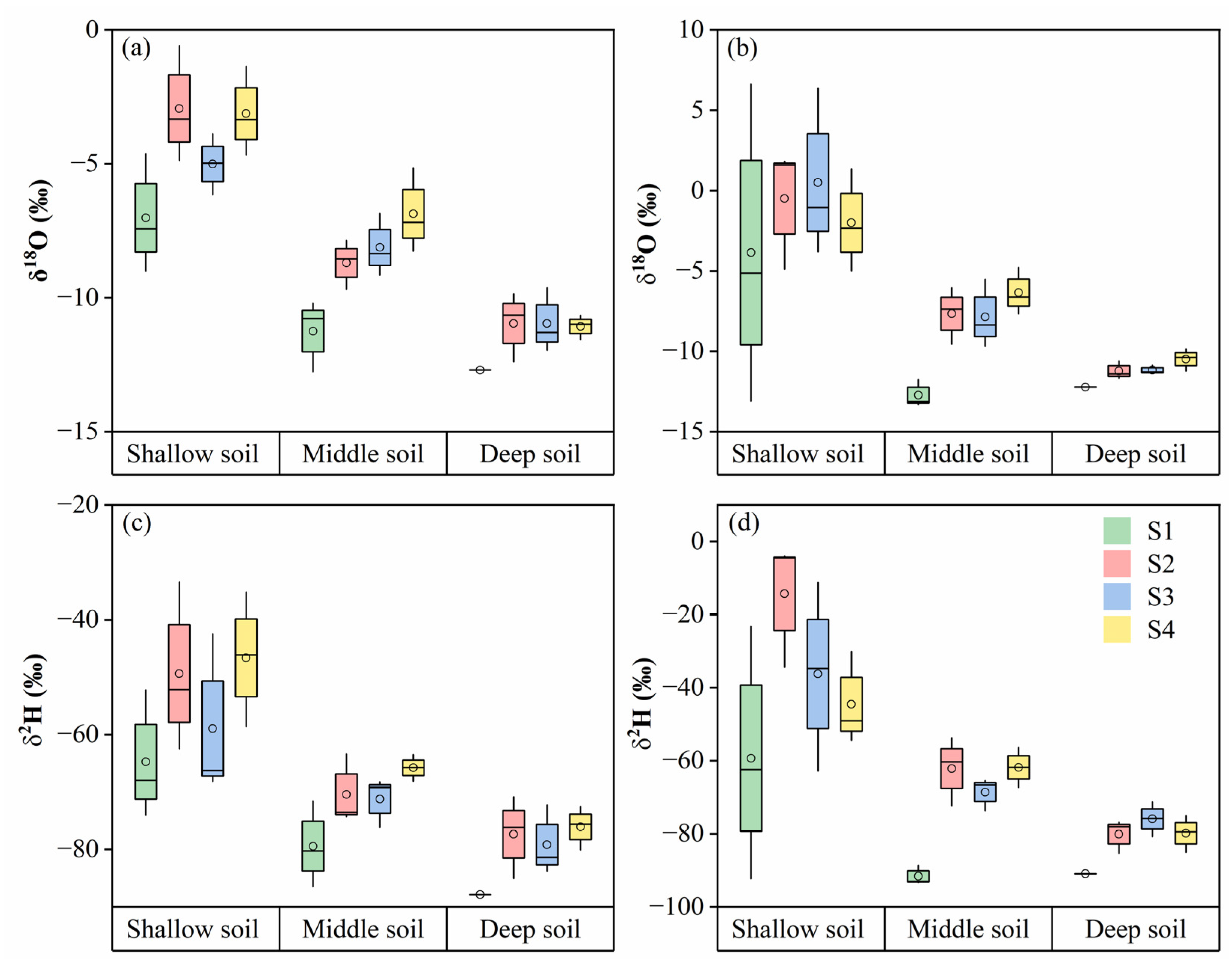
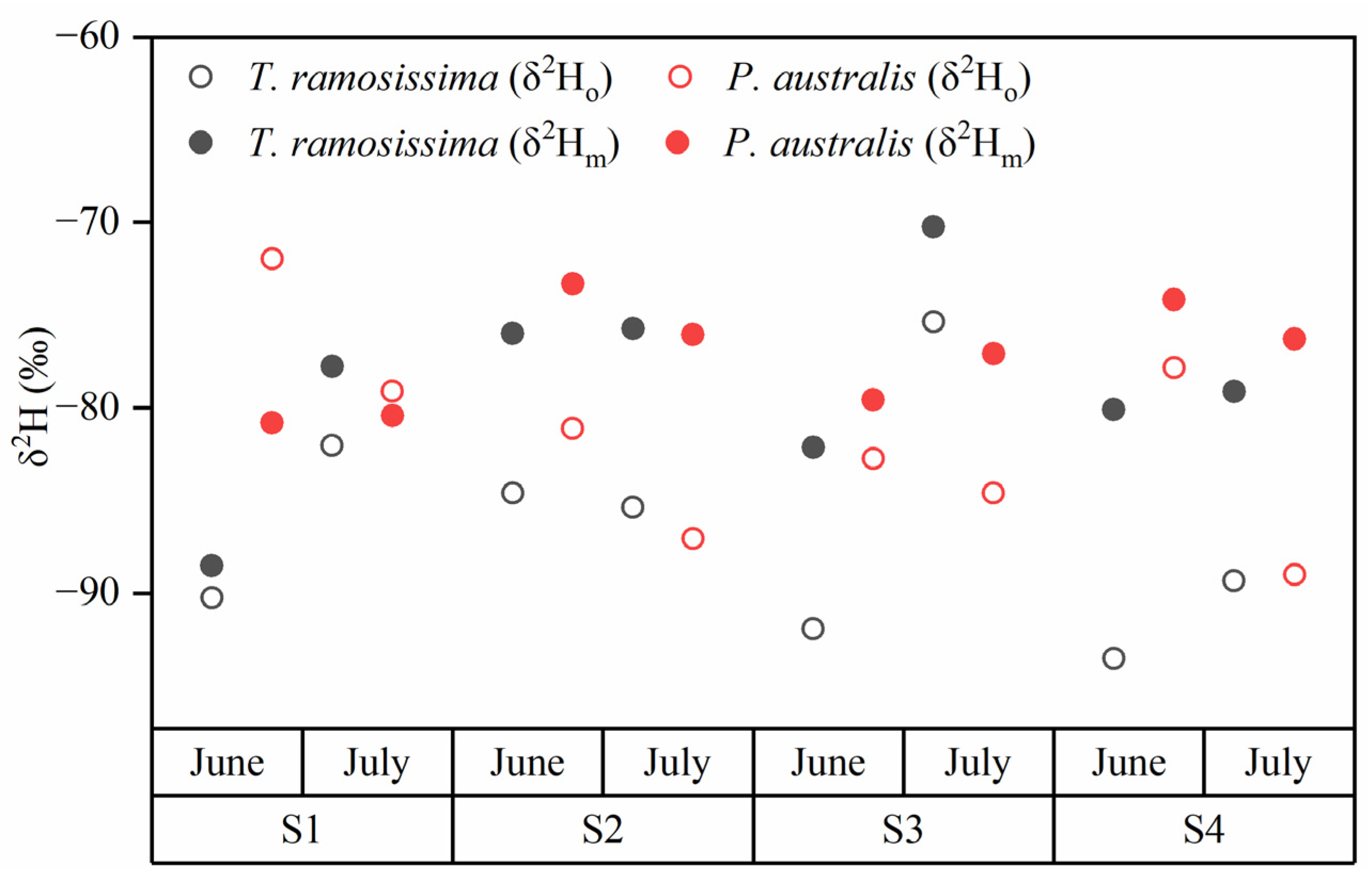
3.2. Stable Isotope Compositions of Plant Water
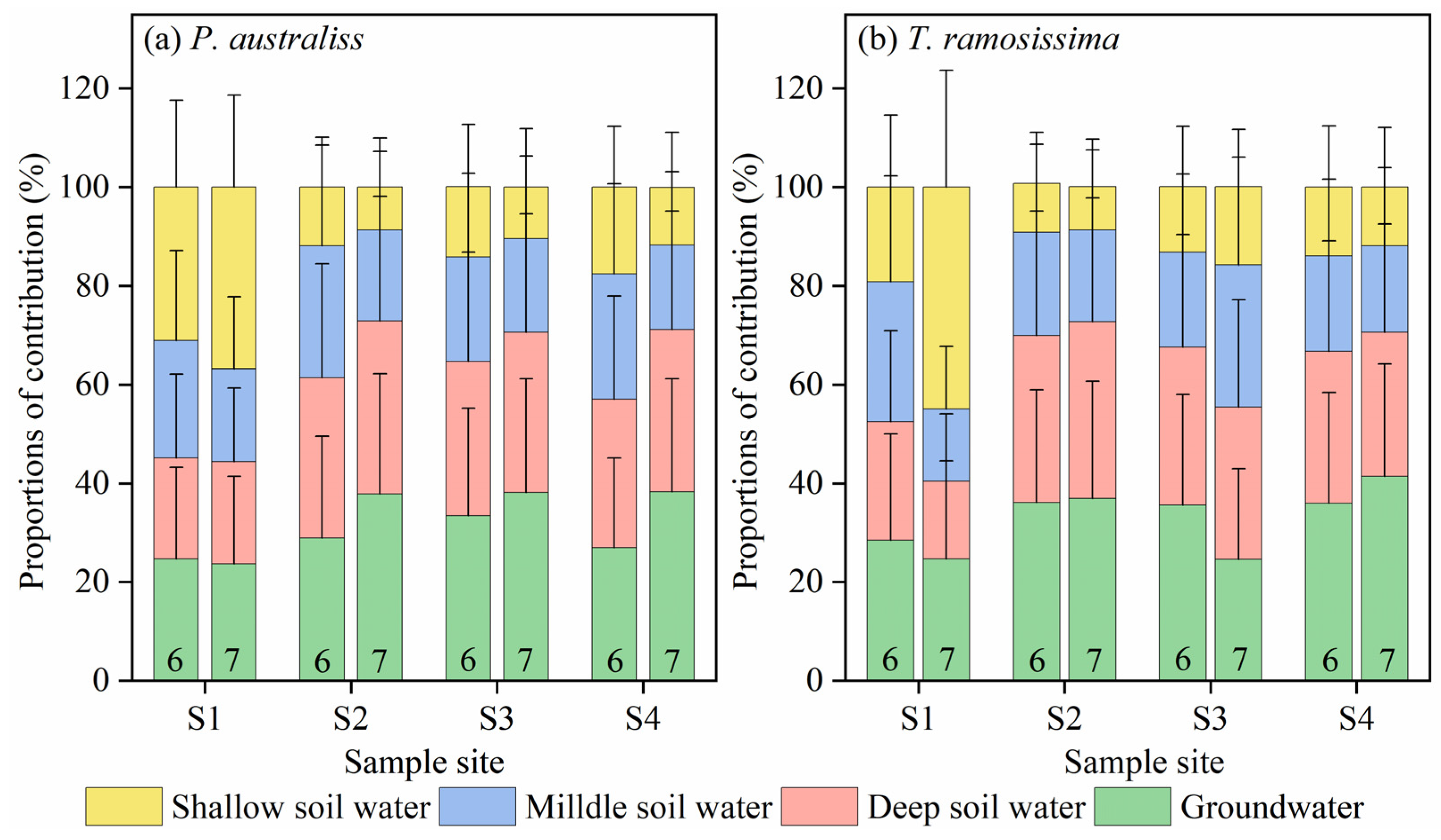
3.3. Contribution Percentages of Potential Water Sources to Plant Water
3.4. Competition for Water between T. ramosissima and P. australis
4. Conclusions
- (1)
- The slope and intercept of the LMWL were smaller compared with the GMWL, indicating that the climate in the study area is dry and subject to strong evaporation. The interconversion of surface water and groundwater occurred mainly in the upper reaches of rivers. Furthermore, the PW-excess values for both T. ramosissima and P. australis are not equal to 0, indicating that hydrogen isotope fractionation occurs during plant water uptake.
- (2)
- The water uptake patterns of T. ramosissima and P. australis in long-term ecological water conveyance samples are to evenly utilize or mainly utilize shallow soil water, and the utilization rate of shallow soil water reaches 30.7 ± 12.6%. In contrast, T. ramosissima and P. australis in intermittent ecological water conveyance samples and non-ecological water conveyance samples mainly absorb and utilize groundwater and deep soil water, with mean values of 34.5 ± 5.1% and 32.2 ± 1.9%, respectively. Therefore, plants can adjust their water uptake patterns to different habitats.
- (3)
- The PSI under all three ecological water conveyance conditions is more than 0.7. The water competition among plants was more intense at sample sites S2-S4, and the long-term ecological water conveyance effectively reduced the water competition among plants.
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bothwell, H.M.; Cushman, S.A.; Woolbright, S.A.; Hersch-Green, E.I.; Evans, L.M.; Whitham, T.G.; Allan, G.J. Conserving threatened riparian ecosystems in the American West: Precipitation gradients and river networks drive genetic connectivity and diversity in a foundation riparian tree (Populus angustifolia). Mol. Ecol. 2017, 26, 5114–5132. [Google Scholar] [CrossRef] [PubMed]
- Stromberg, J.C.; Beauchamp, V.B.; Dixon, M.D.; Lite, S.J.; Paradzick, C. Importance of low-flow and high-flow characteristics to restoration of riparian vegetation along rivers in arid south-western United States. Freshw. Biol. 2007, 52, 651–679. [Google Scholar] [CrossRef]
- Garcia, C.; Amengual, A.; Homar, V.; Zamora, A. Losing water in temporary streams on a Mediterranean island: Effects of climate and land-cover changes. Glob. Planet. Change 2017, 148, 139–152. [Google Scholar] [CrossRef]
- Stromberg, S.C.; Merritt, D.M. Riparian plant guilds of ephemeral, intermittent and perennial rivers. Freshw. Biol. 2016, 61, 1259–1275. [Google Scholar] [CrossRef]
- Pang, Z.; Huang, T.; Chen, Y. Diminished groundwater recharge and circulation relative to degrading riparian vegetation in the middle Tarim River, Xinjiang Uygur, Western China. Hydrol. Process. 2010, 24, 147–159. [Google Scholar] [CrossRef]
- Sun, W.; Song, X.; Mu, X.; Gao, P.; Wang, F.; Zhao, G. Spatiotemporal vegetation cover variations associated with climate change and ecological restoration in the Loess Plateau. Agric. For. Meteorol. 2015, 209, 87–99. [Google Scholar] [CrossRef]
- Huan, L.; Shaorong, Y.; Xiaoming, W. Research Progress on Ecological Restoration Technology Based on River Ecosystem Health. J. Hydro. 2019, 40, 1–6. [Google Scholar]
- Hao, X.; Li, W. Impacts of ecological water conveyance on groundwater dynamics and vegetation recovery in the lower reaches of the Tarim River in northwest China. Environ. Monit. Assess 2014, 186, 7605–7616. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, H. Study on vegetation change of Taitemar Lake during ecological water transfer. Environ. Monit. Assess 2019, 191, 613. [Google Scholar] [CrossRef]
- Costelloe, J.F.; Payne, E.; Woodrow, I.E.; Irvine, E.C.; Western, A.W.; Leaney, F.W. Water sources accessed by arid zone riparian trees in highly saline environments, Australia. Oecologia 2008, 156, 43–52. [Google Scholar] [CrossRef]
- Chen, Y.N.; Li, W.H.; Zhou, H.H.; Chen, Y.P.; Hao, X.M.; Fu, A.H.; Ma, J.X. Analysis of water use strategies of the desert riparian forest plant community in inland rivers of two arid regions in northwestern China. Biogeosci. Discuss. 2014, 11, 14819–14856. [Google Scholar]
- Li, Y.; Ma, Y.; Song, X.; Wang, L.; Yang, L.; Li, X.; Li, B. Contrasting water use characteristics of riparian trees under different water tables along a losing river. J. Hydrol. 2022, 611, 128017. [Google Scholar] [CrossRef]
- Wang, P.; Liu, W.; Zhang, J.; Yang, B.; Singh, A.K.; Wu, J.; Jiang, X.J.E. Seasonal and spatial variations of water use among riparian vegetation in tropical monsoon region of SW China. Ecohydrology 2019, 12, e2085. [Google Scholar] [CrossRef]
- Ellsworth, P.Z.; Williams, D.G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant Soil 2007, 291, 93–107. [Google Scholar] [CrossRef]
- Zimmermann, U.; Münnich, K.; Roether, W.; Kreutz, W.; Schubach, K.; Siegel, O. Tracers determine movement of soil moisture and evapotranspiration. Science 1966, 152, 346–347. [Google Scholar] [CrossRef]
- Rothfuss, Y.; Javaux, M. Reviews and syntheses: Isotopic approaches to quantify root water uptake: A review and comparison of methods. Biogeosciences 2017, 14, 2199–2224. [Google Scholar] [CrossRef] [Green Version]
- Hao, S.; Li, F. Water sources of the typical desert vegetation in Ebinur Lake basin. Acta Geogr. Sin. 2021, 76, 1649–1661. [Google Scholar] [CrossRef]
- Miguez-Macho, G.; Fan, Y. Spatiotemporal origin of soil water taken up by vegetation. Nature 2021, 598, 624–628. [Google Scholar] [CrossRef]
- Dawson, T.E.; Ehleringer, J.R. Streamside trees that do not use stream water. Nature 1991, 350, 335–337. [Google Scholar] [CrossRef]
- Holland, K.L.; Charles, A.H.; Jolly, I.D.; Overton, I.C.; Gehrig, S.; Simmons, C.T. Effectiveness of artificial watering of a semi-arid saline wetland for managing riparian vegetation health. Hydrol. Process. 2009, 23, 3474–3484. [Google Scholar] [CrossRef]
- Si, J.; Feng, Q.; Cao, S.; Yu, T.; Zhao, C. Water use sources of desert riparian Populus euphratica forests. Environ. Monit. Assess. 2014, 186, 5469–5477. [Google Scholar] [CrossRef] [PubMed]
- Rao, W.; Chen, X.; Meredith, K.T.; Tan, H.; Gao, M.; Liu, J. Water uptake of riparian plants in the lower Lhasa River Basin, South Tibetan Plateau using stable water isotopes. Hydrol. Process. 2020, 34, 3492–3505. [Google Scholar] [CrossRef]
- Wu, H.; Zhao, G.; Li, X.-Y.; Wang, Y.; He, B.; Jiang, Z.; Zhang, S.; Sun, W. Identifying water sources used by alpine riparian plants in a restoration zone on the Qinghai-Tibet Plateau: Evidence from stable isotopes. Sci. Total Environ. 2019, 697, 134092. [Google Scholar] [CrossRef] [PubMed]
- O’Grady, A.; Cook, P.; Eamus, D.; Duguid, A.; Wischusen, J.; Fass, T.; Worldege, D. Convergence of tree water use within an arid-zone woodland. Oecologia 2009, 160, 643–655. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Niu, Y.B.; Xun, M.Y.; Jin, J.Y. Responses of water source to seasonal drought of dominant afforestation tree species in the loess hilly region of China. Acta Ecol. Sin. 2022, 42, 4101–4112. [Google Scholar]
- Smith, S.D.; Wellington, A.B.; Nachlinger, J.L.; Fox, C.A. Functional responses of riparian vegetation to streamflow diversion in the eastern Sierra Nevada. Ecol. Appl. 1991, 1, 89–97. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Lu, N.; Zhang, L. Seasonal variation in water uptake patterns of three plant species based on stable isotopes in the semi-arid Loess Plateau. Sci. Total Environ. 2017, 609, 27–37. [Google Scholar] [CrossRef]
- Stromberg, J.C.; Bagstad, K.J.; Leenhouts, J.M.; Lite, S.J.; Makings, E. Effects of stream flow intermittency on riparian vegetation of a semiarid region river (San Pedro River, Arizona). River Res. Appl. 2005, 21, 925–938. [Google Scholar] [CrossRef]
- Wuchao, Y.; Fengmei, J.; Yajun, S.; Zhi, L. Runoff change controlled by combined effects of multiple environmental factors in a headwater catchment with cold and arid climate in northwest China. Sci. Total Environ. 2021, 756, 143995. [Google Scholar]
- Yang, G.; Xue, L.; He, X.; Wang, C.; Long, A.H. Change in land use and evapotranspiration in the Manas River Basin, China with long-term water-saving measures. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Chen, D.; He, X.; Long, A.; Yang, M.; Li, X. Land use change characteristics affected by water saving practices in Manas River Basin, China using Landsat satellite images. Int. J. Agric. Biol. Eng. 2017, 10, 123–133. [Google Scholar]
- Wang, Z.; Wang, W.; Zhang, Z.; Hou, X.; Ma, Z.; Chen, B.J.G.E. River-groundwater interaction affected species composition and diversity perpendicular to a regulated river in an arid riparian zone. Glob. Ecol. Conserv. 2021, 27, e01595. [Google Scholar] [CrossRef]
- Liu, J.; Yan, P.; Zhai, W.; Jiang, J. Floristic characteristics of seed plants in low mountain deserts in the upper and middle reaches of the Manas River in Xinjiang. Pratac. Sci. 2019, 36, 83–92. [Google Scholar]
- Wang, Z.; Wang, W.; Zhang, Z.; Hou, X.; Duan, L.; Yao, D. Assessment of the effect of water-table depth on riparian vegetation along the middle and lower reaches of the Manasi River, Northwest China. Hydrol. J. 2021, 29, 579–589. [Google Scholar] [CrossRef]
- Wang, J.; Liang, X.; Liu, Y.; Jin, M.; Knappett, P.S.; Liu, Y. Hydrogeochemical evolution along groundwater flow paths in the Manas River Basin, Northwest China. Groundwater 2019, 57, 575–589. [Google Scholar] [CrossRef]
- Wang, Z. The Effect of River-Groundwater Interaction on Riparianplants Ecology in Manasi River Baisn; Chang’an University: Xi’an, China, 2021. [Google Scholar]
- De la Casa, J.; Barbeta, A.; Rodríguez-Uña, A.; Wingate, L.; Ogée, J.; Gimeno, T. Isotopic offsets between bulk plant water and its sources are larger in cool and wet environments. Hydrol. Earth Syst. Sci. 2022, 26, 4125–4146. [Google Scholar] [CrossRef]
- Li, Y.; Ma, Y.; Song, X.; Wang, L.; Han, D. A δ2H offset correction method for quantifying root water uptake of riparian trees. J. Hydrol. 2021, 593, 125811. [Google Scholar] [CrossRef]
- Stock, B.C.; Jackson, A.L.; Ward, E.J.; Parnell, A.C.; Phillips, D.L.; Semmens, B.X. Analyzing mixing systems using a new generation of Bayesian tracer mixing models. PeerJ 2018, 6, e5096. [Google Scholar] [CrossRef]
- Colwell, R.K.; Futuyma, D.J. On the measurement of niche breadth and overlap. Ecol. Lett. 1971, 52, 567–576. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, M.; Hughes, C.E.; Zhu, X.; Dong, L.; Ren, Z.; Chen, F. Factors controlling stable isotope composition of precipitation in arid conditions: An observation network in the Tianshan Mountains, central Asia. Tellus B Chem. Phys. Meteorol. 2016, 68, 26206. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Rao, Z.; Zhang, X.; Huang, W.; Chen, J.; Chen, F. Variations in the oxygen isotopic composition of precipitation in the Tianshan Mountains region and their significance for the Westerly circulation. J. Geogr. Sci. 2015, 25, 801–816. [Google Scholar] [CrossRef]
- Li, X.F.; Zhang, M.J.; Li, Y.J.; Wang, S.J.; Huang, X.Y.; Ma, Q.; Ma, X.N. Characteristics of δ18O in Precipitation and Moisture Transports over the Arid Region in Northwest China. Environ. Sci. 2012, 33, 711–719. [Google Scholar]
- Chen, X.; Wang, G.; Wang, F. Classification of Stable Isotopes and Identification of Water Replenishment in the Naqu River Basin, Qinghai-Tibet Plateau. Water 2019, 11, 46. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.D.; Zhang, F.; Zhang, Z.U.; Kung, H.T.; Yushanjiang, A. Hydrogen and Oxygen Isotope Composition and Water Quality Evaluation for Different Water Bodies in the Ebinur Lake Watershed, Northwestern China. Water 2019, 11, 2067. [Google Scholar] [CrossRef] [Green Version]
- Bailing, F.; Dong, Z.; Zhenghua, T.; Zhiqi, Z. Compositions of hydrogen and oxygen isotope values of Yellow River water and the response to climate change. Chin. Environ. Sci. 2017, 37, 9. [Google Scholar]
- Freitas, J.G.; Furquim, S.A.C.; Aravena, R.; Cardoso, E.L. Interaction between lakes’ surface water and groundwater in the Pantanal wetland, Brazil. Environ. Earth Sci. 2019, 78, 139. [Google Scholar] [CrossRef]
- Darling, W.G.; Bath, A.H.; Talbot, J.C. The O and H stable isotope composition of freshwaters in the British Isles. 2. Surface waters and groundwater. Hydrol. Earth Syst. Sci. 2003, 7, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Ma, B.; Jin, M.; Liang, X.; Li, J. Groundwater mixing and mineralization processes in a mountain–oasis–desert basin, northwest China: Hydrogeochemistry and environmental tracer indicators. Hydrol. J. 2018, 26, 233–250. [Google Scholar] [CrossRef]
- Wang, J.; Liang, X.; Ma, B.; Liu, Y.; Jin, M.; Knappett, P.S.K.; Liu, Y. Using isotopes and hydrogeochemistry to characterize groundwater flow systems within intensively pumped aquifers in an arid inland basin, Northwest China. J. Hydrol. 2021, 595, 126048. [Google Scholar] [CrossRef]
- Canham, C.A.; Duvert, C.; Beesley, L.S.; Douglas, M.M.; Setterfield, S.A.; Freestone, F.L.; Clohessy, S.; Loomes, R.C. The use of regional and alluvial groundwater by riparian trees in the wet-dry tropics of northern Australia. Hydrol. Process. 2021, 35, e14180. [Google Scholar] [CrossRef]
- Qing, X.; Ting, W.; Deqiang, G. Application of Carbon, Hydrogen and Oxygen Stable Isotope on Hydrologic Cycle of Grassland Ecosystem: A Review. For. Res. 2019, 32, 130–136. [Google Scholar]
- Li, Y.F.; Yu, J.J.; Kai, L.; Wang, P. Water sources of Populus euphratica and Tamarix ramosissima in Ejina Delta, the lower reaches of the Heihe River, China. Chin. J. Plant Ecol. 2017, 41, 519–528. [Google Scholar]
- Gazis, C.; Feng, X.H. A stable isotope study of soil water: Evidence for mixing and preferential flow paths. Geoderma 2004, 119, 97–111. [Google Scholar] [CrossRef]
- Landgraf, J.; Tetzlaff, D.; Dubbert, M.; Dubbert, D.; Smith, A.; Soulsby, C. Xylem water in riparian willow trees (Salix alba) reveals shallow sources of root water uptake by in situ monitoring of stable water isotopes. Hydrol. Earth Syst. Sci. 2022, 26, 2073–2092. [Google Scholar] [CrossRef]
- Chen, Y.N.; Li, W.H.; Chen, Y.P.; Zhou, H.H.; Hao, X.M.; Zhu, C.G. Water Use Process of Constructive Plants in Desert Riparian Forest. Arid Zone Res. 2018, 35, 130–136. [Google Scholar]
- Wang, Y.; Chen, M.; Yan, L.; Yang, G.; Ma, J.; Deng, W. Quantifying Threshold Water Tables for Ecological Restoration in Arid Northwestern China. Groundwater 2020, 58, 132–142. [Google Scholar] [CrossRef]
- Yong, W.; Chengyi, Z.; Dandan, W.; Sihua, F. Water Use Strategies of Populus euphratica and Tamarix ramosissima at Different Ages in Tarim River Basin. J. Soiland Water Conserv. 2017, 31, 157–163. [Google Scholar]
- Song, T.H.; Ge, M.J.; Yang, J.M.; Zhang, D.; Li, Y.Z.; Liu, J.T.; Yu, J.B.; Guan, B. Stable isotope analysis of water use sources of Phrag·matis australis in heterogeneous water-salt habitats in the Yellow River Delta. Chin. J. Ecol. 2022, 41, 10. [Google Scholar]
- Qi, J.; Markewitz, D.; Radcliffe, D. Modelling the effect of changing precipitation inputs on deep soil water utilization. Hydrol. Process. 2018, 32, 672–686. [Google Scholar] [CrossRef]
- Jiang, L.X.; Li, Y. Comparison on Architecture Characteristics of Root Systems and Leaf Traits for Three Desert Shrubs Adapted to Arid Habitat. Acta Ecol. Sin. 2008, 28, 130–137. [Google Scholar]
- Xu, G.Q.; Yan, L. Roots distribution of three desert shrubs and their response to precipitation under co-occurri ng conditions. Acta Ecol. Sin. 2009, 29, 130–137. [Google Scholar]
- Sun, K.; Niu, J.; Wang, C.; Fu, Q.; Yang, G.; Liang, F.; Wang, Y. Effects of Different Irrigation Modes on the Growth, Physiology, Farmland Microclimate Characteristics, and Yield of Cotton in an Oasis. Water 2022, 14, 1579. [Google Scholar] [CrossRef]
- Liu, H.J.; Kang, Y.H. Effect of sprinkler irrigation on microclimate in the winter wheat field in the North China Plain. Agric. Water Manag. 2006, 84, 3–19. [Google Scholar] [CrossRef]
- Duan, D.Y.; Ouyang, H. Application of stable hydrogen and oxygen isotope in analyzing plant water use sources. Ecol. Environ. 2007, 16, 655–660. [Google Scholar]
- Su, P.; Zhang, M.; Qu, D.; Wang, J.; Zhang, Y.; Yao, X.; Xiao, H. Contrasting water use strategies of Tamarix ramosissima in different habitats in the Northwest of Loess Plateau, China. Water 2020, 12, 2791. [Google Scholar] [CrossRef]
- Wu, G.L.; Jiang, S.W.; Zhou, T.H.; Wang, D.D.; Li, J. Water Use Strategies of Seedlings of Populus euphratica and Tamarix ramosissima under Different Groundwater Depths. Arid Zone Res. 2016, 33, 8. [Google Scholar]
- Chen, Y.; Li, W.; Zhou, H.; Chen, Y.; Hao, X.; Fu, A.; Ma, J. Experimental study on water transport observations of desert riparian forests in the lower reaches of the Tarim River in China. Int. J. Biometeorol. 2017, 61, 1055–1062. [Google Scholar] [CrossRef]
- Zhang, Y.; Shen, Y.; Bai, S.; Wang, Z. Effects of the mixed on root growth and distribution of fraxinus mandshurica and larix gmelinii. Sci. Silv. Sin. 2001, 37, 16–23. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Site | Shallow Soil | Middle Soil | Deep Soil | |||
|---|---|---|---|---|---|---|
| June | July | June | July | June | July | |
| S1 | 6.50% | 10.94% | 7.84% | 19.56% | 7.91% | 21.58% |
| S2 | 5.65% | 3.94% | 3.85% | 5.07% | 12.99% | 7.14% |
| S3 | 6.25% | 4.29% | 7.91% | 6.03% | 11.95% | 7.70% |
| S4 | 4.26% | 2.30% | 5.16% | 2.55% | 6.77% | 2.90% |
| Month | Proportional Similarity Index | |||
|---|---|---|---|---|
| S1 | S2 | S3 | S4 | |
| June | 0.882 | 0.885 | 0.940 | 0.859 |
| July | 0.705 | 0.969 | 0.879 | 0.945 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ran, M.; Tian, H.; Yang, G.; Li, X.; Tian, L.; Kong, C.; Liu, B.; Li, P.; Xue, L.; Li, F.; et al. Source Water Apportionment Using Stable Isotopes for Typical Riparian Plants along the Manas River in Xinjiang, Northwest China. Water 2023, 15, 927. https://doi.org/10.3390/w15050927
Ran M, Tian H, Yang G, Li X, Tian L, Kong C, Liu B, Li P, Xue L, Li F, et al. Source Water Apportionment Using Stable Isotopes for Typical Riparian Plants along the Manas River in Xinjiang, Northwest China. Water. 2023; 15(5):927. https://doi.org/10.3390/w15050927
Chicago/Turabian StyleRan, Maolin, Hao Tian, Guang Yang, Xiaolong Li, Lijun Tian, Chunxian Kong, Bing Liu, Pengfei Li, Lianqing Xue, Fadong Li, and et al. 2023. "Source Water Apportionment Using Stable Isotopes for Typical Riparian Plants along the Manas River in Xinjiang, Northwest China" Water 15, no. 5: 927. https://doi.org/10.3390/w15050927