
Beneath the Aegean Sun: Investigating Dunaliella Strains’ Diversity from Greek Saltworks
 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
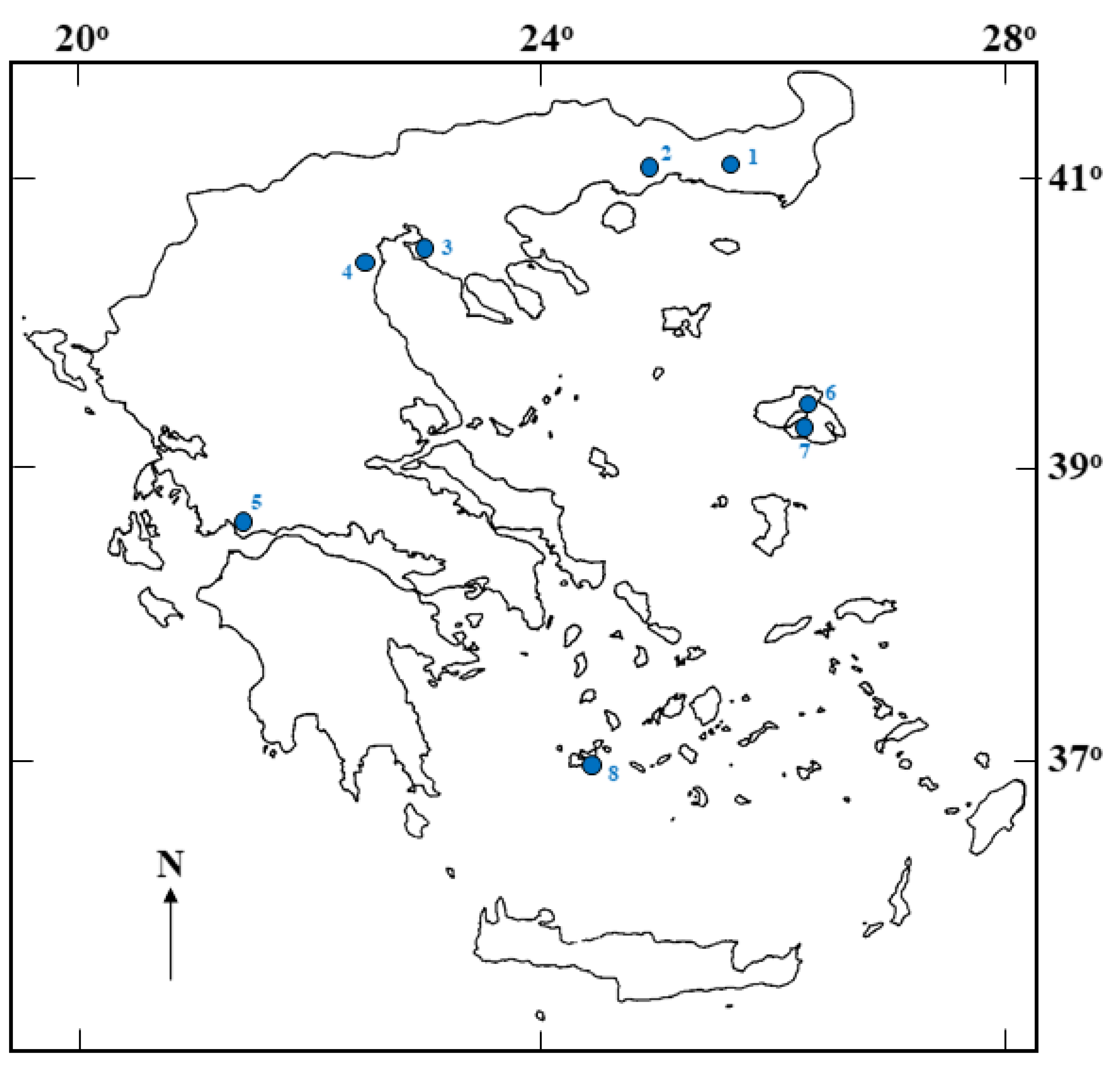
2.1. Site Description, Sampling and Strain Isolation
2.2. Polyphasic Characterization
2.3. Growth Rates and Pigments
3. Results
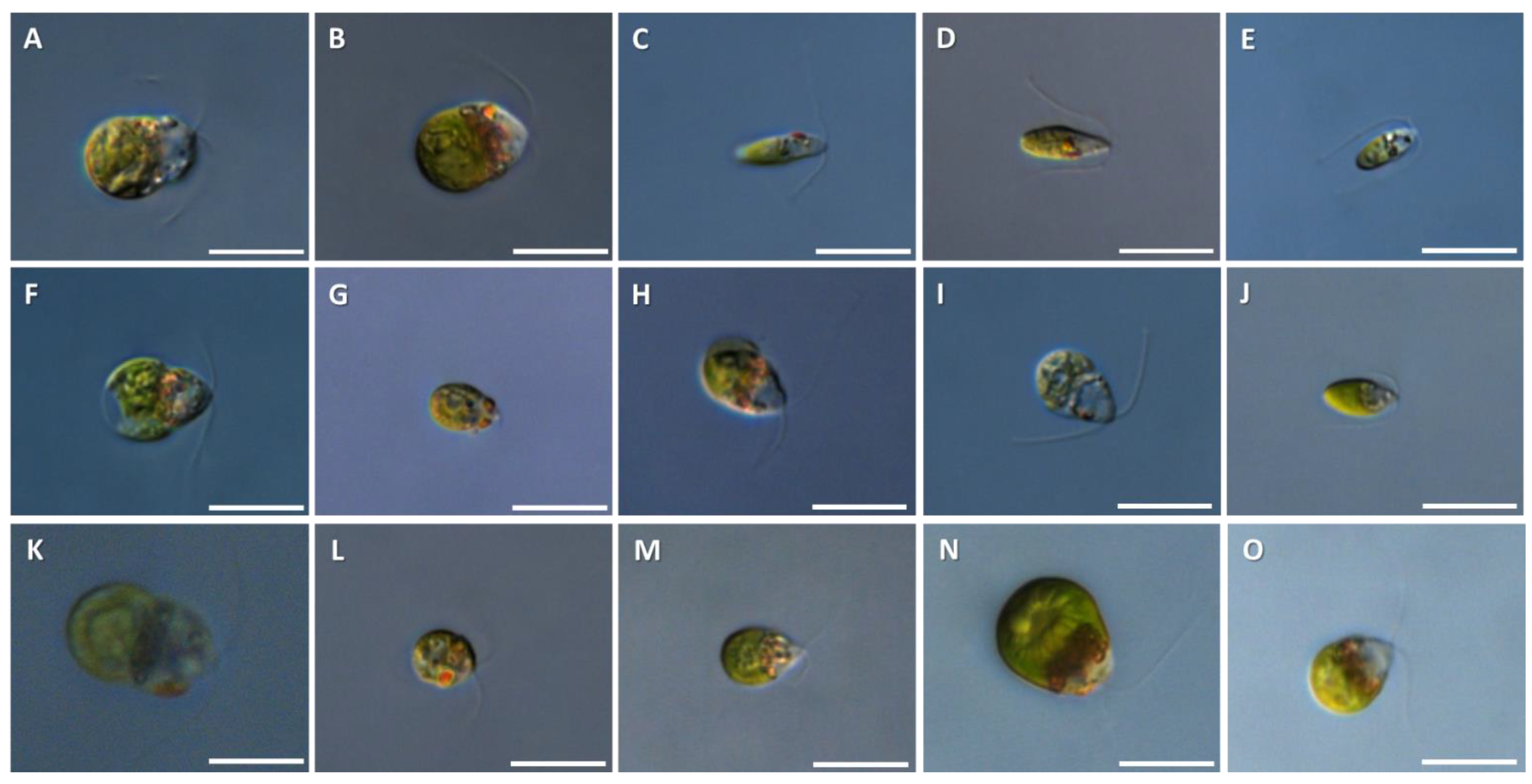
3.1. Morphological Analysis
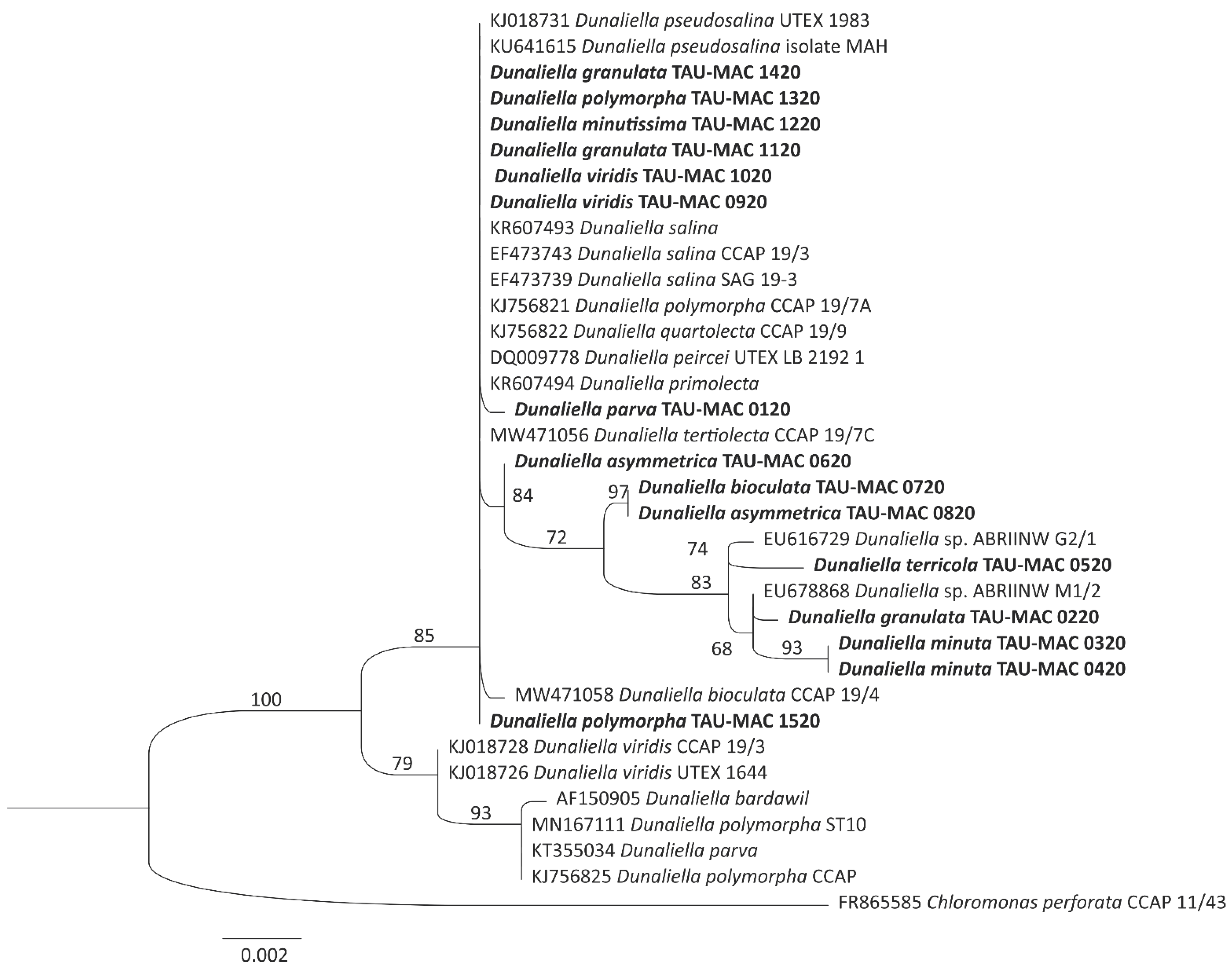
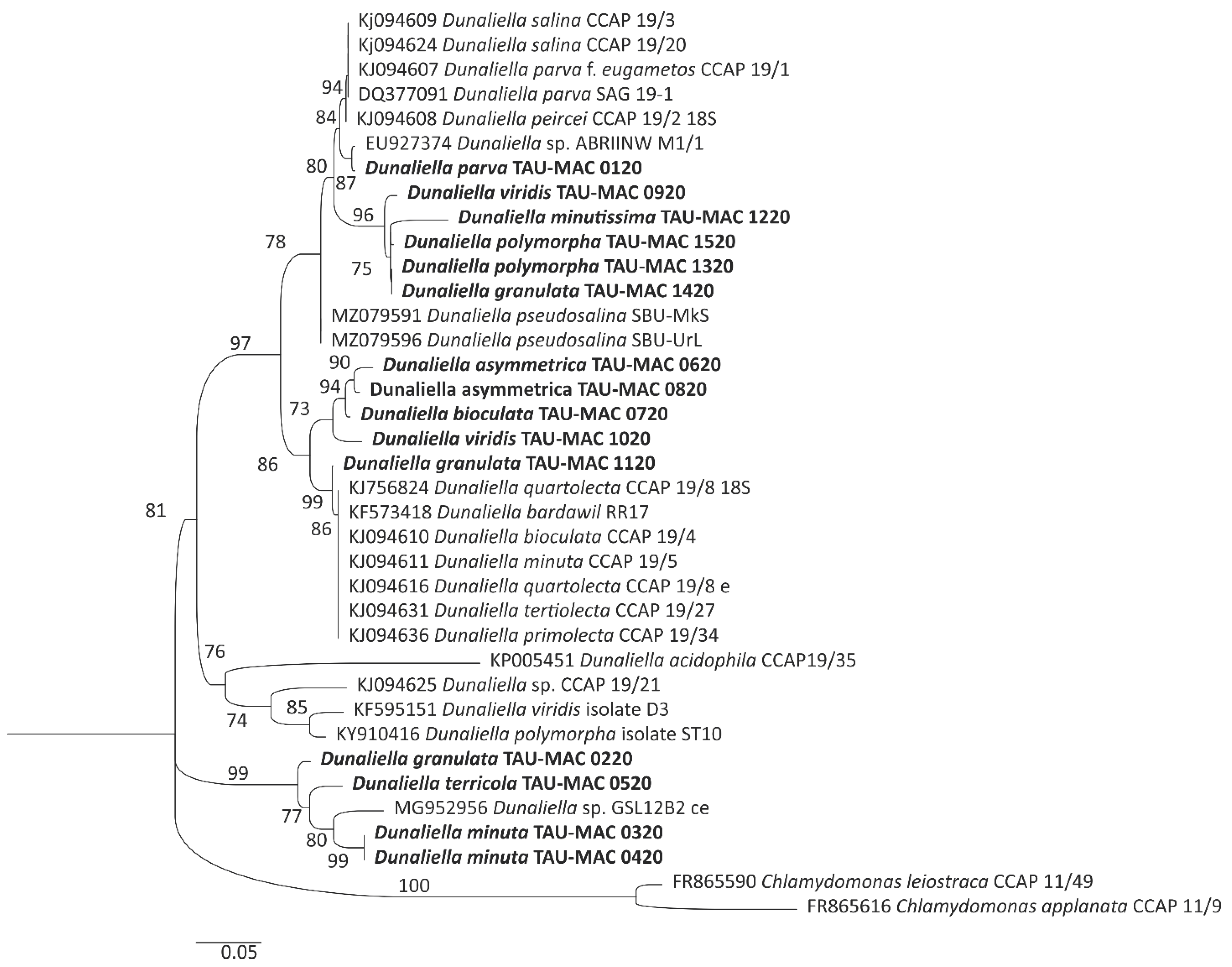
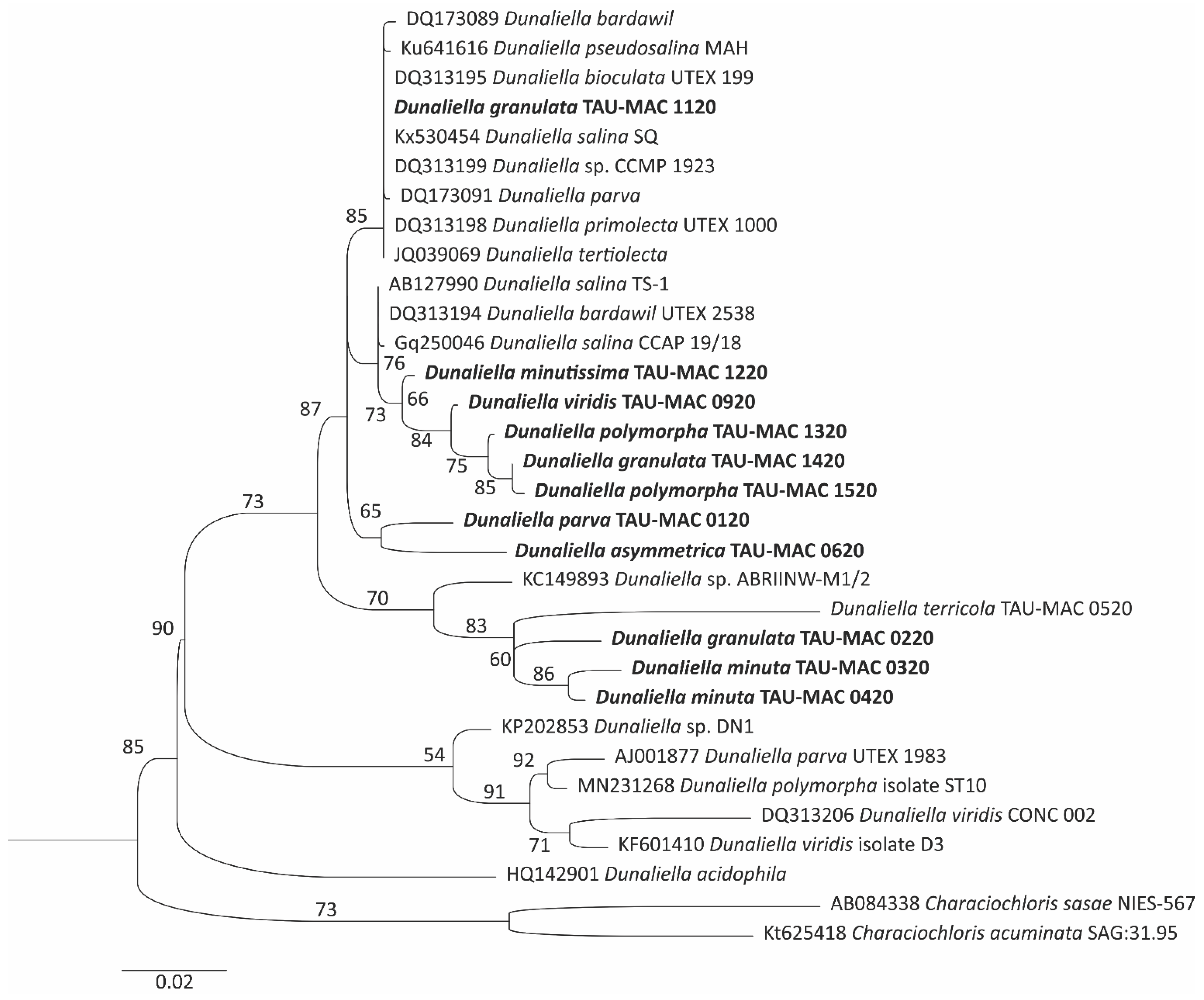
3.2. Phylogenetic Analysis
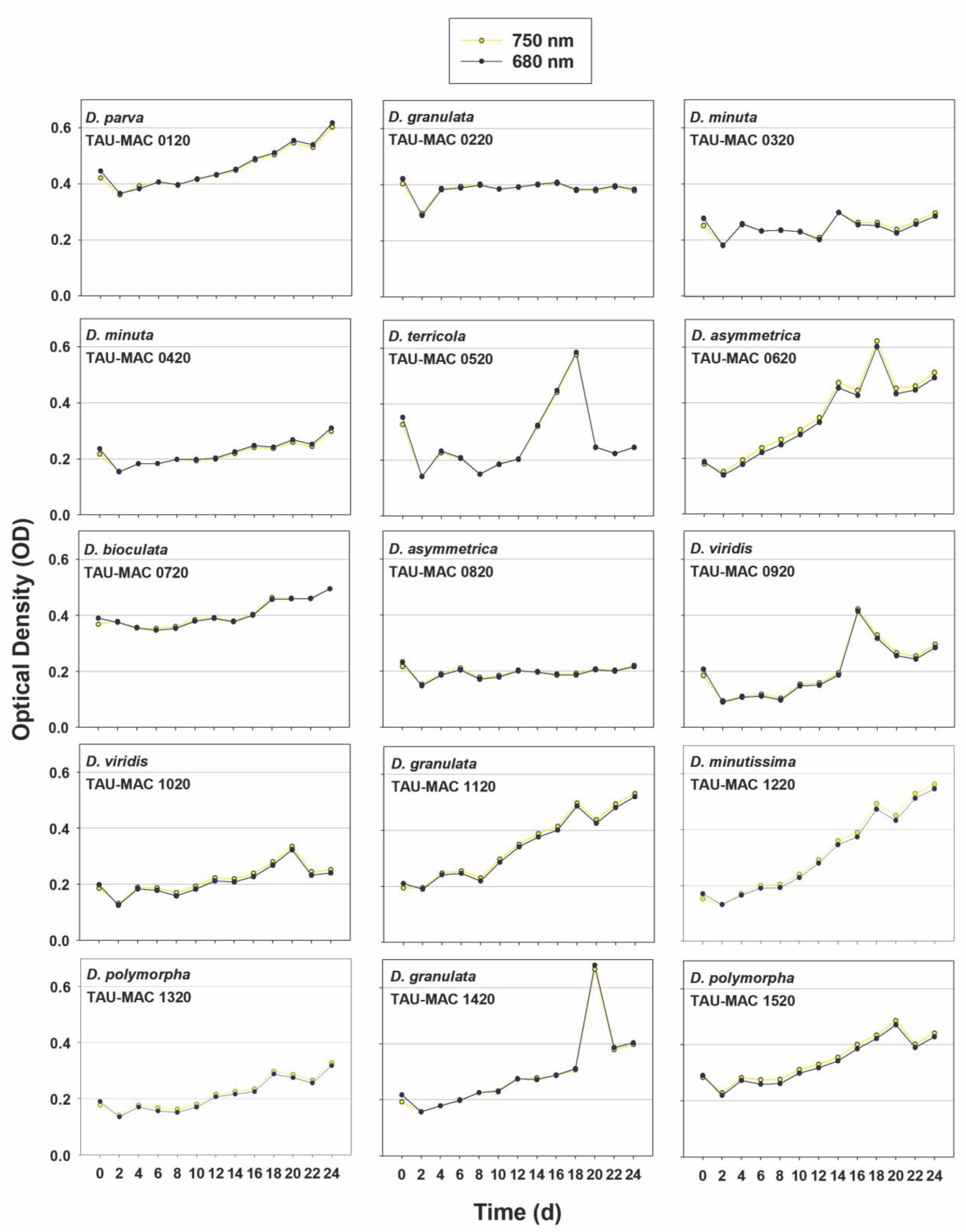
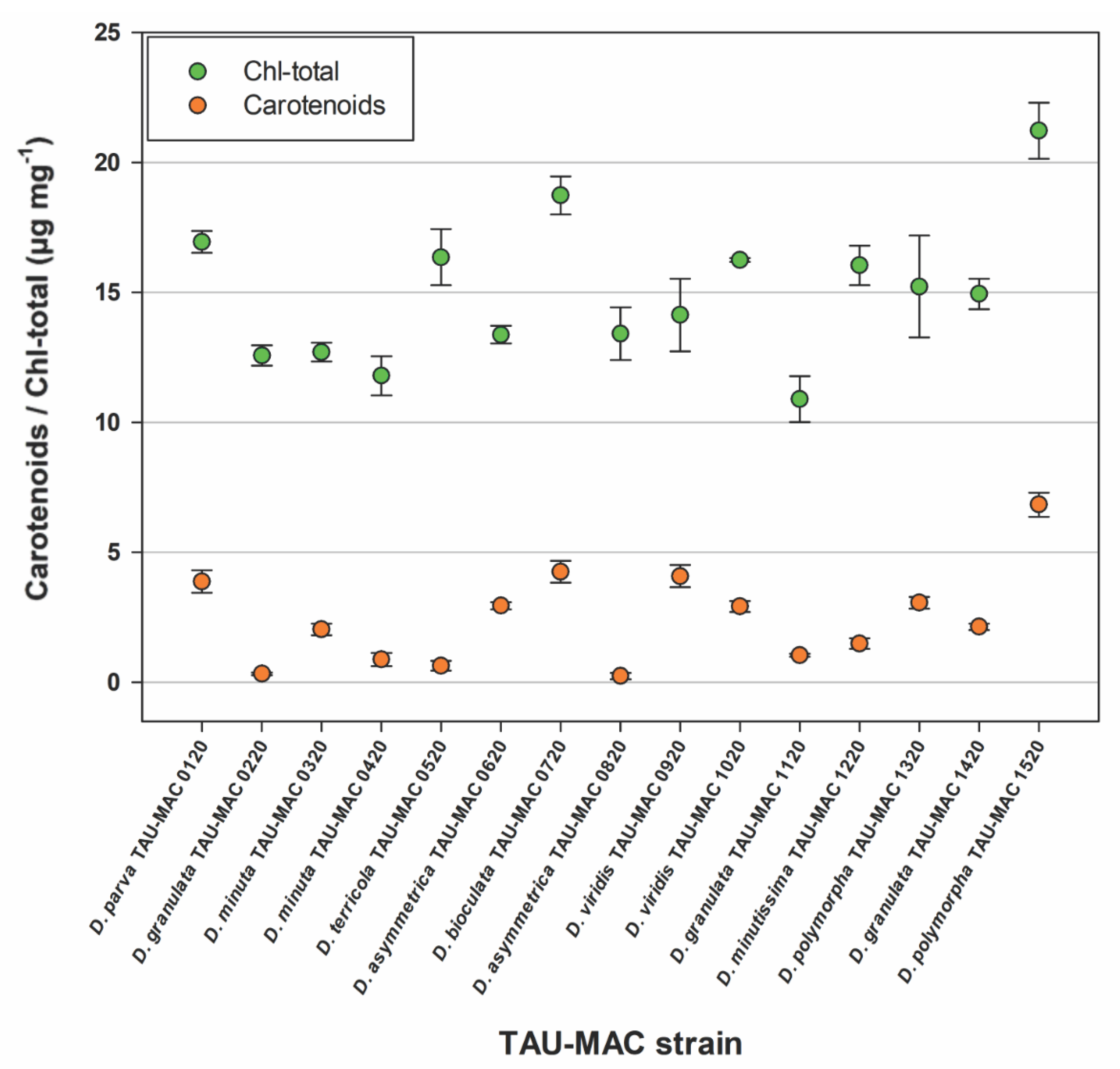
3.3. Growth Rates and Pigments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A. The microbiology of red brines. In Advances in Applied Microbiology; Academic Press Inc.: Cambridge, MA, USA, 2020; Volume 113, pp. 57–110. ISBN 9780128207093. [Google Scholar]
- Borowitzka, M.A.; Siva, C.J. The taxonomy of the genus Dunaliella (Chlorophyta, Dunaliellales) with emphasis on the marine and halophilic species. J. Appl. Phycol. 2007, 19, 567–590. [Google Scholar] [CrossRef]
- da Silva, M.R.O.B.; Moura, Y.A.S.; Converti, A.; Porto, A.L.F.; Viana Marques, D.d.A.; Bezerra, R.P. Assessment of the potential of Dunaliella microalgae for different biotechnological applications: A systematic review. Algal Res. 2021, 58, 102396. [Google Scholar] [CrossRef]
- Oren, A. The ecology of Dunaliella in high-salt environments. J. Biol. Res. 2014, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- González, M.A.; Gómez, P.I.; Polle, J.E.W. Taxonomy and Phylogeny of the Genus Dunaliella. In The Alga Dunaliella; CRC Press: Boca Raton, FL, USA, 2019; pp. 15–44. [Google Scholar]
- Henley, W.J.; Cobbs, M.; Novoveská, L.; Buchheim, M.A. Phylogenetic analysis of Dunaliella (Chlorophyta) emphasizing new benthic and supralittoral isolates from Great Salt Lake. J. Phycol. 2018, 54, 483–493. [Google Scholar] [CrossRef]
- Gao, F.; Nan, F.; Feng, J.; Lv, J.; Liu, Q.; Liu, X.; Xie, S. Comparative morphological, physiological, biochemical and genomic studies reveal novel genes of Dunaliella bioculata and D. quartolecta in response to salt stress. Mol. Biol. Rep. 2022, 49, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Highfield, A.; Ward, A.; Pipe, R.; Schroeder, D.C. Molecular and phylogenetic analysis reveals new diversity of Dunaliella salina from hypersaline environments. J. Mar. Biol. Assoc. UK 2021, 101, 27–37. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Bio, A.; Amat, F.; Vieira, N. Artisanal salt production in Aveiro/Portugal—An ecofriendly process. Saline Syst. 2011, 7, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chekanov, K. Diversity and Distribution of Carotenogenic Algae in Europe: A Review. Mar. Drugs 2023, 21, 108. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, X.; Wang, M.; Zhang, W.; Zhou, B.; Wang, Y. ROS and calcium signaling mediated pathways involved in stress responses of the marine microalgae Dunaliella salina to enhanced UV-B radiation. J. Photochem. Photobiol. B Biol. 2017, 173, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Kong, F.; Chi, Z. ROS Induce β-Carotene Biosynthesis Caused by Changes of Photosynthesis Efficiency and Energy Metabolism in Dunaliella salina Under Stress Conditions. Front. Bioeng. Biotechnol. 2021, 8, 1447. [Google Scholar] [CrossRef]
- Pourkarimi, S.; Hallajisani, A.; Nouralishahi, A.; Alizadehdakhel, A.; Golzary, A. Factors affecting production of beta-carotene from Dunaliella salina microalgae. Biocatal. Agric. Biotechnol. 2020, 29, 101771. [Google Scholar] [CrossRef]
- Gallego-Cartagena, E.; Castillo-Ramírez, M.; Martínez-Burgos, W. Effect of stressful conditions on the carotenogenic activity of a Colombian strain of Dunaliella salina. Saudi J. Biol. Sci. 2019, 26, 1325–1330. [Google Scholar] [CrossRef]
- Xi, Y.; Bian, J.; Luo, G.; Kong, F.; Chi, Z. Enhanced β-carotene production in Dunaliella salina under relative high flashing light. Algal Res. 2022, 67, 102857. [Google Scholar] [CrossRef]
- Hotos, G.N. A Preliminary Survey on the Planktonic Biota in a Hypersaline Pond of Messolonghi Saltworks (W. Greece). Diversity 2021, 13, 270. [Google Scholar] [CrossRef]
- Hotos, G.; Avramidou, D.; Mastropetros, S.G.; Tsigkou, K.; Kouvara, K.; Makridis, P.; Kornaros, M. Isolation, identification, and chemical composition analysis of nine microalgal and cyanobacterial species isolated in lagoons of Western Greece. Algal Res. 2023, 69, 102935. [Google Scholar] [CrossRef]
- Chantzistrountsiou, X.; Ntzouvaras, A.; Papadaki, S.; Tsirigoti, A.; Tzovenis, I.; Economou-Amilli, A. Carotenogenic Activity of Two Hypersaline Greek Dunaliella salina Strains under Nitrogen Deprivation and Salinity Stress. Water 2023, 15, 241. [Google Scholar] [CrossRef]
- Sili, C.; Torzillo, G.; Vonshak, A. Arthrospira (Spirulina). In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Springer: Berlin/Heidelberg, Germany, 2012; Volume 9789400738, pp. 677–705. ISBN 9789400738553. [Google Scholar]
- Waterbury, J.B.; Stanier, R.Y. Patterns of growth and development in pleurocapsalean cyanobacteria. Microbiol. Rev. 1978, 42, 2–44. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.K.; Johnson, E.J.; MacElroy, R.D.; Speer, H.L.; Bruff, B.S. Effects of salts on the halophilic alga Dunaliella viridis. J. Bacteriol. 1968, 95, 1461–1468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chitlaru, E.; Pick, U.; Physiology, S.P.; Oct, N.; Chitlaru, E.; Pick, U. Selection and Characterization of Dunaliella salina Mutants Defective in Haloadaptation Linked references are available on JSTOR for this article: Selection and Characterization of Dunaliella salina Mutants Defective in Haloadaptation. Plant Physiol. 1989, 91, 788–794. [Google Scholar] [CrossRef] [Green Version]
- De Chazal, N.M.; Smaglinski, S.; Smith, G.D. Methods involving light variation for isolation of cyanobacteria: Characterization of isolates from central Australia. Appl. Environ. Microbiol. 1992, 58, 3561–3566. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Fernández Tussy, P.; Zaoutsos, N. Isolation and preliminary characterization of cyanobacteria strains from freshwaters of Greece. Open Life Sci. 2015, 10, 52–60. [Google Scholar] [CrossRef]
- Lortou, U.; Gkelis, S. Polyphasic taxonomy of green algae strains isolated from Mediterranean freshwaters. J. Biol. Res. 2019, 26, 11. [Google Scholar] [CrossRef] [Green Version]
- Gkelis, S.; Panou, M. Capturing biodiversity: Linking a cyanobacteria culture collection to the “scratchpads” virtual research environment enhances biodiversity knowledge. Biodivers. Data J. 2016, 4, e7965-1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atashpaz, S.; Khani, S.; Barzegari, A.; Barar, J.; Vahed, S.Z.; Azarbaijani, R.; Omidi, Y. A robust universal method for extraction of genomic DNA from bacterial species. Microbiology 2010, 79, 538–542. [Google Scholar] [CrossRef]
- Medlin, L.; Elwood, H.J.; Stickel, S.; Sogin, M.L. The characterization of enzymatically amplified eukaryotic 16S-like rRNA-coding regions. Gene 1988, 71, 491–499. [Google Scholar] [CrossRef] [Green Version]
- Lane, D. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematic; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Johnson, J.L.; Fawley, M.W.; Fawley, K.P. The diversity of Scenedesmus and Desmodesmus (Chlorophyceae) in Itasca State Park, Minnesota, USA. Phycologia 2007, 46, 214–229. [Google Scholar] [CrossRef]
- Nozaki, H.; Itoh, M.; Sano, R.; Uchida, H.; Watanabe, M.M.; Kuroiwa, T. Phylogenetic relationships within the colonial Volvocales (Chlorophyta) inferred from rbcL gene sequence data. J. Phycol. 1995, 31, 970–979. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M. UGENE team Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, M.J.; Garcin, C.; van Hille, R.P.; Harrison, S.T.L. Interference by pigment in the estimation of microalgal biomass concentration by optical density. J. Microbiol. Methods 2011, 85, 119–123. [Google Scholar] [CrossRef]
- Wellburn, A.R. The Spectral Determination of Chlorophylls a and b, as well as Total Carotenoids, Using Various Solvents with Spectrophotometers of Different Resolution. J. Plant Physiol. 1994, 144, 307–313. [Google Scholar] [CrossRef]
- Preetha, K.; John, L.; Subin, C.S.; Vijayan, K.K. Phenotypic and genetic characterization of Dunaliella (Chlorophyta) from Indian salinas and their diversity. Aquat. Biosyst. 2012, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Beuzenberg, V.; Smith, K.F.; Packer, M.A. Isolation and characterisation of halo-tolerant Dunaliella strains from Lake Grassmere/Kapara Te Hau, New Zealand. N. Z. J. Bot. 2014, 52, 136–152. [Google Scholar] [CrossRef]
- Assunção, P.; Jaén-Molina, R.; Caujapé-Castells, J.; de la Jara, A.; Carmona, L.; Freijanes, K.; Mendoza, H. Phylogenetic position of Dunaliella acidophila (Chlorophyceae) based on ITS and rbcL sequences. J. Appl. Phycol. 2012, 24, 635–639. [Google Scholar] [CrossRef]
- González, M.A.; Coleman, A.W.; Gómez, P.I.; Montoya, R. Phylogenetic relationship among various strains of Dunaliella (Chlorophyceae) based on nuclear its rDNA sequences. J. Phycol. 2001, 37, 604–611. [Google Scholar] [CrossRef]
- Assunção, P.; Jaén-Molina, R.; Caujapé-Castells, J.; Wolf, M.; Buchheim, M.A.; de la Jara, A.; Freijanes, K.; Carmona, L.; Mendoza, H. Phylogenetic analysis of ITS2 sequences suggests the taxonomic re-structuring of Dunaliella viridis (Chlorophyceae, Dunaliellales). Phycol. Res. 2013, 61, 81–88. [Google Scholar] [CrossRef]
- Polle, J.E.W.; Struwe, L.; Jin, E. Identification and characterization of a new strain of the unicellular green alga Dunaliella salina (Teod.) from Korea. J. Microbiol. Biotechnol. 2008, 18, 821–827. [Google Scholar] [PubMed]
- Hejazi, M.A.; Barzegari, A.; Gharajeh, N.H.; Hejazi, M.S. Introduction of a novel 18S rDNA gene arrangement along with distinct ITS region in the saline water microalga Dunaliella. Saline Syst. 2010, 6, 4. [Google Scholar] [CrossRef] [Green Version]
- Dolapsakis, N.P.; Tafas, T.; Abatzopoulos, T.J.; Ziller, S.; Economou-Amilli, A. Abundance and growth response of microalgae at Megalon Embolon solar saltworks in northern Greece: An aquaculture prospect. J. Appl. Phycol. 2005, 17, 39–49. [Google Scholar] [CrossRef]
- Hotos, G.N.; Avramidou, D. The Effect of Various Salinities and Light Intensities on the Growth Performance of Five Locally Isolated Microalgae [Amphidinium carterae, Nephroselmis sp., Tetraselmis sp. (var. red pappas), Asteromonas gracilis and Dunaliella sp.] in Laboratory Batch Cultures. J. Mar. Sci. Eng. 2021, 9, 1275. [Google Scholar]
- Al-Mhanna, N.; Pistorius, M.; Al Sammarraie, L. Optimization of the Cultivation Conditions of the Green Algae Dunaliella salina by Using Simplex Method. Processes 2023, 11, 292. [Google Scholar] [CrossRef]
- Gan, S.; Liang, S.; Zou, Q.; Shang, C. Optimization of carotenoid extraction of a halophilic microalgae. PLoS ONE 2022, 17, e0270650. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, X.; Luo, L.; Shang, C. Optimization of Extraction Conditions of Carotenoids from Dunaliella parva by Response Surface Methodology. Molecules 2022, 27, 1444. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, M.M.S.; El-Ayouty, Y.M.; Fathey, H.A. Disparity of the carotenoids antioxidant properties of wild-type and D-PSY-transgenic Dunaliella parva strains under three environmental stresses. Physiol. Mol. Biol. Plants 2021, 27, 2151. [Google Scholar] [CrossRef] [PubMed]
- Ismaiel, M.M.S.; El-Ayouty, Y.M.; Said, A.A.; Fathey, H.A. Transformation of Dunaliella parva with PSY gene: Carotenoids show enhanced antioxidant activity under polyethylene glycol and calcium treatments. Biocatal. Agric. Biotechnol. 2018, 16, 378–384. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain (TAU-MAC) | Isolation Site | Geographic Coordinates (N, E) | Collection Date |
|---|---|---|---|
| 0720 | Nea Kessani | 41.019039, 25.077181 | 23 January 2020 |
| 0820 | 26 May 2020 | ||
| 1520 | 10 September 2020 | ||
| 0120 | Mesi | 40.973328, 25.220879 | 23 January 2020 |
| 0220 | 23 January 2020 | ||
| 1320 | 10 September 2020 | ||
| 1220 | Angelochori | 40.491461, 22.821933 | 9 September 2020 |
| 0520 | Kitros | 40.374210, 22.634911 | 17 January 2020 |
| 1420 | 9 September 2020 | ||
| 0920 | Kalloni | 39.213665, 26.252675 | 23 June 2020 |
| 0620 | Polichnitos | 39.114962, 26.176667 | 23 February 2020 |
| 0320 | Messolonghi | 38.399854, 21.415901 | 27 January 2020 |
| 0420 | 27 January 2020 | ||
| 1020 | 12 June 2020 | ||
| 1120 | Adamas, Milos | 36.711771, 24.467830 | 26 August 2020 |
| Primer | Target Gene-Region | Sequence (5′-3′) | Size (bp) | Reference | Conditions |
|---|---|---|---|---|---|
| EukA | 18S rRNA | AACCTGGTTGATCCTGCCAGT | 1750 | [28] | Initial denaturation step at 95 °C for 5 min, 35 cycles consisting of denaturation at 95 °C for 60 s, annealing at 55 °C for 60 s and elongation at 72 °C for 90 s; a final 7-min elongation step at 72 °C was included. |
| EukB | TGATCCTTCTGCAGGTTCACCTAC | ||||
| U1391R | GGGCGGTGTGTACAARGR * | [29] | |||
| ITS-AF | ITS | CGTTTCCGTAGGTGAACCTGC | 700 | [30] | Initial denaturation step at 94 °C for 4 min, 35 cycles consisting of denaturation at 95 °C for 60 s, annealing at 58 °C for 2 min and elongation at 72 °C for 2 min; a final 7 min elongation step at 72 °C was included. |
| ITS-BR | CATATGCTTAAGTTCAGCGGG T | ||||
| rbcL1-20 | rbcL | ATGGTTCCACAAACAGAAAC | 1100 | [31] | |
| rbcL1181-1160 | AAGATTTCAACTAAAGCTGGCA |
| Strain (TAU-MAC) | Taxonomic Assignment | Cells Length and Width (μm) | Morphology |
|---|---|---|---|
| 0120 | D. parva | l: 10–13 w: 6–9 | Cells: green (faint or pale), oval to cylindrical, radially symmetric with widely rounded posterior and slightly narrowed anterior. Flagella: length slightly longer than the cell. Stigma small, red and distinctive. Chloroplast: slightly cup-shaped, with poorly developed lateral lobes that do not reach the anterior end of the cell. Figure 2A |
| 0220 1120 1420 | D. granulata | l: 10–13 w: 6–9 | Cells: intense green, broad oval, ovoid or slightly pyriform, wide and rounded at the posterior end and gradually narrowed and rounded at the anterior end. Chloroplast: cup-shaped with poorly developed lateral lobes, always granulated on the edge. Constant presence of a ring of dark granules at the anterior edge of the chloroplast. Stigma: large, very distinct, protuberant. Figure 2B,K,N |
| 0320 0420 | D. minuta | l: 8–11 w: 2.5–4.5 | Cells: green, narrow-cylindrical, oval or elliptical, usually with rounded anterior and posterior ends. Flagella: length slightly longer than cell length, often with the one of the two flagellums to be shorter than the other. Anterior large elongated rod-shaped, narrow, bright red stigma. Vegetative cysts 7–9 μm diameter. Figure 2C,D |
| 0520 | D. terricola | l: 5.5–9 w: 2–3.5 | Cells: green, wide and rounded at the anterior end, narrow towards the posterior end, elongate spindle shaped, sometimes slightly bent. Flagella: 1.5–2 times cell length. Cup-shaped chloroplast with thin lateral lobes reaching nearly to the cell apex. Pyrenoid axial and central, usually spherical. Apical stigma, usually small. Figure 2E |
| 0620 0820 | D. asymmetrica | l: 8–12 w: 6–9 | Cells: green, asymmetrical, irregularly dorsiventral. They are irregularly ovoid, pyriform from the dorsal side, oval or ellipsoid with widely rounded posterior end and slightly narrowed anterior end. Flagella are slightly longer than the cell, oriented forward or slightly to the side during motion. The cup-shaped chloroplast is usually fragmented at the anterior edge. Pyrenoid is basal, large and is sometimes slightly asymmetrical in form and position. Large stigma, usually located on the ventral side of the cell. Figure 2F,H |
| 0720 | D. bioculata | l: 7–10 w: 3.5–6.5 | Cells: green, oval, ovoid, cylindrical or nearly spherical, wide from the posterior end and gradually narrowing towards the anterior end, with both ends rounded. Flagella (not obvious in the Figure 2G) is 1.5–2 times the cell length. Cup-shaped chloroplast with wide lateral lobes which do not reach the anterior end of the cell. Pyrenoid is axial or basal, spherical or elongated with a distinct amylosphere. Two distinct stigmata at the anterior end of the cell. Figure 2G |
| 0920 1020 | D. viridis | l: 7–11 w: 3.5–7 | Cells: green, pyriform, ellipsoid, oval, ovoid or spindle shaped. Flagella: about 1.3–1.5 times cell length. Chloroplast: cup-shaped with lateral lobes not reaching anterior end. Anterior end of cell is usually transparent. Stigma large, usually elongated and ellipsoidal. Figure 2I,J |
| 1220 | D. minutissima | d: 5–7.0 | Cells: green, spherical, 5–7.0 μm in diameter. Flagella: longer than the cell. Stigma large and distinct. Figure 2L |
| 1320 1520 | D. polymorpha | l: 8–10 w: 5–7 | Cells: green, radially symmetrical, mostly ellipsoidal or pyriform. Flagella: length about 1.5 times the cell length. Stigma: small and medial. Linear or U-shaped zone of irregular refractile granules at anterior sinus of chloroplast. Figure 2M,O |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lortou, U.; Panou, M.; Papapanagiotou, G.; Florokapi, G.; Giannakopoulos, C.; Kavoukis, S.; Iakovou, G.; Zalidis, G.; Triantafyllidis, K.; Gkelis, S. Beneath the Aegean Sun: Investigating Dunaliella Strains’ Diversity from Greek Saltworks. Water 2023, 15, 1037. https://doi.org/10.3390/w15061037
Lortou U, Panou M, Papapanagiotou G, Florokapi G, Giannakopoulos C, Kavoukis S, Iakovou G, Zalidis G, Triantafyllidis K, Gkelis S. Beneath the Aegean Sun: Investigating Dunaliella Strains’ Diversity from Greek Saltworks. Water. 2023; 15(6):1037. https://doi.org/10.3390/w15061037
Chicago/Turabian StyleLortou, Urania, Manthos Panou, Georgia Papapanagiotou, Georgia Florokapi, Christos Giannakopoulos, Savvas Kavoukis, Georgios Iakovou, Giorgos Zalidis, Konstantinos Triantafyllidis, and Spyros Gkelis. 2023. "Beneath the Aegean Sun: Investigating Dunaliella Strains’ Diversity from Greek Saltworks" Water 15, no. 6: 1037. https://doi.org/10.3390/w15061037