
The Effects of Channelization with Low In-Stream Barriers on Macroinvertebrate Communities of Mountain Rivers

1
Institute of Nature Conservation, Polish Academy of Sciences, Adama Mickiewicza 33, 31-120 Krakow, Poland
2
Faculty of Geology, Geophysics and Environmental Protection, University of Science and Technology AGH, Adama Mickiewicza Av.30, 30-059 Krakow, Poland
3
Polish Hydrobiological Society, Krakow Branch, Adama Mickiewicza 33, 31-120 Krakow, Poland
*
Author to whom correspondence should be addressed.
Water 2023, 15(6), 1059; https://doi.org/10.3390/w15061059
Submission received: 20 January 2023
/
Revised: 27 February 2023
/
Accepted: 7 March 2023
/
Published: 10 March 2023
(This article belongs to the Special Issue Monitoring of Immediate and Long-Term Effects of River Restoration Projects)
Abstract
:The effects of channelization with low in-stream barriers in mountain rivers on macroinvertebrate communities are still weakly recognised. We aimed to assess the differences in structure and density of benthic macroinvertebrates between two north Carpathian mountain rivers: one channelized with a cascade of concrete sills and the other one with block ramps. Water was collected five times for physicochemical analysis. Macroinvertebrate fauna and fine bottom sediments were collected three times (spring, summer, autumn) from different habitat types: glides, runs, pools and riffles of the estimated channel surface. The in-stream barriers altered the channel gradient and flow velocity, reducing the area of pools and riffles as well as extending the area of glides with slow current velocities, associated with the highest rate of fine sediment deposition. Channel modifications induced a general shift of investigated fluvial systems toward lentic habitat conditions. Macroinvertebrate communities characteristic of slow-flowing rivers and pools became dominant, while those characteristic for the riffles of mountain rivers were confined to only small areas. The studied section of the Porębianka River channelized with block ramps exhibited greater diversity in the macroinvertebrate community between habitats and significantly higher macroinvertebrate density than the Mszanka River which was channelized with concrete sills.
1. Introduction
Channelization involves engineering work aimed at the modification of a river’s slope, width, depth, or transported sediment load. It can be achieved by a number of methods which vary depending on the hydrology and the type of a river reach [1]. Typically, channelized rivers differ from natural ones in their more uniform width and depth, regular bend geometry and a bank lining reducing lateral erosion. Moreover, channelized river reaches are stabilized vertically by grade control structures in order to reduce river downcutting [2] and slow down the movement of the bed load [3]. In mountain streams, where the processes of bed erosion are particularly active, these in-stream barriers are widespread [4]. They have become the most striking feature of channels in densely inhabited valleys where natural sediment transport and flow hydraulics are imbalanced due to channelization [5].
It has been widely accepted that habitat homogenisation in river channels due to channelization works can, among other environmental stressors, significantly reduce invertebrate density [6,7,8]. Macroinvertebrates can respond to channelization with a decrease of specialisation level associated with a decrease of species with narrow environmental tolerance [9], the expansion of lentic species in impounded river sections upstream of in-stream barriers [10], or reduced longitudinal connectivity even by small in-stream barriers [11]. Bank modifications also can affect aquatic macroinvertebrates by reducing their abundance [12], species richness, or proportion of individual feeding groups [13].
To overcome the negative effects of channelization, efforts toward the restoration of pristine river conditions are often undertaken. Unfortunately, according to the numerous works available, only one third of the studies found positive effects of increasing habitat heterogeneity on invertebrate species richness due to channel reconfiguring with meanders, bars, boulders etc. [14]. Most of them however, found more complicated relationships controlling biodiversity, such as the scale of the riparian zone and upstream river sections transformation [15], a specific substrate type [16], or nutrient load [17]. A considerable number of restoration projects also deal with in-stream barriers of different dimensions from dams, low-head dams, or weirs, to check dams, which are removed in order to facilitate the passage of fish and to provide a riverbed habitat for macroinvertebrates [18,19]. The overall significance of these works for macroinvertebrate richness and abundance is ambiguous. Larger barriers predominantly affect the macroinvertebrate community in a more than moderate degree [20] and their removal leads, in principle, to an increase in aquatic biodiversity [21]. The effects of functioning and restoring channelized river reaches with low in-stream barriers, e.g., check dams or sills, are considered to be insignificant for macroinvertebrate richness [22]. Generally, the role of channelization with check dams or sills as low in-stream barriers in the modification of invertebrate heterogeneity is considered to be small and neglected in investigations and the possible mitigation of low in-stream barrier effects are not recognised.
In our study, we aimed to demonstrate the impact of channelization with low in-stream barriers on the structure and density of benthic macroinvertebrate in two small mountain rivers. One of the rivers was channelized with a cascade of concrete, 1 m high sills, whereas the other was modified with dozens of block ramps forming chutes of a similar elevation of 1 m. Each chute was designed as an environmentally friendly structure substituting concrete sills to ensure river continuity over the ca. 3.5 km reach [23]. Both constructions were applied at the dawn of the 21st century for channels heavily affected by earlier river training works.
2. Materials and Methods
2.1. Study Area
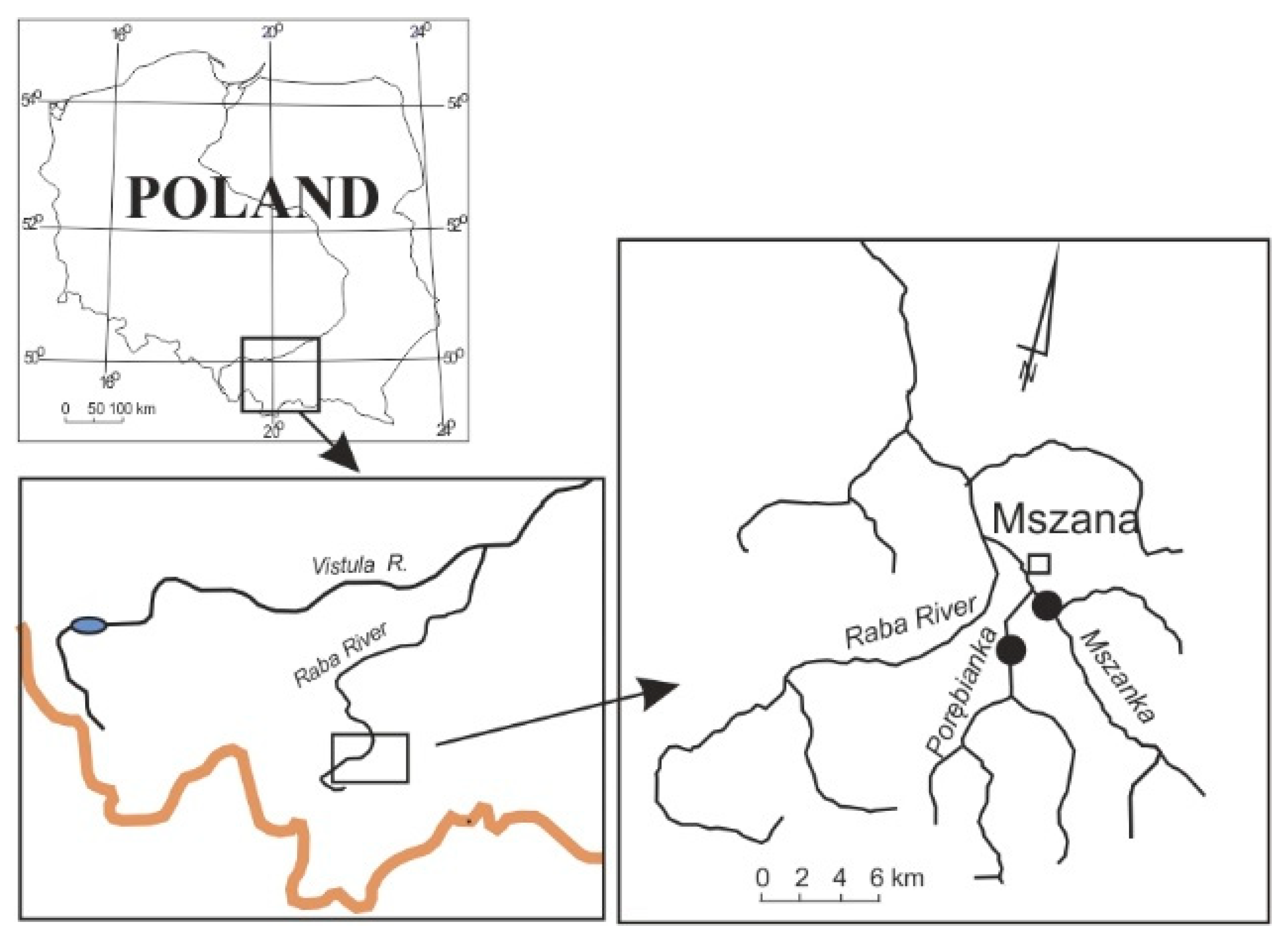
The Porębianka (15.4 km) and Mszanka (19.5 km) rivers are Carpathian tributaries of the Raba River, a right-bank tributary of the Vistula River, in southern Poland (Figure 1 and Figure 2). The catchment basins of both rivers equals 175 km2 and the mean annual discharge downstream of their confluence is 3.3 m3/s [2]. The upper slopes of river catchments are mostly forested and representative of the relief of middle mountains built of flysch with elevations reaching 1300 m asl. The lower parts of the catchments, 700–400 m asl, are densely inhabited and utilised as meadows and cultivated land. Average annual precipitation is 1000 mm, peaking in the summer months.
Prior to channelization in 1959, the Mszanka River was a braided, gravel bed river with a width exceeding 100 m. A debris dam in the middle section of the river and extensive gravel mining with some bank protection since the 1960s induced rapid channel incision, which exceed even 1.5 m, and its narrowing by even 75% over the following years [24]. To limit the incision, the ca. 6-km-long lower reach of the river was engineered with hydraulic drop control structures between the years 1977–2003. The cascade of 25 concrete 1-m-high sills reduced the channel gradient between the consecutive barriers to the designed value 0.004 (Figure 2). Channel width was reduced by even 65% to the uniform value, 30–34 m, and the channel bar surface decreased by 100 times; banks were lined with boulders. Each sill was accompanied by a concrete, ca. 14-m-long stilling basin terminated with a low, ca. 0.3 m high sill, designed to slow down water velocity and reduce stream power [2]. Since the time of construction, the channel bedload started to deposit behind drop structures over 100–150-m-long sections, raising the channel bed to the top of the consecutive sills (Figure 2). Because the channel width was oversized, the river started to deposit narrow side bars which constrained the active channel [24].
The Porębianka is a gravel bed river with a predominance of sandstone pebbles and cobbles characterized by an average diameter ranging between 40 and 65 mm [23]. The river was trained in its middle and lower reaches in the 1960s and 1970s with groynes several metres long for lateral channel stabilization. The concentration of the flow in the middle of the channel and progressive filling of the intergroyne basins with coarse sediment induced river narrowing and incision into alluvia. The large flood of 1970 significantly destroyed the groynes and resulted in channel braiding with an increase in width of up to 150% [5]. In the following years, the channel was trained occasionally to protect a nearby road but after the large floods of 1997 and 2001, a new scheme for channelization was designed and implemented over the next 10 years. In the lower, 3.3 km-long reach, a cascade of 25 boulder ramps was built. The width of the channel was designed at a constant value of 28 m whereas the height of each ramp was set at ca. 1 m. The overall channel gradient was reduced from 1.25 to 0.55 promile [25]. Each ramp was designed as an environmentally friendly structure mimicking natural rapids (Figure 2). In order to facilitate the migration of fish and invertebrates at even low discharges, a 20 cm depression was constructed in the central part of each ramp and over the whole 12 m of the ramp length. Ramps were constructed of boulders ca 1 m in diameter and the slope of the whole ramp bed is 0.083. They end in a stilling basin which is 5 m long and approximately 1 m deep. These basins are characterised by intensive erosion during high water discharge, but the eroded material accumulates immediately downstream, forming central or side bars [25].
2.2. Methods
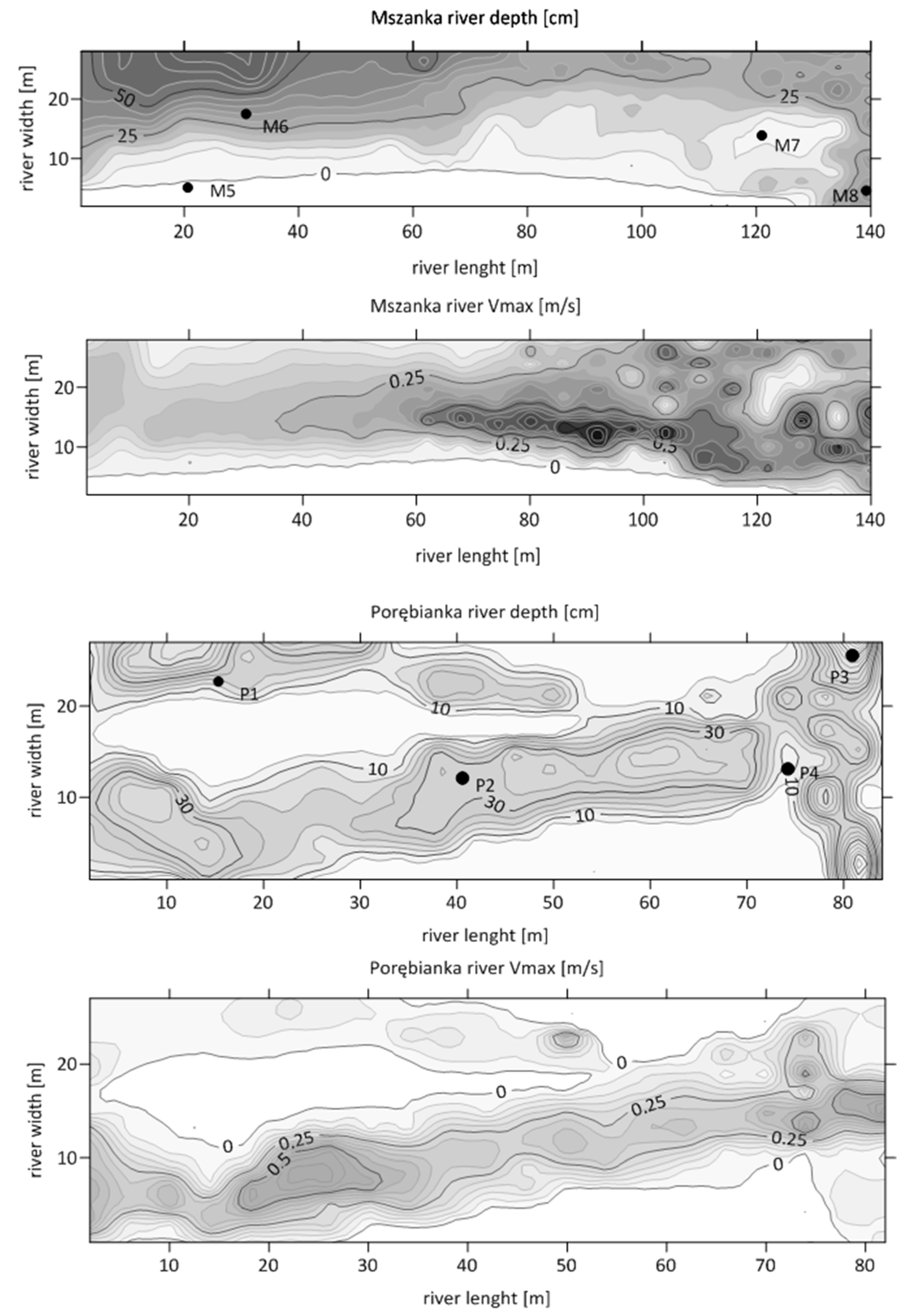
For detailed study, we selected two river segments limited by constructed barriers: one in the lower reach of the Mszanka River, constructed at the end of 1980s, and the other on the Porębianka River, constructed at the beginning of the 21st century (Figure 1). In September 2021, at low average discharges, in each of the two river segments, we performed measurements of water depth and maximum flow velocities in a net 4 × 2 m on the Porębianka and 6 × 4 m on the Mszanka river. We constructed maps of flow velocity and water depth and identified 4 habitat types to be sampled and defined according to [26]: Type 1—Glides (with shallow/slow moving water), sites P1 and M1; Type 2—Runs (with moderately fast flow), sites P2 and M2; Type 3—Pools (deep/low flow velocities or no flow with fine substrate), sites P3 and M3; Type 4—Riffles (with shallow/fast flowing water and with a coarse substrate) (P4 and M4); they are presented in Table 1 and Figure 3. The share of the particular habitats in each river was estimated (Table 1).
Benthic macroinvertebrate was collected in each river segment at low average discharges on three occasions: in the autumn (September 2021), spring (March 2022) and summer (August 2022). The density and biodiversity of macroinvertebrate fauna were analysed for the four selected habitats of the Porębianka (P) and Mszanka (M) rivers (Figure 3, Table 1) varying in flow velocity and depth. Three subsamples were collected from every habitat type, on each occasion (in total, 36 subsamples from each river). The samples of benthic macroinvertebrates were taken with a hand net, 22.5 cm in diameter, covered with a 0.3 mm mesh bolting cloth. The obtained material was preserved in 4% formalin. The larvae were selected, identified to higher taxonomic units, while Diptera were classified according to family, and counted in the laboratory. The density of benthic fauna in the sample was calculated per 1 m2 of the substratum. The presented macroinvertebrate densities for every habitat type (sites 1–4) are given as average values from three subsamples taken on the three sampling occasions.
Simultaneously, at each occasion in each habitat type, the selected parameters of water (flow velocity, water temperature, dissolved oxygen, and oxygen saturation) were measured in situ. In addition, at the same time and additionally in June 2021 and 2022, water samples were collected from the runs (P2 and M2) for the analysis of physicochemical parameters (pH, conductivity, main anions Cl−, SO42−, HCO3− and cations Ca2+, Mg2+, Na+, K+ as well as nutrients NO3−, PO43−, NH4+). Analysis was carried out in the laboratory within 24 h of sampling.
Water temperature, dissolved oxygen and oxygen saturation were measured using a HQ40D multimeter (HACH Lange GmBH, Düsseldorf, Germany), while the flow was gauged using a flowmeter (MiniAir2, Schiltknecht Messtechnik AG, Gossau ZH, Switzerland). The pH and conductivity were measured with a WTW (Multi 340i/SET2, WTW Wissenschaftlich-Technische Werkstatten GmbH, Weilheim, Germany) apparatus. Anions (Cl−, SO42−, HCO3−, NO3−, PO43−) and cations (Ca2+, Mg2+, Na+, K+, and NH4+) were analysed by ion chromatography (DIONEX ICS 1000 and IC DX 320, Dionex Corporation, Sunnyvale, CA, USA). Before chromatography analysis, water samples were filtered through pore-sized syringe filters (Minisart RC 25, Sartorius Stedim Biotech GmbH, Göttingen, Germany) into polyethylene sample tubes.
Samples for fine sediment and organic matter were taken from surface and subsurface layers (up to a depth of 10 cm) into 50 mL vessels during the agitation of water within a Walling`s cylinder inserted into the riverbed, according to the method described in [27]. Samples were then dried, weighted, and fine sediment deposition was calculated in mg/m2. The content of organic matter was expressed as losses on ignition (LOI) after sample ignition at 500 °C for 3 h.
2.3. Statistics
The differences in the values of physicochemical parameters in water between the Porębianka and Mszanka rivers were calculated using the Wilcoxon test. To study the relationship between the values of the environmental variables and the density of the dominant group of benthic macroinvertebrates, Spearman correlations were used.
The mean density of benthic macroinvertebrates in the studied rivers sections were calculated according to the formula of [28]:
where QI, QII, QIII, and QIV denote the mean density of benthic macroinvertebrates in different habitats (Types 1–4), while %I, %II, %III, and %IV represent the percentage of the different habitats in a studied river section.
QM = (QI × %I + QII × % II + QIII × %III + QVI × %IV)/100
The similarity in the density and structure of benthic macroinvertebrate communities at the studied sites/habitats of the Porębianka and Mszanka rivers were classified according to hierarchical cluster analysis. The Euclidean distance and within-groups linkage were used as a grouping method. Dendrograms of similarities were obtained, on the basis of which groups of habitats with similar benthic macroinvertebrates communities were separated. We used STATISTICA 13.1 software for the statistical analyses (Statsoft Inc., Tulsa, OK, USA).
3. Results
3.1. Environmental Parameters
Throughout the study period, the waters of the Porębianka and Mszanka rivers were of overall good quality (Table 2). The water of both rivers was slightly alkaline, and their chemical composition was dominated by the presence of ions of HCO3− and Ca2+. Despite their similar quality, values of conductivity and concentrations of major ions in the Mszanka river (SO42−, HCO3−, Mg2+, Ca2+, Cl−, Na+, and K+) were higher than in the Porębianka river (Table 3). Particularly, the content of Na+ and Cl− ions was up to two times higher in the Mszanka than in the Porębianka. There were no significant differences in the concentrations of nutrients between the studied rivers. However, the slightly higher mean concentrations of NO3−, NH4+, and PO43− found in the Porębianka than in the Mszanka might suggest its accidental pollution.
Water temperature in the sampled habitat types (1–4) of both rivers varied over the studied period in a wide range, from 3.8 to 18.3 °C. The dissolved oxygen content (8.5–13.6 mg/L) and oxygen saturation (91.9–150.3%) were high in all sampled sites (Table 2). In the Porębianka River, the mean flow velocities in glides (type 1) and riffles (type 4) were lower by one third than in the Mszanka River but they were higher in runs (type 2) by even two times.
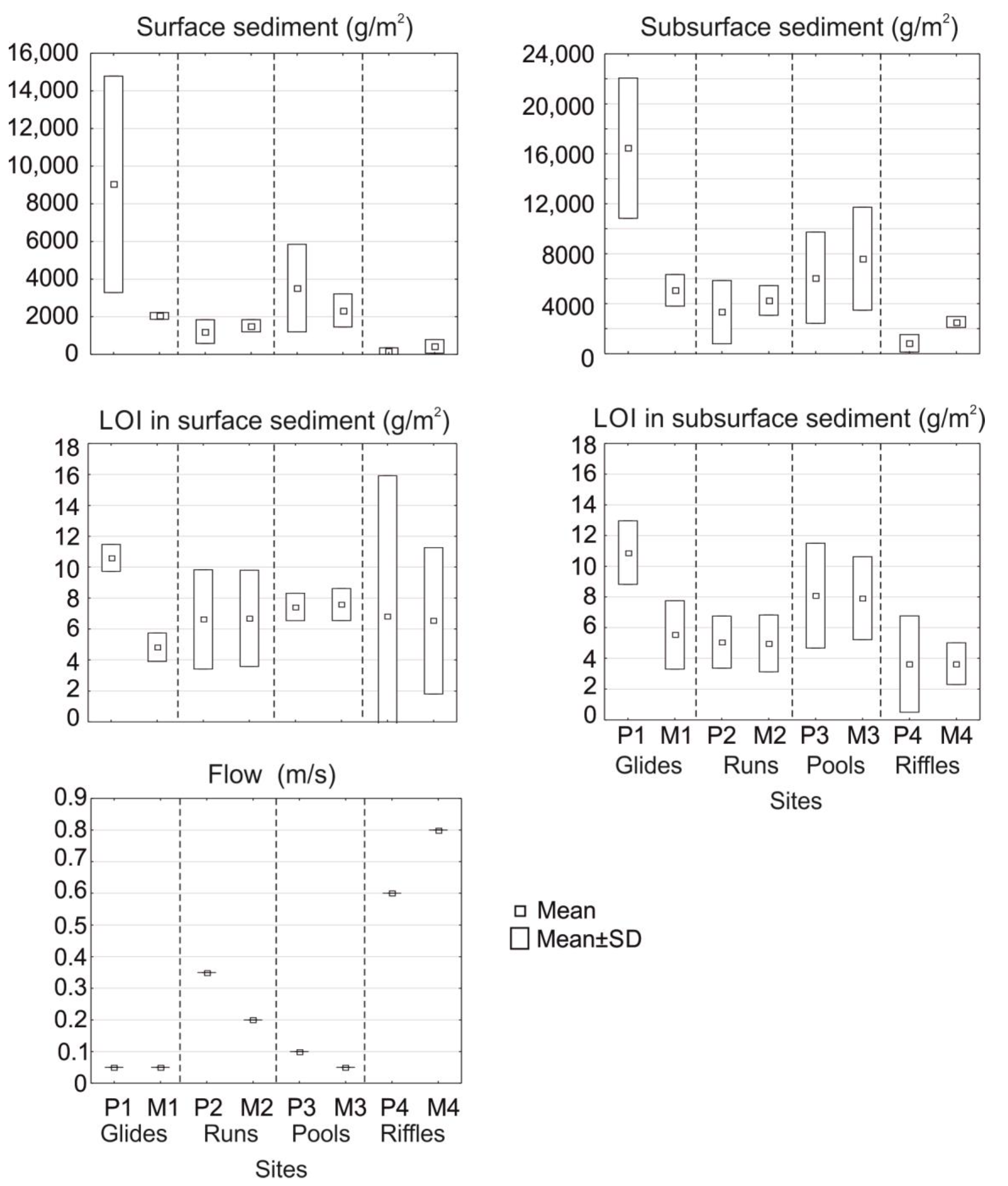
Contents of fine sediment in surface and subsurface layers in the sampled sites ranged between 70 and 15,376 g/m2 and 70–22,544 g/m2, respectively (Figure 4). Differences correspond to average flow velocities: the lowest contents were observed at high flow values in riffles (type 4) and in runs (type 2) and much higher contents of fine sediment were at low velocities in glides at river banks (type 1) and in pools with a slowly circulating current (type 3). These values, however, varied in the studied period, particularly for subsurface sediments in the sites with high sediment deposition rates. The content of organic matter (LOI) also exhibited highly variable values both for surface, 1.0–17.3%, and subsurface sediments, 1–13.1% (Figure 4). In general, contents of sediment and organic matter (LOI) showed positive correlations for subsurface sediment strata, whereas LOI values were extremely variable over the studied period in the surface sediment and for this reason they were not well correlated with fine sediment content (Table 4). Moreover, there were noticeable differences in the sediment content within the particular habitat types between both rivers. Namely, the mean sediment content in the glides of the Porębianka River was higher than in the Mszanka River (on the surface by 400% and by 250% in the subsurface), in pools (in the subsurface by 45%), while it was lower in runs (on the surface by 40% and in the subsurface by 60%) and in riffles (in the subsurface by 50%).
3.2. Benthic Fauna
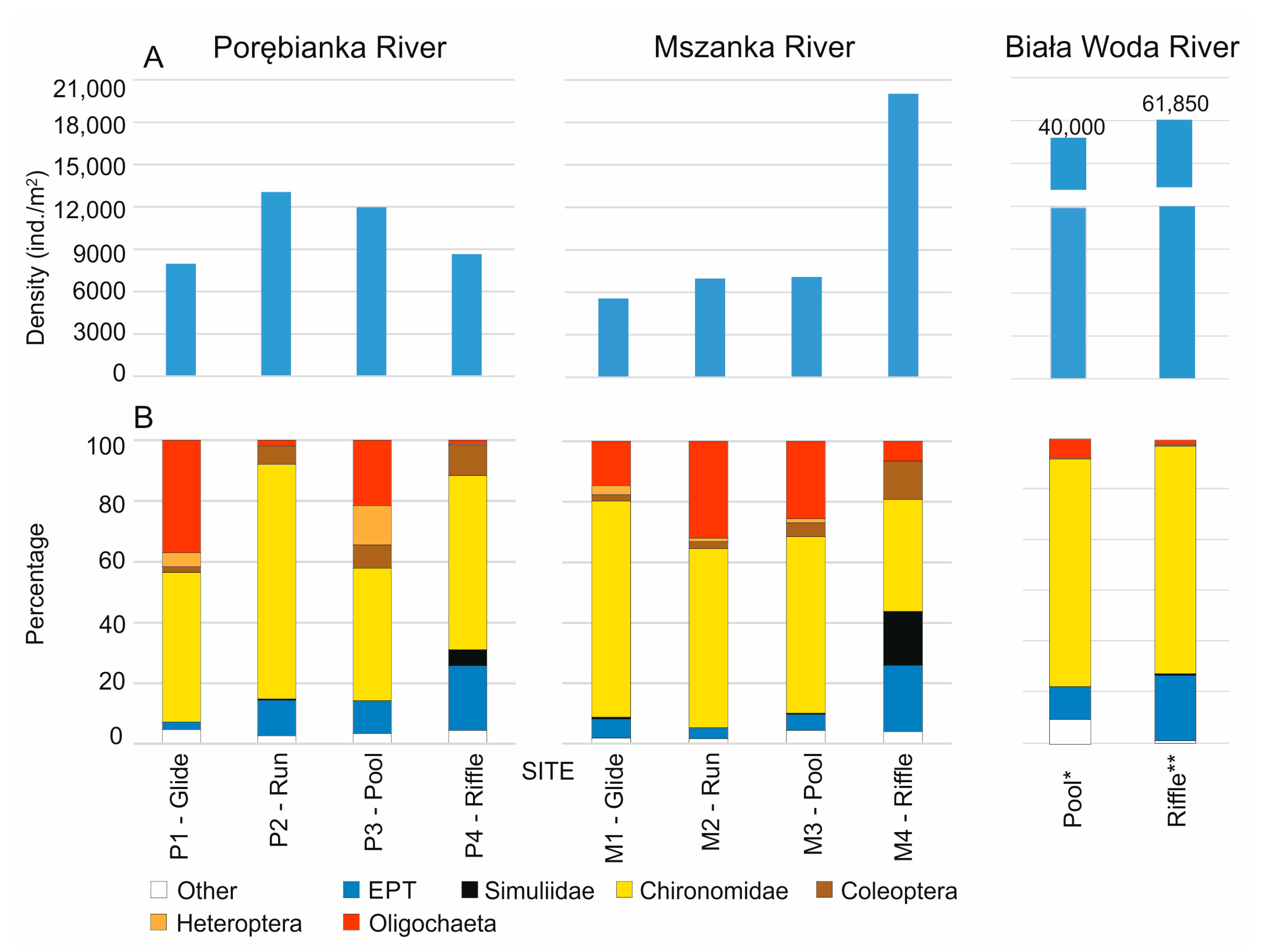
The median density of benthic macroinvertebrates was higher by 30% in the studied section of the Porębianka River than in the Mszanka River (Table 1). Considering the density of benthic macroinvertebrates in individual habitats, it was higher in glides, runs, and pools and lower in riffles of the Porębianka River in comparison to the Mszanka River (Figure 5). The differentiation of macroinvertebrate density in all of the habitats of the Porębianka River was relatively small and ranged from 7992 ind./m2 in glides to 13,057 ind./m2 in runs. There were no clear differences in the density of benthic macroinvertebrates between riffles (8661 ind./m2) and the other habitats. Inversely, the density of benthic macroinvertebrate in the riffles of the Mszanka River (20,029 ind./m2) was significantly higher in comparison to those in the other habitats (5571–7087 ind./m2).
In all habitat types of the Porębianka and Mszanka rivers, Chironomidae larvae were the dominant group of benthic macroinvertebrates (36.8–77.2% of the total) (Figure 5). Oligochaeta were more numerous in glides and pools (14.7–37%) than in riffles (1.3% and 6.6%, respectively). Solely in runs (type 2), the differences in the percentage of Oligochaeta between studied rivers were manifold (2% and 32%). The percentage of Ephemeroptera, Plecoptera, and Trichoptera, characteristic for mountain rivers, did not usually exceed 10%. The percentages of Ephemeroptera and Trichoptera were the highest in riffles (type 4) on the Porębianka River (15.9% and 5.5%, respectively) and on the Mszanka River (9.5% and 12.5%, respectively). A relatively high percentage of Coleoptera in riffles of the Porębianka River (10.1%) and the Mszanka River (12.7%), as well as Heteroptera in pools of the Porębianka River (12.8%), was unusual for a mountain river. The remaining groups of Ceratopogonidae, Hydracarina, Mollusca, Turbelaria, Sialis, Hirudinea, and Gammaridae were only present sporadically in the studied sites.
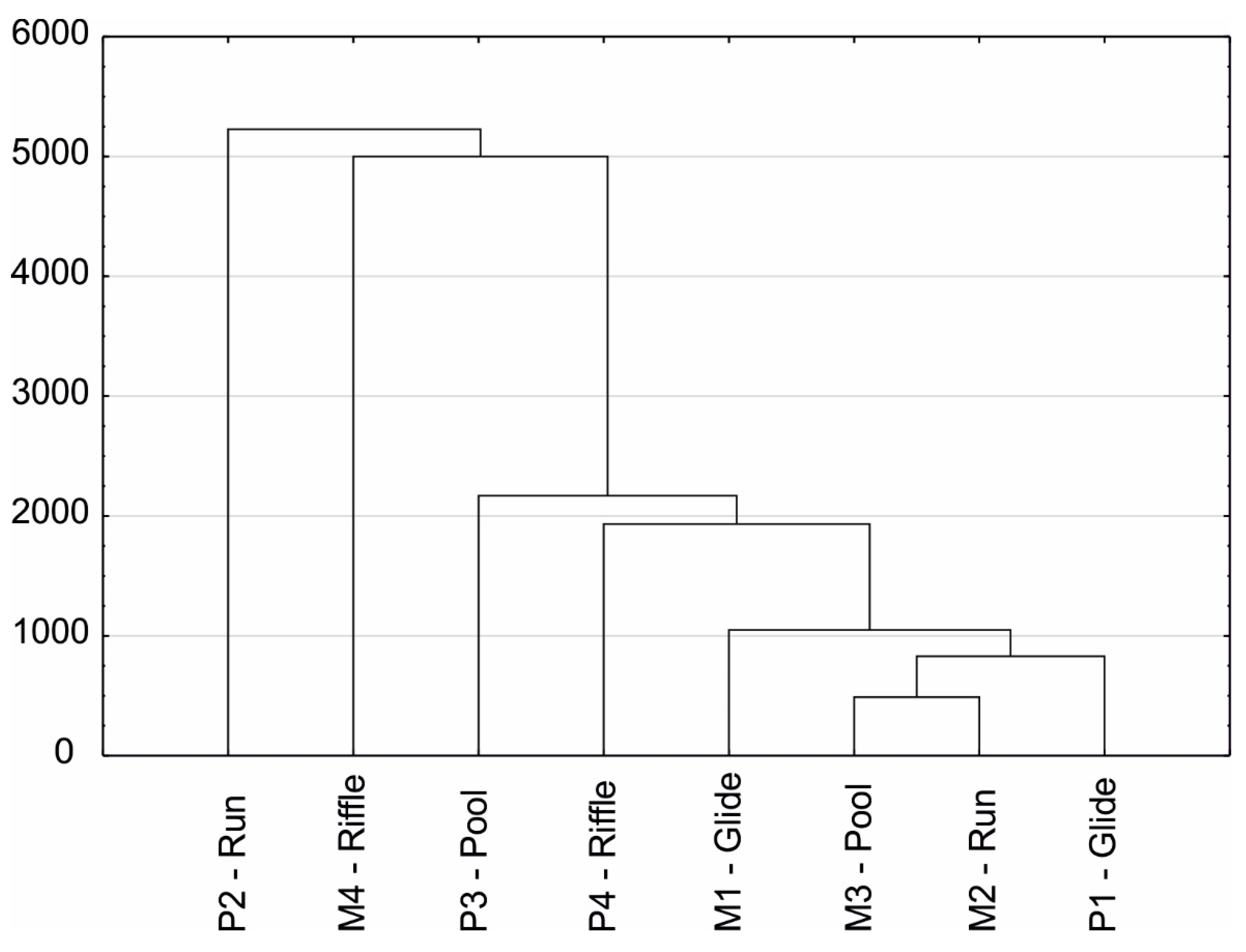
The dendrogram of similarity (Figure 6) exhibited the highest resemblance of the structure and density of benthic macroinvertebrate communities in glides, pools, and runs of the Mszanka River and glides of the Porębianka River. All of these habitats were characterized by the higher percentage of Oligochaeta (14.7–37.0%) and a low percentage (usually <5%) of Ephemeroptera and Trichoptera, which are characteristic of mountain rivers. The similarity of the other habitats was not as clear.
The density of EPT (Ephemeroptara, Plecoptera, Trichoptera) and Simuliidae exhibited a positive correlation with flow velocity, while Oligochaeta had a negative one (Table 4). Moreover, the density of EPT, Simuliidae, and Chironomidae showed a negative correlation, while Oligochaeta were positively correlated with the sediment content. The density of Simuliidae and Ephemeroptera was negatively correlated with the organic matter content in the sediment.
4. Discussion
4.1. Environmental Parameters
The waters of the Porębianka and Mszanka rivers were well oxygenated and had a slightly alkaline pH typical for other Carpathian streams and rivers in Poland [29,30]. The river waters, in principle, were clean. According to [31], the physicochemical class I was defined in terms of dissolved oxygen and nutrients NO3−, NH4+, and PO43− in both of the rivers, as was conductivity in the Porębianka River, whereas the Mszanka was class II in terms of conductivity. Slightly higher mean concentrations of NH4+ and PO43− in the Porębianka River might indicate the possible weak influence of domestic effluent. The samples were collected during low river discharges, when the share of the point source pollution load in total river flow is greater. It seems that the water chemistry was mainly controlled by natural processes, and much higher concentrations of Cl− and Na+ in the Mszanka than in the Porębianka were most probably also of natural origin. This difference might be related to numerous occurrences of natural chloride enrichment in springs or surface waters in this mountain area (up to 13.8 g/dm3) as a result of the local diagenetic dehydration of clay minerals or the leaching of marine water remnants from flysch rocks [32]. The observed differences in the water chemistry of both rivers were not important enough to have a negative impact and to varying degrees affect the benthic macroinvertebrate community.
There is a large variety of possible agents controlling the macroinvertebrate communities at both the landscape scale and in-channel microhabitats [16]. Channel morphology is a crucial factor controlling the diversity of river habitats. In the Porębianka and Mszanka rivers affected by channelization, small in-stream barriers became the main feature modifying channel gradient and flow velocities, albeit in a different way. The gradient of the traditionally engineered Mszanka River was reduced by 2.5 times from the average value of 0.018 to 0.007 in the studied river section. It resulted in the very small area of riffles (<5%), which occur immediately below the sill (Table 1). Pools were also associated with concrete sills where high flow velocities during floods induces the formation of scour holes [2] but they covered a very small area of the studied section (<3%). Most of the area, over two thirds in the middle part of the section (~66%), was associated with runs of moderate flow velocities whereas the areas of glides with very low flow velocities, favouring the highest deposition of fine sediments, extended over as much as 25%, mainly of the lowest parts of the section. Despite the eco-friendly design of the barriers on the Porębianka River, they reduced the natural channel gradient by seven times, and the areas of riffles and pools became even smaller than on the Mszanka River (Table 1). Due to the greater reduction of the channel gradient on the Porębianka River, there was the largest proportion of glides with the slowest currents (<0.15 m/s), exceeding half of the whole surface (~54%) in that river section. Riffles are recognized as local hot spots of macroinvertebrate density in mountain rivers due to high water turbulence and oxygenation [19]. However, as a result of the extensive channelization, the riffles became particularly unique in both studied rivers as their total length in the whole 140 m long reach of the Mszanka River is only in the order of a dozen meters. The reduction of habitats with fast flowing currents upstream of barriers and the expansion of the lentic sub-environment is widespread in impounded river sections [33].
Slow flows favour the deposition of fine sediments clogging spaces in the gravel bed. This process, known as siltation, impacts macroinvertebrate communities and is well known from other anthropogenically modified rivers [34,35]. In the studied river sections, due to their reduced gradient, the siltation of the stony substrate, strongly dependent on the longevity of low flow periods, also became widespread over most of the channel bed. In the Porębianka River, the highest siltation rate (expressed as fine sediment content) was confined to glides with the slowest flows occupying ~54% of the river bed. However, runs (43.3% of area) had considerable faster flows and a lower siltation rate as compared to those in the Mszanka River (66.3% of area) (Figure 2 and Figure 4). With respect to general flow velocity reduction, small in-stream barriers i.e., sills on the Mszanka and boulder ramps on the Porębianka, impacted fluvial ecosystems in a similar manner to dams, weirs or check dams. However, their upstream extent, depending on the height of the barrier, channel gradient, and purpose of the construction, was obviously much smaller [36].
The style in which the in-stream barriers were designed seems to be of minor importance, as all low in-stream barriers potentially only weakly affect benthic habitats [22]. Block ramps, which were designed in order to ensure better connectivity, do not seem to be particularly better than concrete sills as ramps do not provide connectivity even in the case of small fish [37].
The macroinvertebrate communities in rivers can also to some extent be associated with the different degree of the channel and riparian area modification. Its significance was indicated by many other studies documenting the naturalness of the channel and riparian zone. They stressed their role as a refuge for colonising species, being of the utmost importance for every river system and also affecting the effectiveness of many restoration projects [21,38]. The Porębianka and Mszanka rivers showed the different degree of the riparian area modification. The length of the channelized Porębianka river reach upstream of the studied section equals about 700 m and is almost three times shorter than on the Mszanka River. Moreover, the 3-km Porębianka River reach situated further upstream is fully natural, with bed rocks alternating with alluvial bars and also an up to 100 m wide riparian forest zone almost continuously overgrowing the channelized river section. It differs from the more extensively channelized 2-km-long upstream of the Mszanka River, with over 30 sills, with a height of 1 m or 30 cm, constructed over the course of 20 years and which are not accompanied by riparian forest but by occasionally removed willow bushes.
4.2. Benthic Macroinvertebrates
To assess the influence of channelization and flow velocity modifications on the benthic macroinvertebrate structure of both of the studied rivers, we referred our results to the studies of the natural Biała Woda River carried out in the 1970s [28]. This river catchment is representative for Carpathian middle mountain relief and of a very similar hydrology [39] and water chemistry [40]. The percentage of Chironomidae larvae in the investigated habitats of the Biała Woda River was very high (66–74%) and in that respect it resembled their content in the studied rivers (Figure 5). Conversely, Oligochaeta larvae, which occurred sporadically in the Biała Woda pools (without or with slow current; 5.8%) and riffles (1.7%) [28], were numerous (14.7–37%) in the glides, pools, and runs of the Mszanka and in the glides and pools of the Porębianka, whereas their content was only low in riffles. Ephemeroptera, which is an important group of benthic macroinvertebrates in habitats with high flow velocities, were more abundant in the Biała Woda in riffles (15%) [28] than in the studied rivers. In the Porębianka and Mszanka rivers, the similar percentage of Ephemeroptera was only found in riffles with turbulent flows whereas their percentage in runs, glides, and pools was lower (0.9–4.8%). Finally, the Coleoptera larvae occurred numerously in all habitats of the Porębianka and Mszanka rivers in comparison to the riffles and pools of the Biała Woda River, where only single specimens of that group were found [28]. Overall, the density of the benthic macroinvertebrates in the Biała Woda River was 4–6 times higher than in the Porębianka and Mszanka rivers. The comparison of the studied, human altered rivers with the natural Biała Woda River presented above indicates, that the modification of the benthic communities corresponds to the general shift of the investigated fluvial systems toward lentic habitat conditions. In particular, this change is reflected in the increase in the density of Oligochaeta in the studied rivers and a decrease in the share of rheophilic forms, such as Ephemeroptera and Trichoptera. This change can be partially related to the observed siltation of stones, which can favour the occurrence of Oligochaeta [41,42]. In contrast, Ephemeroptera, Plecoptera, and Trichoptera (EPT) are better adapted to habitats with rapid flow velocities [43], so they predominated in the riffles particularly of the Biała Woda River [28]. The dependence of EPT, Simuliidae, and Oligochaeta densities on flow and sediment content was also confirmed by the obtained correlations. EPT and Simuliidae densities were positively correlated with flow currents and negatively correlated with sediment content. Conversely, Oligochaeta densities were negatively correlated with flow and positively correlated with the sediment content.
The obtained result indicates that the Porębianka River represents a more natural habitat for benthic macroinvertebrates as compared to the Mszanka River. Their comparison exhibited a higher variability of the macroinvertebrate communities in the Porębianka than in the Mszanka river. This is evidenced by the dendrogram of the similarities where the studied sites of Porębianka (P1–4) did not form a separate group. Moreover, the higher median density of macroinvertebrates (by 30%) in the studied section of the Porębianka than the Mszanka indicates its more natural-like character and fits better to the other communities found in Carpathian rivers unaffected by extensive channelization [28]. The larger bias of the Mszanka than Porębianka river benthic communities, as compared to that of the natural river, was also evidenced by the generally lower and similar density of benthic macroinvertebrates in glides, runs, and pools (Table 1). This can be partially associated with generally slower flows and a higher siltation rate (express as sediment content) over longer low flow periods which was observed in the runs of the Mszanka River (Figure 2 and Figure 4).
To sum up, the obtained results exhibited the negative influence of channelization with in-stream barriers on benthic communities in both rivers. This can be mainly attributed to the decrease of channel gradients and the domination of relatively slow currents and shallow areas. Channelization, which evidently reduced riffle habitats in both studied rivers in comparison to the natural one, might affect macroinvertebrate density in other river reaches engineered with grade control structures. It seems that the higher density of the benthic macroinvertebrates in the Porębianka than the Mszanka river can also be associated to some extent with the differing degree of the modification of riparian areas.
5. Conclusions
The water chemistry of the studied rivers was typical for the northern Carpathians. The waters were well oxygenated, of the calcium-carbonate type and had a low content of nutrients. Therefore, the chemical composition of the waters of both rivers could not have adversely affected the communities of benthic macroinvertebrates.
The obtained results indicate that both types of the channelization of the Porębianka and Mszanka rivers induced modifications of the structure and density of the macroinvertebrate communities. Channelization of the Mszanka river extended over 2 km upstream and was associated with the periodic removal of riparian coppices. Due to the reduction in channel gradient by the concrete sills and the general slowing of flow currents, almost the whole surface of the investigated channel section became uniform, with runs of moderate depths and velocities occupying the prevailing part of its surface. The siltation of the stony substrate became widespread over most of the surface of the channel bed. The construction of a cascade of sills significantly reduced the surface of pools and riffles. In the Porębianka River, the cascade of block ramps reduced the channel gradient more than on the Mszanka River, but the highest siltation rate was confined to the glides with the slowest flows occupying half of the riverbed surface. The river section channelized with block ramps was accompanied by riparian forest and a natural river reach situated several hundred metres upstream.
The channelization of both rivers altered the macroinvertebrate communities. The river habitats were dominated by communities characteristic for slowly flowing rivers and pools, while communities characteristic for riffles and dominating in unaffected mountain rivers were only found in small areas. The macroinvertebrate community in individual habitats of the Porębianka River exhibited greater diversity and their median density in the studied river section was significantly higher (~30%) than in the Mszanka River. Therefore, overall, the Porębianka River represents more natural habitat conditions.
The results provide insights into the effects of the channelization of mountain river channels. Thus, they are important for making decisions on the selection of cost-effective techniques for the channelization of Carpathian rivers, especially in the case of the presence of not entirely justified interference with channel ecosystems.
Author Contributions
Conceptualization, E.S.-G., D.C. and A.K.; formal analysis, E.S.-G., D.C. and A.K.; investigation, E.S.-G., D.C. and A.K.; writing—original draft, E.S.-G., D.C. and A.K.; writing—review & editing, E.S.-G., D.C. and A.K.; visualization, E.S.-G., D.C. and A.K.; supervision, E.S.-G., D.C. and A.K.; project administration, E.S.-G. and D.C.; funding acquisition, E.S.-G. and D.C. All authors have read and agreed to the published version of the manuscript.
Funding
The research was funded by the AGH University of Science and Technology project no. 16.16.140.315 (D.C.) and by subvention of Institute of Nature Conservation, Polish Academy of Sciences (E.S.-G).
Data Availability Statement
Data requests can be addressed to the corresponding author.
Acknowledgments
We would like to thank Maria Profus for preparing the samples of benthic fauna.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Brookes, A. Channelized Rivers: Perspectives for Environmental Management; Wiley: Chichester, UK, 1988; p. 326. [Google Scholar]
- Korpak, J.; Lenar-Matyas, A.; Radecki-Pawlik, A.; Plesiński, K. Erosion irregularities from series of grade control structures: The Mszanka River, Western Carpathians. Sci. Total Environ. 2021, 799, 149469. [Google Scholar] [CrossRef] [PubMed]
- Casserly, C.M.; Turner, J.N.; O’Sullivan, J.J.; Bruen, M.; Bullock, C.; Atkinson, S.; Kelly-Quinn, M. Effect of low-head dams on reach-scale suspended sediment dynamics in coarse-bedded streams. J. Environ. Manag. 2021, 277, 111452. [Google Scholar] [CrossRef]
- Conesa-García, C.C.; Lenzi, M.A. (Eds.) Check Dams, Morphological Adjustments and Erosion Control in Torrential Streams; Nova Science Publishers. Inc.: New York, NY, USA, 2013. [Google Scholar]
- Korpak, J. The influence of river training on mountain channel changes (Polish Carpathian Mountains). Geomorphology 2007, 92, 166–181. [Google Scholar] [CrossRef]
- Rohasliney, H.; Jackson, D.C. Lignite and stream channelization influences on aquatic macroinvertebrate assemblages along the Natchez Trave Parkway, Mississippi, USA. Hydrobiologia 2008, 598, 149–162. [Google Scholar] [CrossRef]
- Tenchini, R.; Cazzola, M.; Erba, S.; Ballestrini, R.; Belgiore, C.; Pagnotta, R. Detecting the impact of bank and channel modification on invertebrate communities in Mediterranean temporary streams (Sardinia, SW Italy). Sci. Total Environ. 2016, 565, 1138–1150. [Google Scholar]
- Kownacki, A.; Szarek-Gwiazda, E.; Ligaszewski, M.; Urban, J. Communities of freshwater macroinvertebrate and fish in mountain streams and rivers of the Upper Dunajec Catchment (Western Carpathians) including long-term human impact. In Polish River Basins and Lakes: Biological Status and Water Management p. II.; Handbook of Environmental Chemistry Series; Korzeniewska, E., Hamisz, M., Eds.; Springer Nature Switzerland: Cham, Switzerland, 2020; Volume 87, pp. 269–294. [Google Scholar]
- Mykra, H.; Heino, J. Decreased habitat specialization in macroinvertebrate assemblages in anthropogenically disturbed streams. Ecol. Complex. 2017, 31, 181–188. [Google Scholar] [CrossRef]
- Buffagni, A.; Armanini, D.G.; Stefania, E.R.B.A. Does the lentic-lotic character of rivers affect invertebrate metrics used in the assessment of ecological quality? J. Limnol. 2009, 68, 92. [Google Scholar] [CrossRef]
- Jansen, W.; Böhmer, J.; Kappus, B.; Beiter, T.; Breitinger, B.; Hock, C. Benthic invertebrate and fish communities as indicators of morphological integrity in the Enz River (south-west Germany). Hydrobiologia 2000, 422–423, 331–342. [Google Scholar] [CrossRef]
- Kennedy, T.L.; Turner, T.F. River channelization reduces nutrient flow and macroinvertebrate diversity at the aquatic terrestrial transition zone. Ecosphere 2011, 2, 35. [Google Scholar] [CrossRef]
- Horsak, M.; Bojkova, J.; Zahradkova, S.; Omesova, M.; Helesic, J. Impact of reservoirs and channelization on lowland river macroinvertebrates: A case study from Central Europe. Limnologica 2009, 39, 140–151. [Google Scholar] [CrossRef] [Green Version]
- Palmer, M.A.; Menninger, H.L.; Bernhardt, E. River restoration, habitat heterogeneity and biodiversity: A failure of theory or practice? Frehw. Biol. 2010, 55, 205–222. [Google Scholar] [CrossRef]
- Leps, M.; Sundermann, A.; Tonkin, J.D.; Lorenz, A.W.; Haase, P. Time is no healer: Increasing restoration age does not lead to improved benthic invertebrate communities in restored river reaches. Sci. Total Environ. 2016, 557–558, 722–732. [Google Scholar] [CrossRef] [PubMed]
- Verdonschot, R.C.M.; Kail, J.; McKie, B.G.; Verdonschot, P.F.M. The role of benthic microhabitats in determining the effects of hydromorfological river restoration on macroinvertebrates. Hydrobiologia 2016, 769, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Leps, M.; Tonkin, J.D.; Dahm, V.; Haase, P.; Sundermann, A. Disentangling environmental drivers of benthic invertebrate assemblages: The role of spatial scale and riverscape heterogeneity in a multiple stressor environment. Sci. Total Environ. 2015, 536, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Poulos, H.M.; Miller, K.E.; Heinemann, R.; Kraczkowski, M.L.; Whelchel, A.W.; Chernoff, B. Dam removal effects on benthic macroinvertebrate dynamics: A New England stream case study (Connecticut, USA). Sustainability 2019, 11, 2875. [Google Scholar] [CrossRef] [Green Version]
- Cook, D.R.; Sullivan, S.M.P. Associations between riffle development and aquatic biota following lowhead dam removal. Environ. Monit. Assess. 2018, 190, 339. [Google Scholar] [CrossRef] [Green Version]
- Mbaka, J.G.; Mwaniki, M.W. A global review of the downstream effects of small impoundments on stream habitat conditions and macroinvertebrates. Environ. Rev. 2015, 23, 257–262. [Google Scholar] [CrossRef]
- Parkyn, M.S.; Smith, B.J. Dispersal constraintsfor stream invertebrates: Setting realistic timescales for biodiversity restoration. Environ. Manag. 2011, 48, 602–614. [Google Scholar] [CrossRef]
- Goeller, B.; Wolter, C. Performance of bottom ramps to mitigate gravel habitat bottlenecks in a channelized lowland river. Restor. Ecol. 2015, 23, 595–606. [Google Scholar] [CrossRef]
- Pagliara, S.; Radecki-Pawlik, A.; Palermo, M.; Plesiński, K. Block ramps in curved rivers: Morphology analysis and prototype data supported design criteria for mild bed slopes. River Res. Appl. 2017, 33, 427–437. [Google Scholar] [CrossRef]
- Korpak, J. Geomorphologic effects of river engineering structures in Carpathian fluvial systems. Landf. Anal. 2010, 14, 34–44. [Google Scholar]
- Plesiński, K.; Radecki-Pawlik, A.; Kuboń, P.; Tatara, T.; Pachla, F.; Jurowska, N. Bed load transport and alternation of a gravel-bed river morphology within a vicinity of block ramp: Classical and numerical approach. Sustainability 2022, 14, 4665. [Google Scholar] [CrossRef]
- Kalogianni, E.; Vourka, A.; Karaouzas, I.; Vardakas, L.; Laschou, S.; Skoulikidis, N.T. Combined effects of water stress and pollution on macroinvertebrate and fish assemblages in a Mediterranean intermittent river. Sci. Total Environ. 2017, 603, 639–650. [Google Scholar] [CrossRef] [PubMed]
- Duerdoth, C.P.; Arnold, A.; Murphy, J.F.; Naden, P.S.; Scarlett, P.; Collins, A.L.; Sear, D.A.; Jones, J.I. Assessment of a rapid method for quantitative reach-scale estimates of deposited fine sediment in rivers. Geomorphology 2015, 230, 37–50. [Google Scholar] [CrossRef]
- Kownacki, A. Stream ecosystems in mountain grassland (West Carpathians). 12. General conclusion. Acta Hydrobiol. 1982, 24, 413–422. [Google Scholar]
- Szarek-Gwiazda, E.; Mazurkiewicz-Boroń, G.; Gwiazda, R.; Urban, J. Chemical variability of water and sediment over time and along a mountain river subjected to natural and human impact. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 5. [Google Scholar] [CrossRef] [Green Version]
- Szarek-Gwiazda, E.; Gwiazda, R. Impact of flow and damming on water quality of the mountain Raba River (southern Poland)—Long-term studies. Arch. Environ. Prot. 2022, 48, 31–40. [Google Scholar]
- Regulation of the Minister of Maritime Economy and Inland Navigation of 11 October 2019 on the Classification of Ecological Status, Ecological Potential and Chemical Status as Well as the Method of Classifying the Status of Surface Water Bodies as Well as Environmental Quality Standards for Priority Substances. Dz. U. z dnia 7 November 2019 r., poz. 2149, Warszawa. Available online: https://isap.sejm.gov.pl/isap.nsf/DocDetails.xsp?id=WDU20190002148 (accessed on 20 January 2023). (In Polish)
- Zuber, A.; Chowaniec, J.; Borowiec, M. On the origin of chloride waters in the Polish Flysch Carpathians. Bull. State Geol. Surv. Pol. 2010, 441, 201–2008. [Google Scholar]
- Wang, J.; Ding, C.; Heino, J.; Jiang, X.; Tao, J.; Ding, L.; Su, W.; Huang, M.; He, D. What explains the variation in dam impacts on riverine macroinvertebrates? A global quantitative synthesis. Environ. Res. Lett. 2020, 15, 124028. [Google Scholar]
- Wood, P.J.; Armitage, P.D. Biological effects of fine sediment in the lotic environment. Environ. Manag. 1997, 21, 203–217. [Google Scholar] [CrossRef]
- Graf, W.; Leitner, P.; Hanetseder, I.; Ittner, L.D.; Dossi, F.; Hauer, C. Ecological degradation of meandering river by local channelization effects: A case study in an Austrian lowland river. Hydrobiologia 2016, 772, 145–160. [Google Scholar] [CrossRef]
- Poff, N.L.; Zimmerman, J.K.H. Ecological responses to altered flow regimes: A literature review to inform the science and management of environmental flows. Freshw. Biol. 2010, 55, 194–205. [Google Scholar] [CrossRef]
- Plesinski, K.; Bylak, A.; Radecki-Pawlik, A.; Mikolajczyk, T.; Kukula, K. Possibilities of fish passage trough the block ramp: Model-based estimation of permeability. Sci. Total Environ. 2018, 631–632, 1201–1211. [Google Scholar] [CrossRef] [PubMed]
- Lorenz, A.W. Continuous riverine biodiversity changes in a 10-years-post-restoration-study-impacts and pitfalls. River Res. Appl. 2021, 37, 270–282. [Google Scholar] [CrossRef]
- Kurek, S.; Pawlik-Dobrowolski, J. Stream ecosystems in mountain grassland (West Carpathians). 2. Hydrological characteristics. Acta Hydrobiol. 1982, 24, 375–390. [Google Scholar]
- Bombówna, M. Stream ecosystems in mountain grassland (West Carpathians). 3. Chemical composition of water. Acta Hydrobiol. 1982, 24, 321–335. [Google Scholar]
- Dumnicka, E. Habitat preferences of invertebrates (especially Oligochaeta) in a stream. Acta Hydrobiol. 1994, 30, 91–101. [Google Scholar]
- Syrovátka, V.; Schenková, J.; Brabec, K. The distribution of chironomid larvae and oligochaetes within a stony-bottomed river stretch: The role of substrate and hydraulic characteristics. Fundam. Appl. Limnol. 2009, 174, 43. [Google Scholar] [CrossRef]
- Longing, S.D.; Voshell, J.R.; Dolloff, C.A.; Roghair, C.N. Relationships of sedimentation and benthic macroinvertebrate assemblages in headwater streams using systematic longitudinal sampling at the reach scale. Environ. Monit. Assess. 2010, 161, 517–530. [Google Scholar] [CrossRef]
Figure 1.
Location of the study site.

Figure 2.
Characteristic features of the investigated sections of the Porębianka and Mszanka rivers; arrows indicate flow direction.
Figure 2.
Characteristic features of the investigated sections of the Porębianka and Mszanka rivers; arrows indicate flow direction.

Figure 3.
Flow depths and velocities in the studied Mszanka and Porębianka river reaches.

Figure 4.
The values of environmental parameters in the Porębianka and Mszanka rivers.

Figure 5.
The mean density (A) and mean percentage (B) of benthic macroinvertebrate groups in the Porębianka and Mszanka rivers (present study) and in the Biała Woda River (according to [28]). In [28] the description of pool and riffle had a wider meaning: * pool—without or with slow current ** riffle—fast current.
Figure 5.
The mean density (A) and mean percentage (B) of benthic macroinvertebrate groups in the Porębianka and Mszanka rivers (present study) and in the Biała Woda River (according to [28]). In [28] the description of pool and riffle had a wider meaning: * pool—without or with slow current ** riffle—fast current.

Figure 6.
Hierarchical cluster analysis based on the density of macroinvertebrate groups in the Porębianka and Mszanka rivers.
Figure 6.
Hierarchical cluster analysis based on the density of macroinvertebrate groups in the Porębianka and Mszanka rivers.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Sampled stream channel habitats.
| Habitat Area in the Rivers [%] | Macroinvertebrate Density [ind./m2] | |||
|---|---|---|---|---|
| Habitat Types | Porębianka | Mszanka | Porębianka | Mszanka |
| 1. Glides (relatively shallow areas with slow current < 0.15 m/s) | 53.7 | 26.3 | 7972 | 5571 |
| 2. Runs (areas of relatively moderate depth and current 0.15–0.6 m/s) | 43.3 | 66.3 | 13,058 | 6981 |
| 3. Pools (relatively deep areas, >0.6 m with slow current < 0.15 m/s) | 1.7 | 2.7 | 11,967 | 7088 |
| 4. Riffles (relatively shallow areas with rapid current > 0.6 m/s) | 1.4 | 4.7 | 8662 | 20,029 |
| Median density in the studied stream section | 10,314 | 7034 | ||
Table 2.
The values of physicochemical parameters (mean, standard deviation) in the waters of the Porębianka and Mszanka rivers.
Table 2.
The values of physicochemical parameters (mean, standard deviation) in the waters of the Porębianka and Mszanka rivers.
| Parameter | Unit | Porębianka | Mszanka |
|---|---|---|---|
| Dissolved oxygen | mg/L | 11.3 ± 1.1 | 11.3 ± 1.1 |
| Oxygen saturation | % | 105.6 ± 10.0 | 108.2 ± 15.2 |
| Conductivity | µS/cm | 281.8 ± 65.5 | 323.0 ± 66.2 |
| pH | 8.0 ± 0.3 | 8.0 ± 0.2 | |
| Cl− | mg/L | 6.5 ± 3.4 | 11.9 ± 2.4 |
| SO42− | mg/L | 12.6 ± 2.2 | 14.8 ± 2.0 |
| HCO3− | mg/L | 118.5 ± 39.2 | 141.0 ± 30.1 |
| Na+ | mg/L | 4.9 ± 2.8 | 8.7 ± 1.4 |
| K+ | mg/L | 1.8 ± 2.0 | 3.0 ± 0.6 |
| Ca2+ | mg/L | 41.2 ± 9.4 | 45.1 ± 10.1 |
| Mg2+ | mg/L | 7.3 ± 1.9 | 8.5 ± 1.9 |
| NO3− | mg/L | 3.5 ± 1.7 | 2.9 ± 0.9 |
| NH4+ | mg/L | 0.028 ± 0.013 | 0.015 ± 0.018 |
| PO43− | mg/L | 0.008 ± 0.011 | 0.007 ± 0.010 |
Table 3.
Differences in the chemical composition of waters between the Porębianka and Mszanka rivers (Wilcoxon test). Only significant differences are given.
Table 3.
Differences in the chemical composition of waters between the Porębianka and Mszanka rivers (Wilcoxon test). Only significant differences are given.
| Parameter | N | T | Z | p |
|---|---|---|---|---|
| Conductivity | 5 | 0.00 | 2.02 | 0.043 |
| Cl− | 5 | 0.00 | 2.02 | 0.043 |
| SO42− | 5 | 0.00 | 2.02 | 0.043 |
| HCO3− | 5 | 0.00 | 2.02 | 0.043 |
| Na+ | 5 | 0.00 | 2.02 | 0.043 |
| K+ | 5 | 0.00 | 2.02 | 0.043 |
| Ca2+ | 5 | 0.00 | 2.02 | 0.043 |
| Mg2+ | 5 | 0.00 | 2.02 | 0.043 |
Table 4.
Spearman correlation coefficients between the values of environmental variables and density of benthic macroinvertebrates in the Porębianka and Mszanka rivers (in total). Significant at p < 0.05. Only significant correlations are given. Abbreviations: DO—dissolved oxygen, * LOI Surface—LOI in surface sediment, ** LOI SubSed—LOI in subsurface sediment.
Table 4.
Spearman correlation coefficients between the values of environmental variables and density of benthic macroinvertebrates in the Porębianka and Mszanka rivers (in total). Significant at p < 0.05. Only significant correlations are given. Abbreviations: DO—dissolved oxygen, * LOI Surface—LOI in surface sediment, ** LOI SubSed—LOI in subsurface sediment.
| Parameter/Fauna Group | Flow | Dissolved Oxygen | Surface Sediment | Subsurface Sediment | LOI Surface * | LOI SubSed ** |
|---|---|---|---|---|---|---|
| Surface sediment | −0.88 | - | 0.91 | 0.66 | ||
| Subsurface sediment | −0.81 | 0.91 | - | 0.54 | ||
| LOI Surface * | - | 0.68 | ||||
| LOI SubSed ** | −0.61 | 0.66 | 0.54 | 0.68 | - | |
| Oligochaeta | −0.42 | 0.41 | ||||
| Chironomidae | −0.47 | |||||
| Simullidae | 0.49 | −0.47 | −0.46 | −0.47 | −0.56 | |
| Ephemeroptera | 0.56 | −0.56 | −0.42 | −0.43 | ||
| Plecoptera | 0.46 | −0.47 | ||||
| Trichoptera | 0.57 | −0.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Szarek-Gwiazda, E.; Ciszewski, D.; Kownacki, A. The Effects of Channelization with Low In-Stream Barriers on Macroinvertebrate Communities of Mountain Rivers. Water 2023, 15, 1059. https://doi.org/10.3390/w15061059
AMA Style
Szarek-Gwiazda E, Ciszewski D, Kownacki A. The Effects of Channelization with Low In-Stream Barriers on Macroinvertebrate Communities of Mountain Rivers. Water. 2023; 15(6):1059. https://doi.org/10.3390/w15061059
Chicago/Turabian StyleSzarek-Gwiazda, Ewa, Dariusz Ciszewski, and Andrzej Kownacki. 2023. "The Effects of Channelization with Low In-Stream Barriers on Macroinvertebrate Communities of Mountain Rivers" Water 15, no. 6: 1059. https://doi.org/10.3390/w15061059
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.