
Confocal Laser Scanning Microscopy Applied to a New Species Helps Understand the Functioning of the Reproductive Apparatus in Stylet-Bearing Urodasys (Gastrotricha: Macrodasyida) †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Sample Processing
2.2. Microscopical Study
2.2.1. Differential Interference Contrast (DIC)
2.2.2. Confocal Laser Scanning Microscopy (CLSM)
2.3. General Conventions
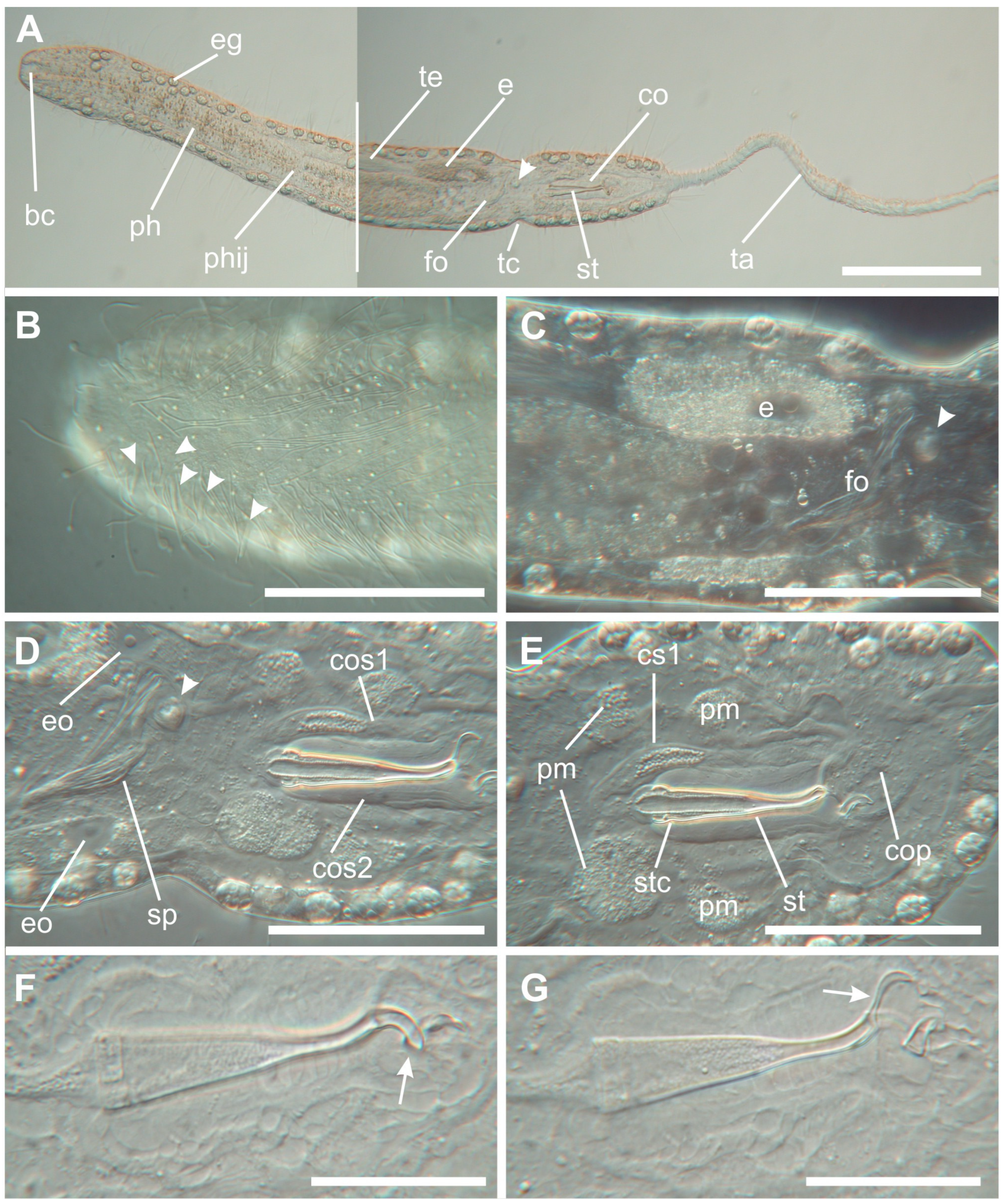
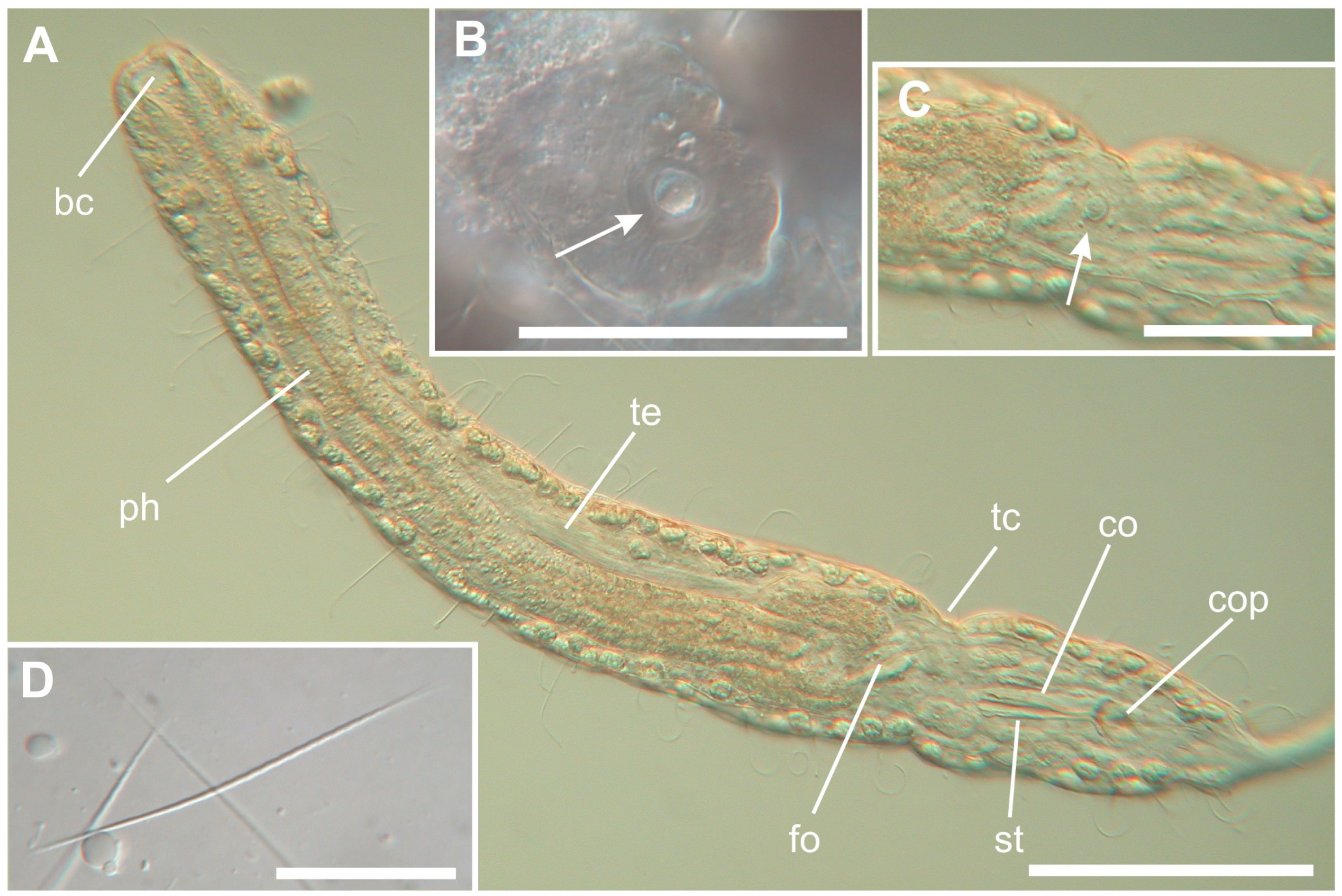
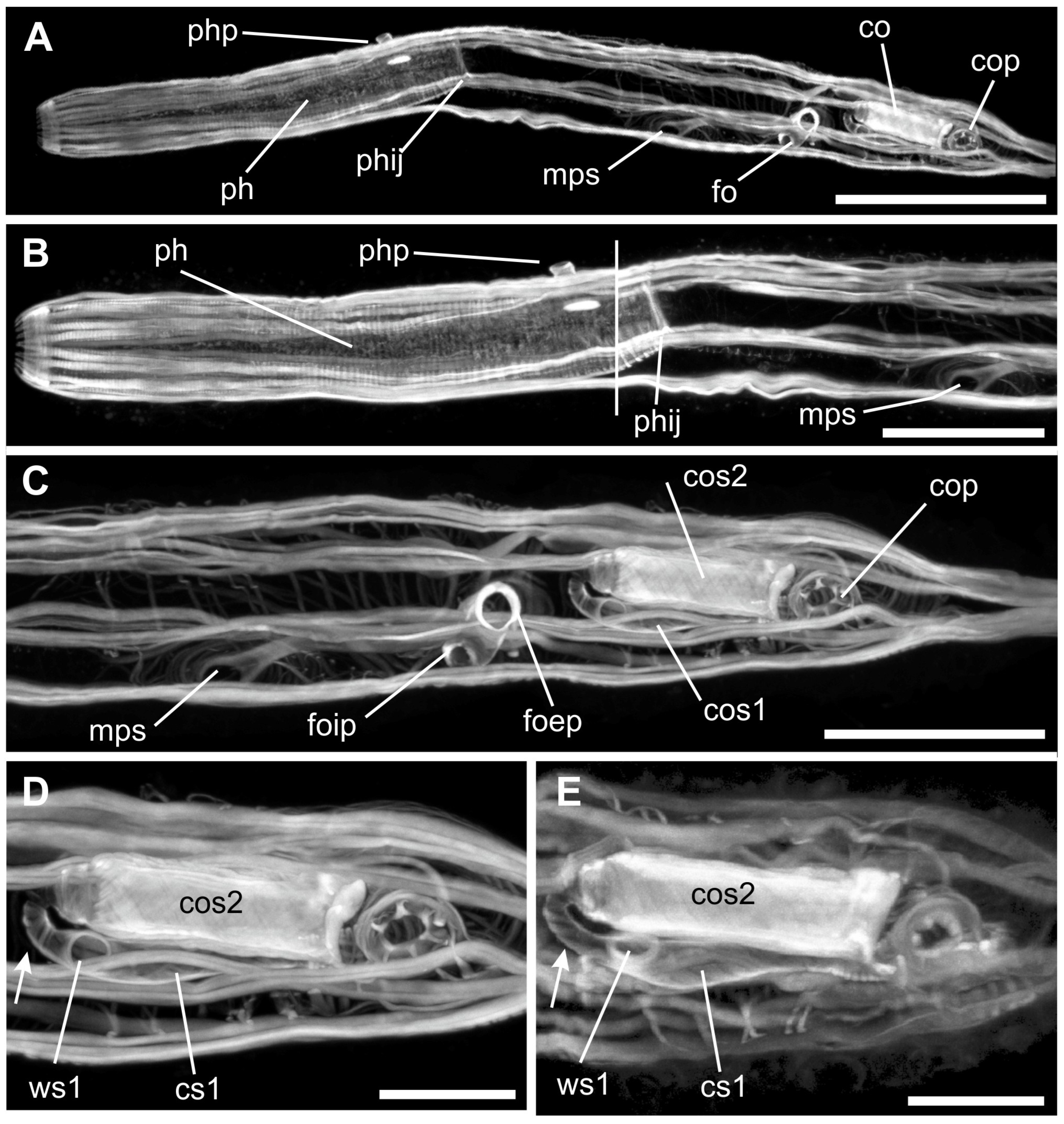
3. Results
Taxonomic Account
4. Discussion
4.1. Taxonomic Affinities
4.2. Reproductive System Functioning
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Todaro, M.A.; Sibaja-Cordero, J.A.; Segura-Bermúdez, O.A.; Coto-Delgado, G.; Goebel-Otárola, N.; Barquero, J.D.; Mariana Cullell-Delgado, M.; Dal Zotto, M. An introduction to the study of Gastrotricha, with a taxonomic key to families and genera of the group. Diversity 2019, 11, 117. [Google Scholar] [CrossRef] [Green Version]
- Kieneke, A.; Todaro, M.A. Discovery of two ‘chimeric’ Gastrotricha and their systematic placement based on an integrative approach. Zool. J. Linn. Soc. 2021, 192, 710–735. [Google Scholar] [CrossRef]
- Balsamo, M.; Artois, T.; Smith, J.P.S.; Todaro, M.A.; Guidi, L.; Leander, B.S.; Van Steenkiste, N.W.L. The curious and neglected soft-bodied meiofauna: Rouphozoa (Gastrotricha and Platyhelminthes). Hydrobiologia 2020, 847, 2613–2644. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.A.; Luporini, P. Not too big for its mouth: Direct evidence of a macrodasyidan gastrotrich preyed in nature by a dileptid ciliate. Eur. Zool. J. 2022, 89, 785–790. [Google Scholar] [CrossRef]
- Dayrat, B. Towards integrative taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Paddock, S.W. (Ed.) Confocal Microscopy: Methods and Protocols; Humana Press: Totowa, NJ, USA, 1999; pp. 1–446. [Google Scholar]
- Leasi, F.; Neves, R.C.; Worsaae, K.; Sørensen, M.V. Musculature of Seison nebaliae Grube, 1861 and Paraseison annulatus (Claus, 1876) revealed with CLSM: A comparative study of the gnathiferan key taxon. Zoomorphology 2012, 131, 185–195. [Google Scholar] [CrossRef]
- Neves, R.C.; Bailly, X.; Leasi, F.; Reichert, H.; Sørensen, M.V.; Kristensen, R.M. A complete three-dimensional reconstruction of the myoanatomy of Loricifera: Comparative morphology of an adult and a Higgins larva stage. Front. Zool. 2013, 10, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Altenburger, A. The neuromuscular system of Pycnophyes kielensis (Kinorhyncha: Allomalorhagida) investigated by confocal laser scanning microscopy. EvoDevo 2016, 7, 25. [Google Scholar] [CrossRef] [Green Version]
- Leasi, F.; Rothe, B.H.; Schmidt-Rhaesa, A.; Todaro, M.A. The musculature of three species of gastrotrichs surveyed with confocal laser scanning microscopy (CLSM). Acta Zool. 2006, 87, 171–180. [Google Scholar] [CrossRef]
- Leasi, F.; Todaro, M.A. The muscular system of Musellifer delamarei (Renaud-Mornant, 1968) and other chaetonotidans with implications for the phylogeny and systematization of the Paucitubulatina (Gastrotricha). Biol. J. Linn. Soc. 2008, 94, 379–398. [Google Scholar] [CrossRef]
- Leasi, F.; Todaro, M.A. Meiofaunal cryptic species revealed by confocal microscopy: The case of Xenotrichula intermedia (Gastrotricha). Mar. Biol. 2009, 156, 1335–1346. [Google Scholar] [CrossRef]
- Todaro, M.A.; Dal Zotto, M.; Leasi, F. An integrated morphological and molecular approach to the description and systematisation of a novel genus and species of Macrodasyida (Gastrotricha). PLoS ONE 2015, 10, e0130278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochberg, R. Musculature of the primitive gastrotrich Neodasys (Chaetonotida): Functional adaptations to the interstitial environment and phylogenetic significance. Mar. Biol. 2005, 146, 315–323. [Google Scholar] [CrossRef]
- Kieneke, A.; Arbizu, P.M.; Riemann, O. Body musculature of Stylochaeta scirtetica Brunson, 1950 and Dasydytes (Setodytes) tongiorgii (Balsamo, 1982) (Gastrotricha: Dasydytidae): A functional approach. Zool. Anz. 2008, 247, 147–158. [Google Scholar] [CrossRef]
- Atherton, S.; Hochberg, R. The evolution of the reproductive system of Urodasys (Gastrotricha: Macrodasyida). Invert. Biol. 2014, 133, 314–323. [Google Scholar] [CrossRef]
- Todaro, M.A.; Cesaretti, A.; Dal Zotto, M. Marine gastrotrichs from Lanzarote, with a description of a phylogenetically relevant species of Urodasys (Gastrotricha, Macrodasyida). Mar. Biodivers. 2019, 49, 2109–2123. [Google Scholar] [CrossRef]
- Schoepfer-Sterrer, C. Five new species of Urodasys and remarks on the terminology of the genital organs in Macrodasyidae (Gastrotricha). Cah. Biol. Mar. 1974, 15, 229–254. [Google Scholar]
- Fregni, E.; Tongiorgi, P.; Faienza, M.G. Two new species of Urodasys (Gastrotricha, Macrodasyidae) with cuticular stylet. Ital. J. Zool. 1998, 65, 377–380. [Google Scholar] [CrossRef]
- Ruppert, E.E. Gastrotricha. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Ruppert, R.R., Eds.; Wiley-Liss: New York, NY, USA, 1991; Volume 4, pp. 41–109. [Google Scholar]
- Kieneke, A.; Schmidt-Rhaesa, A. Gastrotricha. In Handbook of Zoology; Schmidt-Rhaesa, A., Ed.; De Gruyter: Berlin, Germany, 2015; pp. 1–134. [Google Scholar]
- Dal Zotto, M.; Tongiorgi, P.; Todaro, M.A. I Gastrotrichi dell’area marina protetta di Capo caccio-Isola Piana (Sardegna). In Proceedings of the 68° Congress of the Unione Zoologica Italiana, Lecce, Italy, 24–27 September 2007. [Google Scholar]
- Curini-Galletti, M.; Artois, T.; Delogu, V.; De Smet, W.H.; Fontaneto, D.; Jondelius, U.; Leasi, F.; Martinez, A.; Meyer-Wachsmuth, I.; Nilsson, K.S.; et al. Patterns of diversity in soft-bodied Meiofauna: Dispersal ability and Body size matter. PLoS ONE 2012, 7, e33801. [Google Scholar] [CrossRef] [PubMed]
- Todaro, M.A.; Dal Zotto, M.; Kånneby, T.; Hochberg, R. Integrated data analysis allows the establishment of a new, cosmopolitan genus of marine Macrodasyida (Gastrotricha). Sci. Rep. 2019, 9, 7989. [Google Scholar] [CrossRef] [Green Version]
- Remane, A. Neue aberrante Gastrotrichen II: Turbanella cornuta nov.spec. und T. hyalina M. Schultze, 1853. Zool. Anz. 1925, 64, 309–314. [Google Scholar]
- Rao, G.C.; Clausen, C. Planodasys marginalis gen. et sp. nov. and Planodasyidae fam. nov. (Gastrotricha Macrodasyoidea). Sarsia 1970, 42, 73–82. [Google Scholar] [CrossRef]
- Remane, A. Neue aberrante Gastrotrichen. I. Macrodasys buddenbrocki nov. gen. nov. spec. Zool. Anz. 1924, 61, 289–297. [Google Scholar]
- Remane, A. Morphologie und Verwandtschaftbeziehungen der aberranten Gastrotrichen I. Z. Morph. Oekol. Tiere 1926, 5, 625–754. [Google Scholar]
- ICZN. International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- ICZN Declaration 45—Addition of Recommendations to Article 73 and of the term “specimen, preserved” to the Glossary. Bull. Zool. Nomencl. 2017, 73, 2–4.
- Fregni, E.; Faienza, M.G.; Grimaldi-De Zio, S.; Tongiorgi, P.; Balsamo, M. Marine gastrotrichs from the Tremiti Archipelago in the southern Adriatic Sea, with the description of two new species of Urodasys. It. J. Zool. 1999, 66, 183–194. [Google Scholar] [CrossRef] [Green Version]
- Atherton, S. Urodasys poculostylis sp. nov., a new stylet-bearing gastrotrich (Macrodasyida) from Capron Shoal, Florida. Mar. Biol. Res. 2014, 5, 530–536. [Google Scholar] [CrossRef]
- Hummon, W.D. Marine Gastrotricha of the Near East: 1. Fourteen new species of Macrodasyida and a redescription of Dactylopodola agadasys Hochberg, 2003. Zookeys 2011, 94, 1–59. [Google Scholar] [CrossRef] [PubMed]
- Guidi, L.; Balsamo, M.; Grassi, L.; Semprucci, F.; Todaro, M.A. New data on reproductive system and spermatozoa confirm Macrodasys as a model in comparative reproductive analysis in Macrodasyida (Gastrotricha). Water 2022, 14, 3085. [Google Scholar] [CrossRef]
- Ruppert, E.E. The reproductive system of gastrotrichs. II. Insemination in Macrodasys: A unique mode of sperm transfer in Metazoa. Zoomorphologie 1978, 89, 201–228. [Google Scholar]
- Mann, T. Spermatophores: Development, Structure, Biochemical Attributes and Role in the Transfer of Spermatozoa; Springer: Heidelberg, Germany, 1984; pp. 1–217. [Google Scholar]
- Kieneke, A.; Arbizu, P.M.; Ahlrichs, W.H. Anatomy and ultrastructure of the reproductive organs in Dactylopodola typhle (Gastrotricha: Macrodasyida) and their possible functions in sperm transfer. Invertebr. Biol. 2008, 127, 12–32. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cesaretti, A.; Leasi, F.; Todaro, M.A. Confocal Laser Scanning Microscopy Applied to a New Species Helps Understand the Functioning of the Reproductive Apparatus in Stylet-Bearing Urodasys (Gastrotricha: Macrodasyida). Water 2023, 15, 1106. https://doi.org/10.3390/w15061106
Cesaretti A, Leasi F, Todaro MA. Confocal Laser Scanning Microscopy Applied to a New Species Helps Understand the Functioning of the Reproductive Apparatus in Stylet-Bearing Urodasys (Gastrotricha: Macrodasyida). Water. 2023; 15(6):1106. https://doi.org/10.3390/w15061106
Chicago/Turabian StyleCesaretti, Agata, Francesca Leasi, and M. Antonio Todaro. 2023. "Confocal Laser Scanning Microscopy Applied to a New Species Helps Understand the Functioning of the Reproductive Apparatus in Stylet-Bearing Urodasys (Gastrotricha: Macrodasyida)" Water 15, no. 6: 1106. https://doi.org/10.3390/w15061106