
Role of Hydrology in Cyanobacterial Blooms in the Floodplain Lakes

1
Department of Hydrobiology and Protection of Ecosystems, University of Life Sciences in Lublin, 13 Akademicka St., 20-950 Lublin, Poland
2
Department of Hydrology and Climatology, Maria Curie-Skłodowska University, Kraśnicka 2cd, 20-718 Lublin, Poland
*
Author to whom correspondence should be addressed.
Water 2023, 15(8), 1547; https://doi.org/10.3390/w15081547
Submission received: 21 February 2023
/
Revised: 11 April 2023
/
Accepted: 13 April 2023
/
Published: 14 April 2023
(This article belongs to the Section Water Quality and Contamination)
Abstract
:River floodplains are sites of extremely high environmental and ecological value due to high biodiversity. Floodplain lakes (FPLs) are essential parts of complex floodplain ecosystems’ biological and hydro-chemical processes. Clear seasonal cycles, determined by the regime of the parent river and closely linked to climatic conditions, are observed in these lakes. Both the quantity and the quality of water are determined by the functional phases of a floodplain lake; limnophase, when the lake is isolated from the river, and potamophase (inundation), when the floodplain lake is overflowed by the river waters. These phases highly modify lake bathymetry, water balance, chemistry, and biology. Human pressure, mostly dam construction and land use changes, alter the frequency, duration, and intensity of natural cycles of inundation. The majority of large rivers in the Northern Hemisphere have been impacted by human activity. Floodplain lakes are also affected by continuous climate change, due to the alterations of the hydrological regimes. Cyanobacterial blooms in floodplain lakes are often characterized by high qualitative and quantitative annual and long-term variability. The main forces driving cyanobacterial blooms can be found in the flooding dynamics and intensity influencing hydraulic residence time and the concentration of biogenic compounds in lake water. Cyanotoxin production and seasonal dynamics in floodplain lakes have been rarely studied, particularly in connection with the hydrological regime. Moreover, the effects of cyanobacterial blooms and their toxicity to organisms inhabiting floodplain lakes, connected rivers, and floodplains are poorly understood. Therefore, knowledge of the processes controlling floodplain lakes’ ecological conditions is crucial to implementing water management and restoration practices, protecting those fragile and precious ecosystems from degradation and possible extension.
1. Introduction
Floodplains, especially those of natural or quasi-natural rivers, are exceptionally valuable areas, from both environmental and economic points of view, due to their extremely high biodiversity potential [1,2]. Floodplain terraces may be a sink of nutrients and sediments, thus they provide diversified ecological services [3,4]. Floodplain lakes [FPLs] are a basic part of the multidirectional floodplain ecosystems’ biological and hydro-chemical processes [5,6]. However, small FPLs are one of the least studied components of river floodplains [7].
Junk et al. [8] were the first to point out that the dynamic of river water levels (so-called flood pulses) determines the functioning of floodplain ecosystems [9]. This is commonly known as the flood pulse concept. The seasonal fluctuations of hydrological factors (water levels and discharge), affecting pulses of matter and energy, are typical for all natural floodplains [10,11]. They also result in dynamic changes in the bathymetry of a floodplain lake [12,13]. River flood regimes are mostly shaped by the climatic conditions of the drainage system [14]. The significance of the fluctuation of river flood periods in shaping both fauna and flora has been well documented in tropical floodplains [15]. Unfortunately, it is estimated that over 90% of rivers in Europe and North America have been altered due to human impact [16]. Human pressure on river valleys has modified the duration, frequency, and intensity of natural floods [17], disturbing the natural functional phases of the floodplain ecosystems (potamophases and limnophases).
Floodplain lakes (FPLs) may represent lentic or lotic environments, or both in different sections of the lake basin. Both the quantity and quality of water are determined by the cyclically occurring functional phases of the floodplain lake. Generally, four annual or multiannual functional phases may be observed: periods of filling, inundation, drainage, and isolation [18,19]. However, the two-phase cycles (limnophase and potamophase; or inundation and isolation) are also commonly distinguished [20]. The repeatability of those periods is responsible for maintaining FPLs’ hydro-sedimentological regime [10]. During the potamophase period, due to the excess water input as fluvial supply, precipitation, and surface runoff, the bathymetric features of FPLs rapidly change, lake basins can be beyond 10 m deep at that time, whereas during the limnophase, the same lakes significantly lose water due to hydrological isolation from the parent river [21]. Flood periods not only remodel lake morphology (maximum depth, volume, surface area), but also water balance, chemistry, and biology. A significant similarity between the physical-chemical composition of river and floodplain waters is usually observed in the potamophase period. When river waters enter FPL, the lake’s drainage area equals the parent river catchment to the point of the overbank crevasse. During extreme floods (Figure 1), floodplain lakes are unidentifiable, being merely a cavity in the flooded terrace. On the other hand, during prolonged drought periods, floodplain lakes significantly shrink, and many of the small water bodies periodically dry out. The duration, amplitude, and frequency of occurrence of potamophases and limnophases shape the functioning and integrity of the environment of the adjacent floodplain area [22].
In addition to meteorological conditions, both the duration and frequency of functional periods are derivative of lateral connectivity. Thus, in recent years, numerous studies on the subject have been published [23,24,25,26,27]. Worldwide, there are floodplain lakes permanently connected with the main river, others are permanently isolated from the river, and the majority in which annual and/or multi-annual cycles of potamophases and limnophases occur [28]. The connectivity pattern is a primary factor determining floodplain lakes’ biodiversity and water quality [29]. Both matter and energy transferred by the rivers’ water [30] affect the biomass and abundance of plants and animals in the aquatic system of the FPL [31]. Increases in the amount of nitrogen and phosphorus concentrations in freshwater bodies, as well as the increased concentration of organic matter, accelerate eutrophication. Furthermore, the eutrophication process may cause excessive cyanobacterial and algal growth (so-called water blooms), altering color and light penetration in the water column, depleting dissolved oxygen (DO), and even producing toxins and other biologically active algal and, mostly, cyanobacterial compounds [32]. Cyanobacterial blooms (CyanoHABs) alter the relative importance of neutral and selective processes in assembling freshwater biocenoses such as, for instance, the bacterioplankton community [33]. The connection of the floodplain lake and the parent river may be influenced by both natural and anthropogenic factors, among which climate, hydrology, industry, and agriculture, play key roles [34,35]. However, estimations of the impact of these factors on water quality are very difficult, particularly in floodplain lakes, due to a significant seasonal variation in hydrological conditions [36,37]. These hydrologic conditions, representing either the presence or absence of flow-paths between aquatic ecosystems among floodplains, have been defined as hydrologic connectivity [38]. Four types of connectivity may be distinguished: longitudinal (channel-channel), lateral (channel-floodplain), vertical (channel-groundwater), and temporal (in time) [39].
The aim of this paper was to gather, on the basis of the available literature data, the present knowledge regarding the occurrence of cyanobacterial blooms and cyanotoxin production in floodplain lakes. The selection conditions of the lakes were the classification of lakes as floodplain lakes and the availability of data on the abundance and/or biomass of cyanobacteria, chlorophyll-a concentration (as an equivalent of phytoplankton biomass), the share, as well as the occurrence and seasonal dynamics, of particular cyanobacterial bloom-forming species. We also analyzed the available literature data pertaining to toxin production (including seasonal cyanotoxin dynamics) in analyzed lakes and compared them with lake hydrology, where possible.
2. Hydrological Connectivity
River valleys represent both terrestrial and aquatic environments, connected and interdependent due to the fluvial impulse. The exchange of matter (both mineral and organic), energy, and/or wildlife between a river and its floodplain is called ‘hydrological connectivity’. Fours dimensions of the connectivity may be distinguished: longitudinal, lateral, vertical, and temporal [40]. FPLs may be hydrologically isolated from the river, or permanently or temporarily connected to the parent river [41]. Flooding periods are extremely environmentally important, maintaining the ecological integrity of the river valley ecosystems [42]. Obolewski et al. [43] believe that lakes’ morphometry is the main factor driving the process of matter exchange in the river valley. During the potamophase period, river waters reach the floodplain, homogenizing both the physical-chemical and biological conditions of water bodies [44,45]. Thus, it has been observed that the water of frequently inundated floodplain lakes shows similarity to the river water quality [46]. The role of connectivity in shaping biodiversity and maintaining rare species has also been widely recognized [47]. Higher connectivity (favoring a shorter water residence time) allows species to spread within the floodplain ecosystems [48]. Wang et al. [49] observed that flooding increased phytoplankton abundance and diversity. Similarly, Fonseca and Rodrigues [50] found the highest number of phytoplankton species in the Parana River occured during the inundation period. On the other hand, during the drainage phase, after the connection to the river is severed, a heterogeneity in water quality increases among the lakes, influenced by the autochthonous catchment conditions; i.e., geology, soil properties, land cover. Precipitation, groundwater supply, wind, and sedimentation also modify the physical and chemical processes of an isolated floodplain lake [44]. Organic matter in water originates from the in situ processes during that stage [51]. Frequently isolated lakes usually experience massive macrophyte growth [52]. In the last century, the natural connectivity of numerous floodplain lakes has been altered due to human activity [53].
3. Cyanobacterial Blooms in Floodplain Lakes
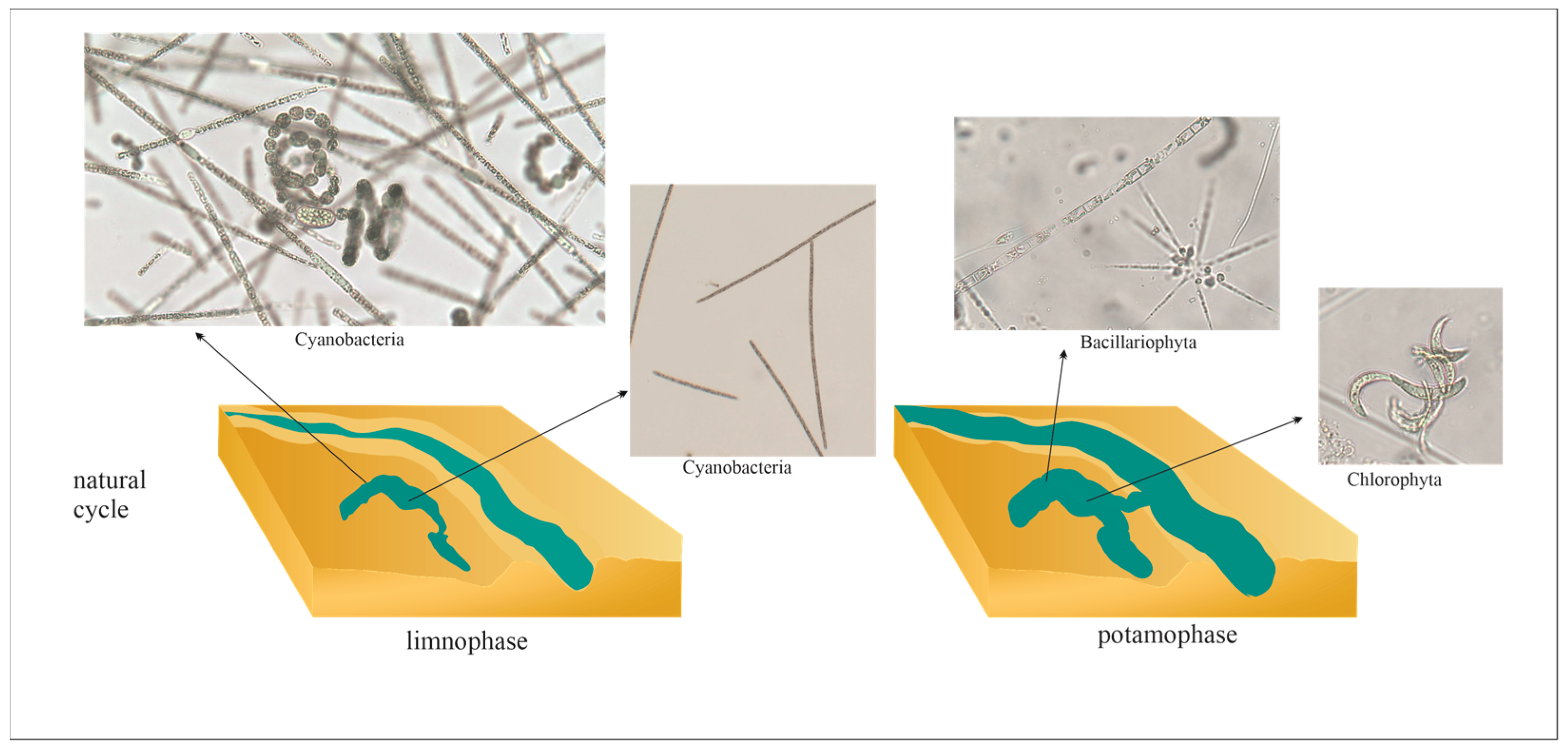
In floodplain lakes, the quantity and quality of water are determined by the functional phases that highly modify lake functioning, including cyanobacterial proliferation. Many studies have pointed out hydrological phases of floodplain lakes as the main factor influencing changes in phytoplankton communities ([49,54] and references therein), showing that some general trends can be featured (Figure 2). During low water (i.e., isolation phase or dry phase), phytoplankton has an autogenic successional sequence with high productivity and biomass, usually with the occurrence of cyanobacterial blooms, whereas during the flooding phase (i.e., high water period), hydrological conditions cause the water column mixing and phytoplankton characterizes higher richness and species diversity. Therefore, in floodplain lakes, which are an integral part of floodplains, phytoplankton composition and biomass frequently change, which reveals the environmental instability of these lakes. Floodplains can be defined as disturbance-dominated systems (harsh ecosystems). What is more, a time shift or the reduction of the disturbance regime, frequency, and magnitude of pulses, can be seen as stress [55]. There is a hypothesis that proposes that a lake’s productivity is driven by the water level based on the “flood pulse” concept, suggesting that seasonally occurring flooding is a major factor in the biotic productivity of the river-floodplain system [56].
In some other aquatic ecosystems, sporadically surprisingly large shifts can occur and they can be attributed to alternative stable states [57]. Water-level fluctuations (WLFs) are considered an important driver not only for the functioning of floodplain lakes [58,59,60], but also for some other shallow lake ecosystems [61], and therefore, extreme levels of water may cause shifts between the turbid and the macrophyte-dominated state [62]. Consequently, dilution and washout can result in significant phytoplankton loss. For instance, Mihaljević and Stević [63] showed that the switch from turbid to a clear state of the Kopački Rit floodplain, one of the largest natural floodplains in the middle section of the Danube River (Europe), occurred in high water levels and nutrient concentrations. This shift was closely related to incidents of extremely high and long-lasting flooding observed during the whole vegetation season. Thus, the hydraulic residence time is one of the main driving forces that shape phytoplankton and its seasonal dynamics in floodplain lakes. It has been determined as a steering force in the phytoplankton dynamics of this type of lakes [64,65]. The massive inflow of floodwater into a lake, (e.g., in the Danube River, the water level increased by 5 m) [63] causes the volume of the lake to rapidly increase, and as a consequence, it overflows, and therefore, a low residence time can be expected [65]. Therefore, dilution and washout significantly affect phytoplankton losses. Among planktonic photosynthetic organisms, cyanobacteria are particularly sensitive to flushing due to the inherently slow rates of cell growth [66], and after flushing they are replaced by smaller and fast-growing algae, mainly diatoms and small chlorococcal species [67].
Harmful cyanobacterial blooms (CyanoHABs) occurring in different fresh and brackish water bodies are of increasing interest to researchers around the world. Cyanobacteria are microscopic and photosynthetic organisms that play an essential role in various, particularly eutrophicated aquatic environments [68,69,70,71]. CyanoHABs, toxins, and other cyanobacterial biologically active metabolites in surface waters are growing global problems escalated by water eutrophication and climate change [69,70]. Huisman and co-authors [32] determined a ‘bloom’ as “a marked visible discoloration of the water that is caused (predominantly) by cyanobacteria” and, for example, according to Nebaeus [72], a chlorophyll-a (Chl-a) concentration over 20 µg/L is treated as a bloom. Bloom-forming cyanobacteria belong to multiple taxa within the coccoid genera Microcystis (Chroococcales), as well as within filamentous genera Dolichospermum (previously Anabaena), Aphanizomenon, Cylindrospermopsis (Nostocales), Planktothrix, and Oscillatoria (Oscillatoriales) [73]. Most of them are capable of producing a wide range of cyanotoxins and other biologically active and potentially harmful metabolites [74,75,76]. In floodplain lakes, blooms of cyanobacteria are mostly created by filamentous species (Table 1) [14,54,63,77,78,79,80,81]. Long-term studies of floodplain lakes located in South America (Argentina) and Europe (Croatia) have revealed very high seasonal and annual variability of cyanobacterial species [54,63,77,78]. Cyanobacteria mostly occurred in combinations of two or three species (Table 1) [14,54,63,77,78].
The success in the predomination of filamentous cyanobacteria may be explained by several competitive characteristics, such as high water temperature, high nutrient concentrations, low water transparency, and mostly by the hydrological regime (Table 1). Cyanobacteria have a high preference for higher water temperatures and concentrations of biogenic compounds, however, these preferences differ among species [82] and among particular functioning groups [83]. For example, nitrogen deficiency was found to favor the occurrence of common N-fixing Nostocales, such as Dolichospermum (previously Anabaena) and Aphanizomenon [83]; on the other hand, Wang et al. [84] showed that N-rich water was also found to support N-fixing Anabaena. Cylindrospermopsis raciborskii, which is an invasive cyanobacterium in Europe, has a particularly high preference for phosphorus and, simultaneously, it has high P-storage capacities [85], whereas Planktothrix agardhii prefers high orthophosphate levels [82]. Oscillatoriales such as P. agardhii and Limnothrix spp., which seasonally co-dominated (often in winter periods) in most of the analyzed floodplain lakes (Table 1) [54,63,77,78,80], prefer turbulent conditions [86]; however, according to Reynolds [83], they are sensitive to flushing. Interestingly, Toporowska and co-authors [71] showed that P. agardhii seemed to be resistant to flushing at very high loads and concentrations of nutrients (mostly P-PO4 and N-NO3). Microcystis spp. were mostly observed as accompanying species [14,77,78] or predominated or co-dominated in large Chinese floodplain lakes [6,58,60]. Large, buoyant Microcystis colonies gain an advantage in the stable water column. For instance, the M. aeruginosa abundance and biovolume negatively correlated with the rate of water flushing in the shallow Mediterranean Spanish Lake Albufera [87]. On the other side, in Poyang Lake, which is one of the few remaining lakes that is freely connected with the Changjiang (Yangtze) River and exhibits highly variable water level fluctuations, Microcystis spp. predominated among cyanobacteria during high water levels (summer seasons); however, the exact cyanobacteria biomass is unknown [58].
Nutrients, both nitrogen and phosphorus, are considered major factors shaping the cyanobacterial abundance and biomass in lakes worldwide. Mihaljević and Stević [63] showed that concentrations of nitrate were significantly lower when the studied floodplain lake was in a turbid state. Thus, very low nitrate concentrations can support the proliferation of species that produce heterocysts [88]. Taxa belonging to the low nitrogen group, such as Dolichospermum, were successful in their blooms in the studied Lake Sakadaš for a short period of time. The blooms usually started with the development of Dolichospermum spp. and Aphanizomenon flos-aquae, and the bloom lasted one month. In the summer, C. raciborskii was dominant or co-dominant. The authors explained that C. raciborskii might produce allelopathic metabolites that could have a strong inhibitory influence on the photosynthetic activity of other phytoplankton species [89]; however, neither cyanotoxins nor other bioactive metabolites were studied [63]. L. redekei accompanied by P. agardhii predominated during the winter bloom. The phenomenon was associated with the low-temperature tolerance of these species [82].
The water temperature is an extremely important environmental factor for most of the bloom-forming cyanobacterial species and their successful proliferation [80], confirmed by CCA analysis for the floodplain of Lake Sakadaš [60]. Mihajevic and Srevic [60] described that the temperature is a very important factor for the development of invasive species, as in shallow and warm floodplain lakes, the highest biomass of C. raciborskii was found at a temperature of 27 °C, which is close to the cyanobacterium temperature requirement in subtropical and tropical areas [90]. On the other hand, a high abundance and biomass of P. agardhii may be observed throughout the whole year, even in winter, despite a preference for this species for higher temperatures, but due to the mentioned tolerance to lower water temperatures [82].

{kind=link}
{kind=link}
Table 1.
Cyanobacterial blooms in floodplain lakes (the selected examples).
| Lakes Trophy, Names and Location, the Study Period | Cyanobacterial Taxa Creating Blooms | Maximal Cyanobacterial Abundance and/or Biomass, Share or Total Phytoplankton Biomass Expressed as the Concentration of Chl-a | Cyanotoxins and Their Concentrations | Factors That Favored Blooms | References |
|---|---|---|---|---|---|
| Eutrophic/hypertrophic Laguna Grande Lake, floodplain Ramsar Paraná de las Palmas and Luján Rivers (Argentina) 1998–2013 | Very high variability of cyanobacterial species from year to year: Planktothrix agardhii and Raphidiopsis mediterranea; Dolichospermum spp. and Anabaenopsis cf. elenkinii; A. cf. elenkinii and Sphaerospermopsis cf. aphanizonemoides accompanied by M. aeruginosa; M. aeruginosa, S. torques-reginae, D. cf. bituri, Cuspidotrix issatschenkoi, S. cf. aphanizonemoides, and R. mediterranea; M. aeruginosa and P. agardhii | 8.5 × 105 colonies/filaments/mL, 670 µg Chl-a/L | l.s. | Extremely low water levels: the effect of water depth, mainly on the acquisition of light and nitrogen. Extreme drought and flood events related to the El Niño Southern Oscillation, which is linked to discharge anomalies in the Paraná River | O’Farrell et al. [77], O’Farrell et al. [78]. |
| Eutrophic/hypertrophic Lake Sakadaš Kopački Rit Danube Floodplain (Croatia) 2003–2008 | Blooms beginning from the development of Nostocales (dominants Dolichospermum spp. and Aphanizomenon flos-aquae) and lasting one month. High variability in patterns of blooming taxa was the main feature of blooms that occurred in summer–autumn periods. C. raciborskii dominanted or co-dominanted in summer. P. agardhii, Pseudanabaena limnetica, L. redekei, Planktolyngbya limnetica was dominant in winter, summer, and autumn. Pseudanabaena limnetica co-occurred with P. agardhii. L. redekei, P. agardhii, and Ps. limnetica were co-dominants or dominants in winter bloom. Only occasionally Merismopedia punctata. | The highest cyanobacterial biomass 61.8 mg/L in the seasons without spring flooding (2003) in comparison with biomass of 10.8 mg/L after the brief spring flood (2007). The bloom absent under conditions of prolonged spring floods (till June 2004 and 2008). The highest biomass equal to 246.6 mg/L | l.s. | High-nutrient concentrations and water column stability, the turbid state during the long-term dry conditions without flooding. The flooding dynamic and intensity as the controlling factor for cyanobacterial blooms in the dynamic river-floodplain system. | Mihaljević, M., and Stević, F. [63]. Stević, F., et al. [54]. |
| Eutrophic/hypertrophic Lakes Dongting and Poyang connected with the river and Lakes Chao and Tai (Taihu) isolated from the river, Yangtze River floodplain (China) 2000–2010 | n.d. * Microcystis spp. | 16.8 µg of Chl-a/L in Poyang Lake 10.0 µg of Chl-a/L in Donting Lake 108 µg of Chl-a/L in Tai Lake 98 µg of Chl-a/L in Poyang Lake | * Dongting Lake: intracellular MC-RR and -LR concentrations were the highest in East Donting Lake * MC-producing Microcystis strains isolated from Lakes Chao and Tai * Lake Tai: First complex study: cyanotoxins predominated by MCs—the highest concentrations were found in November (mean 2.21 µg/L), whereas the lowest amounts were found in February (mean 0.049 µg/L). Cylindrospermopsins concentrations were the highest in May (max. concentration 0.623 µg/L). Saxitoxins only occurred in May (mean concentration 0.019 µg/L) and November (mean 0.199 µg/L). The amount of extracellular T&O was the highest in August—β-cyclocitral predominated (mean 0.241 µg/L) followed by 2-methylisoborneol (mean 0.147 µg/L). | Concentrations of Chl-a considerably higher when relative lake level fluctuations (RLLF) < 100, precipitation < 2.6 mm, wind speed > 2.6 m/s, air temperature > 17.8 °C; Chl-a concentration in river-isolated lakes was more significantly affected by water level amplitude, precipitation, wind speed, and air temperature than in the river-connected lakes | Wu et al. [6]. Huang, J., et al. [60]. Min, et al. [91]. Krüger, et al. [92]. Li, et al. [93] |
| Find floodplain eutrophic lakes including Lake Waaienstein and Lake Ewijk, lower Rhine, (The Netherlands) 1990–1991 | A. flos-aquae predomination in August (summer) only in Lake Waainestein and Lake Ewijk | 110 µg Chl-a/L in Lake Ewijk 55 µg Chl-a/L in Lake Waainetein | l.s. | More frequent events of flood in Lakes Waainestein and Ewijk than in three other studied lakes; floods in winter with river water rich in nutrients | Van den Brink et al. [79]. |
| Eutrophic Iyieke and Ehoma Lakes Cross River floodplain (Nigeria) March 2005–August 2006 | A higher abundance of cyanobacteria during the wet season. Oscillatoria limnetica predominant, O. splendida, Phormidium subcylindrica, Anabaena cylindrica, Aph. flos-aquae, Microcystis flos-aquae, and M. aeruginosa as accompanying taxa | 1.7 × 105 cells/L | l.s. | Nutrient enrichment, water velocity, width, and depth positively influenced cyanobacteria abundance | Okechukwu and Ugwumba Alex O [14]. |
| Smal Winnica 1, Winnica 1a and Martówka Lakes, Larger Lake Port Drzewny, Vistula River (Poland) July and August 2009—before the flood and in July and August 2010-after the flood | Port Drzewny Lake (connected to the river)—annual summer blooms of Aph. flos-aquae, M. wesenbergii and P. agardhii | The highest abundance (17.9 × 106 ind./L) and biomass (4.7 mg/L)in Lake Port Drzewny before the flood | l.s. | Massive flood favored cyanobacterial development in three macrophyte-dominated lakes isolated from the river, but caused a decrease in the cyanobacteria abundance and biomass in lakes that were connected to the river | Dembowska [80]. |
| Eutrophic Lake Poyang Yangtze River floodplain (China) May 2012 to November 2013 | Cyanobacterial dominance in periods of high water levels (from July to August); Microcystis spp. predominated Accompanying taxa: Anabaena spp. Phormidium spp. Planktothrix sp. | 45–93% of the total phytoplankton biomass, the highest peak in August 2013 (93%) | * from January to December 2012: MC-RR was the dominant variant, followed by two isoforms: MC-LR, MC-YR detected in low concentrations. The total concentration of MC (intracellular and extracellular MCs) ranged from 0.0013 to 9.92 µg/L, with an average of 0.47 ng/L. Seasonal variations of MC concentrations. MCs at low concentrations from January to April, then, from June to August, MC concentration rapidly increased and rapidly decreased thereafter. | Large WLFs; high water level observed from April-May to July-August | Liu, X. et al. [58]. Zhang, et al. [94]. |
| Amazonian floodplain eutrophic lakes, Curuai floodplain (Brazil) 2013–2014 | The highest biovolume of cyanobacteria during HW (high water), FL (flushing), and LW (low water) periods. The dominants during HW were Phormidium spp. The dominants during the FL were Dolichospermum spp. and Gleiterinema splendidum. During LW, Oscillatoria spp. and Phormidium spp. predominated. The proportion of cyanobacteria increased along the hydrological cycle from RS to LW | Ca. 98% of the total phytoplankton biomass | l.s. | Nutrient increment in rising periods favored phytoplankton development and increased biomass, which became predominated by cyanobacteria during the low-water period. | Kraus, et al. [81]. |
Note: n.d.—not demonstrated, l.s.—lack of studies, *—data from papers that did not include the impact of hydrology.
Low transparency indicates low light levels as a general habitat template of floodplain lakes during a turbid state. Filamentous cyanobacteria, particularly belonging to Oscillatoriales, can more or less tolerate lower light levels and also create higher turbidity [82,86]. Therefore, a competitive advantage is observed among species, and those with tolerance to high light deficiency, such as Planktothrix, Limnothrix, and C. raciborskii, may be more successful than Aphanizomenon and Dolichosmermum species, which are sensitive to a deficiency of light [54,63,83].
In addition to the influence of in-lake variables on cyanobacteria, the tendency toward cyanobacterial equilibrium in floodplain lakes was pronounced after the extremely long-lasting and high flooding [63,79,80]. Moreover, all cases of steady-state observed in the Bulgarian floodplain lakes were found after drastic changes in the hydrological regime combined with extremely high temperatures [95]. Other studies [54,81] have also indicated that one of the most important environmental factors that controlled the cyanobacterial blooms in floodplain lakes is flooding events, providing the inputs of nutrients that favored cyanobacterial blooms during low-water periods. This also indicates the importance of sediments and their influence on phytoplankton assemblages in wetlands, as the steady state incidents can be associated with it [96] because sediments are part of a lake where the cyanobacterial resting stages survive [97]. Akinetes of Nostocales species (Dolichospermum, Aphanizomenon, Cylindrospermopsis) play a double role as they are inocula for cyanobacterial growth in the following year, and they may assimilate phosphorus (P) during their sedimentary stage which can be used as a P pool of the initial growth [98]. Moreover, for instance, C. raciborskii can form much higher amounts of akinetes per biomass unit than other N-fixing cyanobacteria, which allows its environmental resistance and easy dispersal [94]. In summary, during extremely high flooding occurring in floodplains, the success of cyanobacteria species can be supported by strong interactions between sediments and water [99].
4. Role of Hydrology in the Occurrence of Cyanobacterial Blooms in the Floodplain Lakes
In various water bodies, cyanobacterial predomination and blooms are mostly favored by physical-chemical conditions, including high nutrient concentrations and higher water temperatures [32]. However, in floodplain lakes, except for the abovementioned parameters, the main drivers that affect cyanobacterial blooms are hydrological factors (Table 1). The factors that favor blooms, mainly in less-frequently river-connected lakes, are extremely low water levels, [77,78] a long water residence time, and the stability of the water column [54,63]. On the other hand, in frequently connected lakes, the nutrient enrichment of lakes supplied by river water at simultaneously higher water levels can also drive cyanobacterial blooms in some large floodplain lakes [14,58,79], and particularly in those that are macrophyte-dominated [80]. Nevertheless, in general, both in floodplain lakes and in other non-floodplain lakes, as well as in different flow-through artificial and natural water bodies [70,100,101], eutrophication, high temperatures, and poor mixing are the main factors driving water blooms. For example, in eutrophicated large water bodies with poor vertical mixing [101,102,103] and high water temperatures (>20 °C), the Microcystis abundance can rapidly increase within a few days.
Hydrology, including WLFs, flushing, and water retention time, after eutrophication and temperature conditions, is a very important factor that negatively influences cyanobacterial bloom development and positively influences bloom depletion not only in floodplain lakes (Table 1), but also in flow-through lakes and reservoirs [61,104]. In lakes without outlets, where physical-chemical conditions and area (favoring mixing) are the main drivers of cyanobacterial blooms, the hydrological role is relevant in light of supplying nutrients from the catchment; however, complex lake-catchment processes, rather than single variables, decide the functioning of water bodies, the phytoplankton qualitative and quantitative structure, and water blooms [105].
5. Cyanotoxins in Floodplain Lakes
MCs are toxins that are the most commonly occurring and studied cyanobacterial metabolites [70,71], and they can be supposed to be present in water during the bloom of MC-producers such as Microsycrtis spp., Planktothrix spp., or Dolichospermum spp. [97], observed as dominants in eutrophicated floodplain lakes (Table 1) [91,92,93,94]. MCs are heptapeptides with hepatotoxic properties, but they also exhibit neurotoxicity and nephrotoxicity, and can cause cardiovascular disease, endocrine disruption, and immunomodulation, and they may reveal both reproductive and developmental toxicity [106]. A total of 279 MC variants have been described [106], out of which MC-LR, MC-RR, and MC-YR are the most frequently producing and extensively studied, highly toxic variants. The potential evolutionary roles of cyanotoxin production may be grouped into “competitive advantage” or “physiological aid” [107]. The first is a theory that cyanobacterial toxins and other harmful compounds may play a role in cellular defense mechanisms, in response to grazing pressure and/or resource competition. However, since cyanotoxins were synthetized much earlier than the competitors evolved [108], it seems that the second theory is more possible. It says that cyanobacterial secondary metabolites are produced to improve cellular physiology, through benefits to homeostasis, photosynthetic efficiencies, and accelerated growth rates [107,109].
Studies on the production of cyanotoxins in floodplain lakes are extremely rare [91,92,93,94] and, to the best of our knowledge, there is no information regarding research on the production and seasonal dynamics of cyanobacterial toxins in floodplain lakes in connection with the lake hydrological regime. Zhang and co-authors [94] revealed the seasonal variations of MCs in the floodplain of Poyang Lake (China) in 2012 (Table 1), but they did not study the influence of hydrology. A predominance of microcystin-RR (MC-RR) isoform, followed by MC-LR and MC-YR variants were found, and a maximal total concentration was equal to 9.92 µg/L, but the authors did not include this in their study WLFs and suggested that TN, pH, cyanobacterial biomass, and water temperature might be regulating factors for MC production. MC concentrations were at a low level from January to April, but quickly increased from June to August and dramatically decreased thereafter [94] just after the decrease in water level [58], which suggests that this phenomenon could be an important factor both for cyanobacterial development and MC production. Okechukwu and Ugwumba [14] discussed the possible production of cyanotoxins in the floodplain lakes on the Cross River floodplain in Nigeria (Africa); however, the cyanotoxins were not studied there. Similarly, Affonso et al. [110] suggested that cyanobacterial blooms may be toxic in Amazonian floodplain lakes.
It is important to note that cyanobacteria are able to produce a wide range of toxins different from MCs, and are potential producers of other biologically active compounds. For instance, neurotoxic alkaloids, such as anatoxins and hepatotoxic cylindropsermopsins, are, after MCs, the next very well-known and most common groups of cyanobacterial toxins [74,75]. Cyanotoxins are harmful to aquatic ecosystems as they can exert severe negative effects on ecosystems and organisms belonging to microorganisms, plants, animals, and humans [74]. Since many surface water bodies are used as a source of drinking water and food, and/or are used for recreation, the human health problems connected with cyanotoxicity have been well documented [74,111,112,113,114]. Some planktonic cyanobacteria can produce biologically active metabolites other than well-known cyanotoxins. Some of these compounds, similarly to MCs, mostly belong to non-ribosomal oligopeptides. Cyanobacterial aeruginosins, anabaenopeptins, aerucyclamides, cyanopeptolins, and microviridins are mostly enzyme inhibitors [75]. Other metabolites such as dermatotoxins (lyngbyatoxin-a, aplasiotoxin, debromoaplysiatoxin, lipopolysaccharides), or taste and odor compounds (T&O), that were found for example in the floodplain of Tai Lake (China) [93] may also be harmful to living organisms, the environment, and/or humans [115,116]. Moreover, Miller and co-authors [117] confirmed the deaths of sea otters caused by MC intoxication and suggested that the most probable route of exposure was the MC land–sea flow with trophic transfer through marine invertebrates. This finding indicates that cyanotoxins and/or other bioactive compounds may negatively affect aquatic ecosystems, even far downstream from their origin, because of fluvial transport. Okechukwu and Ugwumba [14] showed that the interaction between the lentic and lotic ecosystems of floodplains during the limnophase and the potamophase leads to river inoculation, with cyanobacteria blooming in the floodplain lakes. In general, in some lakes, phytoplankton can rapidly develop for one or two months following the flood [80,118]. For example, extreme floods occurring in floodplain lakes usually isolated from the river can increase the abundance and biomass of phytoplankton, including cyanobacteria, particularly in less eutrophic, macrophyte-dominated lakes; whereas, in eutrophicated lakes connected with the rivers, the reverse situation can be observed [80]: the phytoplankton abundance and biomass (often predominated by filamentous cyanobacteria) decrease. However, the dynamics of cyanotoxins and other cyanobacterial metabolites in floodplains remain unknown.
6. Role of Hydrology in Cyanotoxin Production in the Floodplain Lakes
Both in floodplain lakes and non-floodplain lakes, there is one main factor that decides the production of cyanotoxins: the presence and development of cyanobacterial strains able to produce MCs and other biologically active metabolites [103,108]. On the other hand, it seems that, in floodplain and non-floodplain lakes, the environmental factors influencing cyanotoxin production and concentration may be different, and mostly depend on hydrology. The role of hydrology has been illustrated in a study [61] carried out in four lakes modified into water storage reservoirs (with periodically regulated water levels) and included in the river drainage canal system (Wieprz-Krzna). The obtained result showed that in two larger lakes, into which nutrient-rich river water was supplied once a year, lower cyanobacterial species diversity, higher biomass of cyanobacteria belonging to Microcystis, Aphanizomenon, and Dolichospermum, and higher MCs concentrations were found. In two other lakes, with several irregular water-level manipulations per year (alternating outputs and inputs), higher species diversity and lower cyanobacterial biomass and MC concentrations were observed. There were also differences in MC variants observed. Intracellular MC-LF, -LR, and -RR produced by Microcystis spp. predominated in the more stable lakes, whereas in the lakes with frequent water manipulations, different Oscillatoriales also produced other MC isoforms (MC-LA, -LY, -LW, and -WR). This study showed that the frequency of water manipulations was the most significant factor for the composition of cyanobacterial communities and that a higher frequency of WLFs, which increases the flushing rate, was beneficial to nutrient-rich water reservoirs. The study carried out in the Mediterranean Lake Albufera [87] also showed that longer water residence time increased total cyanobacterial (M. aeruginosa) biomass, and MC concentrations in the lake water, as well as in seston. Moreover, flows lower than 10 m3/s raised toxicological risk from low to moderate-high. The same phenomena may occur in floodplain lakes, and further studies in this field are urgently required. Data showing low concentrations (in comparison with other water bodies [61,70,100,101]) of cyanotoxins in large Chinese floodplain lakes (Table 1) suggest that the environmental conditions in these lakes are not favorable for high toxin production and concentrations. Moreover, frequent WLFs, which are stress factors, may cause cell breakdown and the release of extracellular toxins. This phenomenon may be dangerous in water bodies which are reservoirs of drinking water, and needs to be explored in floodplain lakes connected to the rivers.
7. Water Management Practices in Floodplain Lakes
The hydrological functioning of floodplain lakes significantly differs from other types of lakes. Thus, the proportion of water balance elements remarkably varies. Regardless of the type of floodplain lake connection to the main river, horizontal water balance elements (fluvial input and output) prevail over the vertical ones, such as precipitation and evaporation [119,120].
Studies have shown that maintaining hydrological connectivity is a key factor in the survival of floodplain lakes [121]. The natural inundation cycles have been disturbed as a result of climate change and human activities. Thus, floodplain areas are shrinking on a global scale, and a large number of FPLs have become permanently isolated from the parent river [122]. The loss of fluvial input within the floodplain may result in a fragmentation of habitats and a decrease in biodiversity [123], as well as a decrease in within-lake habitat heterogeneity [124]. Thus, the remediation of floodplain lakes is usually focused on the preservation of hydrological connectivity (water, matter, and organisms exchange) between lakes and the parent river [125]. Water management practices may result in the creation of open, partial, or closed ecosystems in terms of their connection within the floodplain [126]. The strengthening hydrological link between the water bodies of the floodplain area, by re-establishing flood channels and/or the re-connection of floodplain lakes, is the primary goal of the remediation processes [43,47]. Environmental gains of controlled inundation are equalized WLFs, preventing extreme water shortage and floods of high magnitudes. Furthermore, as a result, the capacity of water and matter storage in floodplain ecosystems is increased, strengthening connections between biotic and abiotic elements of the system [127] and ecosystem services [128]. Regulation techniques aim to increase the self-regulation ability of floodplain sites [129] and their perseverance against human pressure [130]. Studies have shown that restoration practices bring about an increase in the diversity of many groups of organisms; e.g., macroinvertebrates. Maintaining frequent flooding is also crucial for fish sprawling [131], and enables FPLs to act as a filter of phosphorus and nitrogen for the river [132]. On the other hand, the higher the connectivity, the easier the transport of sediments and nutrients [126] and the more increased the mobility of the invasive species [133]. Fruitful and cost-effective management practices have to be controlled with long-term environmental monitoring [134]. Each ecosystem is an individual, and no universal practices have been developed to date [135]. Increasing flood risk and continuous climate change make management practices very difficult.
8. Future Changes of Floodplain Lakes Due to Climate Changes
Many studies of floodplains worldwide have focused on climate change [136], mostly on the dynamic of precipitation and temperature [137]. Usually, small and shallow floodplain lakes have been considered to be extremely sensitive to climate change [7]. Large-scale analyses [138], as well as smaller-scale and regional studies [139,140], have focused on flooding pattern alterations due to climate change, which has already been observed at the macro-scale [141]. Disturbances of natural potamophase and limnophase patterns, prolonged periods of isolation, and extreme floods may be destructive for floodplain lake ecosystems [142]. Estimations of the influence of climate change on extreme hydrologic events is very difficult, due to their interdependence and the impact of other natural factors: ice or snow cover formation, soil conditions, or catchment water resources (soil moisture, surface and ground-water levels) [138]. It is expected that changes in the sea level also influence river inundation cycles [143,144].
A more frequent occurrence of extreme floods and droughts has been observed worldwide [145,146,147]. Studies have shown that altered connectivity patterns increase CO2 and CH4 emissions from floodplains [148]. Barbosa et al. [149] observed high fluxes of methane after a prolonged drought period in the Amazon catchment as a result of disturbed biological processes. In the last two decades, the frequency of occurrence of extreme floods and droughts in the world’s largest river basin has increased, and the far-reaching ecological implications of the process are impossible to predict. An improvement in water quality following periods of extreme floods was observed in some floodplain lakes with regard to the regular flooding cycle [150]. The process of sediment resuspension during extreme floods was also observed [151]. The water quality of the FPLs is also a derivative of lake volume and temperature distribution in the water column [152]. The increased volume may cause a dilution effect, a decrease in cyanobacterial biomass [138], and an increase in the phytoplankton diversity index [152]. On the other hand, low lake volumes bring about higher concentrations of nutrients in the water solution [153]. Furthermore, fluctuations in river water discharge and patterns of potamophases change the water residence time of floodplain lakes [154], a key factor driving the phytoplankton dynamic [155].
Changes in water temperature may have a far-reaching effect on aquatic life [156]. The extension of thermal optima may accelerate cyanobacteria blooms [157]. Increasing global air temperature is expected to cause more rapid precipitation [158] due to the higher water-holding capacity of the atmosphere [159]. Therefore, the increase in sediment load and nutrient load, favoring cyanobacterial occurrence, is expected [160]. Higher surface runoff and matter transport will, in turn, negatively impact light penetration in the water column, favoring cyanobacteria domination [161].
9. Conclusions
The structure and dynamics of cyanobacterial communities and blooms occurring in floodplain lakes have been rarely studied, but it has been well documented that cyanobacterial blooms in floodplain lakes are regulated (next to high levels of nutrients and high temperatures) by the hydrological regime, and the blooms are mostly predominated by filamentous species, which are common cyanotoxin producers. However, to the best of our knowledge, there has been no research carried out on the influence of functional phases on cyanotoxins production and dynamics in floodplains. Consequently, the risk of the negative effects of cyanobacterial blooms on FPLs, their biota, and humans, as well as ecosystems located downstream, seasonally fed by waters containing cyanobacteria and cyanotoxins, is unknown. Therefore, intensive multidisciplinary studies in this field are strongly required. Water level control (e.g., projects like dams), by altering natural potamophases and limnophases cycles, may affect phytoplankton and cyanobacterial biomass and assemblage structure patterns. However, all management activities must be safe for floodplains and planned in accordance with good practices of adaptation to climate change and wetland conservation. Floodplain environments, historically impacted by human pressure (reclamation, eutrophication, drainage) are extremely sensitive to climate changes, especially to more frequently observed major droughts and floods. Extreme flooding usually results in water quality improvement in the FPLs, but extreme and prolonged droughts give converse results.
Author Contributions
Conceptualization, B.F. and M.T.; methodology, M.T., B.F. and J.D.; formal analysis, B.F., M.T. and J.D.; investigation, B.F., M.T. and J.D.; resources, B.F.; writing—original draft preparation, B.F. and M.T.; writing—review and editing, B.F.; validation, B.F.; visualization, B.F. and M.T.; supervision, B.F.; project administration, B.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are available upon request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Thomas, R.F.; Kingsford, R.T.; Lu, Y.; Cox, S.J.; Sims, N.C.; Hunter, S.J. Mapping inundation in the heterogeneous floodplain wetlands of the Macquarie Marshes, using Landsat Thematic Mapper. J. Hydrol. 2015, 524, 194–213. [Google Scholar] [CrossRef]
- Ogbeide, O.; Uhunamure, G.; Uwagboe, L.; Osakpamwan, T.; Glory, M.; Chukwuka, A. Comparative gill and liver pathology of tilapia zilli, Clarias gariepinus and Neochanna diversus in owan river (Nigeria): Relative ecological risks of species in a pesticide-impacted river. Chemosphere 2019, 234, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pettit, N.E.; Naiman, R.J.; Warfe, D.M.; Jardine, T.D.; Douglas, M.M.; Bunn, S.E.; Davies, P.M. Productivity and connectivity in tropical riverscapes of northern Australia: Ecological insights for management. Ecosystems 2017, 20, 492–514. [Google Scholar] [CrossRef]
- Crook, D.A.; Lowe, W.H.; Allendorf, F.W.; Erős, T.; Finn, D.S.; Gillanders, B.M.; Hadwen, W.L.; Harrod, C.; Hermoso, V.; Jennings, S.; et al. Human Effects on Ecological Connectivity in Aquatic Ecosystems: Integrating Scientific Approaches to Support Management and Mitigation. Sci. Total Environ. 2015, 534, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Melack, J.M.; Forsberg, B.R. Biogeochemistry of Amazon Floodplain Lakes and Associated Wetlands: Biogeochemistry of the Amazon Basin; Oxford University Press: Oxford, UK, 2001; pp. 235–274. [Google Scholar]
- Khaki, M.; Awange, J. Improved remotely sensed satellite products for studying Lake Victoria’s water storage changes. Sci. Total Environ. 2018, 652, 915–926. [Google Scholar] [CrossRef]
- Gmitrowicz-Iwan, J.; Ligęza, S.; Pranagal, J.; Smal, H.; Olenderek, H. Floodplain Reservoirs in the Face of Climate Change—Sink or Source of Nutrients? Water 2020, 12, 3423. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The floodpulse concept in river-floodplain systems. Can. Fish. Aquat. Sci. Spec. Publ. 1989, 106, 110–127. [Google Scholar]
- Fantin-Cruz, I.; Loverde-Oliveira, S.; Girard, P. Morphometric characterization and its limnological implications in Northern Pantanal lakes. Acta Sci. Biol. Sci. 2008, 30, 133–140. [Google Scholar]
- Neiff, J.J. Ideas para la interpretacion ecológica del Parana. Interciencia 1990, 15, 424–441. [Google Scholar]
- Thomaz, S.M.; Pagioro, T.A.; Bini, L.M.; Roberto, M.C.; Rocha, R.R.A. The Upper Parana River and Itsfloodplain, Physicalaspects, Ecology and Conservation; Thomaz, S.M., Agostinho, A.A., Hahn, N.S., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2004. [Google Scholar]
- Yao, J.; Zhang, Q.; Ye, X.; Zhang, D.; Bai, P. Quantifying the impact of bathymetric changes on the hydrological regimes in a large floodplain lake: Poyang Lake. J. Hydrol. 2018, 561, 711–723. [Google Scholar] [CrossRef]
- Reisenbüchler, M.; Bui, M.D.; Skublics, D.; Rutschmann, P. An integrated approach for investigating the correlation between floods and river morphology: A case study of the Saalach River, Germany. Sci. Total Environ. 2019, 647, 814–826. [Google Scholar] [CrossRef] [PubMed]
- Okechukwu, I.; Ugwumba, O.; Alex, O. Cyanobacteria abundance and its relationship to water quality in the Mid-Cross River floodplain, Nigeria. Rev. Biol. Trop. 2009, 57, 33–43. [Google Scholar] [CrossRef] [PubMed]
- Câmara Dos Reis, M.; Lacativa Bagatini, I.; de Oliveira Vidal, L.; Bonnet, M.P.; da Motta Marques, D.; Sarmento, H. Spatial heterogeneity and hydrological fluctuations drive bacterioplankton community composition in an Amazon floodplain system. PLoS ONE 2019, 14, e0220695. [Google Scholar] [CrossRef]
- Wolf, S.; Esser, V.; Schüttrumpfl, H.; Lehmkuhl, F. Infuence of 200 years of water resource management on a typical central European river. Does industrialization straighten a river? Environ. Sci. Eur. 2021, 33, 15. [Google Scholar] [CrossRef]
- Souza-Filho, E.E.; Zanetti, K.; Peccinini, J.P.; França, V. A hidroquímica do rio Paraná após a barragem de Porto Primavera. Bol. Geogr. 2010, 28, 101–112. [Google Scholar] [CrossRef]
- Hamilton, S.K.; Lewis, W.M. Basin morphology in relation to chemical and ecological characteristics of lakes on the Orinoco River floodplain, Venezuela. Arch. Hydrobiol. 1990, 119, 393–425. [Google Scholar] [CrossRef]
- García de Emiliani, M.O. Effects of water level fluctuations on phytoplankton in a river-floodplain lake system (Paraná River, Argentina). Hydrobiologia 1997, 357, 1–15. [Google Scholar] [CrossRef]
- Dawidek, J.; Ferencz, B. Hydrological processes in the riverine systems, the origin and classifications of floodplain lakes. Ecológia 2012, 31, 331–340. [Google Scholar] [CrossRef]
- Lesack, L.F.W.; Melack, J.M. Flooding hydrology and mixture dynamics of lake water derived from multiple sources in an Amazon floodplain lake. Water Resour. Res. 1995, 31, 329–345. [Google Scholar] [CrossRef]
- Neiff, J.J. Diversity in some tropical wetland systems of South América. In Biodiversity in Wetlands: Assessment, Function and Conservation; Gopal, B., Junk, W.J., Davis, J.A., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2001; pp. 157–186. [Google Scholar]
- Scott, D.T.; Keim, R.F.; Edwards, B.L.; Jones, C.N.; Kroes, D.E. Floodplain Biogeochemical Processing of Floodwaters in the Atchafalaya River Basin during the Mississippi River Flood of 2011. J. Geophys. Res. Biogeosci. 2014, 119, 537–546. [Google Scholar] [CrossRef]
- Jones, C.N.; Scott, D.T.; Guth, C.; Hester, E.T.; Hession, W.C. Seasonal Variation in Floodplain Biogeochemical Processing in a Restored Headwater Stream. Environ. Sci. Technol. 2015, 49, 13190–13198. [Google Scholar] [CrossRef] [PubMed]
- Reid, M.A.; Reid, M.C.; Thoms, M.C. Ecological significance of hydrological connectivity for wetland plant communities on a dryland floodplain river, Macintyre river, Australia. Aquat. Sci. 2016, 78, 139–158. [Google Scholar] [CrossRef]
- Zhang, Y.; Huang, C.; Zhang, W. The concept, approach, and future research of hydrological connectivity and its assessment at multiscales. Environ. Sci. Pollut. Res. 2021, 28, 52724–52743. [Google Scholar] [CrossRef]
- Saha, S.; Chukwuka, A.V.; Mukherjee, D.; Saha, N.C.; Adeogun, A.O. Hydrological connectivity, surface water quality and distribution of fish species within sub-locations of an urban oxbow lake, East India. Watershed Ecol. Environ. 2022, 4, 44–58. [Google Scholar] [CrossRef]
- Napiórkowski, P.; Bąkowska, M.; Mrozińska, N.; Szymańska, M.; Kolarova, N.; Obolewski, K. The Effect of Hydrological Connectivity on the Zooplankton Structure in Floodplain Lakes of a Regulated Large River (the Lower Vistula, Poland). Water 2019, 11, 1924. [Google Scholar] [CrossRef]
- Jacobson, R.B.; Bouska, K.L.; Bulliner, E.A.; Lindner, G.A.; Paukert, C.P. Geomorphic controls on floodplain connectivity, ecosystem services, and sensitivity to climate change: An example from the lower Missouri River. Water Resour. Res. 2022, 58, e2021WR031204. [Google Scholar] [CrossRef]
- López-Vicente, M.; Ben-Salem, N. Computing structural and functional flow and sediment connectivity with a new aggregated index: A case study in a large Mediterranean catchment. Sci. Total Environ. 2019, 651 Pt 1, 179–191. [Google Scholar] [CrossRef]
- De Melo, M.L.; Bertilsson, S.; Henrique, J.; Amaral, F.; Barbosa, P.M.; Forsberg, B.R.; Sarmentoet, H. Flood pulse regulation of bacterioplankton community composition in an Amazonian floodplain lake. Freshw. Biol. 2019, 64, 108–120. [Google Scholar] [CrossRef]
- Huisman, J.; Codd, G.A.; Paerl, H.W.; Ibelings, B.W.; Verspagen, J.M.; Visser, P.M. Cyanobacterial blooms. Nat. Rev. Microbiol. 2018, 16, 471–483. [Google Scholar] [CrossRef]
- Wang, K.; Razzano, M.; Mou, X. Cyanobacterial blooms alter the relative importance of neutral and selective processes in assembling freshwater bacterioplankton community. Sci. Total Environ. 2020, 706, 135724. [Google Scholar] [CrossRef]
- Whitehead, P.G.; Wilby, R.L.; Battarbee, R.W.; Kernan, M.; Wade, A.J. A review of the potential impacts of climate change on surface water quality. Hydrol. Sci. J. 2009, 54, 101–123. [Google Scholar] [CrossRef]
- Gallardo, B.; Cabezas, A.; Gonzalez, E.; Comínet, F.A. Effectiveness of a Newly Created Oxbow Laketo Mitigate Habitat Loss and Increase Biodiversityin a Regulated Floodplain. Restor. Ecol. 2012, 20, 387–394. [Google Scholar] [CrossRef]
- Gallardo, B.; Dolédec, S.; Paillex, A.; Arscott, D.B.; Sheldon, F.; Zilli, F. Response of benthic macroinvertebrates to gradients in hydrological connectivity: A comparison of temperate, subtropical, Mediterranean and semiarid river floodplains. Freshw. Biol. 2014, 59, 630–648. [Google Scholar] [CrossRef]
- Affonso, A.G.; Queiroz, H.L.; Novo, E.M.L.M. Limnological characterization of floodplain lakes in Mamirauá Sustainable Development Reserve, Central Amazon (Amazonas State, Brazil). Acta Limnol. Bras. 2011, 23, 95–108. [Google Scholar] [CrossRef]
- Larned, S.T.; Datry, T.; Arscott, D.B.; Tockner, K. Emerging concepts in temporary river ecology. Freshw. Biol. 2010, 55, 717–738. [Google Scholar] [CrossRef]
- Freeman, M.C.; Pringle, C.M.; Jackson, C.R. Hydrologic connectivity and the contribution of stream headwaters to ecological integrity at regional scales. J. Am. Water Resour. Assoc. 2007, 43, 5–14. [Google Scholar] [CrossRef]
- Knutti, J.; Braunisch, V.; Pellet, J.; Arlettaz, R. Improving longitudinal habitat connectivity in major river restoration projects through farmland re-allocation. J. Nat. Conserv. 2021, 64, 126062. [Google Scholar] [CrossRef]
- Paira, A.R.; Drago, E.C. Origin, Evolution, and Types of Floodplain Water Bodies. In The Middle Paraná River; Iriondo, M.H., Paggi, J.C., Parma, M.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2007. [Google Scholar] [CrossRef]
- Karpack, M.N.; Morrison, R.R.; McManamay, R.A. Quantitative assessment of floodplain functionality using an index of integrity. Ecol. Indic. 2020, 111, 106051. [Google Scholar] [CrossRef]
- Obolewski, K.; Glińska-Lewczuk, K.; Ożgo, M.; Astel, A. Connectivity restoration of floodplain lakes: An assessment based on macroinvertebrate communities. Hydrobiologia 2016, 774, 23–37. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Bini, L.M.; Bozelli, R.L. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 2007, 579, 1–13. [Google Scholar] [CrossRef]
- MacKinnon, B.D.; Sagin, J.; Baulch, H.M.; Lindenschmidt, K.E.; Jardine, T.D. Influence of hydrological connectivity on winter limnology in floodplain of the Saskatchewan River Delta, Saskatchewan. Can. J. Fish. Aquat. Sci. 2016, 73, 140–152. [Google Scholar] [CrossRef]
- Wiklund, J.A.; Hall, R.I.; Wolfe, B.B.; Edwards, T.W.D.; Farwell, A.J.; Dixon, D.G. Has Alberta oil sands development increased far-field delivery of airborne contaminants to the Peace-Athabasca Delta? Sci. Total Environ. 2012, 433, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Seidel, M.; Voigt, M.; Langheinrich, U.; Hoge-Becker, A.; Gersberg, R.M.; Arévalo, J.R.; Lüderitz, V. Reconnection of oxbow lakes as an effective measure of river restoration. CLEAN Soil Air Water 2017, 45, 1600211. [Google Scholar] [CrossRef]
- Paillex, A.; Castella, E.; zu Ergmassen, P.S.E.; Aldridge, D.C. Testing Predictions of Changes in Alien and Native Macroinvertebrate Communities and Their Interaction After the Restoration of a Large River Floodplain (French Rhône). Freshw. Biol. 2015, 60, 1162–1175. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, Z.; Wang, W.; Zhou, Z.; Ye, X. Effects of Flood on Phytoplankton Diversity and Community Structure in Floodplain Lakes Connected to the Yangtze River. Diversity 2022, 14, 581. [Google Scholar] [CrossRef]
- Fonseca, I.A.; Rodrigues, L. Periphytic cyanobacteria in different environments from the upper Paraná river floodplain, Brazil. Acta Limnol. Bras. 2007, 19, 53–65. [Google Scholar]
- Amaral, J.H.F.; Borges, A.V.; Melack, J.M.; Sarmento, H.; Barbosa, P.M.; Kasper, D.; Forsberg, B.R. Influence of plankton metabolism and mixing depth on CO2 dynamics in an Amazon floodplain lake. Sci. Total Environ. 2018, 630, 1381–1393. [Google Scholar] [CrossRef]
- Sokal, M.A.; Hall, R.I.; Wolfe, B.B. The role of flooding on inter-annual and seasonal variability of lake water chemistry, phytoplankton diatom communities and macrophyte biomass in the Slave River Delta (Northwest Territories, Canada). Ecohydrology 2010, 3, 41–54. [Google Scholar] [CrossRef]
- Amezcua, F.; Rajnohova, J.; Flores-de-Santiago, F. The Effect of Hydrological Connectivity on Fish Assemblages in a Floodplain System From the South-East Gulf of California, Mexico. Front. Mar. Sci. 2019, 6, 240. [Google Scholar] [CrossRef]
- Stević, F.; Mihaljević, M.; Špoljarić, D. Changes of phytoplankton functional groups in a floodplain lake associated with hydrological perturbations. Hydrobiologia 2013, 709, 143–158. [Google Scholar] [CrossRef]
- Tockner, K.; Pusch, M.; Borchardt, D.; Lorang, M.S. Multiple stressors in coupled river–floodplain ecosystems. Freshw. Biol. 2010, 55, 135–151. [Google Scholar] [CrossRef]
- Tockner, K.; Malard, F.; Ward, J.V. An extension of the flood pulse concept. Hydrol. Process. 2000, 14, 2861–2883. [Google Scholar] [CrossRef]
- Scheffer, M.; Carpenter, S.R. Catastrophic regime shifts in ecosystems: Linking theory to observation. Trends Ecol. Evol. 2003, 18, 648–656. [Google Scholar] [CrossRef]
- Liu, X.; Qian, K.; Chen, Y. Effects of water level fluctuations on phytoplankton in a Changjiang River floodplain lake (Poyang Lake): Implications for dam operations. J. Great Lakes Res. 2015, 41, 770–779. [Google Scholar] [CrossRef]
- Wu, S.; Wang, S.; Yang, H.; Xie, P.; Ni, L.; Xu, J. Field studies on the environmental factors in controlling microcystin production in the subtropical shallow lakes of the Yangtze River. Bull. Environ. Contam. Toxicol. 2008, 80, 329–334. [Google Scholar] [CrossRef]
- Huang, J.; Xu, Q.; Wang, X.; Ji, H.; Quigley, E.J.; Sharbatmaleki, M.; Li, C. Effects of hydrological and climatic variables on cyanobacterial blooms in four large shallow lakes fed by the Yangtze River. Environ. Sci. Ecotechnol. 2021, 5, 100069. [Google Scholar] [CrossRef] [PubMed]
- Pawlik-Skowronska, B.; Toporowska, M. How to mitigate cyanobacterial blooms and cyanotoxin production in eutrophic water reservoirs? Hydrobiologia 2016, 778, 45–59. [Google Scholar] [CrossRef]
- Coops, H.; Beklioglu, M.; Crisman, T.L. The role of water-level fluctuations in shallow lake ecosystems–workshop conclusions. Hydrobiologia 2003, 506, 23–27. [Google Scholar] [CrossRef]
- Mihaljević, M.; Stević, F. Cyanobacterial blooms in a temperate river-floodplain ecosystem: The importance of hydrological extremes. Aquat. Ecol. 2011, 45, 335–349. [Google Scholar] [CrossRef]
- Oosterberg, W.; Staras, M.; Bogdan, L.; Buijse, A.D.; Constantinescu, A.; Coops, H.; Hanganu, J.; Ibelings, B.W.; Menting, G.A.M.; Nãvodaru, I.; et al. Ecological Gradients in the Danube Delta Lakes: Present State and Man-Induced Changes: RIZA, Danube Delta National Institute Romania and Danube Delta Biosphere Reserve Authority Romania, The Netherlands; RIZA Rapport no. 2000.015; RIZA: Lelystad, The Netherlands, 2000. [Google Scholar]
- Schagerl, M.; Drozdowski, I.; Angeler, D.G.; Hein, T.; Preiner, S. Water age—A major factor controlling phytoplankton community structure in a reconnected dynamic floodplain (Danube, Regelsbrunn, Austria). J. Limnol. 2009, 68, 274–287. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Petersen, A.C. The distribution of planktonic cyanobacteria in Irish lakes in relation to their trophic states. Hydrobiologia 2000, 424, 91–99. [Google Scholar] [CrossRef]
- Mihaljević, M.; Špoljarić, D.; Stević, F.; Cvijanović, V.; Kutuzović, B.H. The influence of extreme floods from the River Danube in 2006 on phytoplankton communities in a floodplain lake: Shift to a clear state. Limnologica 2010, 40, 260–268. [Google Scholar] [CrossRef]
- Whitton, B.A.; Potts, M. (Eds.) The Ecology of Cyanobacteria: Their Diversity in Time and Space; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2007. [Google Scholar]
- Paerl, H.W.; Huisman, J. Blooms like it hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Mantzouki, E.; Lürling, M.; Fastner, J.; de Senerpont Domis, L.; Wilk-Woźniak, E.; Koreiviene, J.; Seelen, L.; Teurlincx, S.; Verstijnen, Y.; Krztoń, W.; et al. Temperature effects explain continental scale distributon of cyanobacterial toxins. Toxins 2018, 10, 156. [Google Scholar] [CrossRef] [PubMed]
- Toporowska, M.; Pawlik-Skowrońska, B.; Kalinowska, R. Mass development of diazotrophic cyanobacteria (Nostocales) and production of neurotoxic anatoxin-a in a Planktothrix (Oscillatoriales) dominated temperate lake. Water Air Soil Pollut. 2016, 227, 321. [Google Scholar] [CrossRef] [PubMed]
- Nebaeus, M. Algal water-blooms under ice-cover. Int. Ver. Theor. Angew. Limnol. 1984, 22, 719–724. [Google Scholar] [CrossRef]
- Chorus, I.; Welker, M. Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Taylor & Francis: Abingdon-on-Thames, UK, 2021; 858p. [Google Scholar]
- Codd, G.; Bell, S.; Kaya, K.; Ward, C.; Beattie, K.; Metcalf, J. Cyanobacterial toxins, exposure routes and human health. Eur. J. Phycol. 1999, 34, 405–415. [Google Scholar] [CrossRef]
- Sivonen, K.; Börner, T. Bioactive Compounds Produced by Cyanobacteria. In The Cyanobacteria; Caister Academic Press: Poole, UK, 2008; pp. 159–197. [Google Scholar]
- Toporowska, M.; Mazur-Marzec, H.; Pawlik-Skowrońska, B. The effects of cyanobacterial bloom extracts on the biomass, Chl-a, MC and other oligopeptides contents in a natural Planktothrix agardhii population. Int. J. Environ. Res. Public Health 2020, 17, 2881. [Google Scholar] [CrossRef]
- O’Farrell, I.; Izaguirre, I.; Chaparro, G.; Unrein, F.; Sinistro, R.; Pizarro, H.; Tell, G. Water level as the main driver of the alternation between a free-floating plant and a phytoplankton dominated state: A long-term study in a floodplain lake. Aquat. Sci. 2011, 73, 275–287. [Google Scholar] [CrossRef]
- O’Farrell, I.; Vinocur, A.; de Tezanos Pinto, P. Long-term study of bloom-forming cyanobacteria in a highly fluctuating vegetated floodplain lake: A morpho-functional approach. Hydrobiologia 2015, 752, 91–102. [Google Scholar] [CrossRef]
- Van den Brink, F.W.B.; De Leeuw, J.P.H.M.; Van der Velde, G.; Verheggen, G.M. Impact of hydrology on the chemistry and phytoplankton development in floodplain lakes along the Lower Rhine and Meuse. Biogeochemistry 1993, 19, 103–128. [Google Scholar] [CrossRef]
- Dembowska, E.A. The impact of an extreme flood in spring/summer 2010 on phytoplankton communities in oxbow lakes of the lower Vistula River, central Poland. Ann. Limnol. Int. J. Limnol. 2017, 53, 19–26. [Google Scholar] [CrossRef]
- Kraus, C.N.; Bonnet, M.P.; de Souza Nogueira, I.; Morais Pereira Souza Lobo, M.T.; da Motta Marques, D.; Garnier, J.; Cardoso Galli Vieira, L. Unraveling flooding dynamics and nutrients’ controls upon phytoplankton functional dynamics in Amazonian floodplain lakes. Water 2019, 11, 154. [Google Scholar] [CrossRef]
- Dokulil, M.T.; Teubner, K. Cyanobacterial dominance in lakes. Hydrobiologia 2000, 438, 1–12. [Google Scholar] [CrossRef]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli-Flores, L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Wang, K.; Mou, X.; Cao, H.; Struewing, I.; Allen, J.; Lu, J. Co-occurring microorganisms regulate the succession of cyanobacterial harmful algal blooms. Environ. Pollut. 2021, 288, 117682. [Google Scholar] [CrossRef]
- Isvánovics, V.; Shafik, H.M.; Présing, M.; Juhos, S. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow cultures. Freshw. Biol. 2000, 43, 257–275. [Google Scholar] [CrossRef]
- Scheffer, M.; Rinaldi, S.; Gragnani, A.; Mur, L.R.; van Nes, E.H. On the dominance of filamentous cyanobacteria in shallow, turbid lakes. Ecology 1997, 78, 272–282. [Google Scholar] [CrossRef]
- Romo, S.; Soria, J.; Fernández, F.; Ouahid, J.; Barón-Solá, A. Water residence time and the dynamics of toxic cyanobacteria. Freshw. Biol. 2013, 58, 513–522. [Google Scholar] [CrossRef]
- Briand, J.F.; Robillot, C.; Quiblier-Llobéras, C.; Humbert, J.F.; Coute’, A.; Bernard, C. Environmental context of Cylindrospermopsis raciborskii (Cyanobacteria) blooms in a shallow pond in France. Water Res. 2002, 36, 3183–3192. [Google Scholar] [CrossRef]
- Figueredo, C.C.; Giani, A.; Bird, D.F. Does allelopathy contribute to Cylindrospermopsis raciborskii blooms occurrence and geographic expansion? J. Phycol. 2007, 43, 256–265. [Google Scholar] [CrossRef]
- Padisák, J. Cylindrospermopsis raciborskii (Wołoszynska) Seenayya et Subba Raju, an expanding, highly adaptative cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. Suppl. 1997, 107, 563–593. [Google Scholar]
- Min, X.; Xu, X.D.; Hong, G.; Kong, R.Q. High variability of the gvpA-gvpC region in Microcystis. Prog. Nat. Sci. 2007, 17, 1290–1295. [Google Scholar]
- Krüger, T.; Wiegand, C.; Kun, L.; Luckas, B.; Pflugmacher, S. More and more toxins around–analysis of cyanobacterial strains isolated from Lake Chao (Anhui Province, China). Toxicon 2010, 56, 1520–1524. [Google Scholar] [CrossRef]
- Li, H.; Gu, X.; Chen, H.; Mao, Z.; Shen, R.; Zeng, Q.; Ge, Y. Co-occurrence of multiple cyanotoxins and taste-and-odor compounds in the large eutrophic Lake Taihu, China: Dynamics, driving factors, and challenges for risk assessment. Environ. Pollut. 2022, 294, 118594. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, J.; Zhang, D.; Luo, L.; Liao, Q.; Yuan, L.; Wu, N. Seasonal and spatial variations of microcystins in Poyang Lake, the largest freshwater lake in China. Environ. Sci. Pollut. Res. 2018, 25, 6300–6307. [Google Scholar] [CrossRef]
- Stoyneva, M.P. Steady-state phytoplankton assemblages in shallow Bulgarian wetlands. Hydrobiologia 2003, 502, 169–176. [Google Scholar] [CrossRef]
- Ortega-Mayagoitia, E.; Rojo, C.; Rodrigo, M.A. Controlling factors of phytoplankton taxonomic structure in wetlands: An experimental approach. Hydrobiologia 2003, 502, 177–186. [Google Scholar] [CrossRef]
- Rengefors, K.; Gustafsson, S.; Ståhl-Delbanco, A. Factors regulating the recruitment of cyanobacterial and eukaryotic phytoplankton from littoral and profundal sediments. Aquat. Microb. Ecol. 2004, 36, 213–226. [Google Scholar] [CrossRef]
- Isvánovics, V.; Pettersson, K.; Rodrgio, M.A.; Pierson, D.; Padisák, J.; Colom, E. Gloeotrichia echinulata, a colonial cyanobacterium with a unique phosphorus uptake and life strategy. J. Plankton. Res. 1993, 15, 531–552. [Google Scholar] [CrossRef]
- Nixdorf, B.; Mischke, U.; Rücker, J. Phytoplankton assemblages and steady state in deep and shallow eutrophic lakes—An approach to differentiate the habitat properties of Oscillatoriales. Hydrobiologia 2003, 502, 111–121. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Toporowska, M. Blooms of toxin-producing Cyanobacteria—A real threat in small dam reservoirs at the beginning of their operation. Oceanol. Hydrobiol. Stud. 2011, 40, 30–37. [Google Scholar] [CrossRef]
- Grabowska, M. Cyanoprocaryota blooms in the polyhumic Siemianówka dam Reservoir in 1992–2003. Oceanol. Hydrobiol. Stud. 2005, 24, 73–85. [Google Scholar]
- Jöhnk, K.; Huisman, J.; Sharples, J.; Sommeijer, B.; Visser, P.M.; Strooms, J.M. Summer heatwaves promote blooms of harmful cyanobacteria. Glob. Chang. Biol. 2008, 14, 495–512. [Google Scholar] [CrossRef]
- Mankiewicz-Boczek, J.; Urbaniak, M.; Romanowska-Duda, Z.; Izydorczyk, K. Toxic cyanobacteria strains in lowland dam reservoir (Sulejów Res. central Poland): Amplification of MCY genes for detection and identification. Pol. J. Ecol. 2006, 54, 171–180. [Google Scholar]
- Toporowska, M.; Ferencz, B.; Dawidek, J. Impact of lake-catchment processes on phytoplankton community structure in temperate shallow lakes. Ecohydrology 2018, 11, e2017. [Google Scholar] [CrossRef]
- Sivonen, K.; Jones, G. Cyanobacterial toxins. In Toxic Cyanobacteria in Water: A Guide to Their Public Health Consequences, Monitoring and Management; Chorus, I., Bartram, J., Eds.; E and FN Spon: London, UK, 1999; pp. 41–111. [Google Scholar]
- Chen, L.; Giesy, J.P.; Adamovsky, O.; Svirčev, Z.; Meriluoto, J.; Codd, G.A.; Xie, P. Challenges of using blooms of Microcystis spp. in animal feeds: A comprehensive review of nutritional, toxicological and microbial health evaluation. Stoten 2021, 764, 142319. [Google Scholar] [CrossRef]
- Holland, A.; Kinnear, S. Interpreting the possible ecological role (s) of cyanotoxins: Compounds for competitive advantage and/or physiological aide? Mar. Drugs 2013, 11, 2239–2258. [Google Scholar] [CrossRef]
- Welker, M.; Döhren, H. Cyanobacterial peptides—nature’s own combinatorial biosynthesis. FEMS Microbiol. Rev. 2006, 30, 530–563. [Google Scholar] [CrossRef]
- Hu, C.; Rzymski, P. Programmed cell death-like and accompanying release of microcystin in freshwater bloom-forming cyanobacterium Microcystis: From identification to ecological relevance. Toxins 2019, 11, 706. [Google Scholar] [CrossRef]
- Affonso, A.G.; Barbosa, C.; Novo, E.M.L.M. Water quality changes in floodplain lakes due to the Amazon River flood pulse: Lago Grande de Curuaí (Pará). Braz. J. Biol. 2011, 71, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Pawlik-Skowrońska, B.; Toporowska, M.; Mazur-Marzec, H. Toxic oligopeptides in the cyanobacterium Planktothrix agardhii dominated blooms and their effects on duckweed (Lemnaceae) development. KMAE 2018, 419, 41. [Google Scholar] [CrossRef]
- Gilroy, D.J.; Kauffman, K.W.; Hall, R.A.; Huang, X.; Chu, F.S. Assessing potential health risks from microcystin toxins in blue-green algae dietary supplements. Environ. Health Perspect. 2000, 108, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.C.; Vasconcelos, V.M. Microcystin dynamics in aquatic organisms. J. Toxicol. Environ. Health Part B 2009, 12, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Foss, A.J.; Aubel, M.T.; Gallagher, B.; Mettee, N.; Miller, A.; Fogelson, S.B. Diagnosing microcystin intoxication of canines: Clinicopathological indications, pathological characteristics, and analytical detection in postmortem and antemortem samples. Toxins 2019, 11, 456. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedziałek, B. Dermatotoxins synthesized by blue-green algae (Cyanobacteria). Adv. Dermatol. Allergol. 2012, 29, 47–50. [Google Scholar]
- Kaloudis, T.; Triantis, T.M.; Hiskia, A. Taste and odour compounds produced by cyanobacteria. In Handbook of Cyanobacterial Monitoring and Cyanotoxin Analysis; Meriluoto, J., Spoof, L., Codd, G.A., Eds.; Wiley: Hoboken, NJ, USA, 2016; pp. 196–201. [Google Scholar]
- Miller, M.A.; Kudela, R.M.; Mekebri, A.; Crane, D.; Oates, S.C.; Tinker, M.T.; Jessup, D.A. Evidence for a novel marine harmful algal bloom: Cyanotoxin (microcystin) transfer from land to sea otters. PLoS ONE 2010, 5, 12576. [Google Scholar] [CrossRef] [PubMed]
- Schemel, L.E.; Sommer, T.R.; Müller-Solger, A.B.; Harrell, W.C. Hydrologic variability, water chemistry, and phytoplankton biomass in a large floodplain of the Sacramento River, CA, U.S.A. Hydrobiologia 2004, 513, 129–139. [Google Scholar] [CrossRef]
- Dawidek, J.; Ferencz, B. Water Balance of selected floodplain lake basins in the Middle Bug River valley. Hydrol. Earth Syst. Sci. 2014, 18, 1457–1465. [Google Scholar] [CrossRef]
- Dessie, M.; Verhoest, N.E.C.; Pauwels, V.R.N.; Adgo, E.; Deckers, J.; Poesen, J.; Nyssen, J. Water balance of a lake with floodplain buffering: Lake Tana, Blue Nile Basin, Ethiopia. J. Hydrol. 2015, 522, 174–186. [Google Scholar] [CrossRef]
- Brierley, G.J.; Fryirs, K.A.; Jain, V. Landscape connectivity: The geographic basis of geomorphic applications. Area 2006, 38, 165–174. [Google Scholar] [CrossRef]
- Schober, B.; Hauer, C.; Habersack, H. Floodplain losses and increasing flood risk in the context of recent historic land use changes and settlement developments: Austrian case studies. J. Flood Risk Manag. 2020, 13, e12610. [Google Scholar] [CrossRef]
- Shen, M.H.; Liu, X.Q. Assessing the effects of lateral hydrological connectivity alteration on freshwater ecosystems: A meta-analysis. Ecol. Indicat. 2021, 125, 107572. [Google Scholar] [CrossRef]
- Liu, X.Q.; Wang, H.Z. Effect of loss of lateral connectivity of fish functional diversity. Conserv. Biol. J. Soc. Conserv. Biol. 2018, 32, 1336–1345. [Google Scholar] [CrossRef]
- Serra-Llobet, A.; Jähnig, S.C.; Geist, J.; Kondolf, G.M.; Damm, C.; Scholz, M.; Lund, J.; Opperman, J.J.; Yarnell, S.M.; Pawley, A.; et al. Restoring Rivers and Floodplains for Habitat and Flood Risk Reduction: Experiences in Multi-Benefit Floodplain Management from California and Germany. Front. Environ. Sci. 2022, 9, 778568. [Google Scholar] [CrossRef]
- Lemke, M.J.; Hagy, H.M.; Dungey, K.; Casper, A.F.; Lemke, A.M.; VanMiddlesworth, T.D.; Angela Kent, A. Echoes of a flood pulse: Short-term effects of record flooding of the Illinois River on floodplain lakes under ecological restoration. Hydrobiologia 2017, 804, 151–175. [Google Scholar] [CrossRef]
- Kiedrzyńska, E.; Kiedrzyński, M.; Zalewski, M. Sustainable floodplain management for flood prevention and water quality improvement. Nat. Hazards 2015, 76, 955–977. [Google Scholar] [CrossRef]
- Thoms, M.C. Floodplain-river ecosystems: Lateral connections and the implications of human interference. Geomorphology 2003, 56, 335–349. [Google Scholar] [CrossRef]
- Zhang, Z.; Lu, W.X.; Zhao, Y.; Song, W.B. Development tendency analysis and evaluation of the water ecological carrying capacity in the Siping area of Jilin Province in China based on system dynamics and analytic hierarchy process. Ecol. Model. 2014, 275, 9–21. [Google Scholar] [CrossRef]
- Zalewski, M. Ecohydrology for implementation of the EU water framework directive. Proc. Inst. Civ. Eng. Water Manag. 2011, 164, 375–385. [Google Scholar] [CrossRef]
- Opperman, J.J.; Luster, R.; McKenney, B.A.; Roberts, M.; Meadows, A.W. Ecologically functional floodplains: Connectivity, flow regime, and scale. J. Am. Water Resour. Assoc. 2010, 46, 211–226. [Google Scholar] [CrossRef]
- Schilling, K.E.; Kult, K.; Wilke, K.; Streeter, M.; Vogelgesang, J. Nitrate reduction in a reconstructed floodplain oxbow fed by tile drainage. Ecol. Eng. 2017, 102, 98–107. [Google Scholar] [CrossRef]
- Boucek, R.E.; Rehage, J.S. Effects of an Episodic Drought on a Floodplain Subsidy Consumed by Mangrove River Fishes. Trans. Am. Fish. Soc. 2015, 83, 113–116. [Google Scholar]
- Bayley, P.B. Understanding large river-floodplain ecosystems. BioScience 1995, 45, 153–158. [Google Scholar] [CrossRef]
- Mallakpour, I.; Villarini, G. The changing nature of flooding across the central United States. Nat. Clim. Chang. 2015, 5, 250–254. [Google Scholar] [CrossRef]
- Klingelhöfer, D.; Müller, R.; Braun, M.; Brüggmann, D.; Groneberg, D.A. Climate change: Does international research fulfil global demands and necessities? Environ. Sci. Eur. 2020, 32, 137. [Google Scholar] [CrossRef]
- Tabari, H. Climate change impact on flood and extreme precipitation increases with water availability. Sci. Rep. 2020, 10, 13768. [Google Scholar] [CrossRef]
- Kundzewicz, Z.W.; Kanae, S.; Seneviratne, S.I.; Handmer, J.; Nicholls, N.; Peduzzi, P.; Mechler, R.; Bouwer, L.M.; Arnell, N.; Mach, K.; et al. Flood risk and climate change: Global and regional perspectives. Hydrol. Sci. J. 2014, 59, 1–28. [Google Scholar] [CrossRef]
- Mediero, L.; Santillán, D.; Garrote, L.; Granados, A. Detection and attribution of trends in magnitude, frequency and timing of floods in Spain. J. Hydrol. 2014, 517, 1072–1088. [Google Scholar] [CrossRef]
- Ward, R.D.; Friess, D.A.; Day, R.H.; MacKenzie, R.A. Impacts of climate change on mangrove ecosystems: A region by region overview. Ecosyst. Health Sustain. 2016, 2, 11879021. [Google Scholar] [CrossRef]
- Blöschl, G.; Hall, J.; Viglione, A.; Perdigão, R.A.P.; Parajka, J.; Merz, B.; Lun, D.; Arheimer, B.; Aronica, G.T.; Bilibashi, A.; et al. Changing climate both increases and decreases. Eur. River Floods Nat. 2019, 573, 108–111. [Google Scholar] [CrossRef]
- Salimi, S.; Berggren, M.; Scholzet, M. Response of the peatland carbon dioxide sink function to future climate change scenarios and water level management. Glob. Chang. Biol. 2021, 27, 5154–5168. [Google Scholar] [CrossRef] [PubMed]
- Budiyono, Y.; Aerts, J.C.J.H.; Tollenaar, D.; Ward, P.J. River flood risk in Jakarta under scenarios of future change. Nat. Hazard. Earth Sys. 2016, 16, 757–774. [Google Scholar] [CrossRef]
- Hamman, J.J.; Hamlet, A.F.; Lee, S.Y.; Fuller, R.; Grossman, E.E. Combined effects of projected sea level rise, storm surge, and peak river flows on water levels in the Skagit Floodplain. Northwest Sci. 2016, 90, 57–78. [Google Scholar] [CrossRef]
- Berghuijs, W.R.; Aalbers, E.E.; Larsen, J.R.; Trancoso, R.; Woods, R.A. Recent changes in extreme floods across multiple continents. Environ. Res. Lett. 2017, 12, 114035. [Google Scholar] [CrossRef]
- Wiel, K.; Wanders, N.; Selten, F.M.; Bierkens, M.F.P. Added value of large ensemble simulations for assessing extreme river discharge in a 2 °C warmer world. Geophys. Res. Lett. 2019, 46, 2093–2102. [Google Scholar] [CrossRef]
- Wang, A.; Melton, A.E.; Soltis, D.E.; Soltis, P.S. Potential distributional shifts in North America of allelopathic invasive plant species under climate change models. Plant Divers. 2022, 44, 11–19. [Google Scholar] [CrossRef]
- Amaral, J.H.F.; Melack, J.M.; Barbosa, P.M.; MacIntyre, S.; Kasper, D.; Cortés, A.; Silva, T.S.F.; de Sousa, R.N.; Forsberg, B.R. Carbon dioxide fluxes to the atmosphere from waters within flooded forests in the Amazon basin. J. Geophys. Res. Biogeosci. 2020, 125, e2019JG005293. [Google Scholar] [CrossRef]
- Barbosa, P.M.; Melack, J.M.; Amaral, J.H.F.; MacIntyre, S.; Kasper, D.; Cortes, A.C.; Forsberg, B.R. Dissolved CH4 concentrations and fluxes to the atmosphere from a tropical floodplain lake. Biogeochemistry 2020, 148, 129–151. [Google Scholar] [CrossRef]
- Zhang, W.; Zheng, X.; Gu, P.; Wang, N.; Zheng, Z. Distribution and risk assessment of phthalates in water and sediment of the Pearl River Delta. Environ. Sci. Pollut. Res. 2020, 27, 12550–12565. [Google Scholar] [CrossRef]
- Koç, G.; Petrow, T.; Thieken, A.H. Analysis of the Most Severe Flood Events in Turkey (1960–2014): Which Triggering Mechanisms and Aggravating Pathways Can be Identified? Water 2020, 12, 1562. [Google Scholar] [CrossRef]
- Li, Q.; Xiao, J.; Ou, T.; Han, M.S.; Wang, J.F.; Chen, J.G.; Li, Y.L.; Salmaso, N. Impact of water level fluctuations on the development of phytoplankton in a large subtropical reservoir: Implications for the management of cyanobacteria. Environ. Sci. Pollut. Res. 2017, 25, 1306–1318. [Google Scholar] [CrossRef] [PubMed]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Søndergaard, M.; Lauridsen, T.L.; Beklioǧlu, M.; Brucet, S.; Volta, P.; González-Bergonzoni, I.; et al. Climate Change Impacts on Lakes: An Integrated Ecological Perspective Based on a Multi-Faceted Approach, with Special Focus on Shallow Lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef]
- Giblin, S.M.; Gerrish, G.A. Environmental factors controlling phytoplankton dynamics in a large floodplain river with emphasis on cyanobacteria. River Res. Appl. 2020, 36, 1137–1150. [Google Scholar] [CrossRef]
- Gopfen, M. Relation Significance between Hydrological Residence Time and Phytoplankton Dynamics in Lake Kinneret (Israel) January. Open J. Ecol. 2019, 9, 179–492. [Google Scholar] [CrossRef]
- Sorribas, M.V.; Paiva, R.C.D.; Melack, J.; Jones, C.; Carvalho, L.; Bravo, J.M.; Costa, M.H. Projections of climate change effects on discharge and inundation in the Amazon River basin. Clim. Chang. 2016, 136, 555–570. [Google Scholar] [CrossRef]
- Mantzouki, E.; Visser, P.M.; Bormans, M.; Ibelings, B.W. Understanding the key ecological traits of cyanobacteria as a basis for their management and control in changing lakes. Aquat. Ecol. 2016, 50, 333–350. [Google Scholar] [CrossRef]
- Hodgkins, G.A.; Whitfield, P.H.; Burn, D.H.; Hannaford, J.; Renard, B.; Stahl, K.; Fleig, A.K.; Madsen, H.; Mediero, L.; Korhonen, J.; et al. Climate-driven variability in the occurrence of major floods across North America and Europe. J. Hydrol. 2017, 552, 704–717. [Google Scholar] [CrossRef]
- Roderick, T.P.; Wasko, C.; Sharma, A. Atmospheric moisture measurements explain increases in tropical rainfall extremes. Geophys. Res. Lett. 2019, 46, 1375–1382. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Booth, E.G.; Kucharik, C.J.; Lathrop, R.C. Extreme daily loads: Role in annual phosphorus input to a north temperate lake. Aquat. Sci. 2015, 77, 71–79. [Google Scholar] [CrossRef]
- Muhetaer, G.; Asaeda, T.; Jayasanka, S.M.D.H.; Baniya, M.B.; Abeynayaka, H.D.L.; Rashid, M.H.; Yan, H. Effects of Light Intensity and Exposure Period on the Growth and Stress Responses of Two Cyanobacteria Species: Pseudanabaena galeata and Microcystis aeruginosa. Water 2020, 12, 407. [Google Scholar] [CrossRef]
Figure 1.
Floodplain lake during extreme hydrological events (drought and flood).

Figure 2.
Common groups of phytoplankton in the limnophase and potamophase periods.

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ferencz, B.; Toporowska, M.; Dawidek, J. Role of Hydrology in Cyanobacterial Blooms in the Floodplain Lakes. Water 2023, 15, 1547. https://doi.org/10.3390/w15081547
AMA Style
Ferencz B, Toporowska M, Dawidek J. Role of Hydrology in Cyanobacterial Blooms in the Floodplain Lakes. Water. 2023; 15(8):1547. https://doi.org/10.3390/w15081547
Chicago/Turabian StyleFerencz, Beata, Magdalena Toporowska, and Jarosław Dawidek. 2023. "Role of Hydrology in Cyanobacterial Blooms in the Floodplain Lakes" Water 15, no. 8: 1547. https://doi.org/10.3390/w15081547
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.