
Influence of Some Environmental Factors on Summer Phytoplankton Community Structure in the Varna Bay, Black Sea (1992–2019)


Abstract
:1. Introduction
- A low-productive ecosystem before 1970 or the so-called “pristine” reference phase (1954–1970);
- A highly productive eutrophic system during the 1980s or a phase of intensive anthropogenic eutrophication and destruction of the Black Sea ecosystem (1970–1995);
- A transitional, relatively low productive system after the beginning of the 1990s or the so-called post-eutrophication phase in the development of the Black Sea ecosystem.
2. Materials and Methods
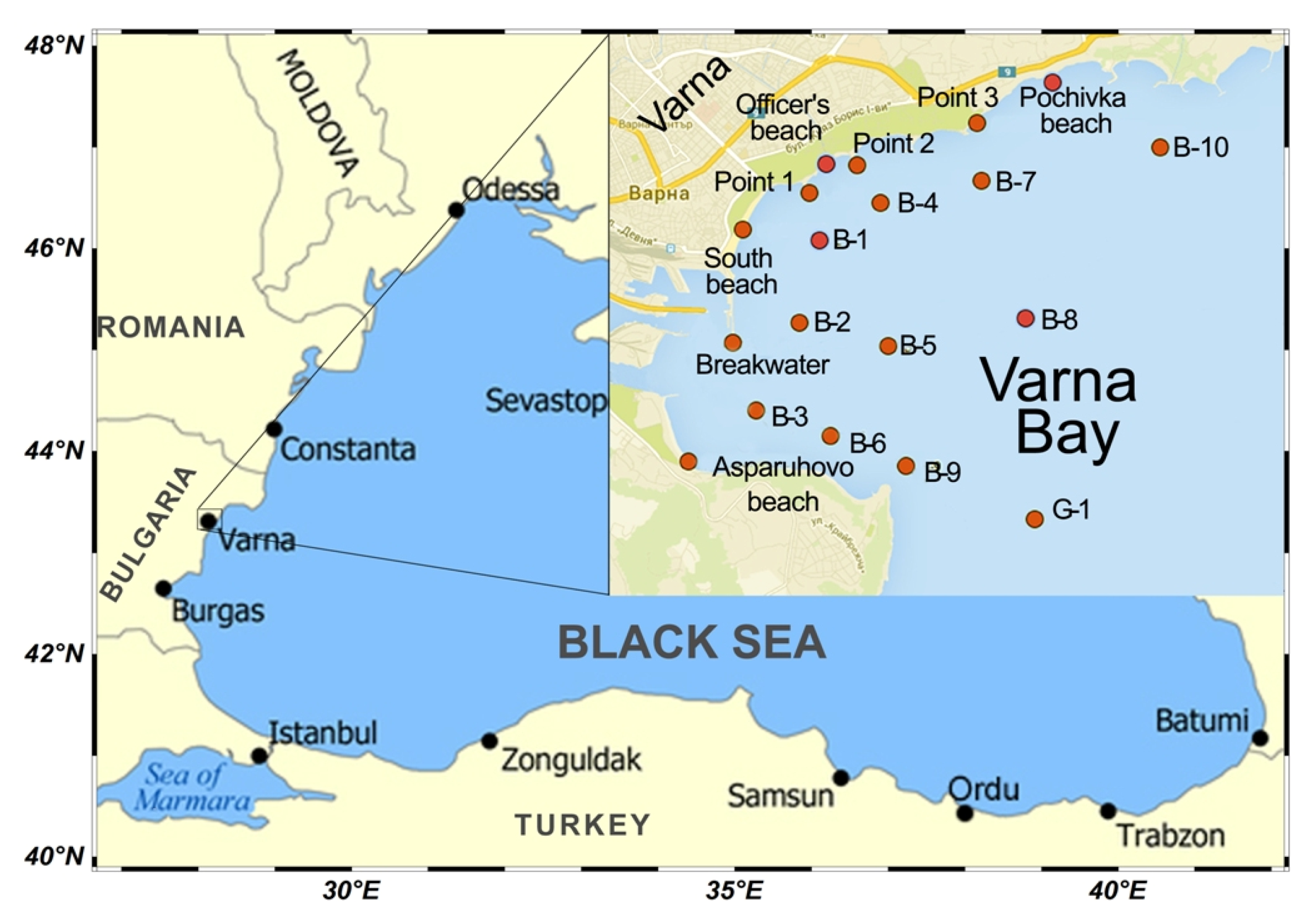
2.1. Study Area: Description of the Varna Bay
2.2. Field Sampling
2.3. Laboratory Treatment of the Phytoplankton Samples
2.4. Morphological Identification and Enumeration
2.5. Methods for Studying the Dependence of Phytoplankton on Sea Surface Temperature (SST) and Nutrients (TP), (DIN)
- The study was conducted only in Varna Bay as an area with uniform hydrologic conditions. This area was also selected because there is a long-term database for the phytoplankton (1631 phytoplankton samples for the whole period 1992–2019).
- To avoid the influence of light on the development of phytoplankton in deep water (from 1 m to 25 m), all samples were taken from the surface water layer (0–1 m).
- Samples from only the period of May–August were analyzed to minimize the effect of the seasonal dynamics on the abundance and biomass of the phytoplankton.
3. Results
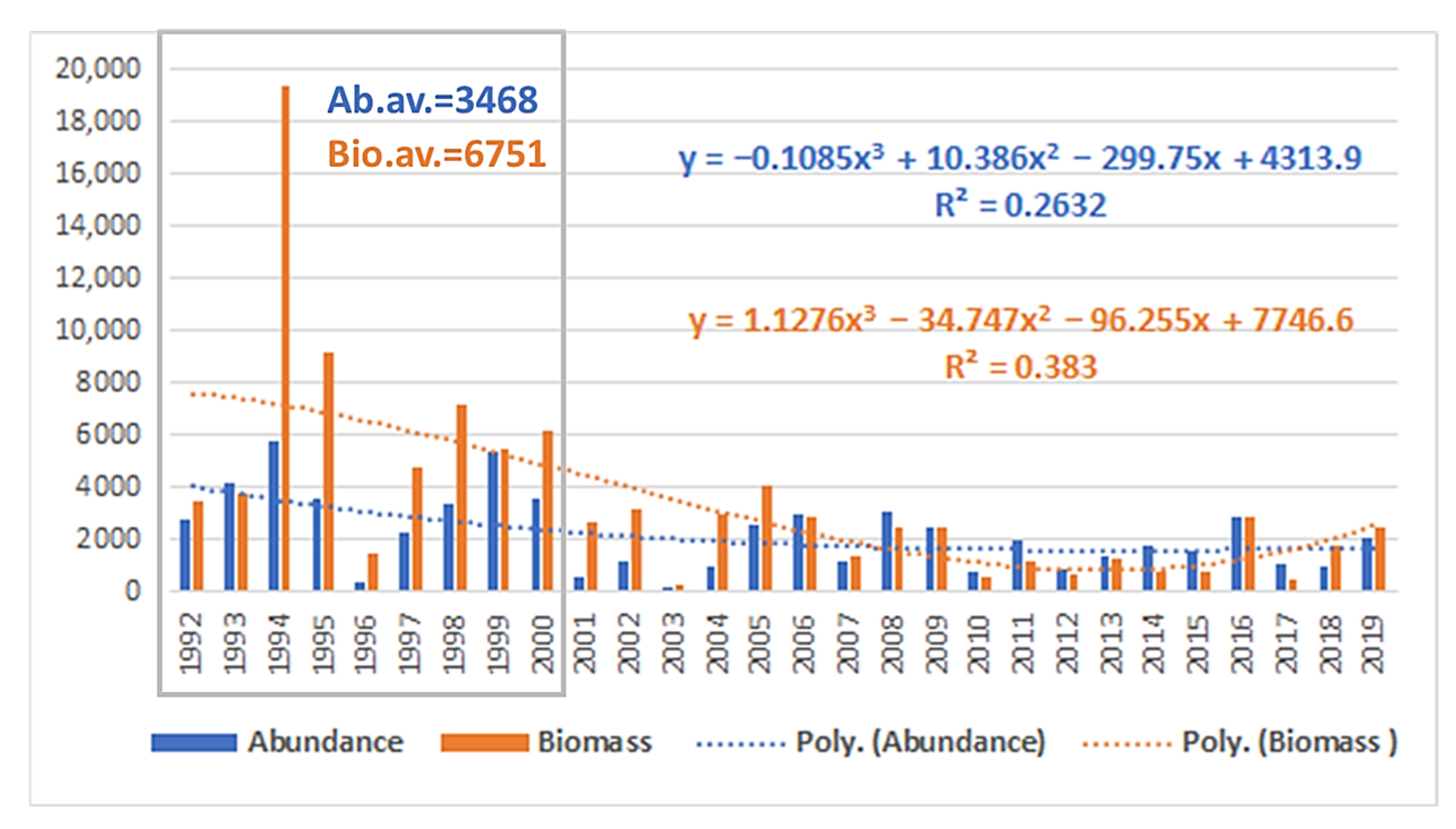
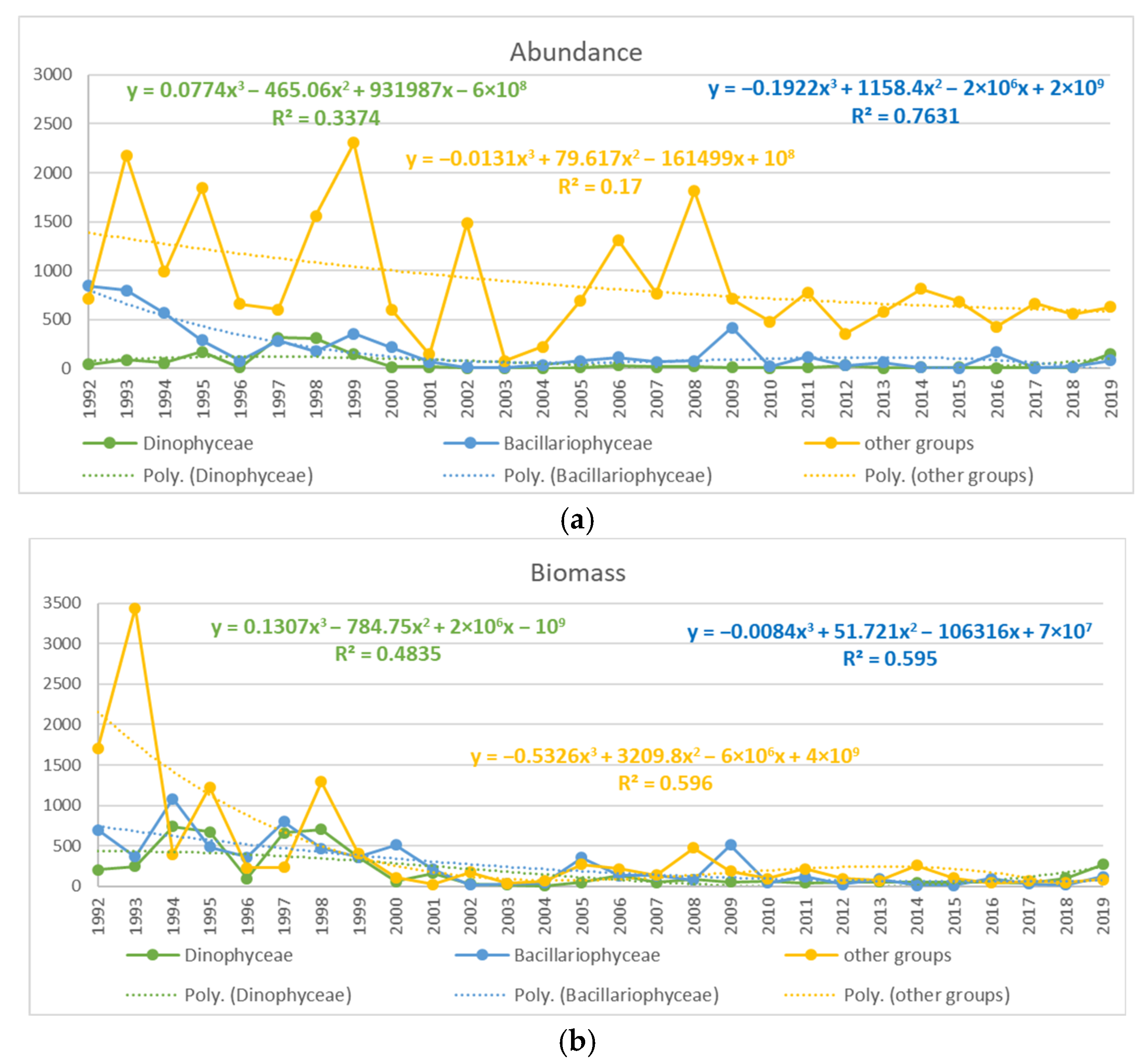
3.1. Tendencies in the Changes of the Phytoplankton Abundance and Biomass in the Varna Bay
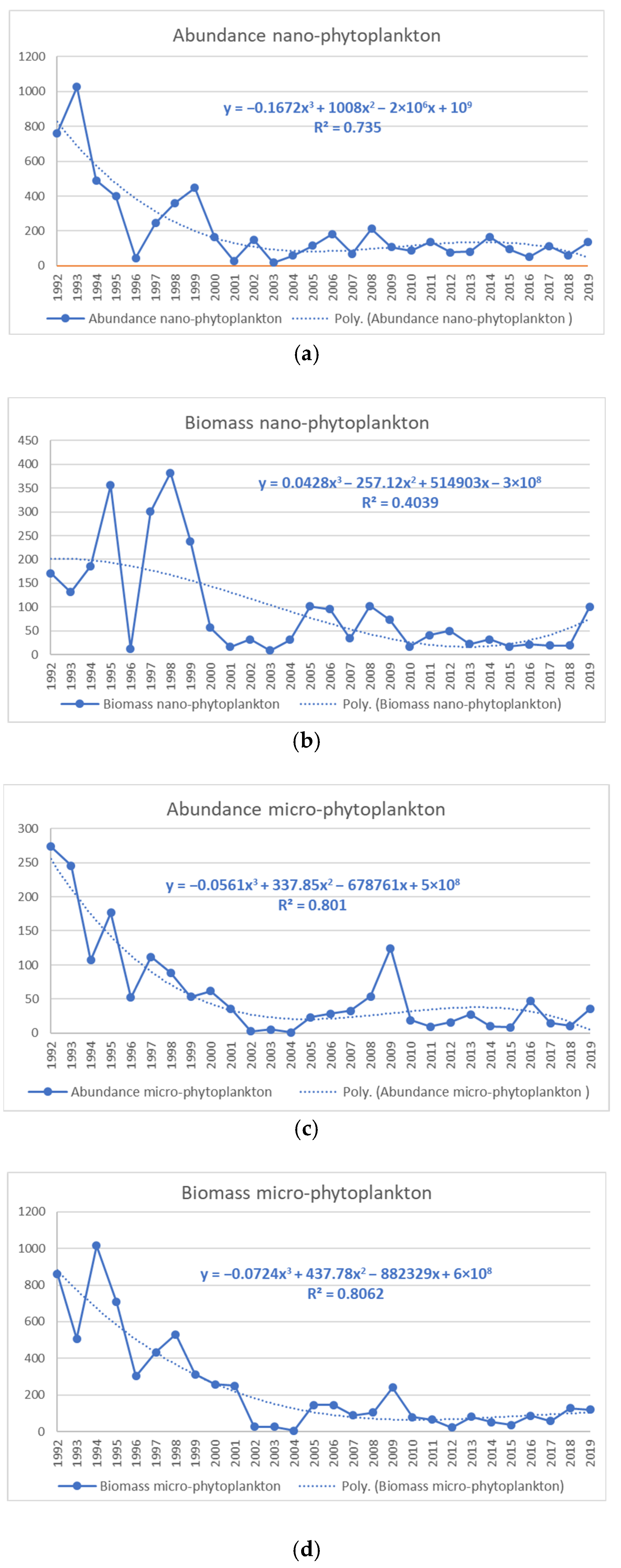
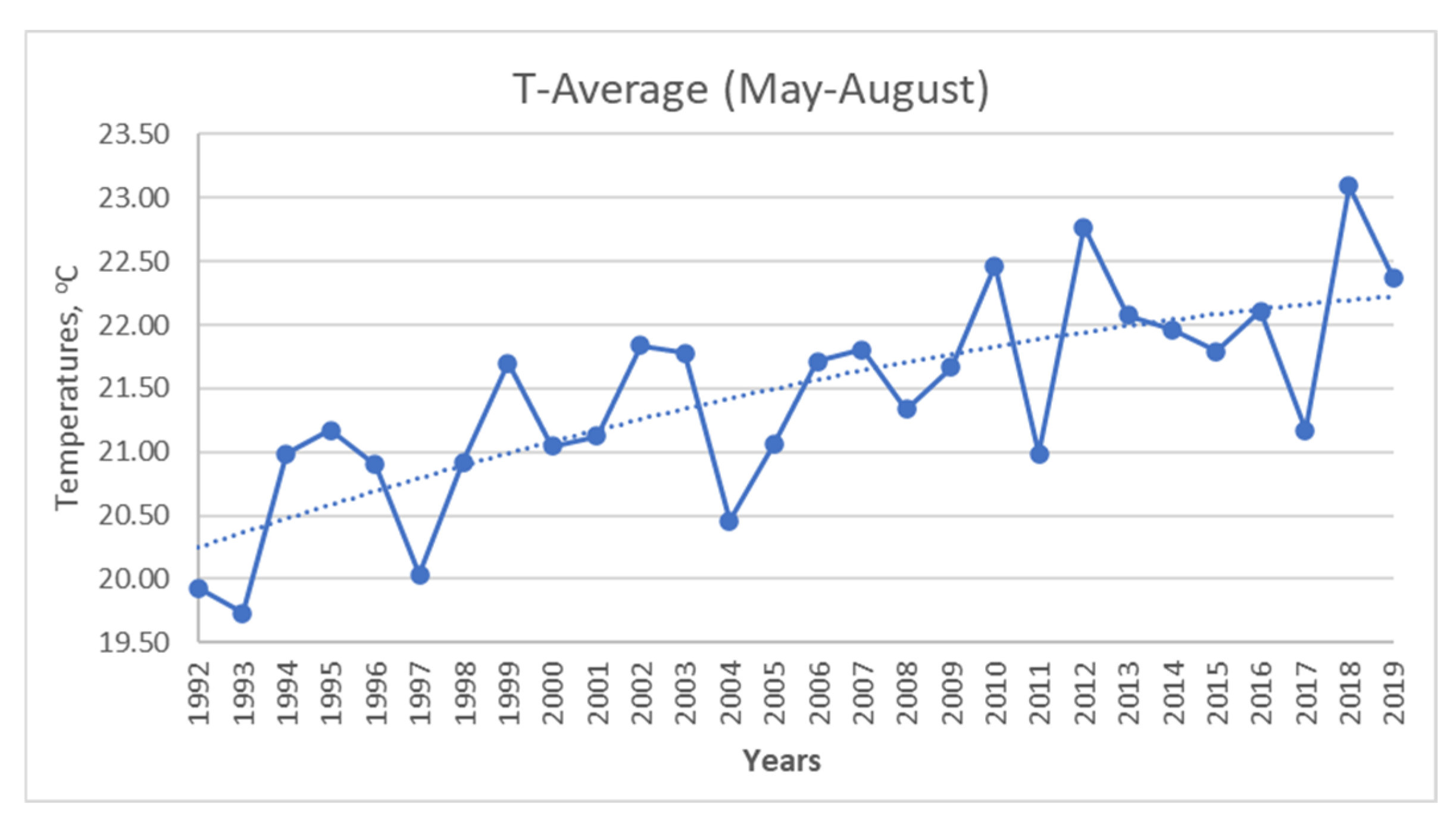
3.2. Relationship between the Sea Surface Temperature (SST), Abundance, and Biomass of Nano- and Micro-Phytoplankton
3.3. Relationship between SST, TP, DIN, and Abundance and Biomass of the Groups Dinophyceae, Bacillariophyceae, and Other
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Angelov, A. General Hydrobiology; Science and Art: Sofia, Bulgaria, 1971; pp. 1–326. (In Bulgarian) [Google Scholar]
- Eppley, R.W. Temperature and phytoplankton growth in the sea. Fish. Bull. 1972, 70, 1063–1085. Available online: https://spo.nmfs.noaa.gov/sites/default/files/pdf-content/1972/704/eppley.pdf (accessed on 6 September 2022).
- Edwards, M.; Richardson, A.J. Impact of climate change on marine pelagic phenology and trophic mismatch. Nature 2004, 430, 881–884. [Google Scholar] [CrossRef] [PubMed]
- Stenseth, N.C.; Mysterud, A.; Ottersen, G.; Hurrell, J.W.; Chan, K.-S.; Lima, M. Ecological effects of climate fluctuations. Science 2002, 297, 1292–1296. [Google Scholar] [CrossRef] [PubMed]
- Uzunov, J.; Kovachev, S. Hydrobiology; Pensoft: Sofia, Bulgaria; Moscow, Russia, 2002; pp. 1–342. ISBN 9789546425195. (In Bulgarian) [Google Scholar]
- Nicklisch, A.; Shatwell, T.; Köhler, J. Analysis and modelling of the interactive effects of temperature and light on phytoplankton growth and relevance for the spring bloom. J. Plankton Res. 2008, 30, 75–91. [Google Scholar] [CrossRef]
- Daskalov, G.M. Long-term changes in fish abundance and environmental indices in the Black Sea. Mar. Ecol. Prog. Ser. 2003, 255, 259–270. [Google Scholar] [CrossRef]
- Konovalov, S.K.; Murray, J.W. Variations in the chemistry of the Black Sea on a time scale of decades (1960–1995). J. Mar. Syst. 2001, 31, 217–243. [Google Scholar] [CrossRef]
- Lancelot, C.L.; Staneva, J.V.; Van Eeckhout, D.; Beckers, J.-M.; Stanev, E.V. Modelling the Danube-influenced North-western continental shelf of the Black Sea. Ecosystem response to changes in nutrient delivery by the Danube River after its damming in 1972. Estuar. Coast. Shelf Sci. 2002, 54, 473–499. [Google Scholar] [CrossRef]
- Mikaelyan, A.S. Long-term variability of phytoplankton communities in open Black Sea in relation to environmental changes. In Sensitivity to Change: Black Sea, Baltic Sea and North Sea NATO-ASI Series 2 Environment 27; Ozsoy, E., Mikaelyan, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1997; pp. 105–116. [Google Scholar] [CrossRef]
- Niermann, U.; Kideys, A.E.; Kovalev, A.V.; Melnikov, V.; Belokopytov, V. Fluctuations of pelagic species of the open Black Sea during 1980-1995 and possible teleconnections. In Environmental Degradation of the Black Sea: Challenges and Remedies NATO ASI Series B Environmental Security 56; Besiktepe, S., Unluata, U., Bologa, A., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999; pp. 147–174. [Google Scholar] [CrossRef]
- Oguz, T. Black Sea ecosystem response to climatic teleconnections. Oceanography 2005, 18, 122–133. [Google Scholar] [CrossRef]
- Yunev, O.A.; Vedernikov, V.I.; Basturk, O.; Yilmaz, A.; Kideys, A.E.; Moncheva, S.; Konovalov, S. Long-term variations of surface chlorophyll-a and primary production in the open Black Sea. Mar. Ecol. Prog. Ser. 2002, 230, 11–28. [Google Scholar] [CrossRef]
- Mee, L.D. The Black Sea in crisis: A need for concerted international action. Ambio 1992, 21, 278–286. [Google Scholar] [CrossRef]
- Oguz, T.; Velikova, V. Abrupt transition of the northwestern Black Sea shelf ecosystem from a eutrophic to an alternative pristine state. Mar. Ecol. Prog. Ser. 2010, 405, 231–242. Available online: http://www.jstor.org/stable/24873891 (accessed on 2 July 2022). [CrossRef]
- Sakalli, A.; Basusta, N. Sea surface temperature change in the Black Sea under climate change: A simulation of the sea surface temperature up to 2100. Int. J. Climatol. 2018, 38, 4687–4698. [Google Scholar] [CrossRef]
- Petrova, V.Y. Phytoplankton in the Black Sea off the Bulgarian coast for the period 1954–1957. Proc. Cent. Res. Inst. Fish Farming Fish. (CRIFFF)-Varna 1963, 3, 31–60. (In Bulgarian) [Google Scholar]
- Petrova, V.Y. Phytoplankton in the Black Sea off the Bulgarian coast during the period 1958–1960. Proc. Inst. Fish Resour. (IFR)-Varna 1964, 5, 5–32. [Google Scholar]
- Petrova-Karadzhova, V.Y. Dynamics of phytoplankton biomass in the Black Sea off the Bulgarian coast during the period 1964–1970. Proc. Inst. Oceanogr. Fish. (IOF)-Varna 1973, 12, 41–66. (In Bulgarian) [Google Scholar]
- Cociasu, A.; Popa, L. Significant changes in Danube nutrient loads and their impact on the Romanian Black Sea coastal waters. Cercet. Mar. 2004, 35, 25–37. Available online: http://www.rmri.ro/Home/Downloads/Publications.RecherchesMarines/2004/paper02.pdf (accessed on 23 January 2020).
- Cociasu, A.; Dorogan, L.; Humborg, C.; Popa, L. Long-term ecological changes in the Romanian coastal waters of the Black Sea. Mar. Pollut. Bull. 1996, 32, 32–38. [Google Scholar] [CrossRef]
- Humborg, C.; Ittekkot, V.; Cociasu, A.; Bodunge, B.V. Effect of Danube River dam on Black Sea biogeochemistry and ecosystem structure. Nature 1997, 386, 385–388. [Google Scholar] [CrossRef]
- Kamburska, L.; Moncheva, S.; Konsulov, A.; Krastev, A.; Prodanov, K. The invasion of Beroe ovata in the Black Sea a warming signal for ecosystem concern. Proc. Inst. Oceanol. 2003, 4, 111–123. Available online: http://www.io-bas.bg/downloads/Ocean_V4_15.pdf (accessed on 22 July 2022).
- Mavrodieva, R.; Michneva, V.; Hiebaum, G.; Venzislav, K. Phytoplankton community in the bay of Bourgas, abiotic ambience and trends in major shifts. In Proceedings of the Seventh International Conference on the Mediterranean Coastal EnvironmentsMEDCOAST, Kushadasi, Turkey, 25–29 October 2005; Ozhan, E., Ed.; pp. 557–568. Available online: https://www·medcoast.net/modul/index/menu/MEDCOAST-05/208 (accessed on 4 July 2022).
- Moncheva, S.; Krastev, A. Some Aspects of Phytoplankton Long-Term Alterations Off Bulgarian Black Sea Shelf. In Sensitivity to Change: Black Sea, Baltic Sea and North Sea NATO ASI Series; Özsoy, E., Mikaelyan, A., Eds.; Springer: Dordrecht, The Netherlands, 1997; Volume 27, pp. 79–94. [Google Scholar] [CrossRef]
- Moncheva, S.; Doncheva, V.; Kamburska, L. On the long-term response of harmful algal blooms to the evolution of eutrophication off the Bulgarian Black Sea coast: Are the recent changes a sign of recovery of the ecosystem-the uncertainties. In Proceedings of the IX International Conference on Harmful Algal Blooms, Hobart, Tasmania, 7–11 February 2000; UNESCO-IOC: Paris, France, 2001; pp. 177–182. Available online: https://wedocs.unep.org/handle/20.500.11822/845?show=full (accessed on 7 June 2022).
- Moncheva, S.P. On the thermohaline development of some mass phytoplankton species from the northern part of the Bulgarian coast. Oceanol. BAS Sofia 1991, 21, 14–20. (In Bulgarian) [Google Scholar]
- Moncheva, S.P. Ecology of Mass Phytoplankton from the Black Sea under Anthropogenic Eutrophication. Ph.D. Thesis, Institute of Botany Bulgarian Academy of Sciences, Sofia, Bulgaria, 1991. (In Bulgarian). [Google Scholar]
- Moncheva, S.P. Quantitative development of phytoplankton species as an indicator of eutrophication of the Bulgarian Black Sea coast. Oceanol. BAS Sofia 1991, 21, 21–30. (In Bulgarian) [Google Scholar]
- Moncheva, S. Phytoplankton blooms and succession under eutrophication. In Proceedings of the First National Conference on Problems of Biological Monitoring, Plovdiv, Bulgaria, 22–24 October 1987; pp. 133–138. [Google Scholar]
- Moncheva, S. Phytoplankton blooms in relation to some abiotic environmental factors. In Proceedings of the Scientific and Practical Conference—State of Research, Rational Utilization and Protection of Natural Resources of the Varna Region, Varna, Bulgaria, 30 October 1989; pp. 134–145. (In Bulgarian). [Google Scholar]
- Petrova, D.; Gerdzhikov, D. Ecological aspects on the distribution of phytoplankton in the Bulgarian part of the Black Sea. In Proceedings of the Seventh International Symposium Ecology-Sustainable Developmentof the Union of Scientists, Vratsa, Bulgaria, 23–25 October 2008; pp. 357–368. (In Bulgarian). [Google Scholar]
- Petrova, D.; Gerdzhikov, D. The phytoplankton community—An indicator of the ecological state along the Bulgarian Black sea coast. Cercet. Mar. 2008, 38, 53–72. [Google Scholar]
- Petrova, D.; Velikova, V.; Gerdzhikov, D. Recent state of phytoplankton community in the Varna bay. Bulg. J. Agric. Sci. 2006, 12, 247–260. Available online: http://www.agrojournal.org/12/02-11-06.pdf (accessed on 10 October 2022).
- Sukhanova, I.; Flint, M.; Hibaum, G.; Karamfilov, V.; Kopylov, A.; Matveeva, E.; Ratkova, T.; Sazhin, A. Exuviaella cordata red tide in Bulgarian coastal waters (May to June 1986). Mar. Biol. 1988, 99, 1–8. [Google Scholar] [CrossRef]
- Velikova, V.; Moncheva, S.; Petrova, D. Phytoplankton dynamics and Red Tides (1987–1997) in the Bulgarian Black Sea. Water Sci. Technol. 1999, 39, 27–36. [Google Scholar] [CrossRef]
- Vinogradova, L.A.; Velikova, V.N. Long-Term Dynamics of the NW Ecosystem of the Black Sea and Its Modeling; Sankt-Petersburg Publishers: Sankt-Petersburg, Russia, 1992; pp. 1–108. (In Russian) [Google Scholar]
- Zaitsev, Y.P.; Alexandrov, B.G.; Minicheva, G.G.; Adobovski, V.V.; Alexandrov, B.G.; Aniulova, L.V.; Babich, E.I.; Berlinskii, N.A.; Birkun, A.A.; Bogatova, U.I.; et al. The Northwest Part of the Black Sea: Biology and Ecology; Naukova Dumka: Kiev, Ukraine, 2006; pp. 1–701. ISBN 966-00-0159-2. (In Russian) [Google Scholar]
- Yunev, O.A.; Carstensen, J.; Moncheva, S.; Khaliulin, A.; Ertebjerg, G.; Nixon, S. Nutrient and phytoplankton trends on the western Black Sea shelf in response to cultural eutrophication and climate changes. Estuar. Coast. Shelf Sci. 2007, 74, 63–76. [Google Scholar] [CrossRef]
- Valkanov, A.; Marinov, H.; Danov, H.; Vladev, P.; Mishev, K.; Popov, V.; Petrova, V.; Valkanov, A.; Valchanov, G.; Dimitrov, D.; et al. Black Sea; Georgi Bakalov Publishers: Varna, Bulgaria, 1978; pp. 1–635. (In Bulgarian) [Google Scholar]
- Zaitsev, Y.; Mamaev, V. Biological Diversity in the Black Sea, A Study of Change and Decline; United Nations Publications: New York, NY, USA, 1997; pp. 1–208. ISBN 92-1-126042-6. [Google Scholar]
- Klisarova, D.; Gerdzhikov, D. Eutrophication of Varna Bay and coastal waters. Bulg. J. Agric. Sci. 2015, 21, 86–89. Available online: https://www.agrojournal.org/21/01s-13.pdf (accessed on 17 May 2022).
- Klisarova, D.; Gerdzhikov, D. Phytoplankton development off the Bulgarian coast (2015–2016). Proc. SU-Varna Mar. Sci. Ser. 2017, 1, 59–67. (In Bulgarian) [Google Scholar]
- Klisarova, D.; Gerdzhikov, D. Features of phytoplankton dynamics in Bulgarian coastal water areas. Proc. IFR-Varna 2017, 28, 63–72. (In Bulgarian) [Google Scholar]
- Petrova-Karadzhova, V.Y. Change of plankton flora in the Bulgarian Black Sea under the influence of eutrophication. Proc. IFR-Varna 1984, 21, 105–112. (In Bulgarian) [Google Scholar]
- Raymont, J.E.G. Plankton and Productivity in the Oceans—Volume 1 Phytoplankton, 2nd ed.; Pergamon Press Ltd.: Oxford, UK, 1980; pp. 1–496. ISBN 9781483297880. [Google Scholar]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG model of a seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. Available online: https://oceanrep.geomar.de/id/eprint/14151 (accessed on 22 January 2022). [CrossRef]
- Stelmakh, L.V.; Babich, I.I.; Tugrul, S.; Moncheva, S.; Stefanova, K. Phytoplankton Growth Rate and Zooplankton Grazing in the Western Part of the Black Sea in the Autumn Period. Oceanology 2009, 49, 83–92. [Google Scholar] [CrossRef]
- Shtereva, G.; Hristova, O.; Dzhurova, B.; Krastev, A. Hydrochemistry of Varna Bay (2000–2008). J. Environ. Prot. Ecol. 2012, 13, 1695–1702. [Google Scholar]
- Shtereva, G.P. Nutrients elements in the water in Bourgas Bay. Proceeding Union Sci.-Varna 2005, 1, 89–93. (In Bulgarian) [Google Scholar]
- Strokal, M.; Kroeze, C. Nitrogen and phosphorus inputs to the Black Sea in 1970–2050. Reg. Environ. Chang. 2013, 13, 179–192. [Google Scholar] [CrossRef]
- Velikova, V.; Petrova, D. Perennial dynamics of phytoplankton in Varna Bay during 1991–1997. Proc. Inst. Fish Resour. 1999, 25, 89–102. (In Bulgarian) [Google Scholar]
- Velikova, V.; Petrova, D.; Mihneva, V.; Dineva, S.; Ouzounova, S. Recent state of the Bulgarian Black Sea—Signs of improvement of the ecosystem. In Proceedings of the Fifth International Conference on the Mediterranean Coastal Environment MEDCOAST, Hammamet, Tunisia, 23–27 October 2001; Ozhan, E., Ed.; pp. 893–905. [Google Scholar]
- Velikova, V.; Cociasu, A.; Popa, L.; Boicenco, L.; Petrova, D. Phytoplankton community and hydrochemical characteristics of the Western Black Sea. Water Sci. Technol. 2005, 51, 9–18. [Google Scholar] [CrossRef]
- Petrova, D.; Ivanova, P.; Velikova, V.; Mihneva, V. The distribution of the plankton of the Black sea. In Proceedings of the Seventh International Conference on the Mediterranean Coastal Environment, MEDCOAST, Kushadasi, Turkey, 25–29 October 2005; Ozhan, E., Ed.; pp. 547–556. [Google Scholar]
- Krutov, A.; Alexandrov, B.; Korshenko, A.; Lavrova, O.; Adobovskiy, V.; Lipchenko, A.; Amalina, A.; Luybimtsev, A.; Arashkevich, E.; Machitadze, N.; et al. State of the Environment of the Black Sea (2009-2014/5); Publications of the Commission on the Protection of the Black Sea Against Pollution (BSC): Istanbul, Turkey, 2019; pp. 1–811. ISBN 978-605-84837-0-5. [Google Scholar]
- Todorova, V.; Monchev, S.; Dzhurova, B.; Doncheva, V.; Peichev, V.; Slabakova, V.; Raykov, V.; Karamfilov, V.; Shtereva, G.P.; Berov, D.; et al. Initial Assessment of the state of the Marine Environment, According to Article 8 of the MSFD and the NOEMS; Publications of the Institute of Oceanology BAS: Varna, Bulgaria, 2013; pp. 1–487. (In Bulgarian) [Google Scholar]
- Yunev, O.; Velikova, V.; Carstensen, J. Reconstructing the trophic history of the Black Sea shelf. Cont. Shelf Res. 2017, 150, 1–9. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Chasovnikov, V.K.; Kubryakov, A.A.; Stanichny, S.V. Phenology and drivers of the winter-spring phytoplankton bloom in the open Black Sea: The application of Sverdrup’s hypothesis and its refinements. Prog. Oceanogr. 2017, 151, 163–176. [Google Scholar] [CrossRef]
- Mikaelyan, A.S.; Pautova, L.A.; Fedorov, A.V. Seasonal evolution of deep phytoplankton assemblages in the Black Sea. J. Sea Res. 2021, 178, 102125. [Google Scholar] [CrossRef]
- Oguz, T.; Gilbert, D. Abrupt transitions of the top-down controlled Black Sea pelagic ecosystem during 1960–2000: Evidence for regime-shifts under strong fishery exploitation and nutrient enrichment modulated by climate-induced variations. Deep. Sea Res. Part I Oceanogr. Res. Pap. 2007, 54, 220–242. [Google Scholar] [CrossRef]
- Petrova, D.; Gerdzhikov, D. Seasonal and inter annual variations of phytoplankton communities in Varna bay—Anthropogenic impact or climate changes. In Proceedings of the IV Balkan Conference BALNIMALCON, Stara Zagora, Bulgaria, 14–16 May 2009; pp. 357–362. [Google Scholar]
- Petrova, D.; Gerdzhikov, D. Development of the traditionally blooming phytoplankton species Prorocentrum cordatum Dodge, 1975 off the Bulgarian coast (2008–2010). Proc. Union Sci.-Varna 2013, 1, 12–17. (In Bulgarian) [Google Scholar]
- Petrova, D.; Gerdzhikov, D. Phytoplankton growth and chlorophyll-a content in the surface layer of the Bulgarian Black Sea coastal waters in 2011. Agric. Sci. Technol. 2013, 5, 99–104. [Google Scholar]
- Oguz, T.; Abaza, V.; Akatov, V.; Aktan, Y.; Arashkevich, E.; Birkun, A.; Boicenco, L.; Chikina, M.V.; Cociasu, A.; Daskalov, G.M.; et al. State of the Environment of the Black Sea (2001-2006/7); Black Sea Commission Publications: Istanbul, Turkey, 2008; pp. 1–421. ISBN 978-9944-245-33-3. [Google Scholar]
- Climate Change 2022: Impacts, Adaptation and Vulnerability. Available online: https://www.ipcc.ch/report/ar6/wg2/ (accessed on 21 June 2022).
- Allan, R.P.; Arias, P.A.; Berger, S.; Canadell, J.G.; Cassou, C.; Chen, D.; Cherchi, A.; Connors, S.L.; Coppola, E.; Cruz, F.A.; et al. Summary for policymakers. In Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Cambridge University Press: Cambridge, UK, 2021; pp. 3–32. [Google Scholar] [CrossRef]
- WMO State of the Global Climate. Available online: https://library.wmo.int/doc_num.php?explnum_id=11178 (accessed on 21 June 2022).
- Mulet, S.; Buongiorno Nardelli, B.; Good, S.; Pisano, A.; Greiner, E.; Monier, M.; Autret, E.; Axell, L.; Boberg, F.; Ciliberti, S.; et al. Ocean temperature and salinity. J. Oper. Oceanogr. 2018, 11, s5–s13. [Google Scholar] [CrossRef]
- Black Sea Time Series and Trend (1997–2020) of Satellite Chlorophyll. Available online: https://marine.copernicus.eu/access-data/ocean-monitoring-indicators/black-sea-chlorophyll-time-series-and-trend-observations (accessed on 26 April 2022).
- Marañon, E.; Holligan, P.M.; Barciela, R.; Gonzalez, N.; Mourino, B.; Pazo, M.J.; Varela, M. Patterns of phytoplankton size structure and productivity in contrasting open-ocean environments. Mar. Ecol. Progr. Ser. 2001, 216, 43–56. [Google Scholar] [CrossRef]
- Bouman, H.A.; Platt, T.; Sathyendranath, S.; Li, W.K.W.; Stuart, V.; Fuentes-Yaco, C.; Maass, H.; Horne, E.; Ulloa, O.; Lutz, V.; et al. Temperature as indicator of optical properties and community structure of marine phytoplankton: Implications for remote sensing. Mar. Ecol. Progr. Ser. 2003, 258, 19–30. [Google Scholar] [CrossRef]
- Robinson, A.; Bouman, H.A.; Tilstone, G.H.; Sathyendranath, S. Size Class Dependent Relationships between Temperature and Phytoplankton Photosynthesis-Irradiance Parameters in the Atlantic Ocean. Front. Mar. Sci. 2018, 4, 435. [Google Scholar] [CrossRef]
- Peter, K.H.; Sommer, U. Phytoplankton Cell Size Reduction in Response to Warming Mediated by Nutrient Limitation. PLoS ONE 2013, 8, e71528. [Google Scholar] [CrossRef] [PubMed]
- Li, W.K.W.; Harrison, W.G.; Head, E.J.H. Coherent assembly of phytoplankton communities in diverse temperate ocean ecosystems. Proc. R. Soc. B 2006, 273, 1953–1960. [Google Scholar] [CrossRef] [PubMed]
- Bopp, L.; Aumont, O.; Cadule, P.; Alvain, S.; Gehlen, M. Response of diatoms distribution to global warming and potential implications: A global model study. Geophys. Res. Lett. 2005, 32, 1–4. [Google Scholar] [CrossRef]
- BSCommision. Available online: http://www.blacksea-commission.org/The%20Black%20Sea/Geography/ (accessed on 15 March 2023).
- Vessel Traffic Service Authority. Available online: https://vtmis.bg/en/structure/vessel-traffic-services-authority-varna/general-information/vts-varna-geographical-information (accessed on 15 March 2023).
- PROVACAT Report of project Provadijska Catchment. Available online: http://www.iia.cnr.it/big_file/EUROCAT/minutes/ProvoCat.pdf (accessed on 15 September 2007).
- Morozova-Vodyanitskaya, N.V. Phytoplankton of the Black Sea, part II. Proc. Sevastopol Biol. Stn. 1954, 8, 11–99. (In Russian) [Google Scholar]
- Moncheva, S.; Parr, B. Manual for Phytoplankton Sampling and Analysis in the Black Sea; Black Sea Commission: Istanbul, Turkey, 2010; pp. 1–68. [Google Scholar]
- Edler, L. Recommendations for Marine Biological Studies in the Baltic Sea Phytoplankton and Chlorophyll; Baltic Marine Biologists: Uppsala, Sweden, 1979; pp. 5–38. [Google Scholar]
- Olenina, I.; Hajdu, S.; Edler, L.; Andersson, A.; Wasmund, N.; Busch, S.; Göbel, J.; Gromisz, S.; Huseby, S.; Huttunen, M.; et al. Biovolumes and size-classes of phytoplankton in the Baltic Sea. HELCOM Balt.Sea Environ. Proc. 2006, 106, 1–144. [Google Scholar]
- Klisarova, D. Phytomar 2.0—Software; Institute of Fish Resources: Varna, Bulgaria, 2008. [Google Scholar]
- WoRMS Editorial Board. World Register of Marine Species. Available online: https://www·marinespecies.org (accessed on 18 November 2021). [CrossRef]
- Guiry, M.D.; Guiry, G.M. AlgaeBase. World-Wide Electronic Publication, National University of Ireland, Galway. Available online: http://www.algaebase.org (accessed on 12 November 2021).
- Zeitzschel, B. Why study phytoplankton. In Phytoplankton Manual; UNESCO: Paris, France, 1978; pp. 1–5. [Google Scholar]
- Banzon, V.; Smith, T.M.; Chin, T.M.; Liu, C.; Hankins, W. A long-term record of blended satellite and in situ sea-surface temperature for climate monitoring, modeling and environmental studies. Earth Syst. Sci. Data 2016, 8, 165–176. [Google Scholar] [CrossRef]
- Pletterbauer, F.; Melcher, A.; Graf, W. Climate Change Impacts in Riverine Ecosystems. In Riverine Ecosystem Management. Aquatic Ecology Series Volume 8; Schmutz, S., Sendzimir, J., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–571. [Google Scholar] [CrossRef]
- Avsar, N.B.; Jin, S.; Kutoglu, S.H. Interannual Variations of Sea Surface Temperature in the Black Sea. In Proceedings of the IGARSS 2018—2018 IEEE International Geoscience and Remote Sensing Symposium, Valencia, Spain, 22–27 July 2018; pp. 5617–5620. [Google Scholar] [CrossRef]
- Shtereva, G.; Velikova, V.; Doncheva, V. Human impact on marine water nutrients enrichment. J. Environ. Prot. Ecol. 2015, 16, 40–48. [Google Scholar]
- Stelmakh, L.; Kovrigina, N.; Gorbunova, T. Phytoplankton Seasonal Dynamics under Conditions of Climate Change and Anthropogenic Pollution in the Western Coastal Waters of the Black Sea (Sevastopol Region). J. Mar. Sci. Eng. 2023, 11, 569. [Google Scholar] [CrossRef]
- Maranon, E.; Cermeno, P.; Latasa, M.; Tadonleke, R.D. Temperature, resources, and phytoplankton size structure in the ocean. Limnol. Oceanogr. 2012, 57, 1266–1278. [Google Scholar] [CrossRef]
- Marinov, I.; Doney, S.C.; Lima, I.D. Response of ocean phytoplankton community structure to climate change over the 21st century: Partitioning the effects of nutrients, temperature and light. Biogeosciences 2010, 7, 3941–3959. [Google Scholar] [CrossRef]
- Moncheva, S.; Slabakova, V.; Doncheva, V. Dominant habitat types in the water column—Phytoplankton. In Initial Assessment of the State of the Marine Environment, According to Article 8 of the WFD and the NOEMS; Publication of Institute of Oceanology: Varna, Bulgaria, 2013; pp. 1–487. (In Bulgarian) [Google Scholar]
- Kroiss, H.; Zessner, M.; Lampert, C. Nutrient Management in the Danube Basin and Its Impact on the Black Sea, daNUbs; Final Report EVK1-CT-2000-00051, Project Duration: 01.02.2001–31.1.2005; Vienna University of Technology: Viena, Austria, 2005; pp. 1–69. Available online: https://cordis.europa.eu/project/id/EVK1-CT-2000-00051 (accessed on 14 September 2007).
- Klisarova, D.; Gerdzhikov, D.; Kostadinova, G.; Petkov, G. Investigation of phytoplankton in the Varna bay (2005–2014). In Proceedings of the Twelfth International Conference on the Mediterranean Coastal Environment MEDCOAST, Varna, Bulgaria, 6–10 October 2015; Özhan, E., Ed.; pp. 299–308. [Google Scholar]
- Moncheva, S. Phytoplankton Shifts in the Black Sea-Driving Forces and Possible Implication for Reference Conditions. In Proceedings of the Joint Workshop on Streamlining the Process of Producing Regional Assessments on Eutrophication for Pan-European Purposes. EC JRC-BSCOM-HELCOM, Istanbul, Turkey, 26–28 October 2005. [Google Scholar]
- Anderson, S.I.; Franze, G.; Kling, J.D.; Wilburn, P.; Kremer, C.T.; Menden-Deuer, S.; Litchman, E.; Hutchins, D.A.; Rynearson, T.A. The interactive effects of temperature and nutrients on a spring phytoplankton community. Limnol. Oceanogr. 2022, 67, 634–645. [Google Scholar] [CrossRef]
- Brandenburg, K.M.; Velthuis, M.; Van de Waal, D.B. Meta-analysis reveals enhanced growth of marine harmful algae from temperate regions with warming and elevated CO2 levels. Glob. Chang. Biol. 2019, 25, 2607–2618. [Google Scholar] [CrossRef] [PubMed]
- Ray, S.; Berec, L.; Straskraba, M.; Jørgensen, S.E. Optimization of exergy and implications of body sizes of phytoplankton and zooplankton in an aquatic ecosystem model. Ecol. Model. 2001, 140, 219–234. [Google Scholar] [CrossRef]
- Gittings, J.A.; Raitsos, D.E.; Brewin, R.J.W.; Hoteit, I. Links between Phenology of Large Phytoplankton and Fisheries in the Northern and Central Red Sea. Remote Sens. 2021, 13, 231. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spearman’s Coefficient | Nano-Phytoplankton | Micro-Phytoplankton | ||
|---|---|---|---|---|
| Abundance (×106 Cells·m−3) | Biomass (mg·m−3) | Abundance (×106 Cells·m−3) | Biomass (mg·m−3) | |
| SST (May-August) | −0.431 * | −0.405 * | −0.471 * | −0.553 * |
| Regression Equation | Coefficient of Determination R2 | Significance | |
|---|---|---|---|
| Nano-phytoplankton (n) | |||
| Abundance (×106 cells·m−3) | (n) = 50,183.57 − 4511.78 × SST + 101.58 × SST2 | 0.509 | 0.0001 |
| Biomass (mg·m−3) | (n) = 2331.002 − 153.55 × SST + 2.293 × SST2 | 0.182 | 0.081 |
| Micro-phytoplankton (m) | |||
| Abundance (×106 cells·m−3) | (m) = 14,530.32 − 1301.47 × SST + 29.18 × SST2 | 0.530 | 0.0001 |
| Biomass (mg·m−3) | (m) = 22,724.72 − 1924.57 × SST + 40.79 × SST2 | 0.332 | 0.007 |
| Spearman’s Coefficient | Dinophyceae | Bacillariophyceae | Other | |||
|---|---|---|---|---|---|---|
| Abundance (×106 Cells·m−3) | Biomass (mg·m−3) | Abundance (×106 Cells·m−3) | Biomass (mg·m−3) | Abundance (×106 Cells·m−3) | Biomass (mg·m−3) | |
| SST (May–August) | −0.322 | −0.287 | −0.563 * | −0.682 * | −0.282 | −0.534 * |
| TP (mg·L−1) | 0.559 * | 0.540 * | 0.600 * | 0.616 * | 0.432 * | 0.637 * |
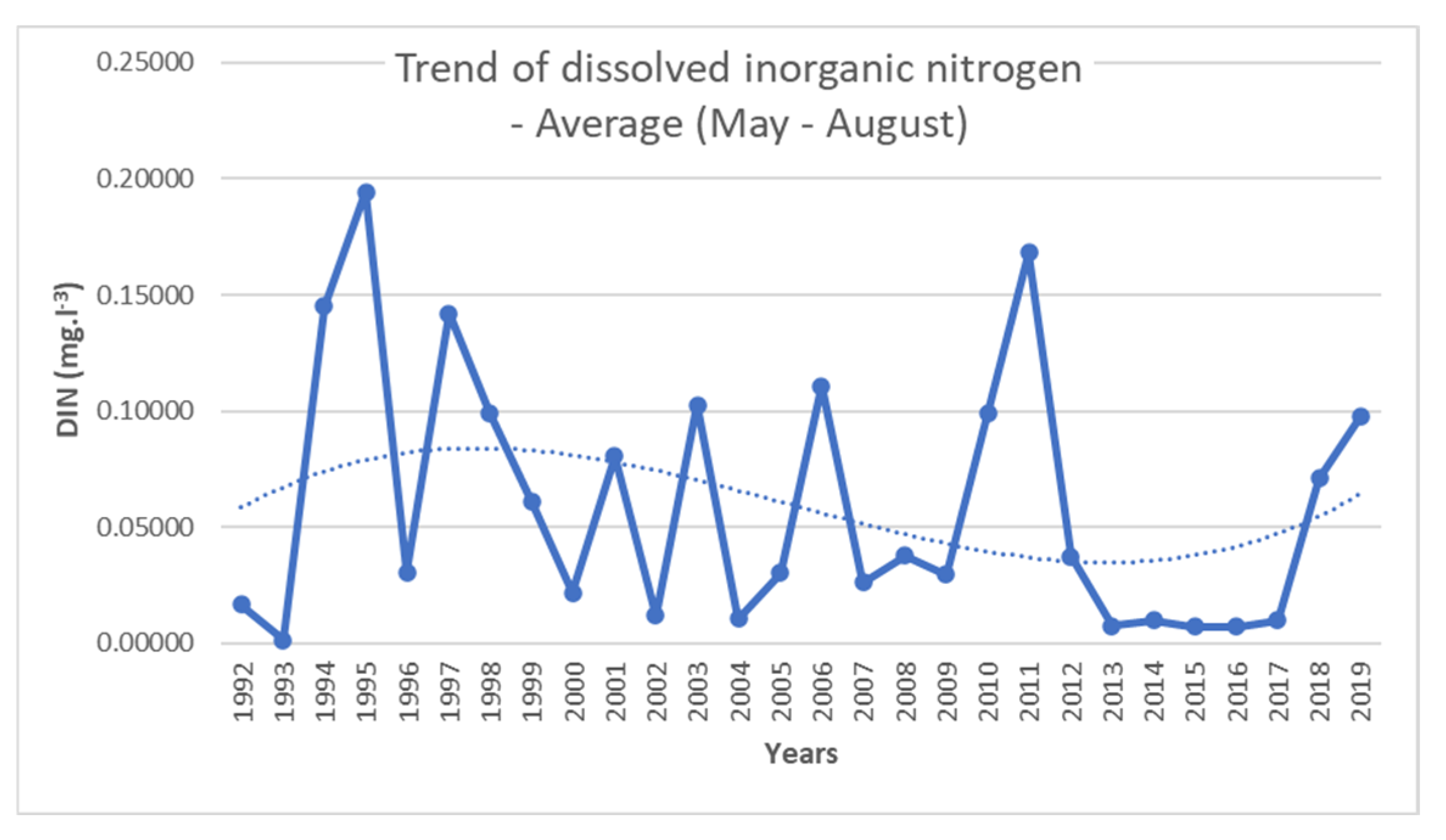
| DIN (mg·L−1) | 0.415 * | 0.450 * | 0.265 | 0.337 | 0.083 | 0.154 |
| Regression Statistics | Abundance | Biomass |
|---|---|---|
| Dinophyceae | ||
| R2 | 0.125 | 0.136 |
| Significance | 0.189 | 0.161 |
| Regression equations | AD = 6997.026 − 616.024 × SST + 13.61 × SST2 | BD = 6585.15 − 496.66 × SST + 9.223 × SST2 |
| Bacillariophyceae | ||
| R2 | 0.495 | 0.356 |
| Significance | 0.0001 | 0.004 |
| Regression equations | AB = 44,305.918 − 3963.06 × SST + 88.72 × SST2 | BB = 79,85.64 − 520.07 × SST + 7.431 × SST2 |
| Other | ||
| R2 | 0.063 | 0.514 |
| Significance | 0.441 | 0.0001 |
| Regression equations | AO = −17,043.57 + 1857.56 × SST − 47.58 × SST2 | BO = 167,448.308 − 15,137.41 × SST + 342.18 × SST2 |
| Regression Statistics | Abundance | Biomass |
|---|---|---|
| Dinophyceae | ||
| R2 | 0.13 | 0.293 |
| Significance | 0.175 | 0.103 |
| Regression equations | AD = −14.704 + 3075.81 × TP − 14,605.36 × TP2 | BD = 41.85 + 4528.87 × TP − 6260.64 × TP2 |
| Bacillariophyceae | ||
| R2 | 0.395 | 0.387 |
| Significance | 0.002 | 0.002 |
| Regression equations | AB = −56.12 + 8830.05 × TP − 28,102.21 × TP2 | BB = 91.63 + 4848.21 × TP − 614.57 × TP2 |
| Other | ||
| R2 | 0.168 | 0.413 |
| Significance | 0.101 | 0.001 |
| Regression equations | AO = 359.17 + 21,831.08 × TP − 94,401.01 × TP2 | BO = −588.40 + 42376.15 × TP − 185,415.56 × TP2 |
| Regression Statistics | Abundance | Biomass |
|---|---|---|
| Dinophyceae | ||
| R2 | 0.237 | 0.408 |
| Significance | 0.034 | 0.001 |
| Regression equations | AD = 5.28 + 1014.33 × DIN − 1551.36 × DIN2 | BD = 52.26 − 1424.08 × DIN + 7095.5 × DIN2 |
| Bacillariophyceae | ||
| R2 | 0.052 | 0.133 |
| Significance | 0.512 | 0.168 |
| Regression equations | AB = 230.37 − 2807.4 × DIN + 17,931.8 × DIN2 | BB = 184.98 − 211.59 × DIN + 9292.15 × DIN2 |
| Other groups | ||
| R2 | 0.057 | 0.100 |
| Significance | 0.482 | 0.269 |
| Regression equations | AO = 960.06 − 6104.9 × DIN + 43,687.8 × DIN2 | BO = 718.71 − 13,704.5 × DIN + 79,298.7 × DIN2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klisarova, D.; Gerdzhikov, D.; Nikolova, N.; Gera, M.; Veleva, P. Influence of Some Environmental Factors on Summer Phytoplankton Community Structure in the Varna Bay, Black Sea (1992–2019). Water 2023, 15, 1677. https://doi.org/10.3390/w15091677
Klisarova D, Gerdzhikov D, Nikolova N, Gera M, Veleva P. Influence of Some Environmental Factors on Summer Phytoplankton Community Structure in the Varna Bay, Black Sea (1992–2019). Water. 2023; 15(9):1677. https://doi.org/10.3390/w15091677
Chicago/Turabian StyleKlisarova, Daniela, Dimitar Gerdzhikov, Nina Nikolova, Martin Gera, and Petya Veleva. 2023. "Influence of Some Environmental Factors on Summer Phytoplankton Community Structure in the Varna Bay, Black Sea (1992–2019)" Water 15, no. 9: 1677. https://doi.org/10.3390/w15091677