
Understanding the Role of Macroalgal Complexity and Allelochemicals Production in Invasive and Non-Invasive Macroalgae in the North-Western Adriatic Sea: Effect on the Associated Communities
 and
and
Abstract
:1. Introduction
2. Materials and Methods
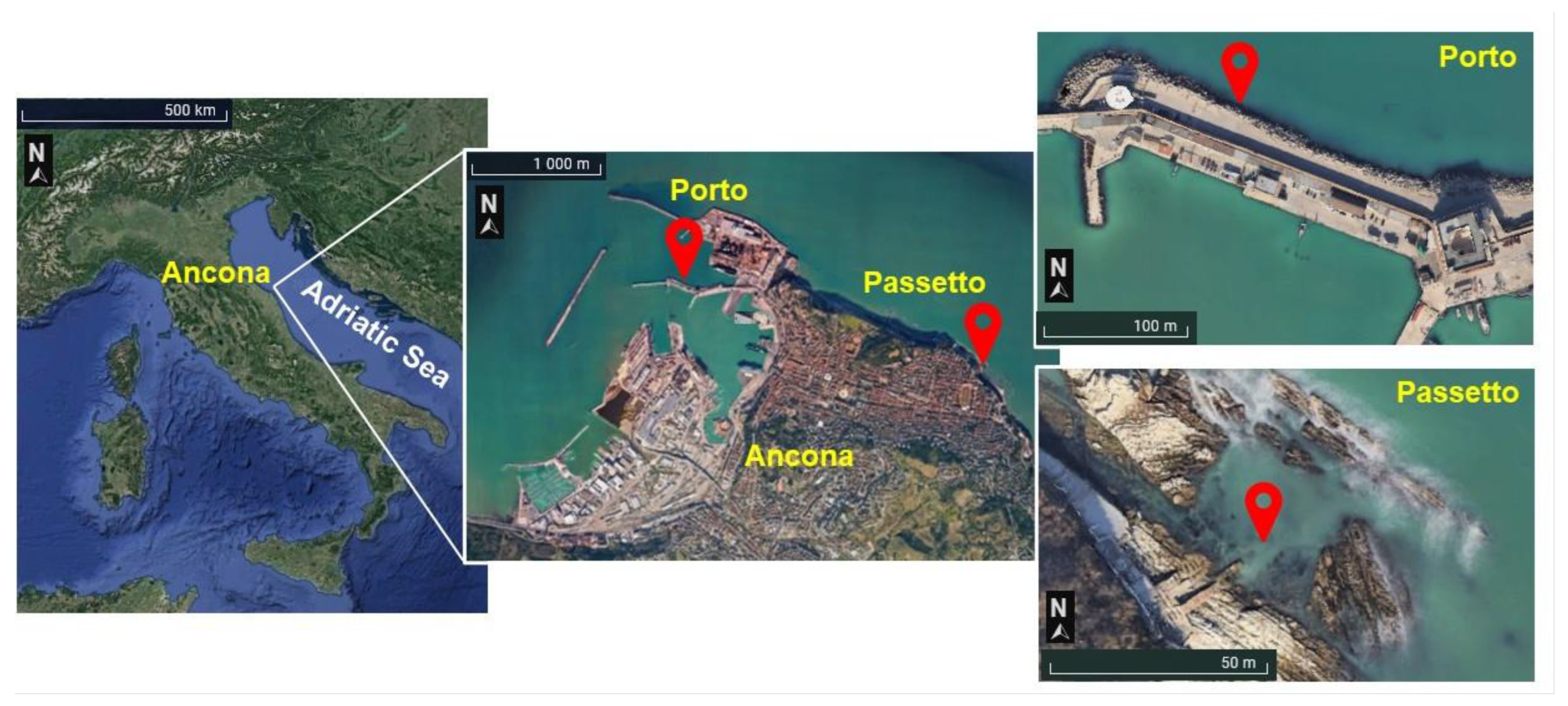
2.1. Study Area
2.2. Morphological Features of the Species Selected
2.3. Sampling and Sample Processing
2.4. Macroalgal Morphology
2.5. Aldehydes (PUAs) Produced by Macroalgae
2.6. Microphytobenthic Assemblages
2.7. Meiobenthic Assemblages
2.8. Data Analysis
3. Results
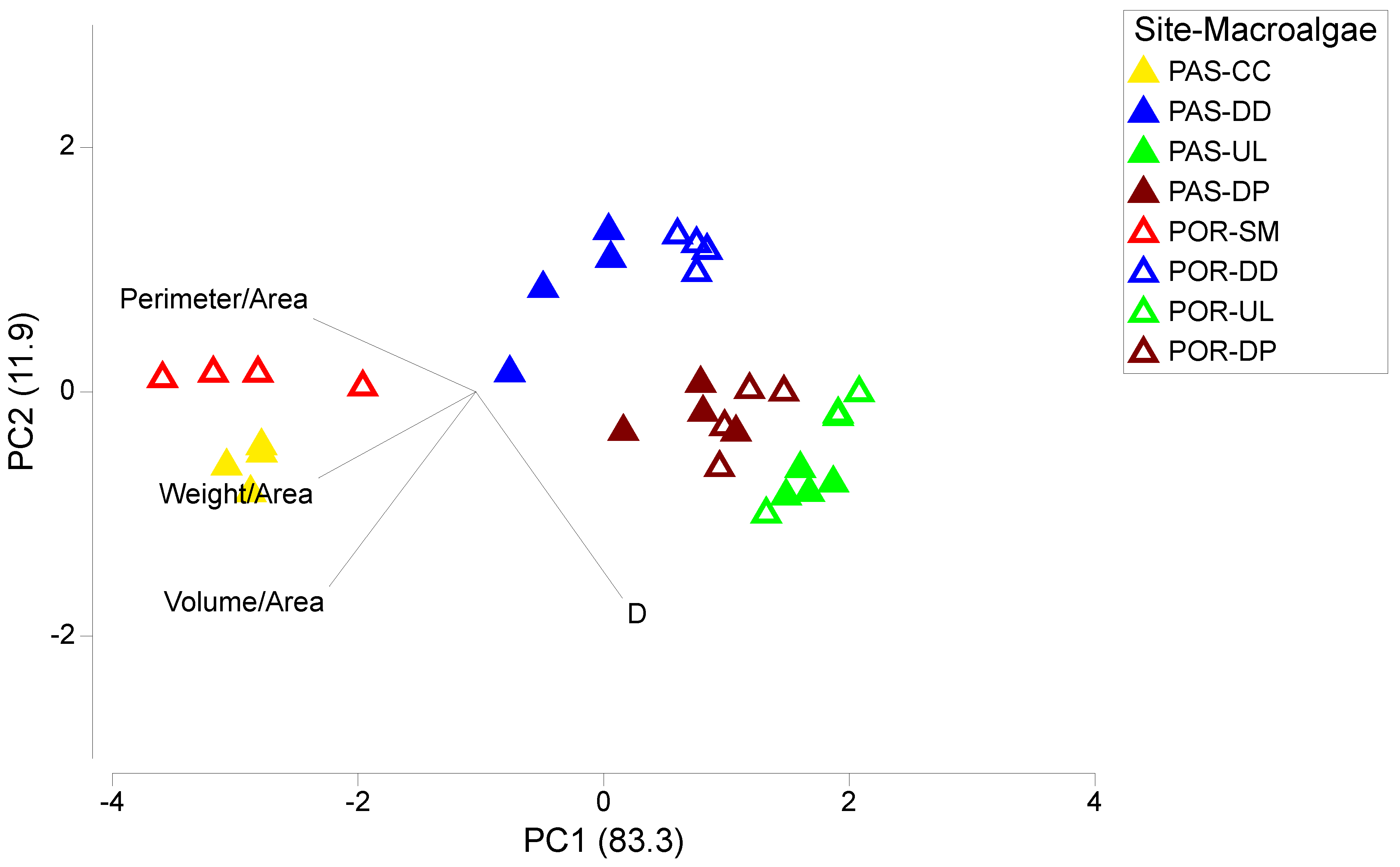
3.1. Macroalgal Morphology
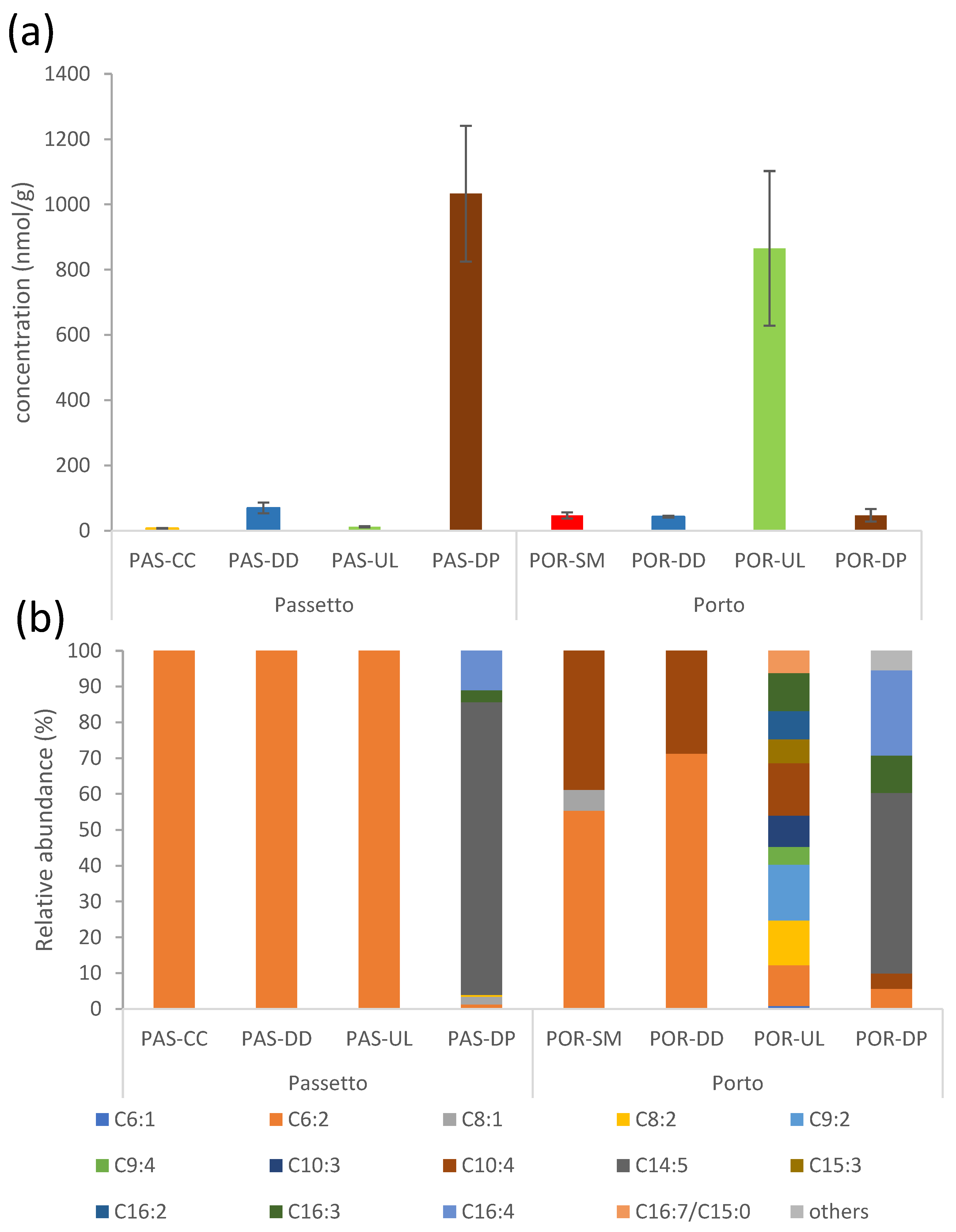
3.2. Unsaturated Aldehydes (PUAs) Produced by Macroalgae
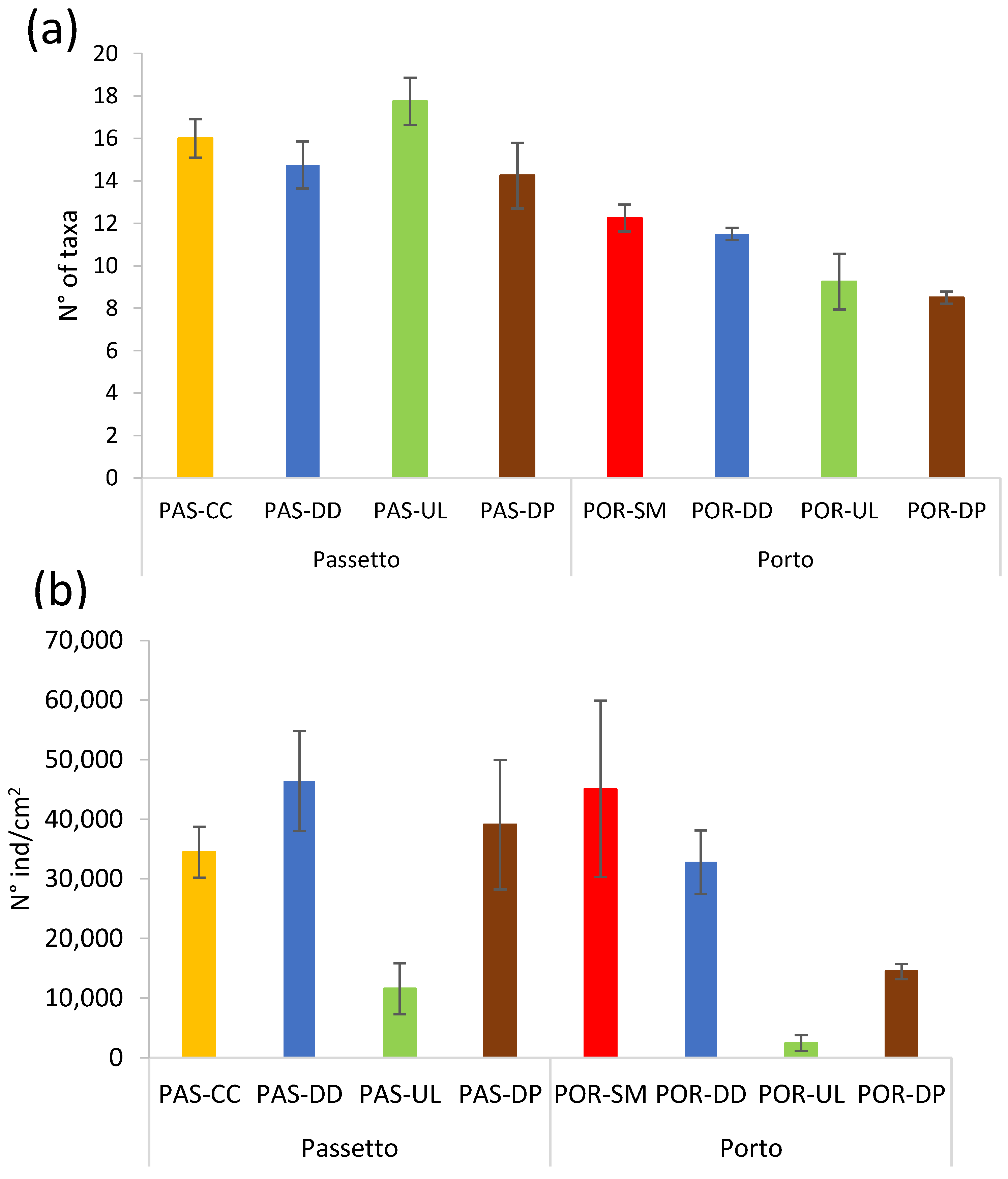
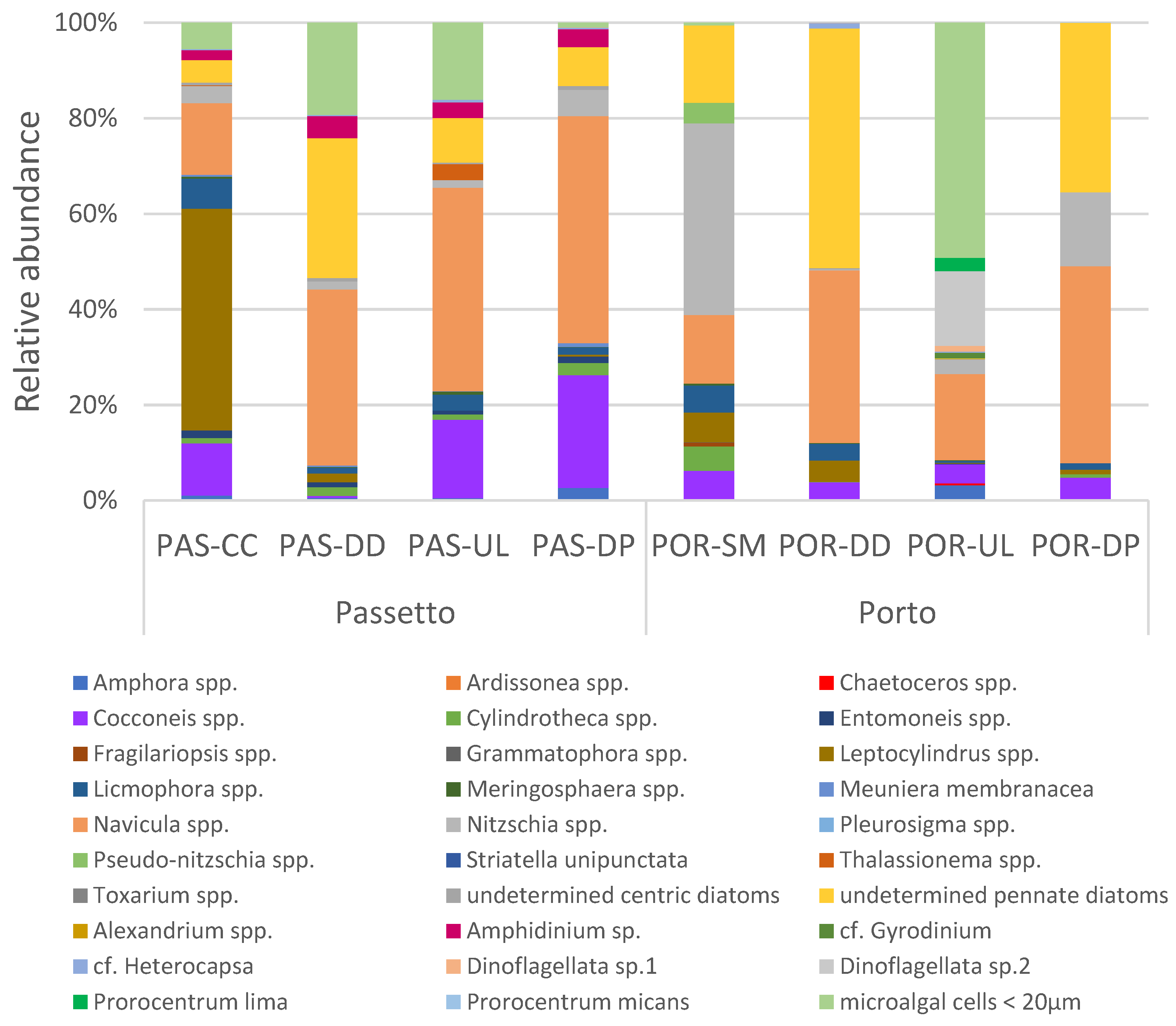
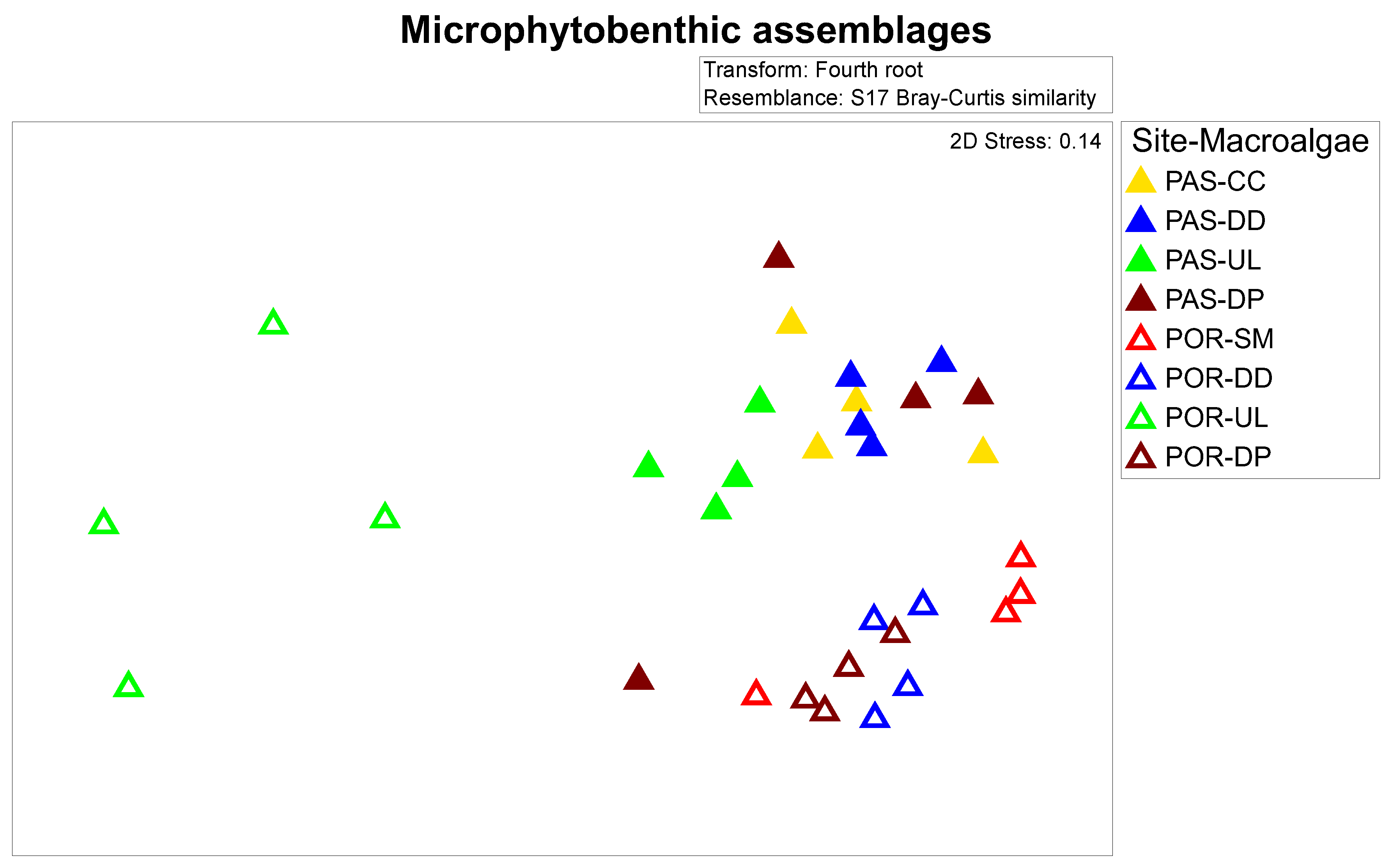
3.3. Microphytobenthic Assemblages
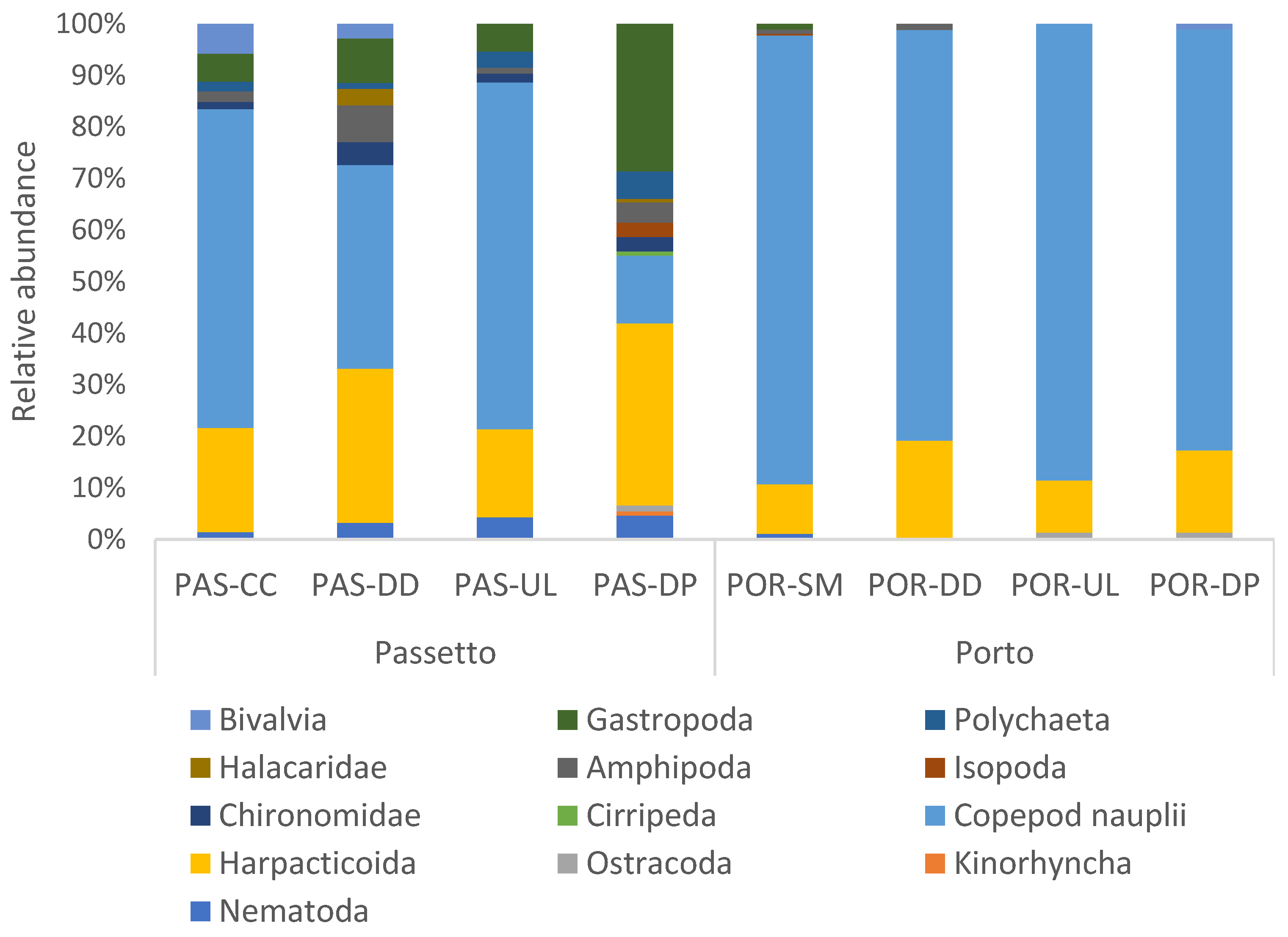
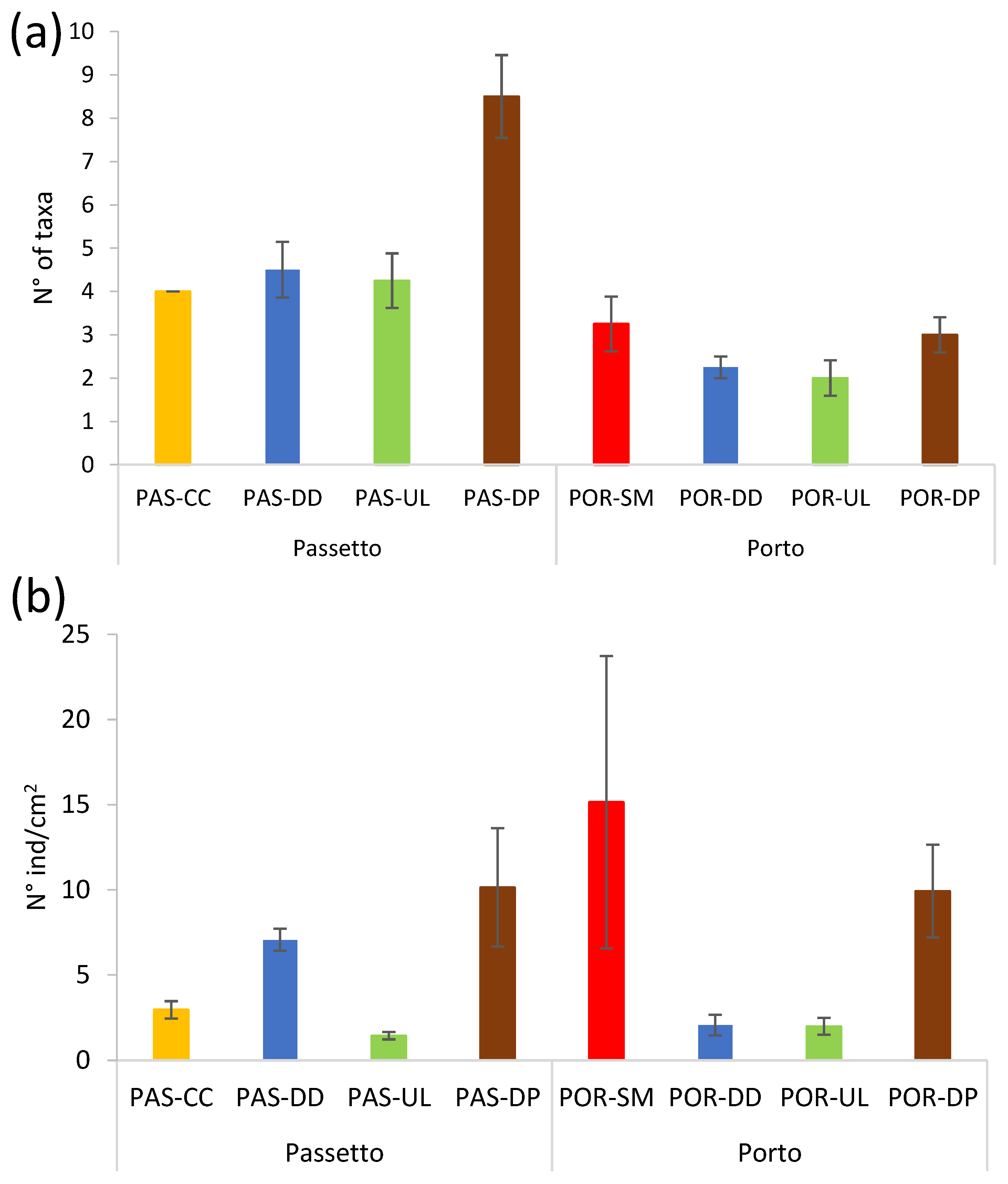
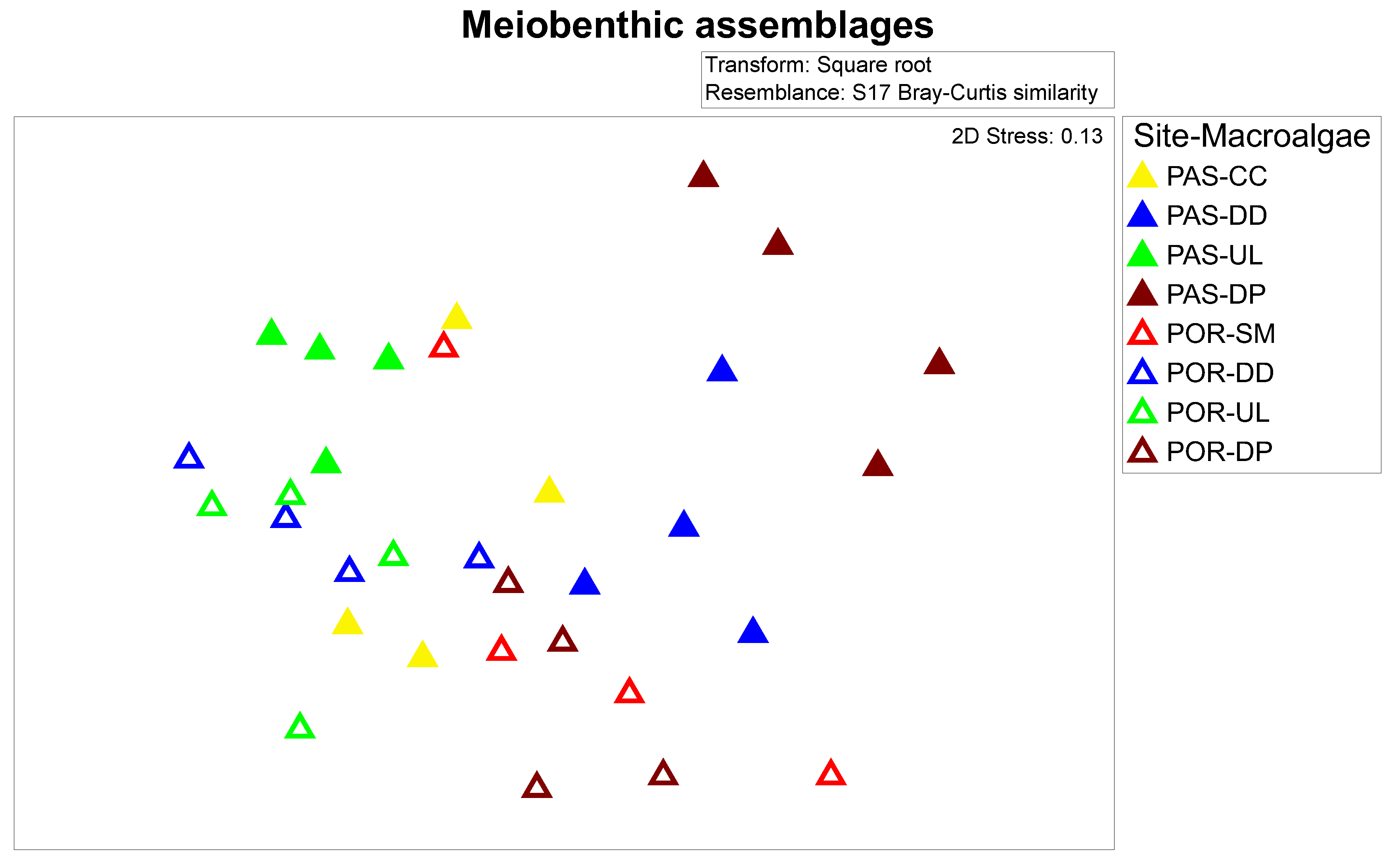
3.4. Meiobenthic Assemblages
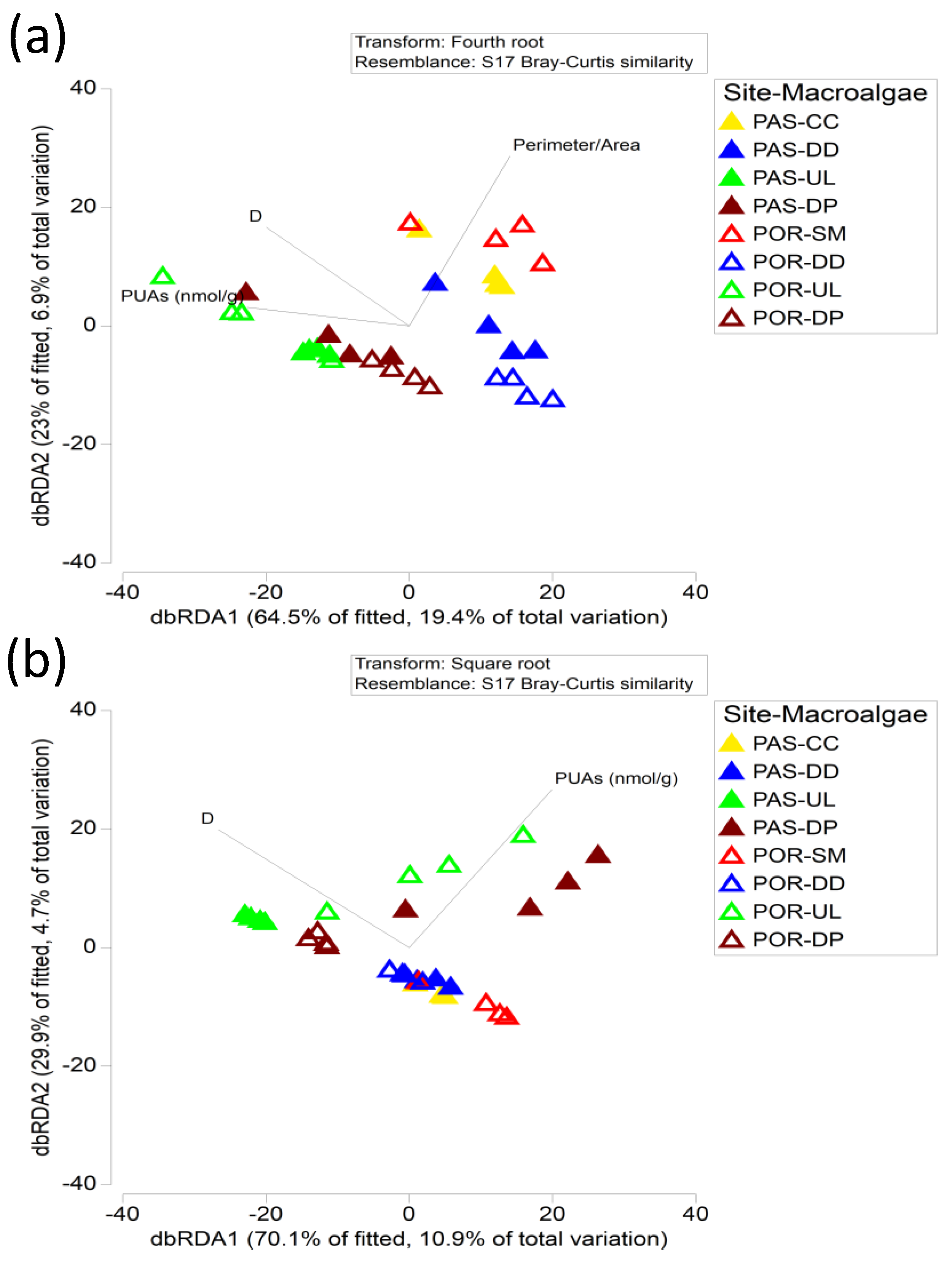
3.5. Relationship between the Microphytobenthos and Meiofauna Assemblages with Macroalgae Complexity and PUAs Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chemello, R.; Milazzo, M. Effect of Algal Architecture on Associated Fauna: Some Evidence from Phytal Molluscs. Mar. Biol. 2002, 140, 981–990. [Google Scholar] [CrossRef]
- Thornber, C.; Jones, E.; Thomsen, M. Epibiont-Marine Macrophyte Assemblages. In Marine Macrophytes as Foundation Species; CRC Press: Boca Raton, FL, USA, 2016; pp. 43–65. [Google Scholar] [CrossRef]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.F.; Pérez, T. Climate Change Effects on a Miniature Ocean: The Highly Diverse, Highly Impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Mannino, A.M.; Balistreri, P.; Deidun, A. The Marine Biodiversity of the Mediterranean Sea in a Changing Climate: The Impact of Biological Invasions. In Mediterranean Identities: Environment, Society, Culture; Books on Demand: Paris, France, 2017. [Google Scholar] [CrossRef] [Green Version]
- Máximo, P.; Ferreira, L.M.; Branco, P.; Lima, P.; Lourenço, A. Secondary Metabolites and Biological Activity of Invasive Macroalgae of Southern Europe. Mar. Drugs 2018, 16, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faria, J.; Prestes, A.C.L.; Moreu, I.; Cacabelos, E.; Martins, G.M. Dramatic Changes in the Structure of Shallow-Water Marine Benthic Communities Following the Invasion by Rugulopteryx Okamurae (Dictyotales, Ochrophyta) in Azores (NE Atlantic). Mar. Pollut. Bull. 2022, 175, 113358. [Google Scholar] [CrossRef] [PubMed]
- Katsanevakis, S.; Wallentinus, I.; Zenetos, A.; Leppäkoski, E.; Çinar, M.E.; Oztürk, B.; Grabowski, M.; Golani, D.; Cardoso, A.C. Impacts of Invasive Alien Marine Species on Ecosystem Services and Biodiversity: A Pan-European Review. Aquat. Invasions 2014, 9, 391–423. [Google Scholar] [CrossRef]
- Grima, E.M.; Pérez, J.A.S.; Camacho, F.G.; Medina, A.R.; Giménez, A.G.; López Alonso, D. The Production of Polyunsaturated Fatty Acids by Microalgae: From Strain Selection to Product Purification. Process Biochem. 1995, 30, 711–719. [Google Scholar] [CrossRef]
- Patil, V.; Källqvist, T.; Olsen, E.; Vogt, G.; Gislerød, H.R. Fatty Acid Composition of 12 Microalgae for Possible Use in Aquaculture Feed. Aquac. Int. 2007, 15, 1–9. [Google Scholar] [CrossRef]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Long Chain Polyunsaturated Fatty Acid Production and Partitioning to Triacylglycerols in Four Microalgae. Phytochemistry 2002, 61, 15–24. [Google Scholar] [CrossRef]
- Ianora, A.; Miralto, A.; Romano, G. Antipredatory Defensive Role of Planktonic Marine Natural Products. In Handbook of Marine Natural Products; Fattorusso, E., Gerwick, W.H., Taglialatela-Scafati, O., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 711–748. ISBN 978-90-481-3834-0. [Google Scholar]
- Pezzolesi, L.; Pichierri, S.; Samorì, C.; Totti, C.; Pistocchi, R. PUFAs and PUAs Production in Three Benthic Diatoms from the Northern Adriatic Sea. Phytochemistry 2017, 142, 85–91. [Google Scholar] [CrossRef]
- Wichard, T.; Poulet, S.A.; Halsband-Lenk, C.; Albaina, A.; Harris, R.; Liu, D.; Pohnert, G. Survey of the Chemical Defence Potential of Diatoms: Screening of Fifty Species for α,β,γ,δ-Unsaturated Aldehydes. J. Chem. Ecol. 2005, 31, 949–958. [Google Scholar] [CrossRef]
- Veiga, P.; Sousa-Pinto, I.; Rubal, M. Meiofaunal Assemblages Associated with Native and Non-Indigenous Macroalgae. Cont. Shelf Res. 2016, 123, 1–8. [Google Scholar] [CrossRef]
- Rindi, F.; Gavio, B.; Díaz-Tapia, P.; Di Camillo, C.G.; Romagnoli, T. Long-Term Changes in the Benthic Macroalgal Flora of a Coastal Area Affected by Urban Impacts (Conero Riviera, Mediterranean Sea). Biodivers. Conserv. 2020, 29, 2275–2295. [Google Scholar] [CrossRef]
- Falace, A.; Alongi, G.; Cormaci, M.; Furnari, G.; Curiel, D.; Cecere, E.; Petrocelli, A. Changes in the Benthic Algae along the Adriatic Sea in the Last Three Decades. Chem. Ecol. 2010, 26, 77–90. [Google Scholar] [CrossRef]
- El Atouani, S.; Belattmania, Z.; Kaidi, S.; Engelen, A.H.; Serrão, E.A.; Chaouti, A.; Reani, A.; Sabour, B. Spatiotemporal Patterns of Phenology of the Alien Phaeophyceae Sargassum Muticum on the Atlantic Coast of Morocco. Sci. Mar. 2021, 85, 103–111. [Google Scholar] [CrossRef]
- Gestoso, I.; Olabarria, C.; Troncoso, J.S. Effects of Macroalgal Identity on Epifaunal Assemblages: Native Species versus the Invasive Species Sargassum Muticum. Helgol. Mar. Res. 2012, 66, 159–166. [Google Scholar] [CrossRef] [Green Version]
- Mollo, E.; Cimino, G.; Ghiselin, M.T. Alien Biomolecules: A New Challenge for Natural Product Chemists. Biol. Invasions 2015, 17, 941–950. [Google Scholar] [CrossRef]
- Pichierri, S.; Accoroni, S.; Pezzolesi, L.; Guerrini, F.; Romagnoli, T.; Pistocchi, R.; Totti, C. Allelopathic Effects of Diatom Filtrates on the Toxic Benthic Dinoflagellate Ostreopsis cf. ovata. Mar. Environ. Res. 2017, 131, 116–122. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Gobler, C.J. Allelopathic Effects of Cochlodinium Polykrikoides Isolates and Blooms from the Estuaries of Long Island, New York, on Co-Occurring Phytoplankton. Mar. Ecol. Prog. Ser. 2010, 406, 19–31. [Google Scholar] [CrossRef]
- Fistarol, G.O.; Legrand, C.; Selander, E.; Hummert, C.; Stolte, W.; Granéli, E. Allelopathy in Alexandrium spp.: Effect on a Natural Plankton Community and on Algal Monocultures. Aquat. Microb. Ecol. 2004, 35, 45–56. [Google Scholar] [CrossRef]
- Legrand, C.; Rengefors, K.; Fistarol, G.O.; Granéli, E. Allelopathy in Phytoplankton-Biochemical, Ecological and Evolutionary Aspects. Phycologia 2003, 42, 406–419. [Google Scholar] [CrossRef] [Green Version]
- Lenzo, D.; Pezzolesi, L.; Samorì, C.; Rindi, F.; Pasteris, A.; Pistocchi, R.; Colangelo, M.A. Allelopathic Interactions between Phytobenthos and Meiofaunal Community in an Adriatic Benthic Ecosystem: Understanding the Role of Aldehydes and Macroalgal Structural Complexity. Sci. Total Environ. 2022, 807, 150827. [Google Scholar] [CrossRef] [PubMed]
- Giere, O. Meiobenthology: The Microscopic Motile Fauna of Aquatic Sediments; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–527. [Google Scholar] [CrossRef]
- Sandulli, R.; Semprucci, F.; Balsamo, M. Taxonomic and Functional Biodiversity Variations of Meiobenthic and Nematode Assemblages across an Extreme Environment: A Study Case in a Blue Hole Cave. Ital. J. Zool. 2014, 81, 508–516. [Google Scholar] [CrossRef] [Green Version]
- Semprucci, F.; Balsamo, M.; Sandulli, R. Assessment of the Ecological Quality (EcoQ) of the Venice Lagoon Using the Structure and Biodiversity of the Meiofaunal Assemblages. Ecol. Indic. 2016, 67, 451–457. [Google Scholar] [CrossRef]
- Chardy, P.; Dauvin, J.C. Carbon Flows in a Subtidal Fine Sand Community from the Western English Channel: A Simulation Analysis. Mar. Ecol. Prog. Ser. 1992, 81, 147–161. [Google Scholar] [CrossRef]
- Accoroni, S.; Romagnoli, T.; Pichierri, S.; Totti, C. Effects of the Bloom of Harmful Benthic Dinoflagellate Ostreopsis cf. ovata on the Microphytobenthos Community in the Northern Adriatic Sea. Harmful Algae 2016, 55, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, M.J. The Impact of Sediment Accumulations, Relative Habitat Complexity and Elevation on Rocky Shore Meiofauna. J. Exp. Mar. Bio. Ecol. 1988, 122, 225–241. [Google Scholar] [CrossRef]
- Pezzolesi, L.; Accoroni, S.; Rindi, F.; Samorì, C.; Totti, C.; Pistocchi, R. Survey of the Allelopathic Potential of Mediterranean Macroalgae: Production of Long-Chain Polyunsaturated Aldehydes (PUAs). Phytochemistry 2021, 189, 112826. [Google Scholar] [CrossRef] [PubMed]
- Hughey, J.R.; Gabrielson, P.W.; Maggs, C.A.; Mineur, F. Genomic Analysis of the Lectotype Specimens of European Ulva Rigida and Ulva Lacinulata (Ulvaceae, Chlorophyta) Reveals the Ongoing Misapplication of Names. Eur. J. Phycol. 2022, 57, 143–153. [Google Scholar] [CrossRef]
- Alsufyani, T.; Engelen, A.H.; Diekmann, O.E.; Kuegler, S.; Wichard, T. Prevalence and Mechanism of Polyunsaturated Aldehydes Production in the Green Tide Forming Macroalgal Genus Ulva (Ulvales, Chlorophyta). Chem. Phys. Lipids 2014, 183, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.A.; Engelen, A.H.; Santos, R.O.P. Macro- and Mesoherbivores Prefer Native Seaweeds over the Invasive Brown Seaweed Sargassum Muticum: A Potential Regulating Role on Invasions. Mar. Biol. 2009, 156, 2505–2515. [Google Scholar] [CrossRef]
- Veiga, P.; Rubal, M.; Sousa-Pinto, I. Structural Complexity of Macroalgae Influences Epifaunal Assemblages Associated with Native and Invasive Species. Mar. Environ. Res. 2014, 101, 115–123. [Google Scholar] [CrossRef]
- Utermöhl, H. Methods of Collecting Plankton for Various Purposes Are Discussed. SIL Commun. 1953–1996 1958, 9, 1–38. [Google Scholar] [CrossRef]
- Edler, L.; Elbrächter, M. Microscopic and Molecular Methods for Quantitative Phytoplankton Analysis; Karlson, B., Cusack, C., Bresnan, E., Eds.; UNESCO: Paris, France, 2010; pp. 13–20. [Google Scholar]
- Tomas, C.R. Identifying Marine Phytoplankton; Academic Press: Cambridge, MA, USA, 1997. [Google Scholar]
- Kraberg, A.; Baumann, M.; Dürselen, C.-D. Coastal Phytoplankton: Photo Guide for Northern European Seas. 2010. Available online: https://repozitorij.uni-lj.si/IzpisGradiva.php?id=37653 (accessed on 20 February 2023).
- Bertalot, H.L.; Witowski, A.; Metzeltin, D. Diatom Flora of Marine Coasts. In Vol. 1 Iconographia Diatomologica: Annotated Diatom Micrographs Vol 7 Diversity-Taxonomy-Identification; Lange, H.E., Ed.; Koeltz: Grafenau, Germany, 2000. [Google Scholar]
- Horiguchi, E. Marine Benthic Dinoflagellates-Unveiling Their Worldwide Biodiversity; Schweizerbart Science Publishers: Stuttgart, Germany, 2014. [Google Scholar]
- Guiry, M.D.; Guiry, G.M. AlgaeBase—World-Wide Electronic Publication; National University of Ireland: Galway, Ireland, 2013. [Google Scholar]
- Anderson, M.J.; Robinson, J. Permutation Tests for Linear Models. Aust. N. Z. J. Stat. 2001, 43, 75–88. [Google Scholar] [CrossRef]
- Anderson, M.J.; Connell, S.D.; Gillanders, B.M.; Diebel, C.E.; Blom, W.M.; Saunders, J.E.; Landers, T.J. Relationships between Taxonomic Resolution and Spatial Scales of Multivariate Variation. J. Anim. Ecol. 2005, 74, 636–646. [Google Scholar] [CrossRef]
- Clarke, K.R.; Tweedley, J.R.; Valesini, F.J. Simple Shade Plots Aid Better Long-Term Choices of Data Pre-Treatment in Multivariate Assemblage Studies. J. Mar. Biol. Assoc. U. K. 2014, 94, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Clarke, K.R. Non-parametric Multivariate Analyses of Changes in Community Structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Getting Started with PRIMER v7; PRIMER-E: Plymouth Marine Laboratory: Plymouth, UK, 2015; 20p. [Google Scholar]
- Anderson, M.; Gorley, R.; Clarke, K. PERMANOVA for PRIMER: Guide to Software and Statistical Methods; PRIMER–E Ltd.: Plymouth, UK, 2008. [Google Scholar]
- Torres, A.C.; Veiga, P.; Rubal, M.; Sousa-Pinto, I. The Role of Annual Macroalgal Morphology in Driving Its Epifaunal Assemblages. J. Exp. Mar. Bio. Ecol. 2015, 464, 96–106. [Google Scholar] [CrossRef]
- Johnson, S.C.; Scheibling, R.E. Reproductive Patterns of Harpacticoid Copepods on Intertidal Macroalgae (Ascophyllum Nodosum and Fucus Vesiculosus) in Nova Scotia, Canada. Can. J. Zool. 1987, 65, 129–141. [Google Scholar] [CrossRef]
- Losi, V.; Sbrocca, C.; Gatti, G.; Semprucci, F.; Rocchi, M.; Bianchi, C.N.; Balsamo, M. Sessile Macrobenthos (Ochrophyta) Drives Seasonal Change of Meiofaunal Community Structure on Temperate Rocky Reefs. Mar. Environ. Res. 2018, 142, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Ape, F.; Gristina, M.; Chemello, R.; Sarà, G.; Mirto, S. Meiofauna Associated with Vermetid Reefs: The Role of Macroalgae in Increasing Habitat Size and Complexity. Coral Reefs 2018, 37, 875–889. [Google Scholar] [CrossRef] [Green Version]
- Akakabe, Y.; Matsui, K.; Kajiwara, T. 2,4-Decadienals Are Produced via (R)-11-HPITE from Arachidonic Acid in Marine Green Alga Ulva Conglobata. Bioorg. Med. Chem. 2003, 11, 3607–3609. [Google Scholar] [CrossRef] [PubMed]
- Andrade, P.B.; Barbosa, M.; Matos, R.P.; Lopes, G.; Vinholes, J.; Mouga, T.; Valentão, P. Valuable Compounds in Macroalgae Extracts. Food Chem. 2013, 138, 1819–1828. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Biologically Active Oxylipins from Enzymatic and Nonenzymatic Routes in Macroalgae. Mar. Drugs 2016, 14, 23. [Google Scholar] [CrossRef] [Green Version]
- Kajiwara, T.; Matsui, K.; Akakabe, Y. Biogeneration of Volatile Compounds via Oxylipins in Edible Seaweeds. In Biotechnology for Improved Foods and Flavors; ACS Symposium Series; Takeoka, G.R., Teranishi, R., Williams, P.J., Kobayashi, A., Eds.; American Chemical Society: Washington, DC, USA, 1996; pp. 146–166. [Google Scholar]
- Melton, J.T.; Lopez-Bautista, J.M. Diversity of the Green Macroalgal Genus Ulva (Ulvophyceae, Chlorophyta) from the East and Gulf Coast of the United States Based on Molecular Data. J. Phycol. 2021, 57, 551–568. [Google Scholar] [CrossRef] [PubMed]
- Hayden, H.S.; Blomster, J.; Maggs, C.A.; Silva, P.C.; Stanhope, M.J.; Waaland, J.R. Linnaeus Was Right All along: Ulva and Enteromorpha Are Not Distinct Genera. Eur. J. Phycol. 2003, 38, 277–294. [Google Scholar] [CrossRef]
- Steinhagen, S.; Karez, R.; Weinberger, F. Cryptic, Alien and Lost Species: Molecular Diversity of Ulva Sensu Lato along the German Coasts of the North and Baltic Seas. Eur. J. Phycol. 2019, 54, 466–483. [Google Scholar] [CrossRef] [Green Version]
- Totti, C.; Cucchiari, E.; De Stefano, M.; Pennesi, C.; Romagnoli, T.; Bavestrello, G. Seasonal Variations of Epilithic Diatoms on Different Hard Substrates, in the Northern Adriatic Sea. J. Mar. Biol. Assoc. U. K. 2007, 87, 649–658. [Google Scholar] [CrossRef]
- D’Alelio, D.; Cante, M.T.; Russo, G.F.; Totti, C.; De Stefano, M. Epizoic Diatoms on Gastropod Shells. When Substrate Complexity Selects for Microcommunity Complexity. In All Flesh Is Grass: Plant-Animal Interrelationships; Springer: Berlin/Heidelberg, Germany, 2011; pp. 345–364. [Google Scholar] [CrossRef]
- Pinckney, J.; Zingmark, R.G. Biomass and Production of Benthic Microalgal Communities in Estuarine Habitats. Estuaries 1993, 16, 887–897. [Google Scholar] [CrossRef]
- Paterson, D.M.; Wiltshire, K.H.; Miles, A.; Blackburn, J.; Davidson, I.; Yates, M.G.; McGrorty, S.; Eastwood, J.A. Microbiological Mediation of Spectral Reflectance from Intertidal Cohesive Sediments. Limnol. Oceanogr. 1998, 43, 1207–1221. [Google Scholar] [CrossRef]
- Car, A.; Witkowski, A.; Dobosz, S.; Jasprica, N.; Ljubimir, S.; Zgłobicka, I. Epiphytic Diatom Assemblages on Invasive Caulerpa taxifolia and Autochthonous Halimeda tuna and Padina sp. Seaweeds in the Adriatic Sea-Summer/Autumn Aspect. Oceanol. Hydrobiol. Stud. 2019, 48, 209–226. [Google Scholar] [CrossRef]
- Wiltshire, K.H. Algae and Associated Pigments of Intertidal Sediments, New Observations and Methods. Limnologica 2000, 30, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Goericke, R.; Montoya, J.P. Estimating the Contribution of Microalgal Taxa to Chlorophyll a in the Field-Variations of Pigment Ratios under Nutrient- and Light-Limited Growth. Mar. Ecol. Prog. Ser. 1998, 169, 97–112. [Google Scholar] [CrossRef]
- Da Rocha, C.M.C.; Venekey, V.; Bezerra, T.N.C.; Souza, J.R.B. Phytal Marine Nematode Assemblages and Their Relation with the Macrophytes Structural Complexity in a Brazilian Tropical Rocky Beach. Hydrobiologia 2006, 553, 219–230. [Google Scholar] [CrossRef]
- Mancuso, F.P.; Milazzo, M.; Sarà, G.; Chemello, R. Bi- and Three-Dimensional Fractal Analysis of the Brown Seaweed Gongolaria Montagnei and Their Relationship with Gastropod Molluscs Assemblage. Mar. Pollut. Bull. 2023, 186, 114396. [Google Scholar] [CrossRef]
- Russo, A.R. The Role of Seaweed Complexity in Structuring Hawaiian Epiphytal Amphipod Communities. Hydrobiologia 1990, 194, 1–12. [Google Scholar] [CrossRef]
- Richardson, M.; Stephens, T. Meiofaunal Diversity on Invasive Sargassum Muticum versus Native Seaweeds. 2014, pp. 1–22. Available online: https://digital.lib.washington.edu/researchworks/handle/1773/27315 (accessed on 15 February 2023).
- Somerfield, P.J.; Yodnarasri, S.; Aryuthaka, C. Relationships between Seagrass Biodiversity and Infaunal Communities: Implications for Studies of Biodiversity Effects. Mar. Ecol. Prog. Ser. 2002, 237, 97–109. [Google Scholar] [CrossRef]
- Hicks, G.R.F. Structure of Phytal Harpacticoid Copepod Assemblages and the Influence of Habitat Complexity and Turbidity. J. Exp. Mar. Bio. Ecol. 1980, 44, 157–192. [Google Scholar] [CrossRef]
- Gestoso, I.; Olabarria, C.; Troncoso, J.S. Variability of Epifaunal Assemblages Associated with Native and Invasive Macroalgae. Mar. Freshw. Res. 2010, 61, 724–731. [Google Scholar] [CrossRef]
- McAbendroth, L.; Ramsay, P.M.; Foggo, A.; Rundle, S.D.; Bilton, D.T. Does Macrophyte Fractal Complexity Drive Invertebrate Diversity, Biomass and Body Size Distributions? Oikos 2005, 111, 279–290. [Google Scholar] [CrossRef]
- Hansen, J.P.; Sagerman, J.; Wikström, S.A. Effects of Plant Morphology on Small-Scale Distribution of Invertebrates. Mar. Biol. 2010, 157, 2143–2155. [Google Scholar] [CrossRef]
- Hansen, J.P.; Wikström, S.A.; Axemar, H.; Kautsky, L. Distribution Differences and Active Habitat Choices of Invertebrates between Macrophytes of Different Morphological Complexity. Aquat. Ecol. 2011, 45, 11–22. [Google Scholar] [CrossRef]
- Cremona, F.; Planas, D.; Lucotte, M. Biomass and Composition of Macroinvertebrate Communities Associated with Different Types of Macrophyte Architectures and Habitats in a Large Fluvial Lake. Fundam. Appl. Limnol. 2008, 171, 119–130. [Google Scholar] [CrossRef]
- Schneider, F.I.; Mann, K.H. Species Specific Relationships of Invertebrates to Vegetation in a Seagrass Bed. II. Experiments on the Importance of Macrophyte Shape, Epiphyte Cover and Predation. J. Exp. Mar. Biol. Ecol. 1991, 145, 119–139. [Google Scholar] [CrossRef]
- Bologna, P.A.X.; Heck, K.L. Macrofaunal Associations with Seagrass Epiphytes Relative Importance of Trophic and Structural Characteristics. J. Exp. Mar. Biol. Ecol. 1999, 242, 21–39. [Google Scholar] [CrossRef]
- Cacabelos, E.; Olabarria, C.; Incera, M.; Troncoso, J.S. Effects of Habitat Structure and Tidal Height on Epifaunal Assemblages Associated with Macroalgae. Estuar. Coast. Shelf Sci. 2010, 89, 43–52. [Google Scholar] [CrossRef]
- Kadiene, E.U.; Bialais, C.; Ouddane, B.; Hwang, J.S.; Souissi, S. Differences in Lethal Response between Male and Female Calanoid Copepods and Life Cycle Traits to Cadmium Toxicity. Ecotoxicology 2017, 26, 1227–1239. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site–Macroalgae | Weight (g) | Volume (mL) | Area (cm2) | Perimeter (cm) | D | Weight/Area | Perimeter/Area | Volume/Area |
|---|---|---|---|---|---|---|---|---|
| PAS-CC1 | 0.80 ± 0.07 | 1.05 ± 0.06 | 9.74 ± 0.77 | 185.10 ± 14.39 | 1.74 ± 0.01 | 0.08 ± 0.002 | 19.03 ± 0.57 | 0.11 ± 0.00 |
| PAS-DD1 | 0.13 ± 0.01 | 0.28 ± 0.09 | 5.16 ± 0.39 | 68.09 ± 9.30 | 1.76 ± 0.01 | 0.03 ± 0.002 | 13.06 ± 0.86 | 0.05 ± 0.01 |
| PAS-UL1 | 0.45 ± 0.03 | 1.23 ± 0.09 | 25.68 ± 2.23 | 49.70 ± 4.70 | 1.94 ± 0.00 | 0.02 ± 0.000 | 1.99 ± 0.26 | 0.05 ± 0.00 |
| PAS-DP1 | 0.34 ± 0.02 | 0.93 ± 0.05 | 15.71 ± 1.26 | 56.98 ± 2.76 | 1.84 ± 0.01 | 0.02 ± 0.001 | 3.69 ± 0.32 | 0.06 ± 0.01 |
| POR-SM1 | 0.66 ± 0.08 | 1.18 ± 0.12 | 11.12 ± 1.28 | 257.45 ± 29.51 | 1.70 ± 0.02 | 0.06 ± 0.003 | 23.22 ± 1.06 | 0.11 ± 0.01 |
| POR-DD1 | 0.32 ± 0.01 | 0.50 ± 0.00 | 19.59 ± 0.52 | 150.48 ± 7.08 | 1.78 ± 0.01 | 0.02 ± 0.001 | 7.68 ± 0.30 | 0.03 ± 0.00 |
| POR-UL1 | 0.28 ± 0.06 | 1.38 ± 0.24 | 35.84 ± 7.70 | 66.42 ± 13.77 | 1.92 ± 0.00 | 0.01 ± 0.000 | 1.88 ± 0.16 | 0.04 ± 0.01 |
| POR-DP1 | 0.36 ± 0.06 | 0.93 ± 0.22 | 18.75 ± 2.98 | 68.40 ± 11.78 | 1.88 ± 0.01 | 0.02 ± 0.000 | 3.63 ± 0.06 | 0.05 ± 0.01 |
| Dead Copepod nauplii | Dead Harpacticoids | Expelled Egg Sacs | |
|---|---|---|---|
| PAS-CC | 0.06 ± 0.06 | 8.25 ± 0.06 | |
| PAS-DD | 0.10 ± 0.06 | 16.02 ± 0.06 | |
| PAS-UL | 0.05 ± 0.02 | 1.11 ± 0.02 | |
| PAS-DP | 0.31 ± 0.17 | 0.37 ± 0.12 | 2.84 ± 0.12 |
| POR-SM | 0.11 ± 0.11 | 0.05 ± 0.03 | 4.03 ± 0.03 |
| POR-DD | 0.03 ± 0.03 | 0.03 ± 0.02 | 0.08 ± 0.02 |
| POR-UL | 0.03 ± 0.02 | 0.01 ± 0.01 | 0.11 ± 0.01 |
| POR-DP | 0.08 ± 0.04 | 0.09 ± 0.03 | 0.91 ± 0.03 |
| (a) | Variable | Pseudo-F | P | Prop |
|---|---|---|---|---|
| Marginal test | D | 5.441 | 0.0001 | 0.154 |
| Weight/Area | 3.850 | 0.0033 | 0.114 | |
| Perimeter/Area | 4.288 | 0.0011 | 0.125 | |
| Volume/Area | 3.184 | 0.0072 | 0.096 | |
| PUAs (nmol/g) | 3.745 | 0.0061 | 0.111 | |
| Model | AICc | R2 | RSS | |
| Best solution | 222.0300 | 0.30 | 24533 | |
| D; Perimeter/Area; PUAs | ||||
| (b) | Variable | Pseudo-F | P | Prop |
| Marginal test | D | 1.598 | 0.1684 | 0.051 |
| Weight/Area | 0.640 | 0.6566 | 0.021 | |
| Perimeter/Area | 0.942 | 0.4193 | 0.030 | |
| Volume/Area | 0.706 | 0.6045 | 0.023 | |
| PUAs (nmol/g) | 1.438 | 0.2063 | 0.046 | |
| Model | AICc | R2 | RSS | |
| Best solution | 227.72 | 0.05 | 34342 | |
| D | ||||
| (c) | Variable | Pseudo-F | P | Prop |
| Marginal test | D | 1.598 | 0.165 | 0.051 |
| Weight/Area | 0.640 | 0.648 | 0.021 | |
| Perimeter/Area | 0.942 | 0.427 | 0.030 | |
| Volume/Area | 0.706 | 0.593 | 0.023 | |
| PUAs (nmol/g) | 1.438 | 0.204 | 0.046 | |
| Abund Microphyto | 3.610 | 0.010 | 0.107 | |
| Model | AICc | R2 | RSS | |
| Best solution | 225.60 | 0.18 | 29777 | |
| PUAs; Abund Microphyto | ||||
| (d) | Variable | Pseudo-F | P | Prop |
| D | 2.153 | 0.0561 | 0.067 | |
| Weight/Area | 1.764 | 0.1056 | 0.056 | |
| Perimeter/Area | 1.542 | 0.1617 | 0.049 | |
| Volume/Area | 1.172 | 0.3007 | 0.038 | |
| PUAs (nmol/g) | 2.226 | 0.0485 | 0.069 | |
| Model | AICc | R2 | RSS | |
| Best solution | 233.70 | 0.16 | 38351 | |
| D; PUAs | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lenzo, D.; Colangelo, M.A.; Pasteris, A.; Rindi, F.; Pistocchi, R.; Pezzolesi, L. Understanding the Role of Macroalgal Complexity and Allelochemicals Production in Invasive and Non-Invasive Macroalgae in the North-Western Adriatic Sea: Effect on the Associated Communities. Water 2023, 15, 1697. https://doi.org/10.3390/w15091697
Lenzo D, Colangelo MA, Pasteris A, Rindi F, Pistocchi R, Pezzolesi L. Understanding the Role of Macroalgal Complexity and Allelochemicals Production in Invasive and Non-Invasive Macroalgae in the North-Western Adriatic Sea: Effect on the Associated Communities. Water. 2023; 15(9):1697. https://doi.org/10.3390/w15091697
Chicago/Turabian StyleLenzo, Denise, Marina Antonia Colangelo, Andrea Pasteris, Fabio Rindi, Rossella Pistocchi, and Laura Pezzolesi. 2023. "Understanding the Role of Macroalgal Complexity and Allelochemicals Production in Invasive and Non-Invasive Macroalgae in the North-Western Adriatic Sea: Effect on the Associated Communities" Water 15, no. 9: 1697. https://doi.org/10.3390/w15091697