
Research Progress of Zebrafish Model in Aquatic Ecotoxicology

Institute of Life Sciences & Biomedicine Collaborative Innovation Center of Zhejiang Province, College of Life and Environmental Science, Wenzhou University, Wenzhou 325035, China
*
Authors to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Water 2023, 15(9), 1735; https://doi.org/10.3390/w15091735
Submission received: 29 March 2023
/
Revised: 24 April 2023
/
Accepted: 28 April 2023
/
Published: 30 April 2023
(This article belongs to the Section Water, Agriculture and Aquaculture)
Abstract
:In recent years, with the aggravation of environmental pollution, the study of aquatic ecotoxicology has become a hot topic. Some aquatic organisms, such as large fleas, toads, and zebrafish, have been developed and applied as model organisms. They have been increasingly used to study the bioaccumulation and toxicity of environmental pollutants due to the advantages of their small size, easy reproduction, short life cycle, low maintenance cost, and ability to combine genetic, cellular, and whole organism endpoints. This review summarizes the methods of zebrafish, as a model in aquatic ecotoxicology, using its developmental toxicity, biomarkers, genomics, and phenotype analysis to evaluate the toxicity of environmental pollutants, providing a reference value for researchers to select model animals for aquatic ecotoxicology research.
1. Introduction
The study of aquatic ecotoxicology focuses on the effects of environmental pollutants on biological communities in aquatic ecosystems, and the application of biological models is an effective means to reveal the toxicological mechanisms of exogenous compounds. In recent years, with the continuous emergence of research results in the field of aquatic ecotoxicology, some aquatic organisms, such as Scenedesmus obliquus, Daphnia magna, toads, and zebrafish, have been developed and used as model organisms [1,2]. Because of its clear genetic background and multiple advantages, the zebrafish has been selected by the Organization for Economic Cooperation and Development (OECD) and the International Organization for Standardization (ISO) as the standard experimental fish. In China, zebrafish are used in water quality testing according to the “Methods for Determination of Acute Toxicity of Water Quality Substances to Freshwater fish (zebrafish)” issued in 1992. Since 2010, the National Zebrafish Model Animal South Center (Shanghai), the National Zebrafish Model Animal North Center (Beijing), and the National Zebrafish Resource Center (Wuhan) have been successively established to provide a more powerful driving force for the research and application of zebrafish.
The main advantage of zebrafish as a model organism is that it is easy to rear, handle, observe, and analyze. The research results have crucial reference value, and zebrafish have become an ideal model organism in developmental biology, biogenetics, and molecular biology research fields, such as drug screening, disease research, and evaluation of the ecological environment [3,4]. The zebrafish genome shares 87% homology with the human genome, and its signal transduction pathway, physiological structure, and function are highly similar to those of mammals. Therefore, the results obtained by zebrafish toxicity tests are also applicable to humans in most cases [5]. Compared with in vitro toxicity tests, in vivo toxicity tests of zebrafish embryos, larvae or adults can more accurately reflect the absorption, distribution, metabolism, and excretion of the tested substances in vivo [6]. As an ideal animal model, zebrafish can be used not only for behavioral observation but also for observing organ changes and immune responses and for analyzing the entire process of absorption, metabolism, and distribution of poisons (drugs) in the body [7]. Therefore, due to the above characteristics, zebrafish, as a typical vertebrate in aquatic ecosystems, have been widely used in the field of aquatic ecotoxicology and show unique advantages. In this review, we detail the application of zebrafish as a model organism in aquatic ecotoxicology. Methods of aquatic ecotoxicology assessment based on developmental toxicity of zebrafish, molecular markers, the genomic and phenotypic analyses highlighted the advantages, and universality of zebrafish models in aquatic ecotoxicology and provided reference value for researchers to select model animals.
The three stages of zebrafish development—embryo, juvenile, and adult—can be used as experimental subjects for aquatic ecotoxicology research. Zebrafish embryos are produced by parental eggs and used for developmental toxicity tests. After toxicant exposure, lethal and teratogenic effects such as egg condensation, delayed hatching, and morphological deformity are often observed as the endpoint [8]. Zebrafish can move freely and feed autonomously after 96–120 h of embryo incubation, so behavior is often used as an evaluation index for toxicity tests at this stage [9]. Zebrafish at 3 months old are adult fish, and the tissues, organs, and endocrine system in the body have been developed, so a variety of biomarkers and endpoints can be observed, and organs in the body can be removed to evaluate the effects of exogenous substances on different tissues and organs [10] (Table 1). In recent years, with the development of transgenic technology and the improvement of zebrafish genome sequencing, an increasing number of transgenic lines of zebrafish have been established. Some studies have established a line of transparent adult fish without stripes because the stripes in adult zebrafish make direct observation difficult, and the changes in tissues and organs in adult zebrafish under the influence of exogenous substances can be observed with the naked eye or through stereoscopic microscopy [11]. Furthermore, a transgenic strain of zebrafish using the promoter of heat shock protein 70 (HSP70) as a response element to drive the expression of green fluorescent protein (GFP) gene was established, which could even detect cadmium pollution as low as 0.2 μmol/L in an effluent environment [12]. The cms1Tg/+ (AB) mitochondrial fluorescent transgenic zebrafish construction can directly judge the degree of mitochondrial function and structural damage caused by environmental stress [13]. Compared with wild-type zebrafish, the use of transgenic technology to obtain results in terms of sensitivity and specificity has greatly increased, enhancing the monitoring stability of the low concentration range and promoting the specificity of single material testing. At the same time, the experimental equipment design, operation, and subsequent data analysis are more convenient, save manpower, material resources, and time, and pay more attention to ecological and environmental protection.
2. Evaluating the Effect of Ecological Water Pollution on Zebrafish
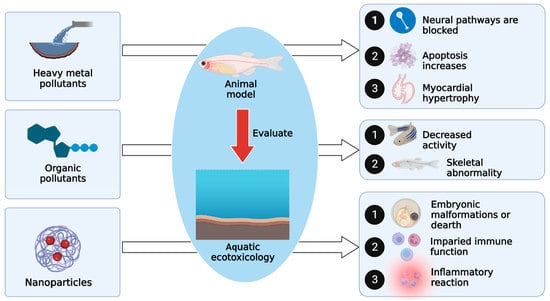
2.1. Evaluating the Toxicological Effects of Heavy Metal Pollution on Zebrafish in Water Environments
Heavy metals are major contaminants in the aqueous environment and include elemental metals (such as Al, Se, and Te), oxides (such as titanium dioxide), and methylates (such as methylmercury). They have high application value in food, materials, environmental protection, chemistry, and biomedicine but are highly toxic and have substantial side effects on the environment [25,26,27]. Biotoxicity test objects include cells, bacteria, algae, water fleas, fish, laboratory mice, and plants, among which zebrafish have great advantages, and common heavy metal pollutants, such as Cd, have different toxicological effects on zebrafish at different developmental stages (Figure 1). Some studies have used metal response elements (MREs) and other response elements to drive the expression of reporter genes (such as fluorescent proteins) to monitor metal pollutants. Endpoints and biomarkers, such as vital signs, tissue distribution of trace elements, and expression of enzymes related to oxidation or degradation reactions, were used to analyze the action pathways of pollutants, and the efficiency and specificity of monitoring were improved through innovation, fusion, and optimal repetition attempts of reaction elements [28]. In an experiment to explore the effect of 16 rare earth elements (REEs) on the embryonic development of zebrafish, one study used transcriptome technology to analyze zebrafish exposed to residual mass concentrations of lanthanum and praseodymium (1.1–10 mg/L) in the water environment of severely polluted areas. The results showed that the two elements had similar mechanisms of action, including 5-hydroxytryptamine neurotransmitter signaling pathway interference, cardiac tissue DNA damage, increased apoptotic activity, myocardial hypertrophy, and impaired myocardial contractile function and cardiac function, which affect embryonic cardiovascular morphology, locomotor behavior, and cardiac physiological abnormalities [29].
2.2. Evaluating the Toxicological Effects of Organic Pollutants on Zebrafish in Water Environments
Another large category of water pollution is organic pollution. Common categories include aromatic hydrocarbon pollutants, chiral organic endocrine-disrupting chemicals (EDCs), chiral organic flame retardants (FRs), antibiotics, pesticides, pharmaceutical and personal care products (PPCPs), and commercial synthetic organic compounds, and the frequency of detection of these synthetic organic compounds in environmental pollution monitoring is gradually increasing [32]. If organic pollutants can be absorbed by organisms but are not easily metabolized and decomposed, they will accumulate in organisms and thus accumulate along the food chain [33]. Some pollutants can interfere with the endocrine system and are called environmental hormones or environmental endocrine disruptors. Environmental hormones can affect the physiological activities of reproduction, development, endocrine function, and immunity of organisms, and some environmental hormones can even induce cancer [34]. More than 70 environmental hormones have been found, most of which are organic substances. It is difficult to judge environmental hormones based only on their chemical structure. Zebrafish is sensitive to environmental hormones in water and is suitable to be used as model animal to detect and evaluate the degree of environmental hormones pollution in water ecology [35].
Some studies exposed adult zebrafish to flame retardants (tritoluene phosphate, toluene diphenyl phosphate) and insecticides for 15 days at exposure concentrations of 5 µL/L and 25 µL/L. During the exposure period, zebrafish motility and biological rhythm were quantitatively monitored to study the adaptability of zebrafish to different environments. The results showed that all organic pollutants in the experiments impaired the locomotor behavior of zebrafish, with little effect on the biological rhythm. The higher the concentration of organic pollutants, the more severe the locomotor damage was. The damaging effects of flame retardants and insecticides were similar [36]. There are scholars studied the toxicological effects of long-term (0–5 DPF (days post fertilization)) and short-term (4–5 DPF) exposures to estrone and bisphenol A and showed that long-term exposure to estrone reduced the locomotor activity of zebrafish. Long-term and short-term exposure to bisphenol A caused more significant bone abnormalities, motor behavior changes and transcriptome responses than estrone. Short-term exposure to bisphenol A at a concentration greater than 100 nmol/L can affect the expression of estrogen genes related to neurological diseases [37]. Therefore, it is of great significance to use zebrafish to improve the sensitivity of water environmental pollution detection and understand the toxic mechanism of pollutants in organisms for pollution monitoring and damage identification.
2.3. Evaluating the Toxicity of Nanoparticles and Microplastics on Zebrafish in Water Environments
With the wide application of nanomaterials, an increasing number of nanoparticles with different particle sizes have been released into the environment. Therefore, the potential ecotoxicity of nanomaterials has attracted increasing attention from ecotoxicologists. Nanotoxicology, a new branch of ecotoxicology, has become a new research focus, and its main research direction is to evaluate the safety of engineered nanostructures and nanodevices [38]. When zebrafish were used as the object of an aquatic ecotoxicology study of nanomaterials, it was found that nano-ZnO had concentration-dependent toxic effects on zebrafish embryos and larvae [39]. The accumulation of nanoparticles in organisms may be related to their particle size. After zebrafish zygotes were exposed to 200 nm and 60 nm fluorescent silica nanoparticles (FSNPs) for 96 h, it was found that FSNPs only adsorbed on the eggshell and did not enter the oil gland [40]. Zebrafish embryos are often used to evaluate the aquatic ecotoxicity of nanoparticles and can also be used as a model organism to study the bioenergetic impact of environmental pollutants on brain and muscle mitochondria [41]. Toxic gold nanoparticles (AUNPs) have wide application prospects in medical diagnosis and treatment. However, the metabolism and toxic effects of AUNPs in vertebrates are still unknown, and rodents are usually used as a vertebrate model for research [42]. Recently, researchers have used zebrafish expressing green fluorescent protein as an experimental vertebrate model to establish a rapid and real-time analytical technique for evaluating the toxic effects of nanoparticles [43]. Adult zebrafish have been used to test the toxicity of nanoparticles. The effect of titanium nanoparticles on the reproductive process of adult zebrafish showed that after 14 days of exposure, the adult fish could reproduce, but the number of viable embryos was lower than that of the experimental group in the 21-day recovery experiment. When the concentration of titanium nanoparticles was ≥10 mg/L, death and malformation of zebrafish embryos could occur [44].
Microplastics are widely present in the water environment, and they can be eaten by aquatic organisms and cause potential harm. The role of freshwater systems as the main source of microplastics in the marine environment cannot be ignored, so the pollution of water bodies by microplastics has become a global environmental threat [45]. Fish are a staple source of protein for humans, so microplastic pollution in fish and its potential effects are of particular concern. Microplastics in natural water will have a certain impact on fish after they are ingested [46,47]. At present, most research mainly focuses on the effects of microplastics on fish feeding, reproduction, and growth, and zebrafish account for a significant proportion of this kind of research. After ingesting microplastic particles, they not only accumulate in the body but also cause harm at the tissue, cellular, and molecular levels. For example, ingestion of microplastics can cause inflammatory responses in fish, alter metabolism, or damage the immune system [48,49]. Researchers exposed zebrafish larvae to polyethylene fragments (5, 50, 500 mg/L). The expression levels of apoptosis-related Casp9 and Casp3a and oxidative stress-related catalase on the 20th day of exposure were lower than those on the 10th day, indicating that oxidative stress had occurred in juvenile zebrafish [50]. Several studies have shown that there are three main factors affecting the accumulation of microplastics in aquatic animals: particle size of microplastics, exposure mode and exposure time, and shape and type of microplastics [51]. The particle size of microplastics is the key factor affecting bioaccumulation in zebrafish. The clearance and distribution of microplastics in tissues had size–effect and time–effect relationships (Figure 2).
3. Aquatic Ecotoxicological Evaluation Methods Based on Zebrafish
3.1. Aquatic Ecotoxicological Evaluation Based on the Physiological Toxicity of Zebrafish
The basic model of zebrafish for toxicological research is as follows: in first step, toxicant species and concentrations are monitored; in the second step, the biological behavior of zebrafish is evaluated, and data are obtained; in the third step, the phenomenon and data obtained are explained; and in the fourth step, the explanation is confirmed. After the explanation and confirmation from biological behavior to molecular mechanism, the behavior and molecule can be used as endpoints and biomarkers for subsequent monitoring [53]. The experimental design of the zebrafish model requires targeted design and selection of toxicant exposure, exposure duration, exposure pathway, and toxicant concentration on various aspects of the zebrafish development stages. The experimental construction of the water environment involves direct exposure of fish to the medium, including pure and mixed solutions such as wastewater and sewage. The range of endpoints and biomarkers is very wide, covering multiple levels of appearance and behavior, metabolism, and molecular structure, and each level has a number of observable indicators. the final purpose of the experiment is to establish a flowchart of the interaction effects between the poison and the organism, starting from the interaction mechanism of different links, and to apply this information to toxicological monitoring, environmental protection, healthcare, and other fields [54,55,56]. The detection of zebrafish exposure toxicity is generally divided into acute toxicity and chronic toxicity. Acute toxicity refers to short-term toxic effects on the body after exposure to a large dose of exogenous chemicals once or several times within 24 h. Lethal effects, developmental toxicity, behavioral abnormalities, and vital signs, including respiration and heart rate, are commonly observed endpoints of acute toxicity in zebrafish models [57]. The growth cycle of postnatal zebrafish can be divided into the embryonic stage (0–3 d), yolk sac stage (4–7 d), and juvenile stage (7–23 d), followed by the normal growth period. At 120 days after birth, zebrafish are considered sexually mature adult fish and can be used for mating and spawning. Exposure tests can be carried out at the different growth stages of zebrafish (Figure 3).
A 24 h continuous short-term exposure test can be performed in each zebrafish stage, or multiple short-term exposure tests within 24 h can be carried out in each stage after the embryonic stage. Through the short-term exposure test, the short-term acute semi-lethal concentration of the test substance is obtained and can lay the foundation for the long-term exposure test concentration [61]. Short-term embryo or juvenile stage experiments are widely used in preliminary toxicological evaluations. The commonly used observation endpoints in the general embryonic stage are egg condensation, somite formation, tail extension, heartbeat, pigmentation, hatching rate, and malformation rate [62]. The median lethal dose in each period is a vital index to evaluate and compare the toxicity of the test substance (Table 2). It is crucial for researchers to determine sublethal concentrations [63] Embryonic zebrafish, as a result of outer membrane barrier protection, can isolate some foreign substances in the external environment, and after the membrane is removed, zebrafish come into direct contact with the external subject, so tolerance in the embryonic period of the toxicity test may be slightly higher than that in the yolk sac stage; at the same time, the tolerance of the adult stage was generally higher than that of the embryonic stage, yolk sac stage, and juvenile stage [64]. Therefore, in acute toxicity tests of zebrafish, it is necessary to carry out toxicity evaluation at different periods according to the characteristics of the test materials and research requirements.
Chronic toxicity is a toxic effect that is gradually produced by long-term exposure to low concentrations of exogenous chemicals. In animal experiments, it refers to the damage caused by exposure to chemicals during most of the normal life cycle. Chronic toxicity experiments can more realistically simulate the toxic effects of environmental chemicals on organisms, and the research results have greater application value in environmental pollution control and nonaccidental injury identification [75]. Long-term exposure experiments in zebrafish can simulate the chronic poisoning process of animals. In long-term chronic exposure experiments, the exposure occurs from the embryonic stage to the adult stage (120–160 days after birth), and the exposed adult fish can mate with unexposed fish of the same age to spawn according to the research needs. Thus, the effects of the test objects on the offspring under long-term exposure conditions are obtained, and the observation indicators are generally the number of eggs laid, the survival rate, and the sex differentiation [76]. In long-term exposure tests, it is necessary to ensure the survival rate of embryos, juveniles, and adults throughout the exposure period, so the maximum concentration of the test substance is generally not higher than 10% of the EC50 in the short-term exposure test. In addition, because the environmentally detectable amount of the test substance is often low, the environmentally detectable amount of the test substance can be selected as the dose for long-term exposure experiments when studying the environmental effect of the test substance. Although the long-term exposure test costs much more manpower and material resources than the short-term acute exposure test, the obtained results can better reflect the environmental effects of the test substance. For example, long-term exposure studies have been carried out using this method in studies of perfluorooctane sulfonate (PFOS) and bisphenol A (BPA) [77,78].
3.2. Evaluation of Water Ecotoxicology Based on Molecular Biomarkers in Zebrafish
In the study of zebrafish, in addition to the typical developmental characteristics commonly used, hormone levels, gene levels, and protein levels are also included in subsequent studies. In environmental toxicology, the endocrine-disrupting effect of the tested substances on zebrafish is a crucial component of this kind of research [79]. Vitellogenin (VTG), a precursor of yolk, is synthesized in the liver of female fish. It is a specific estrogen analog that can stimulate the production of vitellogenin by binding to estrogen receptors. Since the vitellogenin gene is a more sensitive biomarker than the vitellogenin protein, detection of the expression of the vitellogenin gene Vtg1 in the liver of male fish is a common method to evaluate the estrogen effect of endocrine disruptors in the environment [80]. In addition, the reproductive ability of fish is generally regulated by the hypothalamic–pituitary–gonadal axis (HPG axis) and the liver, and gamete development and oocyte maturation are regulated using estradiol and testosterone [81]. The HPG axis is a novel analytical model that has been used to study the ecotoxicogenomics of endocrine disruptors and the effects of steroid hormone synthesis genes on the fertility of gonadal tissues [82]. Theoretically, any damage to the HPG axis by exogenous substances would have adverse effects on reproductive capacity. Therefore, it is common to study the changes in androgen and estrogen content in zebrafish, as well as changes in the expression of vital genes in different hormonal pathways [83,84]. Due to the small amount of blood available from individual zebrafish, tail vein blood is generally collected, and zebrafish hormones are determined using a dedicated ELISA kit. Because of the small variety of ELISA kits available for zebrafish assays and the high cost of research, the determination of changes in the expression of specific genes for corresponding hormones and proteins is widely used in zebrafish studies. For example, the hypothalamus, pituitary, gonad, and other tissues can be extracted, and the related hormone genes can be determined by real-time quantitative PCR [85,86,87]. Commonly used hormone genes include hormone receptor genes, gonadotropin-releasing hormone (GnRH) genes, and aromatic CYP genes. GnRHs are crucial hormones that control reproductive ability through the HPG axis. It exerts its corresponding effects by regulating the synthesis and release of gonadotropin-releasing hormone, which corresponds to GNRH class genes. Luteinizing hormone (LH) and follicle-stimulating hormone (FSH), the corresponding FSHβ and LSHβ genes, play a key role in regulating the expression of steroidogenic genes and hormone-dependent feedback [88]. The upregulation of the FSHβ gene in brain tissue indicates that related chemicals can interfere with sperm development [89]. The aromatic CYP19B gene has a feedback effect on the automatic regulation of estradiol stimulation in brain tissue, and upregulation of the CYP19B gene may also lead to an increase in estradiol levels in adult fish [90]. The aromatic CYP19A gene catalyzes the conversion of androgens to estrogens, and upregulation of CYP19A disrupts kinin homeostasis in the body [91]. CYP11A1 is a protease that regulates the conversion of cholesterol to progesterone, and CYP17 plays a main regulatory role in the conversion of 17α-hydroxyprogesterone to androstenedione in zebrafish testes [92]. In addition, hormone synthesis in female and male fish is related to the regulation of steroid hormone synthesis acute regulator protein (StAR), whose mRNA mainly controls the response mechanism of hormone genes [93]. This aspect of research is also seen in a large number of studies.
In zebrafish, changes in the activities of oxidoreductases and related enzymes usually show a dose-dependent effect with the stress of environmental toxic substances, which can be used as one of the evaluation criteria for toxic substances. Continuous exposure of zebrafish embryos to 500 μg/L polystyrene, 5 μg/L Cd, and their combination for 30 days inhibited the antioxidant system of zebrafish larvae, including the active enzyme activities of superoxide dismutase (SOD) and catalase (CAT). A large amount of ROS accumulated in the cells and tissues of juvenile fish, the cell membrane system showed peroxidation, juvenile fish showed a large number of spinal deformities and organ edema, and the juvenile survival rate decreased greatly [94]. Some studies have found that the insecticide cypermethrin significantly affected antioxidant enzyme activities in larval zebrafish in a concentration-dependent manner [95]. SOD activity and malondialdehyde (MDA) content were significantly increased in zebrafish embryos and larvae treated with 2,2′,4,4′-tetrabromodiphenyl ether (BDE-47), and there was a significant dose-response relationship between CAT activity and BDE-47 toxicity concentration [96].
3.3. Aquatic Ecotoxicological Evaluation Based on Genomic and Phenotypic Analysis of Zebrafish Toxicology
Toxicogenomics is a science that studies how organisms respond to environmental factors and toxicants at the genomic level. Using zebrafish as model animals, the toxicogenomics of the impact of bisphenol A on the fish’s early life was studied, and early exposure to bisphenol A in zebrafish was found to change the phenotype in the following ways: cardiac edema, skull deformity, swim bladder distention failure, reduced tactile responses, and dysplasia accompanied by spinal neurons. This indicates that there are many related signaling pathways involved in this process, including ephrin receptor-associated clathrin-mediated endocytosis and the deregulation of long-term potentiation of synapses. Six of 28 genes (ncl1, apoeb, mdm1, mycl1b, sp4, U1SNRNPBP homolog) were found to be sensitive to early BPA exposure and could be used as biomarkers for BPA toxicity in fish [77]. Liu et al. exposed zebrafish embryos to 30 μm NaHS solution and observed severe pericardial edema, spinal curvature and severe oxidative stress in zebrafish larvae exposed to H2S through fluorescent staining [13]. Transcriptome sequencing analysis showed that the antioxidant system of zebrafish larvae was seriously damaged. The apoptotic signaling pathway was highly activated, in which mitochondria are damaged and regulate apoptosis. Because zebrafish have mainly carbohydrates in their diet, their metabolic patterns are similar to those of humans, and zebrafish embryos exposed to 1.5% glucose solution for 10 days developed into zebrafish larvae with the typical characteristics of diabetes. Using the third generation of transcriptome sequencing analysis found that sugar disrupts the metabolic process of zebrafish larvae, causing the larval mortality rates to rise sharply [97]. Therefore, genomic and phenotypic analysis based on zebrafish toxicology can not only determine the health of aquatic ecosystems but also evaluate their effects on aquatic organisms and related toxicological response mechanisms.
4. Application of the Zebrafish Model in Environmental Toxicology Evaluation
4.1. Reproductive Toxicity
The occurrence of germ cells is the basis of the development and inheritance of organisms, and environmental pollutants often cause malformations or damage to the reproductive system. The reproductive toxicity of environmental pollutants can be detected by partial life cycle tests or whole life cycle tests in zebrafish. The commonly used detection indicators include vitellogenin, histological examination of the gonads and gonadal index, and sperm plasma membrane integrity [98,99]. Previous studies have shown that environmental estrogens can lead to feminization of male zebrafish, a change in the sex ratio, a decrease in male zebrafish semen volume, sperm total number, sperm quality, and other reproductive function damage [100]. Some studies suggest that with increasing treatment concentrations (2–10 ng/L ethinyl estradiol), ovarian development is more significantly inhibited [101]. In view of the many reproductive hazards of environmental chemicals, Chen et al. cloned the vitellogenin gene promoter, constructed the vitellogenin promoter–green fluorescent protein fusion gene, and established a transgenic fish line, ere-zvtg:gfp, that can directly monitor environmental estrogen pollution by microinjection of fertilized zebrafish eggs [102]. This method can roughly quantify the expression level of green fluorescent protein, detect a variety of environmental estrogen substances, and provide environmental warning for environmental estrogen. In addition, this model can also be used to study the mechanism of environmental estrogen action.
4.2. Teratogenic Toxicity of Embryos
The use of zebrafish embryos to detect the teratogenic effects of environmental chemicals has the characteristics of low cost, few influencing factors, good repeatability, easy operation, high sensitivity, and the ability to record multiple toxicity indicators (such as lethal, sublethal and teratogenic). Compared with the toxicity evaluation of other systems or target organs, zebrafish are relatively mature organisms for evaluation of the embryonic developmental toxicity of environmental pollutants. Due to the characteristics of in vitro fertilization, in vitro development, and easy operation, zebrafish are suitable for image-based detection, and a number of teratogenic toxicity indicators can be recorded, including embryonic development events such as cell movement in the gastrula stage, brain region formation, heartbeat, and blood circulation, which can be used as the end point of toxicological evaluation [103,104]. In addition, the teratogenic test cycle of zebrafish embryos is short, usually completed within 1 week, which is suitable for large-scale gene mutation screening and analysis [105].
For the observation of toxicity characterization of zebrafish embryos, generally four developmental time points, 24, 48, 72, and 96 hours post-fertilization (hpf), are selected, including spontaneous twitch (24 hpf), hatching rate (48 hpf), heartrate (72 hpf), and pigmentation, body axis bending and pericardial cysts (96 hpf). The time period beginning with 48 hpf is a key window of embryonic development and a crucial feature for the evaluation of teratogenic toxicity of embryos. One study found that for six kinds of triazole compounds with known developmental toxicity, the zebrafish embryo toxicity test was better than the European Center for Research on Alternative Experimental Methods rat whole embryo breeding and mouse embryonic stem cell test methods [106]. Studies have selected 31 positive teratogens to verify in the zebrafish embryo toxicity test, and 87% were consistent with the mammalian test results; only two false-positive and false-negative cases were observed [107]. Another study used zebrafish eggs immersed in 1.5% glucose solution for 10 days and found that high glucose toxicity had serious lethal and teratogenic effects on the development of zebrafish embryos to juveniles [97]. One study used the zebrafish embryo toxicity test to detect 27 compounds for verification in the in vitro substitution test, and the EC50, LC50 and teratogenic index (TI) data were obtained through the drug efficacy curve. The results showed that the sensitivity of the zebrafish embryo toxicity test was 72%, and the specificity was 100% [108]. In a study on the correlation between teratogenic effects on zebrafish embryos and teratogenic effects on mammals, exposure of zebrafish embryos to valproic acid and its nine analogs showed that sublethal dose exposure of four compounds could cause microphthalmia and craniofacial deformities in zebrafish, which were related to neural tube defects in mammals [109].
4.3. Neurodevelopmental Toxicity and Behavioral Changes
The zebrafish is an excellent model for high-throughput screening of environmental chemical neurotoxicity, which can be used to detect the potential toxicity of environmental chemicals on nervous system development. Juvenile fish can swim freely after 5 days of fertilization because the brain tissue of juvenile fish has differentiated into telencephalon, diencephalon, mesencephalon, hindbrain, and rhomboencephalon segments, which allows for evaluation of the movement, learning and memory behavior of the photoelectric stimulation response indicators [110,111]. In addition, similar to the mammalian model, biochemical indicators, morphological changes, neuronal necrosis and apoptosis can be used to evaluate the neurodevelopmental toxicity of environmental chemicals on zebrafish embryos. Exposure of zebrafish fertilized eggs to different concentrations of alcohol for 4–24 h after fertilization resulted in significant changes in ultrahigh pressure, dopamine and serotonin and their metabolites DOPAC and 5-HIAA in the brain [112]. Cd exposure can induce brain hypoplasia, such as cerebellar malformations, in zebrafish during early embryonic development, which affects the orientation and differentiation of neural progenitor cells of the brain. The number of sensory ganglia was reduced with developmental defects in forebrain and hindbrain interneurons [113]. A recent study found that zebrafish embryos exposed to 800 µmol/L 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, pyridine (MPTP). Two days later, in situ hybridization of zebrafish brain tissue showed that the exposure of zebrafish embryos to MPTP specifically damaged dopaminergic neurons [114]. Continuous exposure of zebrafish embryos to nonteratogenic concentrations (≤4 µmol/L) of aristolochic acid could lead to impaired expression of genes related to visual perception in larval zebrafish [115]. In the study of the neurotoxicity mechanism of titanium dioxide nanoparticles (nano-TiO2), exposure of zebrafish embryos to a concentration gradient of particles (0.01, 0.1, 1.0 mg/L) showed that microparticles can adversely affect the growth of the axons of zebrafish embryonic motor neurons. Gene expression detection confirmed that microparticle exposure affects the expression of genes related to neuron genesis and axon growth, and high concentrations of microparticles can reduce the length and weight of zebrafish larvae [116].
Neurodevelopmental toxicity often leads to behavioral changes. As an observational approach, behavioral changes have the advantages of many observable items and simple operation. However, the high complexity of behavioral reactions means there is a current lack of normative standards for the operation and evaluation of behavioral experiments, which makes the evaluation of the validity of the results difficult. Zebrafish that continuously ingested large amounts of sugars in the environment developed diabetic symptoms accompanied by depressive behavior, and increasing their aerobic exercise could significantly change depressive behavior [117]. The experimental results of the impact of pesticides containing glyphosate on water environment ecology showed that short-term (2 weeks) exposure to glyphosate at environmental-related concentrations (0.3, 3 µg/L) could reduce exploration behavior and change the social behavior of adult zebrafish, which further adversely affected the survival of zebrafish [118]. A study on the interference of diazepam residues in the water environment on the behavior of zebrafish showed that the courtship-related experimental behaviors of female and male zebrafish were significantly affected after 21 days of exposure to diazepam at 21 µg/L and 120 µg/L (both in the sublethal concentration range) [119].
4.4. Cardiac and Cardiovascular Developmental Toxicity
Zebrafish embryos have obvious advantages in the toxicological evaluation of cardiac development after exposure to environmental chemicals. The zebrafish heart is similar to the human embryonic heart, with one atrium and one ventricle and a valve between them. Regular heartbeat begins at 36 hpf, and changes in heart morphology and rhythm, such as heartbeat, blood vessel morphology, and cell activity in blood vessels, can be directly observed by microscopy, or even on clear photographic images. All these factors provide great convenience for toxicity evaluation and toxicological research of environmental chemicals [120]. In recent years, transgenic zebrafish with gene promoters only expressed in the cardiovascular system have been successfully established and can be used for real-time monitoring and quantitative and qualitative evaluation of cardiovascular damage caused by specific environmental chemical toxicity [121]. Zebrafish embryos are particularly suitable for toxicological evaluation of cardiac development by environmental chemicals. Because the embryo obtains oxygen through diffusion in the first week, embryos with cardiovascular development defects can obtain oxygen to survive until oxygen diffusion in the water body has difficulty maintaining the needs of embryonic development, which is convenient for the dynamic monitoring of embryonic heart development. Studies used the antimicrobial pesticide mancozeb (MZ) to study the developmental toxicity of zebrafish embryonic heart. Heart tissue sections showed that MZ has a series of toxic effects on the heart development of zebrafish embryos, such as pericardial edema, slow heart rate, cardiac congestion, and myocardial fibrosis. This is consistent with the characteristics of toxic effects in mammalian cardiac development [122]. When adult zebrafish were exposed to benzo(a)pyrene, the ventricular heart rate was reduced, leading to cardiopulmonary injury in zebrafish [123]. Studies have verified the zebrafish cardiotoxicity test and found that the sensitivity of the zebrafish model was 78%–100%, and its specificity was 77%–100%, which met the requirements of the European Center for Research on Alternative Experimental Methods for alternative tests (“sufficient” (65–75% predictivity); “good” (75–85% predictivity)), indicating that the zebrafish model is correlated with the mammalian model and has the potential to evaluate cardiotoxicity induced by environmental chemicals [124].
The molecular pathways regulating the hematopoietic system in zebrafish are quite conserved, especially in the early development process of the cardiovascular system, which is very similar to that of humans. Mutants with defective cardiovascular systems can still survive for a long time, providing extremely favorable conditions for studying the developmental toxicity of the cardiovascular system caused by environmental pollutants [120]. The effects of seven known cardiotoxic drugs, such as aspirin, and two noncardiotoxic drugs, gentamicin sulfate and tetracycline hydrochloride, on the cardiovascular development of zebrafish embryos and juvenile fish were detected with six phenotypic endpoints, including heart rate, cardiac rhythm, and thrombus. The results showed that the seven drugs with known toxicity caused various cardiovascular developmental defects, such as tachycardia, bradycardia, and atrioventricular block, in zebrafish larvae, while gentamycin sulfate and tetracycline hydrochloride did not show cardiovascular toxicity in zebrafish [125].
4.5. Hepatorenal Toxicity
The liver is the main metabolic organ of the body, has a crucial function in the body and is easily damaged by the toxicity of various chemicals, thus affecting the normal operation of the body. The response of the zebrafish liver to chemicals in the early developmental stage is similar to that of humans, so it is an ideal model for studying chemical hepatotoxicity [126]. Zebrafish have been used as model organisms to study the toxic effects of octocrylene (OC). The results showed that octocrylene (OC) had toxic effects on zebrafish and its embryos, and the liver was a vital target organ of polychlorinated biphenyls [127]. Another study investigated the hepatotoxicity of tris (1,3-dichloro-2-propyl) phosphate (TDCIPP) in juvenile zebrafish using liver fluorescence transgenic zebrafish developing 72 hpf. The results showed that TDCIPP was hepatotoxic to juvenile zebrafish [128]. Zhang also found that the paralytic shellfish toxin secreted by filamentous algae has a toxic effect on the liver of zebrafish. After exposure, the juvenile zebrafish showed abnormal liver morphology, darker liver color, obvious liver atrophy, and other toxic reactions [129].
The morphology, physiology, and function of the kidney are highly conserved among different vertebrates. Using zebrafish embryos as a model to study the mechanism, degree of injury, detoxification, and recovery process of environmental pollutants on kidney toxicity is helpful for researchers to have a deep understanding of renal injury and renal failure to better protect human health [130]. Ding et al. treated zebrafish embryos with 10 mg/L aristolochic acid (AA) for 24 h. Under a microscope, abnormal morphological phenomena, such as anterior renal canal, cystic degeneration of the anterior renal arch and glomerular atrophy, were observed, which proved that AA had a toxic effect on embryonic kidney development [131].
4.6. Endocrine-Disrupting Toxicity
The thyroid gland is a vital hub of hormone regulation, and the hypothalamic–pituitary–thyroid axis (HPTA) is a key factor in maintaining body homeostasis. A toxicity study of the antibiotic oxytetracycline (OTC) confirmed that the environmental concentration OTC significantly increased plasma levels of triiodothyronine in adult zebrafish [132,133]. A study on the effect of bisphenol fluorene, which is used in the production of plastic cups, on HPTA showed that bisphenol fluorene could affect the level of thyroid hormone by affecting the expression of HPTA-related genes [134].
4.7. Environmental Carcinogenicity
Zebrafish are also recognized as model organisms for cancer research. Due to the high conservation between zebrafish tumor models and human tumors, many human tumor-related pathways have homologs in zebrafish tumors. Zebrafish were used in tumor research as early as 2005. Recently, the emergence of transgenic zebrafish and tumor transplant zebrafish has made it a potential animal tumor model. Zebrafish are more similar to humans than invertebrate nematodes and fruit flies and are used for rapid screening of tumor proliferation, division, metastasis, and antineoplastic drugs in vivo [135]. Many carcinogens in the environment can induce tumors in zebrafish, and the liver is the organ most prone to tumors, because the liver keeps growing throughout the lifespan [136]. DMBA (dimethyl benzanthracene) and MNNG (N-methyl-N’-nitro-N-nitrosoguanidine) can induce tumors not only in the digestive system but also in the immune system and nervous system [137]. Zebrafish infected with pseudocapillary nematodes were fed DMBA, and the incidence of tumors was 50% higher than that of uninfected zebrafish [138]. Spitsbergen et al. evaluated the effects of carcinogens with different structures, such as N-nitroso dimethylamine, aflatoxin B, N-methyl-N-nitro-N-nitrosoguanidine, and 7,12-dimethyl-phenylanthracene, on different developmental stages of wild-type zebrafish. When zebrafish were fed with feed containing carcinogens for 3–9 months, tumors in the epithelium, stroma, nerve tissue occurred in embryos or young zebrafish, among which liver tumors were the most common [139,140]. Zebrafish can also provide an effective and economical research system for the evaluation of the carcinogenicity of industrial sewage, the study of tumorigenesis mechanisms and the screening of new anticancer drugs.
5. Conclusions and Perspectives
With the gradual development and utilization of water resources, the environmental pollutants in aquatic chemicals are becoming more and more complex, and the water ecological pollution is becoming more and more serious. The effects of environmental chemicals on and signaling pathways of zebrafish and higher mammals are basically the same. Recently, it has been considered that the changes caused by environmental chemicals can be used as markers of characteristic toxic reactions. It is an effective tool for predicting the toxicity of chemical substances and can also complement the mammalian testing system as a first step in modeling the toxicity of pollutants to spinal animals and environmental monitoring. Zebrafish have received increasing attention from toxicologists because of their excellent biological characteristics and have become one of the most vital model organisms in environmental toxicology, ecotoxicology, drug toxicology, and other toxicological research. Zebrafish have been recommended as a new alternative animal by the European Experimental Substitution Research Center and have become an efficient and universal environmental toxicology model organism in the 21st century. They can be used to carry out multidimensional ecological risk assessments and have great value in optimizing and reducing mammalian in vivo experiments and environmental chemical toxicity mechanism tests. This paper briefly introduces the application of zebrafish as an animal model in the study of aquatic ecotoxicology that can be used to evaluate the toxicological effects of metal elements, organic pollutants, nanoparticles, and microplastics in aquatic ecology. In addition, the developmental process of zebrafish can also be used in aquatic ecotoxicological assessment, using typical developmental characteristics, biomarkers, and genome levels to study the reproductive toxicity, embryonic teratogenic toxicity, and liver and kidney toxicity of environmental pollutants in zebrafish (Figure 4).
Although wild-type zebrafish are widely used in toxicology research, they also have many shortcomings. Specifically, zebrafish lack some of the mammalian organs such as the lung, prostate, skin, and mammary gland, and so the toxic effects of environmental pollution on humans and other mammals cannot fully be predicted. With the development of genomics and other technologies, the use of transgenic zebrafish with specific phenotypes for different kinds of pollutants to study the toxicology of environmental pollutants has become a new focus of ecotoxicology research. At the same time, the database of molecular biomarkers of different pollutants is constantly expanded to provide more data support for environmental monitoring and early warning of pollutants. In the future, the extensive application of traditional techniques and instruments and the rapid development of microanalytical techniques will continue to tap the potential of zebrafish in toxicity analysis, so as to make the database of pollutant-induced zebrafish toxicity and toxicity phenotypes more comprehensive. The continuous development of various specific transgenic zebrafish will provide a more convenient, reliable, flexible and cost-effective vertebrate model for chemists and biologists. Furthermore, more specific phenotypes of zebrafish will also promote the research process of aquatic ecotoxicology, so as to help humans solve the problem of water pollution more quickly.
Author Contributions
H.L. and Y.L. contributed equally. H.L. and Y.L.: investigation, data curation, writing—original draft, writing—review and editing. Q.C.: software, supervision, validation. L.J. and R.P.: conceptualization, writing—review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Natural Science Foundation of Zhejiang Province (Grant numbers: LQ20C020003).
Data Availability Statement
No new data were created.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
OECD: the Organization for Economic Cooperation and Development; ISO: the International Organization for Standardization; HSP70: heat shock protein 70; GFP: green fluorescent protein; ROS: reactive oxygen species; SGR: specific growth rate; E2: 17β-estradiol; VTG: vitellogenin; GSI: gonadosomatic index; MgHP: magnesium–hesperidin complex; CAT: catalase; CYP450: cytochrome P450; SOD: superoxide dismutase; DAO: diamine oxidase; AChE: acetylcholinesterase; ATP: adenosine triphosphate; MDA: malondialdehyde; ssDNA: single-stranded DNA; DA: dopamine; GABA: γ-aminobutyric acid; MREs: metal response elements; REEs: rare earth elements; EDCs: endocrine-disrupting chemicals; FRs: flame retardants; PPCPs: pharmaceutical and personal care products; DPF: days post-fertilization; FSNPs: fluorescent silica nanoparticles; AUNPs: gold nanoparticles; 6:2 FTAB: 6:2 fluorotelomer sulfonamide alkylbetaine; MB-5G: Maxilon Blue 5G; RB-203: Reactive Blue 203; PFOS: perfluorooctane sulfonate; BPA: bisphenol A; HPG axis: hypothalamic–pituitary–gonadal axis; GnRH: gonadotropin-releasing hormone; LH: luteinizing hormone; FSH: follicle-stimulating hormone; StAR: synthesis acute regulator protein; BDE-47: 2,2’,4,4’-tetrabromodiphenyl ether; TI: teratogenic index; MPTP: 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine–pyridine; MZ: mancozeb; OC: octocrylene; TDCIPP: tris(1,3-dichloro-2-propyl) phosphate; AA: aristolochic acid; HPTA: hypothalamic–pituitary–thyroid axis; OTC: oxytetracycline; DMBA: dimethyl benzanthracene; MNNG: N-methyl-N’-nitro-N-nitrosoguanidine.
References
- Ledón-Rettig, C.C.; Pfennig, D.W. Emerging Model Systems in Eco-Evo-Devo: The Environmentally Responsive Spadefoot Toad. Evol. Dev. 2011, 13, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Huang, X.; Wu, Q.; Shi, J.; Zhang, X.; Ouyang, L.; Crump, D.; Zhang, X.; Zhang, R. Acute Toxicity of Polychlorinated Diphenyl Ethers (PCDEs) in Three Model Aquatic Organisms (Scenedesmus Obliquus, Daphnia Magna, and Danio Rerio) of Different Trophic Levels. Sci. Total Environ. 2022, 805, 150366. [Google Scholar] [CrossRef] [PubMed]
- Bambino, K.; Chu, J. Zebrafish in Toxicology and Environmental Health. Curr. Top. Dev. Biol. 2017, 124, 331–367. [Google Scholar] [CrossRef] [PubMed]
- MacRae, C.A.; Peterson, R.T. Zebrafish as Tools for Drug Discovery. Nat. Rev. Drug Discov. 2015, 14, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Zanandrea, R.; Bonan, C.D.; Campos, M.M. Zebrafish as a Model for Inflammation and Drug Discovery. Drug Discov. Today 2020, 25, 2201–2211. [Google Scholar] [CrossRef]
- Raldúa, D.; Piña, B. In Vivo Zebrafish Assays for Analyzing Drug Toxicity. Expert Opin. Drug Metab. Toxicol. 2014, 10, 685–697. [Google Scholar] [CrossRef]
- Kanungo, J.; Cuevas, E.; Ali, S.F.; Paule, M.G. Zebrafish Model in Drug Safety Assessment. Curr. Pharm. Des. 2014, 20, 5416–5429. [Google Scholar] [CrossRef]
- Lantz-McPeak, S.; Guo, X.; Cuevas, E.; Dumas, M.; Newport, G.D.; Ali, S.F.; Paule, M.G.; Kanungo, J. Developmental Toxicity Assay Using High Content Screening of Zebrafish Embryos. J. Appl. Toxicol. 2015, 35, 261–272. [Google Scholar] [CrossRef]
- Bauer, B.; Mally, A.; Liedtke, D. Zebrafish Embryos and Larvae as Alternative Animal Models for Toxicity Testing. Int. J. Mol. Sci. 2021, 22, 13417. [Google Scholar] [CrossRef]
- Audira, G.; Siregar, P.; Chen, J.-R.; Lai, Y.-H.; Huang, J.-C.; Hsiao, C.-D. Systematical Exploration of the Common Solvent Toxicity at Whole Organism Level by Behavioral Phenomics in Adult Zebrafish. Environ. Pollut. 2020, 266 Pt 1, 115239. [Google Scholar] [CrossRef]
- Krauss, J.; Astrinidis, P.; Astrinides, P.; Frohnhöfer, H.G.; Walderich, B.; Nüsslein-Volhard, C. Transparent, a Gene Affecting Stripe Formation in Zebrafish, Encodes the Mitochondrial Protein Mpv17 That Is Required for Iridophore Survival. Biol. Open 2013, 2, 703–710. [Google Scholar] [CrossRef]
- Blechinger, S.R.; Warren, J.T.; Kuwada, J.Y.; Krone, P.H. Developmental Toxicology of Cadmium in Living Embryos of a Stable Transgenic Zebrafish Line. Environ. Health Perspect 2002, 110, 1041–1046. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Q.; Li, Y.; Bi, L.; Lin, S.; Ji, H.; Sun, D.; Jin, L.; Peng, R. Hydrogen Sulfide-Induced Oxidative Stress Mediated Apoptosis via Mitochondria Pathway in Embryo-Larval Stages of Zebrafish. Ecotoxicol. Environ. Saf. 2022, 239, 113666. [Google Scholar] [CrossRef] [PubMed]
- Domingues, I.; Oliveira, R.; Lourenço, J.; Grisolia, C.K.; Mendo, S.; Soares, A.M.V.M. Biomarkers as a Tool to Assess Effects of Chromium (VI): Comparison of Responses in Zebrafish Early Life Stages and Adults. Comp. Biochem. Physiol. C Toxicol. Pharm. 2010, 152, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Richetti, S.K.; Rosemberg, D.B.; Ventura-Lima, J.; Monserrat, J.M.; Bogo, M.R.; Bonan, C.D. Acetylcholinesterase Activity and Antioxidant Capacity of Zebrafish Brain Is Altered by Heavy Metal Exposure. Neurotoxicology 2011, 32, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Olson, A.J.; Cyphers, T.; Gerrish, G.; Belby, C.; King-Heiden, T.C. Using Morphological, Behavioral, and Molecular Biomarkers in Zebrafish to Assess the Toxicity of Lead-Contaminated Sediments from a Retired Trapshooting Range within an Urban Wetland. J. Toxicol. Environ. Health A 2018, 81, 924–938. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, A.-P.; Li, W.-F.; Shi, R.; Jin, H.-T.; Wei, J.-F. Sensitive Biomarkers Identification for Differentiating Cd and Pb Induced Toxicity on Zebrafish Embryos. Environ. Toxicol. Pharmacol. 2017, 56, 340–349. [Google Scholar] [CrossRef]
- Zheng, J.-L.; Peng, L.-B.; Xia, L.-P.; Li, J.; Zhu, Q.-L. Effects of Continuous and Intermittent Cadmium Exposure on HPGL Axis, GH/IGF Axis and Circadian Rhythm Signaling and Their Consequences on Reproduction in Female Zebrafish: Biomarkers Independent of Exposure Regimes. Chemosphere 2021, 282, 130879. [Google Scholar] [CrossRef]
- Bonomo, M.M.; Fernandes, J.B.; Carlos, R.M.; Fernandes, M.N. Biochemical and Genotoxic Biomarkers and Cell Cycle Assessment in the Zebrafish Liver (ZF-L) Cell Line Exposed to the Novel Metal-Insecticide Magnesium-Hespiridin Complex. Chemosphere 2020, 250, 126416. [Google Scholar] [CrossRef]
- Domingues, I.; Oliveira, R.; Musso, C.; Cardoso, M.; Soares, A.M.V.M.; Loureiro, S. Prochloraz Effects on Biomarkers Activity in Zebrafish Early Life Stages and Adults. Environ. Toxicol. 2013, 28, 155–163. [Google Scholar] [CrossRef]
- Jeon, H.-J.; Lee, Y.-H.; Kim, M.-J.; Choi, S.-D.; Park, B.-J.; Lee, S.-E. Integrated Biomarkers Induced by Chlorpyrifos in Two Different Life Stages of Zebrafish (Danio Rerio) for Environmental Risk Assessment. Environ. Toxicol. Pharmacol. 2016, 43, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, R.; McDonough, S.; Ladewig, J.C.L.; Soares, A.M.V.M.; Nogueira, A.J.A.; Domingues, I. Effects of Oxytetracycline and Amoxicillin on Development and Biomarkers Activities of Zebrafish (Danio Rerio). Environ. Toxicol. Pharmacol. 2013, 36, 903–912. [Google Scholar] [CrossRef] [PubMed]
- Qiao, R.; Sheng, C.; Lu, Y.; Zhang, Y.; Ren, H.; Lemos, B. Microplastics Induce Intestinal Inflammation, Oxidative Stress, and Disorders of Metabolome and Microbiome in Zebrafish. Sci. Total Environ. 2019, 662, 246–253. [Google Scholar] [CrossRef]
- Sarasamma, S.; Audira, G.; Siregar, P.; Malhotra, N.; Lai, Y.-H.; Liang, S.-T.; Chen, J.-R.; Chen, K.H.-C.; Hsiao, C.-D. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. Int. J. Mol. Sci. 2020, 21, 1410. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshini, E.; Priyadarshini, S.S.; Pradhan, N. Heavy Metal Resistance in Algae and Its Application for Metal Nanoparticle Synthesis. Appl. Microbiol. Biotechnol. 2019, 103, 3297–3316. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Pandit, S.; Mokkapati, V.R.S.S.; Garg, A.; Ravikumar, V.; Mijakovic, I. Gold Nanoparticles in Diagnostics and Therapeutics for Human Cancer. Int. J. Mol. Sci. 2018, 19, 1979. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and the Environment. Exp. Suppl. 2012, 101, 133–164. [Google Scholar] [CrossRef] [PubMed]
- Lee, O.; Green, J.M.; Tyler, C.R. Transgenic Fish Systems and Their Application in Ecotoxicology. Crit. Rev. Toxicol. 2015, 45, 124–141. [Google Scholar] [CrossRef]
- Zhao, Y.; Liang, J.; Meng, H.; Yin, Y.; Zhen, H.; Zheng, X.; Shi, H.; Wu, X.; Zu, Y.; Wang, B.; et al. Rare Earth Elements Lanthanum and Praseodymium Adversely Affect Neural and Cardiovascular Development in Zebrafish (Danio Rerio). Environ. Sci. Technol. 2021, 55, 1155–1166. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Q.; Li, Y.; Bi, L.; Jin, L.; Peng, R. Toxic Effects of Cadmium on Fish. Toxics 2022, 10, 622. [Google Scholar] [CrossRef]
- Min, E.K.; Lee, A.N.; Lee, J.-Y.; Shim, I.; Kim, P.; Kim, T.-Y.; Kim, K.-T.; Lee, S. Advantages of Omics Technology for Evaluating Cadmium Toxicity in Zebrafish. Toxicol. Res. 2021, 37, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Olisah, C.; Adams, J.B.; Rubidge, G. The State of Persistent Organic Pollutants in South African Estuaries: A Review of Environmental Exposure and Sources. Ecotoxicol. Environ. Saf. 2021, 219, 112316. [Google Scholar] [CrossRef]
- Li, C.; Yang, L.; Shi, M.; Liu, G. Persistent Organic Pollutants in Typical Lake Ecosystems. Ecotoxicol. Environ. Saf. 2019, 180, 668–678. [Google Scholar] [CrossRef] [PubMed]
- Brody, J.G.; Rudel, R.A. Environmental Pollutants and Breast Cancer. Environ. Health Perspect. 2003, 111, 1007–1019. [Google Scholar] [CrossRef] [PubMed]
- Spaan, K.; Haigis, A.-C.; Weiss, J.; Legradi, J. Effects of 25 Thyroid Hormone Disruptors on Zebrafish Embryos: A Literature Review of Potential Biomarkers. Sci. Total Environ. 2019, 656, 1238–1249. [Google Scholar] [CrossRef] [PubMed]
- Ren, Z.; Poopal, R.-K.; Ramesh, M. Synthetic Organic Chemicals (Flame Retardants and Pesticides) with Neurotoxic Potential Induced Behavioral Impairment on Zebrafish (Danio Rerio): A Non-Invasive Approach for Neurotoxicology. Environ. Sci. Pollut. Res. Int. 2021, 28, 37534–37546. [Google Scholar] [CrossRef]
- Wu, C.-C.; Shields, J.N.; Akemann, C.; Meyer, D.N.; Connell, M.; Baker, B.B.; Pitts, D.K.; Baker, T.R. The Phenotypic and Transcriptomic Effects of Developmental Exposure to Nanomolar Levels of Estrone and Bisphenol A in Zebrafish. Sci. Total Environ. 2021, 757, 143736. [Google Scholar] [CrossRef]
- Oberdörster, G.; Oberdörster, E.; Oberdörster, J. Nanotoxicology: An Emerging Discipline Evolving from Studies of Ultrafine Particles. Environ. Health Perspect. 2005, 113, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Wang, S.; Wu, Y.; You, H.; Lv, L. Acute ZnO Nanoparticles Exposure Induces Developmental Toxicity, Oxidative Stress and DNA Damage in Embryo-Larval Zebrafish. Aquat. Toxicol. 2013, 136–137, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Fent, K.; Weisbrod, C.J.; Wirth-Heller, A.; Pieles, U. Assessment of Uptake and Toxicity of Fluorescent Silica Nanoparticles in Zebrafish (Danio Rerio) Early Life Stages. Aquat. Toxicol. 2010, 100, 218–228. [Google Scholar] [CrossRef]
- Bourdineaud, J.-P.; Rossignol, R.; Brèthes, D. Zebrafish: A Model Animal for Analyzing the Impact of Environmental Pollutants on Muscle and Brain Mitochondrial Bioenergetics. Int. J. Biochem. Cell Biol. 2013, 45, 16–22. [Google Scholar] [CrossRef]
- Boisselier, E.; Astruc, D. Gold Nanoparticles in Nanomedicine: Preparations, Imaging, Diagnostics, Therapies and Toxicity. Chem. Soc. Rev. 2009, 38, 1759–1782. [Google Scholar] [CrossRef]
- Pan, Y.; Leifert, A.; Graf, M.; Schiefer, F.; Thoröe-Boveleth, S.; Broda, J.; Halloran, M.C.; Hollert, H.; Laaf, D.; Simon, U.; et al. High-Sensitivity Real-Time Analysis of Nanoparticle Toxicity in Green Fluorescent Protein-Expressing Zebrafish. Small 2013, 9, 863–869. [Google Scholar] [CrossRef]
- Ramsden, C.S.; Henry, T.B.; Handy, R.D. Sub-Lethal Effects of Titanium Dioxide Nanoparticles on the Physiology and Reproduction of Zebrafish. Aquat. Toxicol. 2013, 126, 404–413. [Google Scholar] [CrossRef]
- Landrigan, P.J.; Stegeman, J.J.; Fleming, L.E.; Allemand, D.; Anderson, D.M.; Backer, L.C.; Brucker-Davis, F.; Chevalier, N.; Corra, L.; Czerucka, D.; et al. Human Health and Ocean Pollution. Ann. Glob. Health 2020, 86, 151. [Google Scholar] [CrossRef]
- Ajith, N.; Arumugam, S.; Parthasarathy, S.; Manupoori, S.; Janakiraman, S. Global Distribution of Microplastics and Its Impact on Marine Environment-a Review. Environ. Sci. Pollut. Res. Int. 2020, 27, 25970–25986. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, J.; Zang, L.; Nishimura, N.; Shimada, Y. Zebrafish: An Emerging Model to Study Microplastic and Nanoplastic Toxicity. Sci. Total Environ. 2020, 728, 138707. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Duan, Z.; Wu, Y.; Wang, Y.; Zhang, H.; Shi, Y.; Zhang, H.; Wei, Y.; Sun, H. Immunotoxicity Responses to Polystyrene Nanoplastics and Their Related Mechanisms in the Liver of Zebrafish (Danio Rerio) Larvae. Environ. Int. 2022, 161, 107128. [Google Scholar] [CrossRef]
- Yang, H.; Lai, H.; Huang, J.; Sun, L.; Mennigen, J.A.; Wang, Q.; Liu, Y.; Jin, Y.; Tu, W. Polystyrene Microplastics Decrease F-53B Bioaccumulation but Induce Inflammatory Stress in Larval Zebrafish. Chemosphere 2020, 255, 127040. [Google Scholar] [CrossRef] [PubMed]
- Karami, A.; Groman, D.B.; Wilson, S.P.; Ismail, P.; Neela, V.K. Biomarker Responses in Zebrafish (Danio Rerio) Larvae Exposed to Pristine Low-Density Polyethylene Fragments. Environ. Pollut. 2017, 223, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Cózar, A.; Martí, E.; Duarte, C.M.; García-de-Lomas, J.; van Sebille, E.; Ballatore, T.J.; Eguíluz, V.M.; González-Gordillo, J.I.; Pedrotti, M.L.; Echevarría, F.; et al. The Arctic Ocean as a Dead End for Floating Plastics in the North Atlantic Branch of the Thermohaline Circulation. Sci. Adv. 2017, 3, e1600582. [Google Scholar] [CrossRef] [PubMed]
- Pei, X.; Heng, X.; Chu, W. Polystyrene Nano/Microplastics Induce Microbiota Dysbiosis, Oxidative Damage, and Innate Immune Disruption in Zebrafish. Microb. Pathog. 2022, 163, 105387. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Inoue, A.; Sasagawa, S.; Koiwa, J.; Kawaguchi, K.; Kawase, R.; Maruyama, T.; Kim, S.; Tanaka, T. Using Zebrafish in Systems Toxicology for Developmental Toxicity Testing. Congenit. Anom. 2016, 56, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Dooley, K.; Zon, L.I. Zebrafish: A Model System for the Study of Human Disease. Curr. Opin. Genet. Dev. 2000, 10, 252–256. [Google Scholar] [CrossRef]
- Tran, S.; Chow, H.; Tsang, B.; Facciol, A.; Gandhi, P.; Desai, P.; Gerlai, R. Zebrafish Are Able to Detect Ethanol in Their Environment. Zebrafish 2017, 14, 126–132. [Google Scholar] [CrossRef]
- Williams, T.D.; Mirbahai, L.; Chipman, J.K. The Toxicological Application of Transcriptomics and Epigenomics in Zebrafish and Other Teleosts. Brief. Funct. Genom. 2014, 13, 157–171. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, R.; Jin, Y. Differential Responses of Larval Zebrafish to the Fungicide Propamocarb: Endpoints at Development, Locomotor Behavior and Oxidative Stress. Sci. Total Environ. 2020, 731, 139136. [Google Scholar] [CrossRef]
- Hu, H.; Su, M.; Ba, H.; Chen, G.; Luo, J.; Liu, F.; Liao, X.; Cao, Z.; Zeng, J.; Lu, H.; et al. ZIF-8 Nanoparticles Induce Neurobehavioral Disorders through the Regulation of ROS-Mediated Oxidative Stress in Zebrafish Embryos. Chemosphere 2022, 305, 135453. [Google Scholar] [CrossRef]
- Li, X.; Zhou, S.; Qian, Y.; Xu, Z.; Yu, Y.; Xu, Y.; He, Y.; Zhang, Y. The Assessment of the Eco-Toxicological Effect of Gabapentin on Early Development of Zebrafish and Its Antioxidant System. RSC Adv. 2018, 8, 22777–22784. [Google Scholar] [CrossRef]
- Yin, J.; Wang, A.-P.; Li, W.-F.; Shi, R.; Jin, H.-T.; Wei, J.-F. Time-Response Characteristic and Potential Biomarker Identification of Heavy Metal Induced Toxicity in Zebrafish. Fish. Shellfish. Immunol. 2018, 72, 309–317. [Google Scholar] [CrossRef]
- Cao, F.; Liu, X.; Wang, C.; Zheng, M.; Li, X.; Qiu, L. Acute and Short-Term Developmental Toxicity of Cyhalofop-Butyl to Zebrafish (Danio Rerio). Environ. Sci. Pollut. Res. Int. 2016, 23, 10080–10089. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, F.S.; Souza, T.M.; Vieira, L.R.; Marchi, F.C.; Nascimento, A.P.; Farias, D.F. Toxicity Testing of Pesticides in Zebrafish-a Systematic Review on Chemicals and Associated Toxicological Endpoints. Environ. Sci. Pollut. Res. Int. 2020, 27, 10185–10204. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Ibrahim, K.E.; Hussain, S.A.; Abdel-Daim, M.M. Role of ROS Generation in Acute Genotoxicity of Azoxystrobin Fungicide on Freshwater Snail Lymnaea luteola L. Environ. Sci. Pollut. Res. 2021, 28, 5566–5574. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Duan, X.; Zhao, S.; Wang, X.; Wang, J.; Liu, Y.; Peng, Y.; Gong, Z.; Wang, L. Barrier Function of Zebrafish Embryonic Chorions against Microplastics and Nanoplastics and Its Impact on Embryo Development. J. Hazard. Mater. 2020, 395, 122621. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.-Y.; Yu, X.-Y.; Wang, D.-L.; Yan, H.-J.; Liu, X.-J. Acute Toxicity to Zebrafish of Two Organophosphates and Four Pyrethroids and Their Binary Mixtures. Pest. Manag. Sci. 2010, 66, 84–89. [Google Scholar] [CrossRef] [PubMed]
- Techer, D.; Milla, S.; Fontaine, P.; Viot, S.; Thomas, M. Acute Toxicity and Sublethal Effects of Gallic and Pelargonic Acids on the Zebrafish Danio Rerio. Environ. Sci. Pollut. Res. Int. 2015, 22, 5020–5029. [Google Scholar] [CrossRef]
- Shi, G.; Xie, Y.; Guo, Y.; Dai, J. 6:2 Fluorotelomer Sulfonamide Alkylbetaine (6:2 FTAB), a Novel Perfluorooctane Sulfonate Alternative, Induced Developmental Toxicity in Zebrafish Embryos. Aquat. Toxicol. 2018, 195, 24–32. [Google Scholar] [CrossRef]
- Köktürk, M.; Altindağ, F.; Ozhan, G.; Çalimli, M.H.; Nas, M.S. Textile Dyes Maxilon Blue 5G and Reactive Blue 203 Induce Acute Toxicity and DNA Damage during Embryonic Development of Danio Rerio. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 242, 108947. [Google Scholar] [CrossRef]
- Huang, H.; Huang, C.; Wang, L.; Ye, X.; Bai, C.; Simonich, M.T.; Tanguay, R.L.; Dong, Q. Toxicity, Uptake Kinetics and Behavior Assessment in Zebrafish Embryos Following Exposure to Perfluorooctanesulphonicacid (PFOS). Aquat. Toxicol. 2010, 98, 139–147. [Google Scholar] [CrossRef]
- Chow, W.S.; Chan, W.K.-L.; Chan, K.M. Toxicity Assessment and Vitellogenin Expression in Zebrafish (Danio Rerio) Embryos and Larvae Acutely Exposed to Bisphenol A, Endosulfan, Heptachlor, Methoxychlor and Tetrabromobisphenol A. J. Appl. Toxicol. 2013, 33, 670–678. [Google Scholar] [CrossRef]
- Yang, L.; Feng, J.; Gao, Y.; Zhu, L. Role of Toxicokinetic and Toxicodynamic Parameters in Explaining the Sensitivity of Zebrafish Larvae to Four Metals. Environ. Sci. Technol. 2021, 55, 8965–8976. [Google Scholar] [CrossRef] [PubMed]
- Pasqualetti, S.; Banfi, G.; Mariotti, M. The Effects of Strontium on Skeletal Development in Zebrafish Embryo. J. Trace Elem. Med. Biol. 2013, 27, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chen, S.; Du, M.; Tang, S.; Chen, M.; Wang, W.; Yang, H.; Chen, Q.; Chen, J. Toxic Effect of Palladium on Embryonic Development of Zebrafish. Aquat. Toxicol. 2015, 159, 208–216. [Google Scholar] [CrossRef]
- Trestrail, C.; Walpitagama, M.; Hedges, C.; Truskewycz, A.; Miranda, A.; Wlodkowic, D.; Shimeta, J.; Nugegoda, D. Foaming at the Mouth: Ingestion of Floral Foam Microplastics by Aquatic Animals. Sci. Total Environ. 2020, 705, 135826. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.A.; Hall, L.W. Environmental Concentrations and Aquatic Toxicity Data on Diflubenzuron (Dimilin). Crit. Rev. Toxicol. 1992, 22, 45–79. [Google Scholar] [CrossRef] [PubMed]
- Péan, S.; Daouk, T.; Vignet, C.; Lyphout, L.; Leguay, D.; Loizeau, V.; Bégout, M.-L.; Cousin, X. Long-Term Dietary-Exposure to Non-Coplanar PCBs Induces Behavioral Disruptions in Adult Zebrafish and Their Offspring. Neurotoxicol. Teratol. 2013, 39, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Lam, S.H.; Hlaing, M.M.; Zhang, X.; Yan, C.; Duan, Z.; Zhu, L.; Ung, C.Y.; Mathavan, S.; Ong, C.N.; Gong, Z. Toxicogenomic and Phenotypic Analyses of Bisphenol-A Early-Life Exposure Toxicity in Zebrafish. PLoS ONE 2011, 6, e28273. [Google Scholar] [CrossRef]
- Du, Y.; Shi, X.; Liu, C.; Yu, K.; Zhou, B. Chronic Effects of Water-Borne PFOS Exposure on Growth, Survival and Hepatotoxicity in Zebrafish: A Partial Life-Cycle Test. Chemosphere 2009, 74, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Kinnberg, K.L.; Petersen, G.I.; Albrektsen, M.; Minghlani, M.; Awad, S.M.; Holbech, B.F.; Green, J.W.; Bjerregaard, P.; Holbech, H. Endocrine-Disrupting Effect of the Ultraviolet Filter Benzophenone-3 in Zebrafish, Danio Rerio. Environ. Toxicol. Chem. 2015, 34, 2833–2840. [Google Scholar] [CrossRef]
- Bakos, K.; Kovacs, R.; Balogh, E.; Sipos, D.K.; Reining, M.; Gyomorei-Neuberger, O.; Balazs, A.; Kriszt, B.; Bencsik, D.; Csepeli, A.; et al. Estrogen Sensitive Liver Transgenic Zebrafish (Danio Rerio) Line (Tg(Vtg1:MCherry)) Suitable for the Direct Detection of Estrogenicity in Environmental Samples. Aquat. Toxicol. 2019, 208, 157–167. [Google Scholar] [CrossRef]
- Johnston, T.K.; Perkins, E.; Ferguson, D.C.; Cropek, D.M. Tissue Explant Coculture Model of the Hypothalamic-Pituitary-Gonadal-Liver Axis of the Fathead Minnow (Pimephales Promelas) as a Predictive Tool for Endocrine Disruption. Environ. Toxicol. Chem. 2016, 35, 2530–2541. [Google Scholar] [CrossRef] [PubMed]
- Wirbisky, S.E.; Freeman, J.L. Atrazine Exposure and Reproductive Dysfunction through the Hypothalamus-Pituitary-Gonadal (HPG) Axis. Toxics 2015, 3, 414–450. [Google Scholar] [CrossRef] [PubMed]
- Fetter, E.; Smetanová, S.; Baldauf, L.; Lidzba, A.; Altenburger, R.; Schüttler, A.; Scholz, S. Identification and Characterization of Androgen-Responsive Genes in Zebrafish Embryos. Environ. Sci. Technol. 2015, 49, 11789–11798. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Sheng, N.; Wang, M.; Zhang, H.; Dai, J. Zebrafish Reproductive Toxicity Induced by Chronic Perfluorononanoate Exposure. Aquat. Toxicol. 2016, 175, 269–276. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Dai, X.; Chen, X.; He, J.; Yin, Z. Zebrafish Pituitary Gene Expression before and after Sexual Maturation. J. Endocrinol. 2014, 221, 429–440. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Fu, Y.; Zhao, C.; Fan, L.; Hu, H.; Yin, S. Ciprofloxacin and Enrofloxacin Can Cause Reproductive Toxicity via Endocrine Signaling Pathways. Ecotoxicol. Environ. Saf. 2022, 244, 114049. [Google Scholar] [CrossRef]
- Zhang, X.; Zhong, Y.; Tian, H.; Wang, W.; Ru, S. Impairment of the Cortisol Stress Response Mediated by the Hypothalamus-Pituitary-Interrenal (HPI) Axis in Zebrafish (Danio Rerio) Exposed to Monocrotophos Pesticide. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2015, 176–177, 10–16. [Google Scholar] [CrossRef]
- Gründker, C.; Emons, G. Role of Gonadotropin-Releasing Hormone (GnRH) in Ovarian Cancer. Cells 2021, 10, 437. [Google Scholar] [CrossRef]
- Recchia, K.; Jorge, A.S.; Pessôa, L.V.d.F.; Botigelli, R.C.; Zugaib, V.C.; de Souza, A.F.; Martins, D.d.S.; Ambrósio, C.E.; Bressan, F.F.; Pieri, N.C.G. Actions and Roles of FSH in Germinative Cells. Int. J. Mol. Sci. 2021, 22, 10110. [Google Scholar] [CrossRef]
- Pellegrini, E.; Menuet, A.; Lethimonier, C.; Adrio, F.; Gueguen, M.-M.; Tascon, C.; Anglade, I.; Pakdel, F.; Kah, O. Relationships between Aromatase and Estrogen Receptors in the Brain of Teleost Fish. Gen. Comp. Endocrinol. 2005, 142, 60–66. [Google Scholar] [CrossRef]
- Tong, S.-K.; Chung, B. Analysis of Zebrafish Cyp19 Promoters. J. Steroid Biochem. Mol. Biol. 2003, 86, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Luu-The, V. Assessment of Steroidogenesis and Steroidogenic Enzyme Functions. J. Steroid Biochem. Mol. Biol. 2013, 137, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Alsop, D.; Vijayan, M.M. Molecular Programming of the Corticosteroid Stress Axis during Zebrafish Development. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2009, 153, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Peng, L.-B.; Wang, D.; Zhu, Q.-L.; Zheng, J.-L. Combined Effects of Polystyrene Microplastics and Cadmium on Oxidative Stress, Apoptosis, and GH/IGF Axis in Zebrafish Early Life Stages. Sci. Total Environ. 2022, 813, 140396. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.; Gu, A.; Ji, G.; Li, Y.; Di, J.; Jin, J.; Hu, F.; Long, Y.; Xia, Y.; Lu, C.; et al. Developmental Toxicity of Cypermethrin in Embryo-Larval Stages of Zebrafish. Chemosphere 2011, 85, 1010–1016. [Google Scholar] [CrossRef]
- Shi, X.; Wu, R.; Wang, X.; Huang, W.; Zheng, S.; Zhang, Q.; Peng, J.; Tan, W.; Wu, K. Effects of 2,2′,4,4′-Tetrabromodiphenyl Ether (BDE-47) on Reproductive and Endocrine Function in Female Zebrafish (Danio Rerio). Ecotoxicol. Environ. Saf. 2022, 248, 114326. [Google Scholar] [CrossRef]
- Li, Y.; Chen, Q.; Liu, Y.; Bi, L.; Jin, L.; Xu, K.; Peng, R. High Glucose-Induced ROS-Accumulation in Embryo-Larval Stages of Zebrafish Leads to Mitochondria-Mediated Apoptosis. Apoptosis 2022, 27, 509–520. [Google Scholar] [CrossRef]
- Cao, F.; Zhu, L.; Li, H.; Yu, S.; Wang, C.; Qiu, L. Reproductive Toxicity of Azoxystrobin to Adult Zebrafish (Danio Rerio). Environ. Pollut. 2016, 219, 1109–1121. [Google Scholar] [CrossRef]
- Lopes, F.M.; Varela Junior, A.S.; Corcini, C.D.; da Silva, A.C.; Guazzelli, V.G.; Tavares, G.; da Rosa, C.E. Effect of Glyphosate on the Sperm Quality of Zebrafish Danio Rerio. Aquat. Toxicol. 2014, 155, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Islinger, M.; Willimski, D.; Völkl, A.; Braunbeck, T. Effects of 17a-Ethinylestradiol on the Expression of Three Estrogen-Responsive Genes and Cellular Ultrastructure of Liver and Testes in Male Zebrafish. Aquat. Toxicol. 2003, 62, 85–103. [Google Scholar] [CrossRef]
- Xu, H.; Yang, J.; Wang, Y.; Jiang, Q.; Chen, H.; Song, H. Exposure to 17alpha-Ethynylestradiol Impairs Reproductive Functions of Both Male and Female Zebrafish (Danio Rerio). Aquat. Toxicol. 2008, 88, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Hu, J.; Yang, J.; Wang, Y.; Xu, H.; Jiang, Q.; Gong, Y.; Gu, Y.; Song, H. Generation of a Fluorescent Transgenic Zebrafish for Detection of Environmental Estrogens. Aquat. Toxicol. 2010, 96, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Alsakran, A.; Kudoh, T. Zebrafish as a Model for Fetal Alcohol Spectrum Disorders. Front. Pharmacol. 2021, 12, 721924. [Google Scholar] [CrossRef]
- Torres-Ruiz, M.; De la Vieja, A.; Gonzalez, M.d.A.; Lopez, M.E.; Calvo, A.C.; Portilla, A.I.C. Toxicity of Nanoplastics for Zebrafish Embryos, What We Know and Where to Go Next. Sci. Total Environ. 2021, 797, 149125. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Huang, Z.; Zhao, L.; Liu, W.; Chen, X.; Meng, P.; Lin, Q.; Chi, Y.; Xu, M.; Ma, N.; et al. Large-Scale Forward Genetic Screening Analysis of Development of Hematopoiesis in Zebrafish. J. Genet. Genom. 2012, 39, 473–480. [Google Scholar] [CrossRef]
- de Jong, E.; Barenys, M.; Hermsen, S.A.B.; Verhoef, A.; Ossendorp, B.C.; Bessems, J.G.M.; Piersma, A.H. Comparison of the Mouse Embryonic Stem Cell Test, the Rat Whole Embryo Culture and the Zebrafish Embryotoxicity Test as Alternative Methods for Developmental Toxicity Testing of Six 1,2,4-Triazoles. Toxicol. Appl. Pharmacol. 2011, 253, 103–111. [Google Scholar] [CrossRef]
- Brannen, K.C.; Panzica-Kelly, J.M.; Danberry, T.L.; Augustine-Rauch, K.A. Development of a Zebrafish Embryo Teratogenicity Assay and Quantitative Prediction Model. Birth Defects Res. B Dev. Reprod. Toxicol. 2010, 89, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Selderslaghs, I.W.T.; Blust, R.; Witters, H.E. Feasibility Study of the Zebrafish Assay as an Alternative Method to Screen for Developmental Toxicity and Embryotoxicity Using a Training Set of 27 Compounds. Reprod Toxicol. 2012, 33, 142–154. [Google Scholar] [CrossRef]
- Brotzmann, K.; Wolterbeek, A.; Kroese, D.; Braunbeck, T. Neurotoxic Effects in Zebrafish Embryos by Valproic Acid and Nine of Its Analogues: The Fish-Mouse Connection? Arch. Toxicol. 2021, 95, 641–657. [Google Scholar] [CrossRef]
- Carrillo, A.; McHenry, M.J. Zebrafish Learn to Forage in the Dark. J. Exp. Biol. 2016, 219 Pt 4, 582–589. [Google Scholar] [CrossRef]
- Yashina, K.; Tejero-Cantero, Á.; Herz, A.; Baier, H. Zebrafish Exploit Visual Cues and Geometric Relationships to Form a Spatial Memory. iScience 2019, 19, 119–134. [Google Scholar] [CrossRef]
- Facciol, A.; Bailleul, C.; Nguyen, S.; Chatterjee, D.; Gerlai, R. Developmental Stage-Dependent Deficits Induced by Embryonic Ethanol Exposure in Zebrafish: A Neurochemical Analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 2020, 99, 109859. [Google Scholar] [CrossRef] [PubMed]
- Chow, E.S.H.; Hui, M.N.Y.; Lin, C.C.; Cheng, S.H. Cadmium Inhibits Neurogenesis in Zebrafish Embryonic Brain Development. Aquat. Toxicol. 2008, 87, 157–169. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.; Korzh, V.; Strahle, U. Zebrafish Embryos Are Susceptible to the Dopaminergic Neurotoxin MPTP. Eur. J. Neurosci. 2005, 21, 1758–1762. [Google Scholar] [CrossRef]
- Chen, J.; Kong, A.; Shelton, D.; Dong, H.; Li, J.; Zhao, F.; Bai, C.; Huang, K.; Mo, W.; Chen, S.; et al. Early Life Stage Transient Aristolochic Acid Exposure Induces Behavioral Hyperactivity but Not Nephrotoxicity in Larval Zebrafish. Aquat. Toxicol. 2021, 238, 105916. [Google Scholar] [CrossRef]
- Gu, J.; Guo, M.; Huang, C.; Wang, X.; Zhu, Y.; Wang, L.; Wang, Z.; Zhou, L.; Fan, D.; Shi, L.; et al. Titanium Dioxide Nanoparticle Affects Motor Behavior, Neurodevelopment and Axonal Growth in Zebrafish (Danio Rerio) Larvae. Sci. Total Environ. 2021, 754, 142315. [Google Scholar] [CrossRef]
- Wang, L.; Ma, J.; Wu, W.; Fang, Y.; Liu, F.; Yang, Q.; Hu, X.; Gu, X.; He, Z.; Sun, D.; et al. Effect of Aerobic Exercise as a Treatment on Type 2 Diabetes Mellitus with Depression-like Behavior Zebrafish. Life Sci. 2022, 300, 120578. [Google Scholar] [CrossRef]
- Faria, M.; Bedrossiantz, J.; Ramírez, J.R.R.; Mayol, M.; García, G.H.; Bellot, M.; Prats, E.; Garcia-Reyero, N.; Gómez-Canela, C.; Gómez-Oliván, L.M.; et al. Glyphosate Targets Fish Monoaminergic Systems Leading to Oxidative Stress and Anxiety. Environ. Int. 2021, 146, 106253. [Google Scholar] [CrossRef]
- Chen, K.; Wu, M.; Chen, C.; Xu, H.; Wu, X.; Qiu, X. Impacts of Chronic Exposure to Sublethal Diazepam on Behavioral Traits of Female and Male Zebrafish (Danio Rerio). Ecotoxicol. Environ. Saf. 2021, 208, 111747. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, H.C.; Amindari, A.; Butcher, J.T.; Althani, A.; Yacoub, M. Heart Function and Hemodynamic Analysis for Zebrafish Embryos. Dev. Dyn. 2017, 246, 868–880. [Google Scholar] [CrossRef]
- Sun, M.; Ding, R.; Ma, Y.; Sun, Q.; Ren, X.; Sun, Z.; Duan, J. Cardiovascular Toxicity Assessment of Polyethylene Nanoplastics on Developing Zebrafish Embryos. Chemosphere 2021, 282, 131124. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, Z.; Fan, Z.; Fang, Y.; He, L.; Peng, M.; Chen, Y.; Hu, Z.; Zhao, K.; Zhang, H.; et al. Cardiac Developmental Toxicity and Transcriptome Analyses of Zebrafish (Danio Rerio) Embryos Exposed to Mancozeb. Ecotoxicol. Environ. Saf. 2021, 226, 112798. [Google Scholar] [CrossRef] [PubMed]
- Cj, G.; Lp, W. Comparison of the Acute Effects of Benzo-a-Pyrene on Adult Zebrafish (Danio Rerio) Cardiorespiratory Function Following Intraperitoneal Injection versus Aqueous Exposure. Aquat. Toxicol. 2015, 165, 19–30. [Google Scholar] [CrossRef]
- Eimon, P.M.; Rubinstein, A.L. The Use of in Vivo Zebrafish Assays in Drug Toxicity Screening. Expert Opin. Drug Metab. Toxicol. 2009, 5, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.-J.; Xu, Y.-Q.; He, J.-H.; Yu, H.-P.; Huang, C.-J.; Gao, J.-M.; Dong, Q.-X.; Xuan, Y.-X.; Li, C.-Q. Human Cardiotoxic Drugs Delivered by Soaking and Microinjection Induce Cardiovascular Toxicity in Zebrafish. J. Appl. Toxicol. 2014, 34, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Katoch, S.; Patial, V. Zebrafish: An Emerging Model System to Study Liver Diseases and Related Drug Discovery. J. Appl. Toxicol. 2021, 41, 33–51. [Google Scholar] [CrossRef]
- Meng, Q.; Yeung, K.; Chan, K.M. Toxic Effects of Octocrylene on Zebrafish Larvae and Liver Cell Line (ZFL). Aquat. Toxicol. 2021, 236, 105843. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Dang, Y.; Gong, Z.; Letcher, R.J.; Liu, C. Progression of Liver Tumor Was Promoted by Tris(1,3-Dichloro-2-Propyl) Phosphate through the Induction of Inflammatory Responses in KrasV12 Transgenic Zebrafish. Environ. Pollut. 2019, 255 Pt 2, 113315. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.L.; Liu, S.Y.; Zhang, J.; Hu, C.X.; Li, D.H.; Liu, Y.D. Antioxidative Responses in Zebrafish Liver Exposed to Sublethal Doses Aphanizomenon Flos-Aquae DC-1 Aphantoxins. Ecotoxicol. Environ. Saf. 2015, 113, 425–432. [Google Scholar] [CrossRef]
- Drummond, I.A.; Davidson, A.J. Zebrafish Kidney Development. Methods Cell Biol. 2016, 134, 391–429. [Google Scholar] [CrossRef]
- Ding, Y.-J.; Chen, Y.-H. Developmental Nephrotoxicity of Aristolochic Acid in a Zebrafish Model. Toxicol. Appl. Pharmacol. 2012, 261, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Nilsson, M.; Fagman, H. Development of the Thyroid Gland. Development 2017, 144, 2123–2140. [Google Scholar] [CrossRef] [PubMed]
- Yu, K.; Li, X.; Qiu, Y.; Zeng, X.; Yu, X.; Wang, W.; Yi, X.; Huang, L. Low-Dose Effects on Thyroid Disruption in Zebrafish by Long-Term Exposure to Oxytetracycline. Aquat. Toxicol. 2020, 227, 105608. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Dang, J.; Paudel, Y.N.; Wang, X.; Wang, B.; Wang, L.; Li, P.; Sun, C.; Liu, K. The Possible Hormetic Effects of Fluorene-9-Bisphenol on Regulating Hypothalamic-Pituitary-Thyroid Axis in Zebrafish. Sci. Total Environ. 2021, 776, 145963. [Google Scholar] [CrossRef] [PubMed]
- Fazio, M.; Ablain, J.; Chuan, Y.; Langenau, D.M.; Zon, L.I. Zebrafish Patient Avatars in Cancer Biology and Precision Cancer Therapy. Nat. Rev. Cancer 2020, 20, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.-W.; Hsia, Y.; Tu, H.-C.; Hsiao, Y.-C.; Yang, W.-Y.; Wang, H.-D.; Yuh, C.-H. Liver Development and Cancer Formation in Zebrafish. Birth Defects Res. Part C Embryo Today Rev. 2011, 93, 157–172. [Google Scholar] [CrossRef]
- Hwang, K.L.; Goessling, W. Baiting for Cancer: Using the Zebrafish as a Model in Liver and Pancreatic Cancer. Adv. Exp. Med. Biol. 2016, 916, 391–410. [Google Scholar] [CrossRef]
- Spitsbergen, J.M.; Tsai, H.W.; Reddy, A.; Miller, T.; Arbogast, D.; Hendricks, J.D.; Bailey, G.S. Neoplasia in Zebrafish (Danio Rerio) Treated with 7,12-Dimethylbenz[a]Anthracene by Two Exposure Routes at Different Developmental Stages. Toxicol. Pathol. 2000, 28, 705–715. [Google Scholar] [CrossRef]
- Spitsbergen, J.M.; Buhler, D.R.; Peterson, T.S. Neoplasia and Neoplasm-Associated Lesions in Laboratory Colonies of Zebrafish Emphasizing Key Influences of Diet and Aquaculture System Design. ILAR J. 2012, 53, 114–125. [Google Scholar] [CrossRef]
- Spitsbergen, J.M.; Tsai, H.W.; Reddy, A.; Miller, T.; Arbogast, D.; Hendricks, J.D.; Bailey, G.S. Neoplasia in Zebrafish (Danio Rerio) Treated with N-Methyl-N’-Nitro-N-Nitrosoguanidine by Three Exposure Routes at Different Developmental Stages. Toxicol. Pathol. 2000, 28, 716–725. [Google Scholar] [CrossRef]
Figure 1.
Toxicological effects of Cd on zebrafish at different developmental stages. Zebrafish at different developmental stages were used as model organisms to detect the toxic effects of Cd, a common metal pollutant, even in the same organ, and the toxic effects were different at different developmental stages [30,31].
Figure 1.
Toxicological effects of Cd on zebrafish at different developmental stages. Zebrafish at different developmental stages were used as model organisms to detect the toxic effects of Cd, a common metal pollutant, even in the same organ, and the toxic effects were different at different developmental stages [30,31].

Figure 2.
Different types of zebrafish disease models caused by exposure to microplastics (a) and toxic effects of microplastics of different particle sizes on zebrafish (b) [52].
Figure 2.
Different types of zebrafish disease models caused by exposure to microplastics (a) and toxic effects of microplastics of different particle sizes on zebrafish (b) [52].

Figure 3.
Different growth stages of zebrafish can be subjected to corresponding long−term or shor−term exposure experiments (a) and under the condition of exposure to pollutants, the antioxidant system in zebrafish cells is damaged and a large amount of ROS is produced (b) [58,59,60].

Figure 4.
Application of the zebrafish model in environmental toxicology evaluation. Changes in multilevel endpoints and biomarkers such as appearance, metabolism, and molecular structure can be used to evaluate the embryonic toxicity, reproductive toxicity, neurotoxicity, and liver and kidney toxicity of environmental pollutants in zebrafish.
Figure 4.
Application of the zebrafish model in environmental toxicology evaluation. Changes in multilevel endpoints and biomarkers such as appearance, metabolism, and molecular structure can be used to evaluate the embryonic toxicity, reproductive toxicity, neurotoxicity, and liver and kidney toxicity of environmental pollutants in zebrafish.


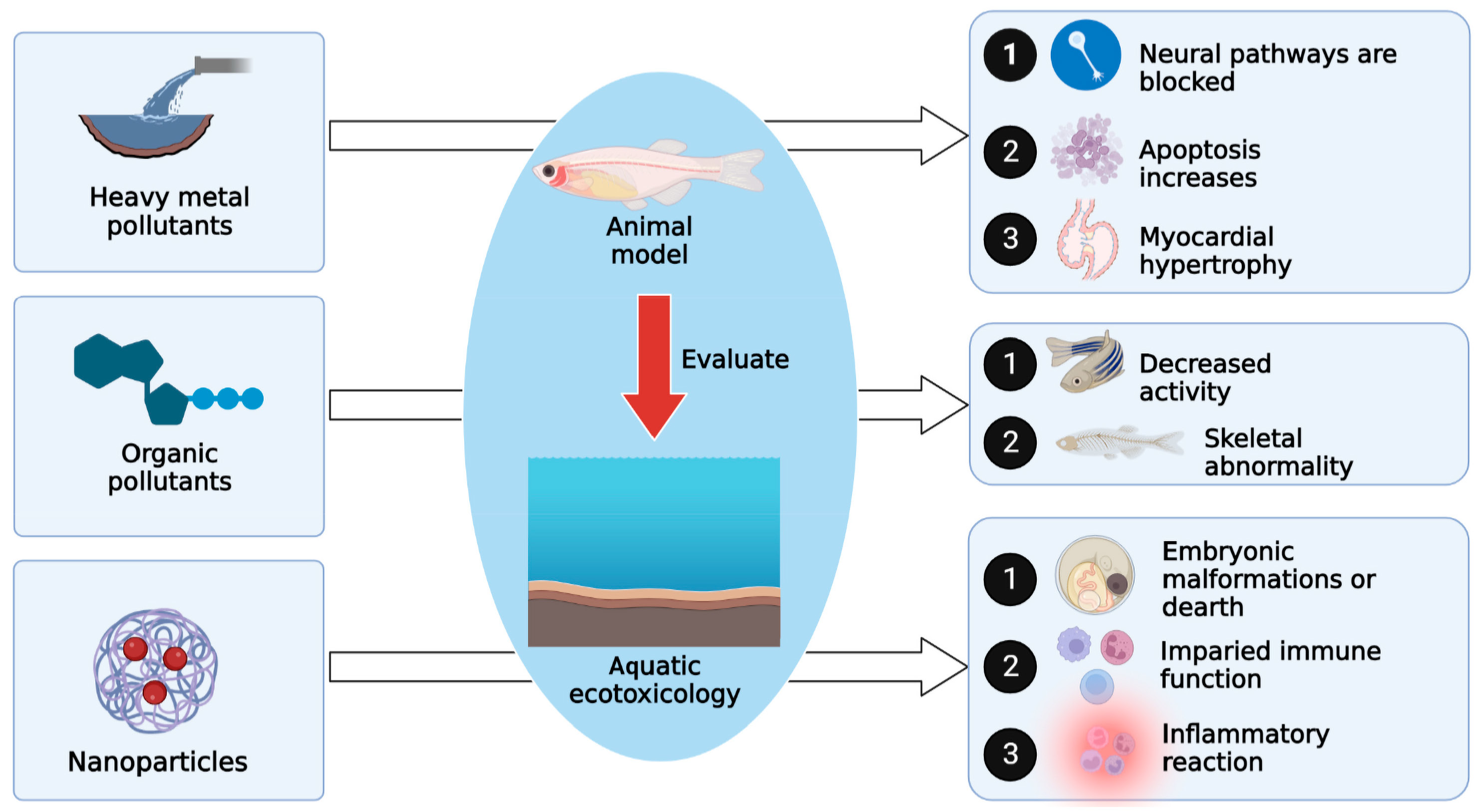{kind=link}
{kind=link}
{kind=link}
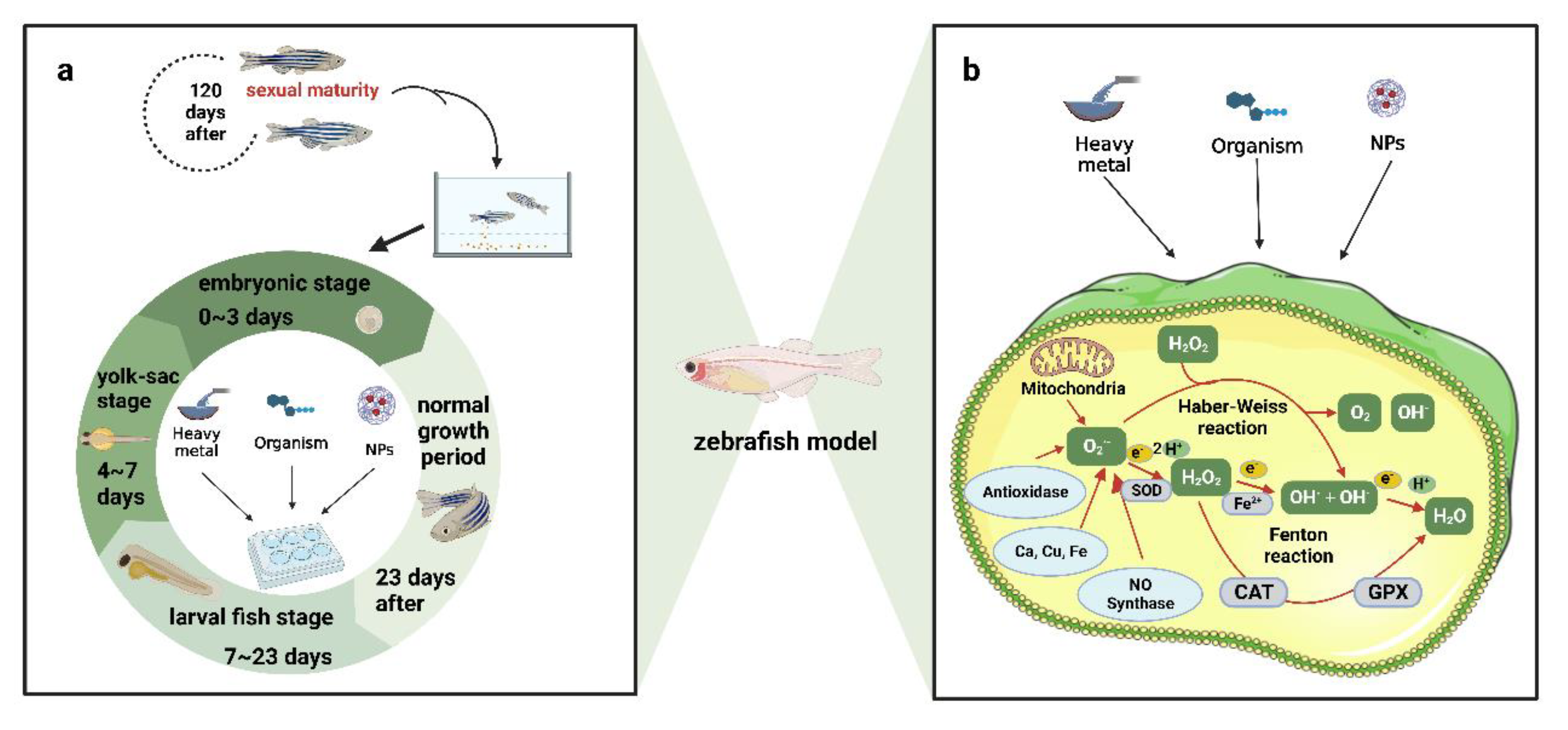{kind=link}
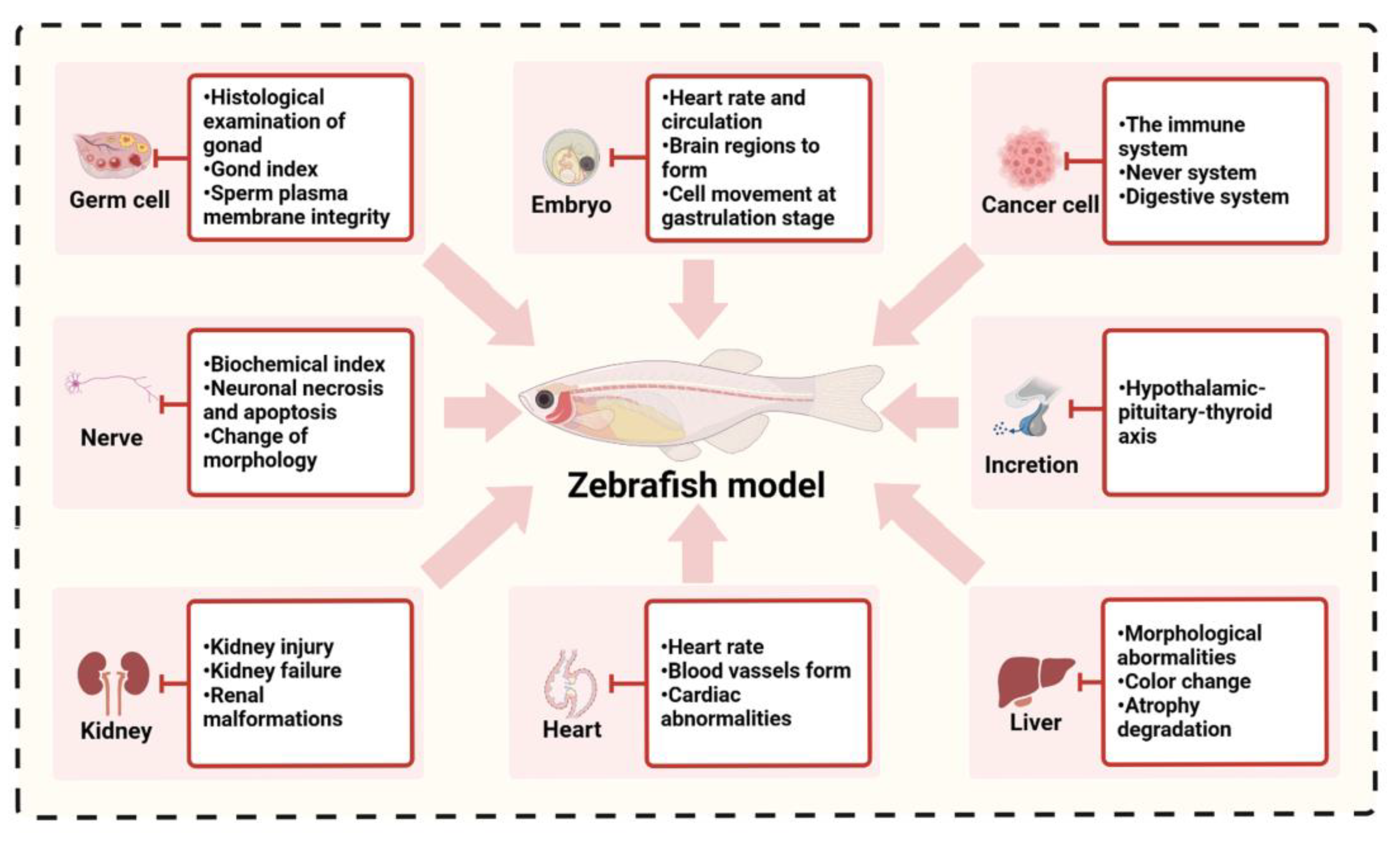{kind=link}
Table 1.
Biomarkers/end points for exposure experiments of zebrafish to different pollutants.
| Pollutants Classes | Contaminants | Biomarkers/Endpoints | Reference |
|---|---|---|---|
| Heavy metal | Cr (VI) | Cholinesterase, glutathione S-transferase and lactate dehydrogenase | [14] |
| Cadmium acetate, lead acetate, mercury chloride, zinc chloride | Acetylcholinesterase activity, gene expression pattern, antioxidant competence in the brain | [15] | |
| Lead contaminated sediments | Survival rate, C-start response, skeletal malformations and bent spines, pericardial edema and yolk sacmalformations, ALA-D gene expression | [16] | |
| Cd, Pb | Embryo lethality and teratogenicity, oxygen species (ROS), Nrf2 and IFN-γ gene expression | [17] | |
| Cd | specific growth rate (SGR), 17β-estradiol (E2) and vitellogenin (VTG) concentrations in plasma, gonadosomatic index (GSI), egg-laying amount, spawning percentage, and hatching and mortality rate of embryos, vtg1 gene expression | [18] | |
| Organic pollutant | MgHP | Catalase (CAT), reactive oxygen species (ROS), glutathione, metallothionein values | [19] |
| Prochloraz (PCZ) | Cholinesterase, glutathione S-transferase, and lactate dehydrogenase | [20] | |
| Chlorpyrifos (CHL) | Rate of deformity, cytochrome P450 (CYP450), NR1I2 gene, UGT1a1, and MDR1 expression level | [21] | |
| Oxytetracycline, amoxicillin | Premature hatch, delayed hatching of embryos, CAT, glutathione-S-transferases, lactate dehydrogenase | [22] | |
| NPs | MPs | CAT, superoxide dismutase (SOD), D-lactate, Diamine oxidase (DAO) activity | [23] |
| PS-NPs | Acetylcholinesterase (AChE), ROS, oxytocin, Adenosine triphosphate (ATP), malondialdehyde (MDA), single stranded DNA (ssDNA), VTG, dopamine (DA), CYP1A1, γ-aminobutyric acid (GABA) | [24] |
Table 2.
LC50 for exposure of zebrafish to different pollutants.
| Pollutant | Time | LC50 | Ref. | |
|---|---|---|---|---|
| Organic pollutant | Permethrin | 24 h | 0.0052 (0.0041–0.0066) mg/L | [65] |
| 48 h | 0.0030 (0.0019–0.0038) mg/L | |||
| 72 h | 0.0026 (0.0018–0.0033) mg/L | |||
| 96 h | 0.0025 (0.0017–0.0032) mg/L | |||
| Tetramethrin | 24 h | 0.0782 (0.0728–0.0838) mg/L | ||
| 48 h | 0.0517 (0.0470–0.0572) mg/L | |||
| 72 h | 0.0484 (0.0436–0.0596) mg/L | |||
| 96 h | 0.0460 (0.0436–0.0481) mg/L | |||
| Bifenthrin | 24 h | 0.0065 (0.0051–0.0093) mg/L | ||
| 48 h | 0.0039 (0.0026–0.0050) mg/L | |||
| 72 h | 0.0034 (0.0020–0.0046) mg/L | |||
| 96 h | 0.0032 (0.0021–0.0041) mg/L | |||
| Etofenprox | 24 h | 0.0969 (0.0701–0.121) mg/L | ||
| 48 h | 0.0865 (0.0613–0.109) mg/L | |||
| 72 h | 0.0825 (0.0561–0.105) mg/L | |||
| 96 h | 0.0791 (0.0564–0.0988) mg/L | |||
| Dichlorvos | 24 h | 51.3 (47.2–55.8) mg/L | ||
| 48 h | 32.3 (28.7–36.0) mg/L | |||
| 72 h | 15.0 (12.1–18.4) mg/L | |||
| 96 h | 13.0 (10.4–16.1) mg/L | |||
| Gallic acid | 96 h | >100 mg/L | [66] | |
| Pelargonic acid | 96 h | 81.2 mg/L | ||
| 6:2 fluorotelomer sulfonamide alkylbetaine (6:2 FTAB) | 120 h | 43.73 ± 3.24 mg/L | [67] | |
| Maxilon blue 5G (MB-5G) | 96 h | 166.04 | [68] | |
| Reactive Blue 203 (RB-203) | 96 h | 278.32 | ||
| perfluorooctane sulfonate (PFOS) | 120 h | 2.20 mg/L | [69] | |
| Bisphenol A(BPA) | 96 h | 8.041 mg/L | [70] | |
| Endosulfan | 96 h | 0.238 mg/L | ||
| Heptachlor | 96 h | 1.738 mg/L | ||
| Heavy metal | Cd | 72 h | 1.823 ± 0.125 μmol/L | [71] |
| Pb | 72 h | 292.6 μg/L | ||
| Cu | 72 h | 0.38 ± 0.092 μmol/L | ||
| Zn | 72 h | 12.432 ± 0.497 μmol/L | ||
| 90Sr | 6 d | 6 mM | [72] | |
| Pd | 48 h | 292.6 μg/L | [73] | |
| NPs | Regular foam Phenol-formaldehyde microplastics | 96 h (170.4 ± 147.5 µm) | 27.1 mg/mL | [74] |
| Biofoam Phenol-formaldehydemicroplastics | 96 h (155.9 ± 56.4 µm) | 43.8 mg/mL | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Li, H.; Liu, Y.; Chen, Q.; Jin, L.; Peng, R. Research Progress of Zebrafish Model in Aquatic Ecotoxicology. Water 2023, 15, 1735. https://doi.org/10.3390/w15091735
AMA Style
Li H, Liu Y, Chen Q, Jin L, Peng R. Research Progress of Zebrafish Model in Aquatic Ecotoxicology. Water. 2023; 15(9):1735. https://doi.org/10.3390/w15091735
Chicago/Turabian StyleLi, Huiqi, Yinai Liu, Qianqian Chen, Libo Jin, and Renyi Peng. 2023. "Research Progress of Zebrafish Model in Aquatic Ecotoxicology" Water 15, no. 9: 1735. https://doi.org/10.3390/w15091735
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.