
Growth and Muscle Quality of Grass Carp (Ctenopharyngodon idella) in In-Pond Raceway Aquaculture and Traditional Pond Culture

1
Key Laboratory of Exploitation and Utilization of Aquatic Genetic Resources, Ministry of Education, Shanghai Ocean University, Shanghai 201306, China
2
China-ASEAN “The Belt and Road” Joint Laboratory of Marine Culture Technology (Shanghai), Shanghai Ocean University, Shanghai 201306, China
3
Shanghai Engineering Research Center of Aquaculture, Shanghai Ocean University, Shanghai 201306, China
*
Authors to whom correspondence should be addressed.
Water 2023, 15(9), 1771; https://doi.org/10.3390/w15091771
Submission received: 31 March 2023
/
Revised: 27 April 2023
/
Accepted: 30 April 2023
/
Published: 5 May 2023
Abstract
:In-pond raceway aquaculture (IPRA) is the rational prescription for water eutrophication and improves the pond environment, enhancing production and the quality of fish. This experiment explored the growth performances and muscle quality of grass carp with stocking densities of 32 tail/m3 and 0.07 tail/m2 in IPRA and traditional pond culture (TPC), respectively. The hepatosomatic index, visceral mass ratio, and correction factor were statistically similar in IPRA compared to TPC. While the weight gain rate (p < 0.001) and the content of crude lipid (p < 0.05) in the flesh of grass carp were observed to be statistically promising in IPRA, the pH and water holding capacity, as well as hardness and chewiness, in grass carp muscle were not significantly different between the two culture systems. However, the 2-MIB concentration in the muscle was observed to increase continuously for the complete culture period in IPRA. The abundance of Proteobacteria was found to be higher in TPC (p < 0.05), while the richness of Planctomycetes was superior in IPRA (p < 0.05). Despite the high stocking density, the off-flavor in IPRA-produced grass carp had less of an impact on the flesh aesthetic quality compared to TPC. Considering all these facts, the results of this study show that grass carp with a better muscle quality can be produced from IPRA.
1. Introduction
Production intensification is an alternative for increasing production and productivity and for ensuring the sustainable supply of aquaculture production [1,2]. In fact, pond culture is a key practice globally for maximizing production, protein supply, and income [3]. The production of grass carp (Ctenopharyngodon idella) ranks highly among the freshwater aquaculture industries [4], accounting for11.8% of global aquaculture production [5].
The growth of fish is affected by multiple factors, from the rearing environment and food nutrition to genetics [6,7]. IPRA ensures the better control of water movement, enhances water quality, and allows for high-density stocking, thus increasing production and productivity compared to TPC. Wang et al. [8] documented a high potential for more controllable and efficient production of bluntnose black bream, channel catfish, yellow catfish, and largemouth bass in IPRA, but information on the production efficiency of grass carp in IPRA is scarce. Similarly, Fatima et al. [9] reported a higher average biomass (57.33 kg/m3) of GIFT tilapia from IPRA and 0.38 kg/m3 from traditional ponds in Pakistan.
Wholesomeness, freshness, and integrity are the major aesthetic characteristics that describe fish muscle quality [10]. In other terms, these characteristics define the hygienic, nutritional, and sensory features, as well as the serviceability of the fish. The freshness of fish muscle is reflected in its appearance, flavor, and texture, while nutritional constituents and water-holding capacity (WHC) also have considerable impacts on fish muscle quality. These characteristics are affected by the species, age, and size of fish and other factors (available nutrients, period, water condition and environment, and consumers) as well as the methods of slaughter, storage, and processing [7,11,12,13]. Thus, modulation of the culture structure and environment to enable higher productivity with improved quality is a prime concern of aquaculture at present [7,14].
The nutritional characteristics of fish include protein, lipid, fatty acids, amino acids, and minerals [15]. Wen et al. [16] reported elevated muscle protein content and WHC with depressed moisture, lipid, and ash contents, as well as improved dietary phosphorus in grass carp. In addition, muscle pH has a linear relationship with softness and WHC [13], which describes the characteristics of muscle for avoiding moisture loss. Similarly, water quality parameters were allied with the growth, nutritional, and sensory values of aquatic species, including fish [17,18]; thus, grass carp cultured in poor-quality water yields a low grade of fish quality [19].
With the rapid development of aquaculture, water pollution that negatively affects the water environment and fish quality is becoming an important concern [20,21]. Therefore, water quality deterioration and eutrophication are serious concerns in intensive aquaculture, as they encourage cyanobacterial blooms and result in the production of earthy-musty compounds in fish flesh [14,22]. The common odorous compounds in fish muscle are 2-methylisoborneol (MIB; 1,2,7,7-tetramethylexo-bicyclo[2.2.1]heptan-2-ol) and geosmin (trans-1,10-dimethyl-trans-9-decalol), which degrade quality and reduce the consumer acceptability and market development [23]. Qin et al. [24] and Tang et al. [13] reported microbial diversity of pond water and described Proteobacteria, Bacteriodetes, Actinobacteria, and Planctomycetes as the dominant microorganisms in different culture systems, while Lukassen et al. identified Proteobacteria as a leading geosminproducer [25]. On such a backdrop, improved culture models (such as a split-plot/partition aquaculture system and in-pond raceway system) have been implemented in traditional culture ponds with more efficient water usage, lower environmental impacts, and high-quality aquatic products with no off-flavor complaints; these models have received more attention in aquaculture [26,27,28]. IPRA has been used in recent years as an aquaculture engineering approach and environmental remediation key for water eutrophication. Thus, IPRA is modulated in just 2–5% of pond areas for intensive culture, making the remaining spaces for water purification more productive and easily manageable, ultimately resulting in better water quality [29,30].
Grass carp is a popular fish species with a large global freshwater production, providing an inexpensive source of high-class animal protein for consumers [4,28], and is a good candidate for IPRA. In recent years, the muscle quality of cultured grass carp has been declining, mainly due to pollution and compromised pond water quality [7]. Poor water quality can be a significant driver for reductions in production, product quality, and profit [17,31]; thus, shifting to and adopting IPRA could purify the eutrophic water and improve the culture environment, yielding a higher fish quality. Furthermore, proper feeding management in IPRA for monitoring the amount of residual feed can be a useful approach for enhancing the muscle quality of grass carp [32,33].Thus, this work aimed to investigate the growth and muscle quality of grass carp in IPRA and TPC.
2. Materials and Methods
2.1. Experimental Layout Set-Up
The experiment was carried out in IPRA and TPC for 83 days at Songjian, Shanghai, with the experimental species being grass carp (Ctenopharyngodon idella). The IPRA was installed in a 2.0 ha traditional pond and consisted of three rectangular raceways (out of ten raceways; 25 m × 5 m × 2.5 m) that were interrelated with each other viaa common pathway. Each raceway unit had airlift, fish culturing, and waste transfer compartments. The fish culture area was separated by barriers (1.0 cm × 1.0 cm mesh) that were set up at the two ends of each cell. Stocking density was maintained at 32 tail/m3; surface and bottom aeration were provided. By contrast, TPC was maintained in three 0.33 ha ponds, each of which had the same water depth and a stocking density of 0.07 tail/m2. For all ponds, the standard feed and feeding techniques were used, and occasional topping of a small quantity of water was carried out to compensate for evaporation loss.
2.2. Water Quality Measurement
Water sample collection was performed in an integrated manner using a Plexiglas water collector before 8 a.m. in a fortnightly interval for physicochemical parameters, and the water was subsequently transported to the laboratory for analysis in a cool box. The dissolved oxygen level of both culture systems was more than 6 mg/L for the whole experimental period, which was optimal for the water temperatures of 22–34 °C and pHs of 7.2 to 7.8.
For chemical analysis, water samples were filtered with Whatman GF/C glass fiber (0.45 μm), and nitrite nitrogen (NO2-N) was examined by using spectrophotometry; nitrate nitrogen (NO3-N) was examined by using sulfamic acid ultraviolet spectrophotometry; ammonia nitrogen was examined by using nesslerization colorimetry; and phosphorus (PO43-P) was examined by using phospho-molybdenum blue spectrophotometry. Unfiltered samples were analyzed for total phosphorus (TP) by using ammonium molybdate spectrophotometry method and total nitrogen (TN)was examined by using the alkaline potassium persulfate digestion UV spectrophotometry method. Total suspended solids (TSS) were determined per APHA (2005) and chemical oxygen demand (COD) by using the magnesium persulfate method (GB 1189289). Similarly, chlorophyll-a analysis was performed according to ISO (1992) and WHO (1999). The details of the water quality status in IPRA and TPC are depicted in Table 1.
2.3. Sample Collection and Analysis
Three grass carp fish from each raceway and pond (n = 18) were sampled and transported to the laboratory at 4 °C. The morphological traits of the fish were evaluated, and the Condition Factor (CF) was determined by CF = (TW/SL3) × 100. Weight gain was determined by final weight/initial weight. The hepatosomatic index and the visceral mass ratio were calculated as (LW/TW) × 100 and (VW/TW) × 100, respectively, after identifying the liver weight (LW) and visceral mass weight (VW). Immediate dissection from the dorsal white muscle of the body was performed for proximate nutrient analysis of the fish, and subsamples (54, triplicates of each sample) were preserved frozen (−20 °C) till investigation. The chemical composition of muscle was analyzed according to the National Food Safety Standard, China. Moisture content was determined by using method GB 5009.3-2016; ash by using method GB 5009.4-2016; crude protein by using the Kjeldahl method (GB 5009.5-2016); and the lipid content by using the Soxtherm extraction system (SOX 416 Macro; Gerhardt, Germany). Moisture and ash content were determined within 12 h of postmortem by using the standard method. The fish homogenate, prepared by thoroughly mixing 2 g of muscle in 10 mL of distilled water, was used to measure pH using a Cyberscan model 500 pH meter (Euteon Instruments, Jurong, Singapore) and WHC by using the filter paper method. Muscle quality was evaluated via a texture analyzer (Universal TA, Shanghai, China) using (1 × 1 × 1 cm3) pieces from the dorsal part of the fish. The hardness (g), springiness (mm), cohesiveness, stickiness (g), chewiness (g × mm), and adhesiveness were evaluated through exponential linking software.
2.4. Microbial Diversity in Pond Water
The water sample (500 mL) was filtered through a 0.2 µm polycarbonate filter with a diameter of 47 mm (Millipore, Billerica, MA, USA) and then stored at −20 °C until processing. The DNA was extracted and purified from filtered samples using the soil DNA kit (Omega Biotech, Norcross, GA, USA). A nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Wilmington, NC, USA) was used to evaluate the nucleotide concentration, and the DNA quality was checked by 1% agarose gel electrophoresis. The purified genomic DNA (20 ng/mL) was sent to Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai, China), and the MiSeq benchtop sequencer (Illumina, San Diego, CA, USA) was used for 16S rRNA gene-based amplicon sequencing. The bacterial 16S rRNA gene was analyzed via polymerase chain reaction (PCR) using bacterial primers (338F-ACTCCTACGGAGCAGCAG and 806R-GGACTACHVGGGTWTCTAAT) [35].
PCR amplification in triplicate was performed using a 4.0 µL 5FastPfu buffer, 2.0 µL deoxyribonucleotide triphosphate (dNTP) (2.5 mM), 0.8 µL forward and reverse primers (5.0 mM), 0.4 µL FastPfu polymerase, and 10.0 ng template DNA, respectively. Finally, PCR-grade water was added to the above mixture to obtain a final volume of 20.0 µL. The PCR thermal program was set at 95 °C for 3 min; then 27 cycles were performed at 95 °C, 55 °C, and 72 °C for 30, 30, and 45 s, respectively.
The PCR product was extracted from a 2% agarose gel and further purified using the AxyPrep DNA gel extraction kit (Axygen Biosciences, Union City, CA, USA). The MiSeq technology was used to analyze microbial communities by high-throughput pyrosequencing (Illumina, San Diego, CA, USA). The 16S rDNA gene cloning and sequencing were performed in accordance with the instructions provided by Majorbio Bio-pharm Biotechnology Co., Ltd. (Shanghai, China). The right-sized PCR products were chosen at random, and the National Center for Biotechnology Information (NCBI’s) search engine was used to compare the sequenced 16S rDNA genes and find the gene that most closely matched it. The sequence was compared with the sequence in the GenBank database, and the basic local alignment search tool algorithm was used to determine the approximate phylogenetic attribution. All the sequences used in this study are available from the NCBI Sequence Read Archive (SRA) under accession number MJ20190808066.
2.5. Statistical Analysis
Data analyses were performed using SPSS version20.0 statistical software (IBM, Armonk, NY, USA). Results were shown as mean with standard deviation (mean ± SD). The differences between the IPRA and TPC were evaluated by one-way analysis of variance (ANOVA) followed by Duncan’s multiple comparison tests. For all analyses, statistically significant differences were determined at p < 0.05.
3. Results
3.1. Water Quality Analysis
The dynamics in water quality indicators during the whole experimental period are shown in Figure 1. All parameters illustrated an increasing tendency during the whole culture period. The increasing trends of ammonia nitrogen, TN, PO43−P, and TP were consistent in both culture systems, while the inclinations of NO3-N and NO2-N were different depending on the system.
Among the various forms of nitrogen observed, nitrate nitrogen, ammonia nitrogen, and total nitrogen values were found to have an increasing trend in IPRA and were observed to have a highly significant difference (p < 0.01), except for a few samplings in TPC. The concentrations of nitrite nitrogen were significantly higher (p < 0.01) in IPRA at all sampling days, except at 83 days (p < 0.05). The ascending trend of nitrate nitrogen concentrations in IPRA showed a highly significant difference (p < 0.01) at 0 and 83 days and a significant difference (p < 0.05) at 70 days compared to TPC. Similarly, concentrations of ammonia nitrogen also showed a rising trend in both culture systems; however, it was significantly higher (p < 0.01) at15, 45, and 83 days (p < 0.05) in IPRA. The phosphate concentration was significantly higher (p < 0.01) for the complete culture period in IPRA. Likewise, the concentrations of total nitrogen, total phosphorous, and chemical oxygen demand were significantly higher (p < 0.01) in IPRA. However, chlorophyll–a was higher (p < 0.01) in TPC (Table 1).
3.2. Growth Performance
The growth performances of grass carp were recorded to be higher in TPC than in IPRA during the experimental period. The harvest weight (p < 0.05) and the length (p < 0.05) were significant (p < 0.01) in TPC, while the weight gain rate was significantly better in IPRA. The non- significant differences were observed for the HSI, VMR, and CF of fish in IPRA and TPC systems, but higher values of HSI and VMR were observed in IPRA. The growth performances and biometric measurements of grass carp in IPRA and TPC are depicted in Table 3.
3.3. Nutrient Composition of Muscle
The crude lipid content in grass carp muscle was found to be statistically higher (p < 0.05) in the IPRA (1.71 ± 0.47) as compared to the TPC (1.38 ± 0.21). However, no statistical differences in moisture, mineral ash, and crude protein between the two culture systems during the whole experimental period were documented. The nutritional composition of grass carp flesh is depicted in Table 4.
3.4. Muscle Texture Profiles
The comprehensive texture characteristics of grass crap muscle are presented in Table 5. The flesh adhesiveness was evidently statistically higher (p < 0.05) in TPC than in IPRA. There were no significant differences in physicochemical characteristics (pH and WHC) in grass carp muscle between the two culture systems. The fish muscles from the TPC were observed to have a higher pH level, while the WHC was recorded as being higher in IPRA after 6−10 h of postmortem, but the differences in both attributes were non-significant.
3.5. 2-MIB and Geosmin in Muscle
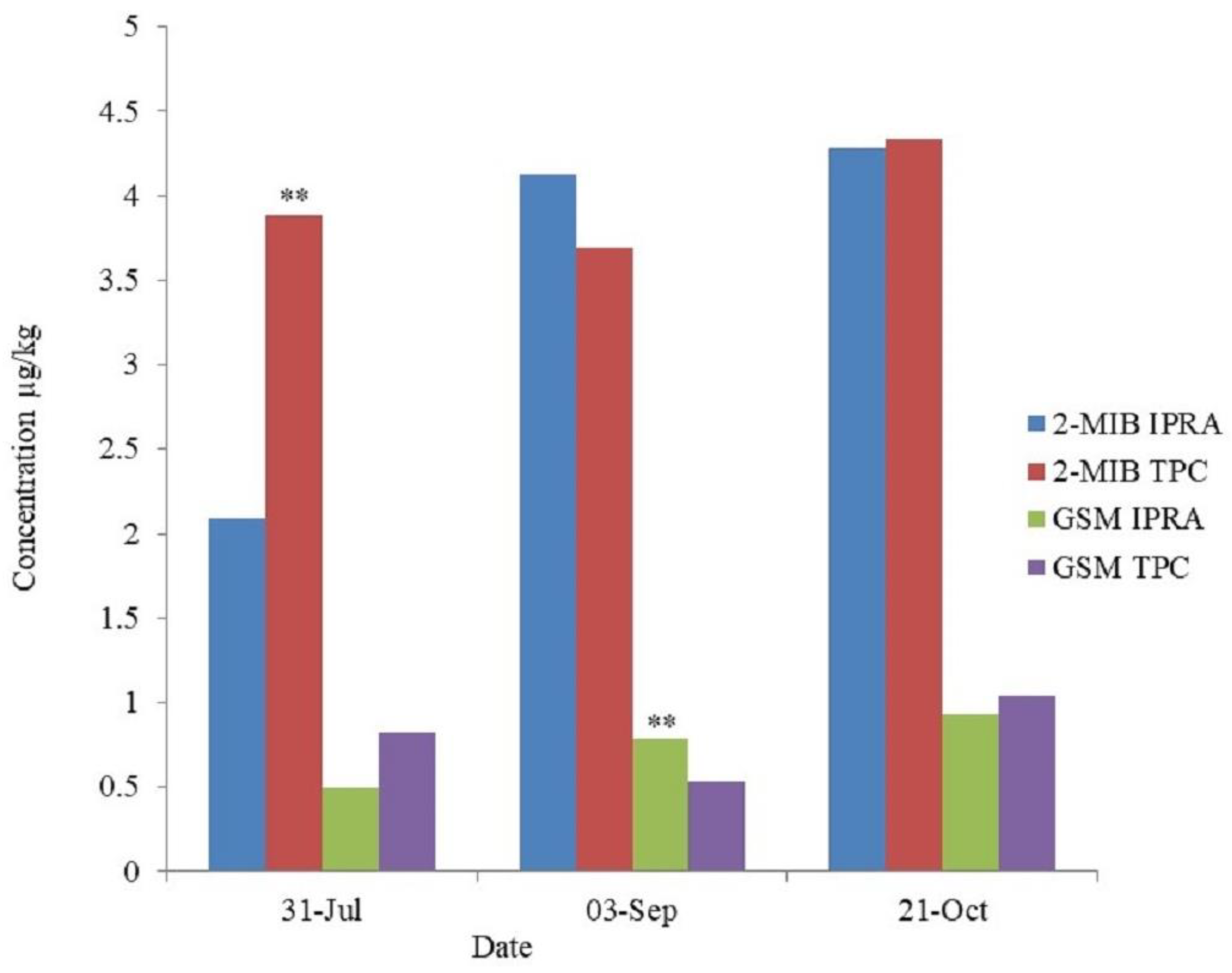
Off-flavor compounds, 2-MIB, and geosmin levels in grass carp muscle from the IPRA and TPC are shown in Figure 2. The muscles were observed to contain significantly higher (p < 0.01) 2-MIB in TPC (3.88 ± 0.24 µg/kg) than that in IPRA (2.09 ± 0.16 µg/kg) at first sampling (0 day/July) but were statistically similar during second (33 days/September) and third sampling (83 days/October). Similarly, the geosmin content of flesh was found to be significantly higher (p < 0.01) in IPRA (0.79 ± 0.05 µg/kg) than in TPC (0.53 ± 0.01 µg/kg) at the second sampling (33 days) but was not significantly different during the first (0 day) and third sampling (83 days). The2-MIB content in the grass carp muscle gradually increased during the whole culture period in IPRA, but in TPC, the geosmin content in the muscle got more depressed in September than in July and October.
3.6. Microbial Diversity in Pond Water
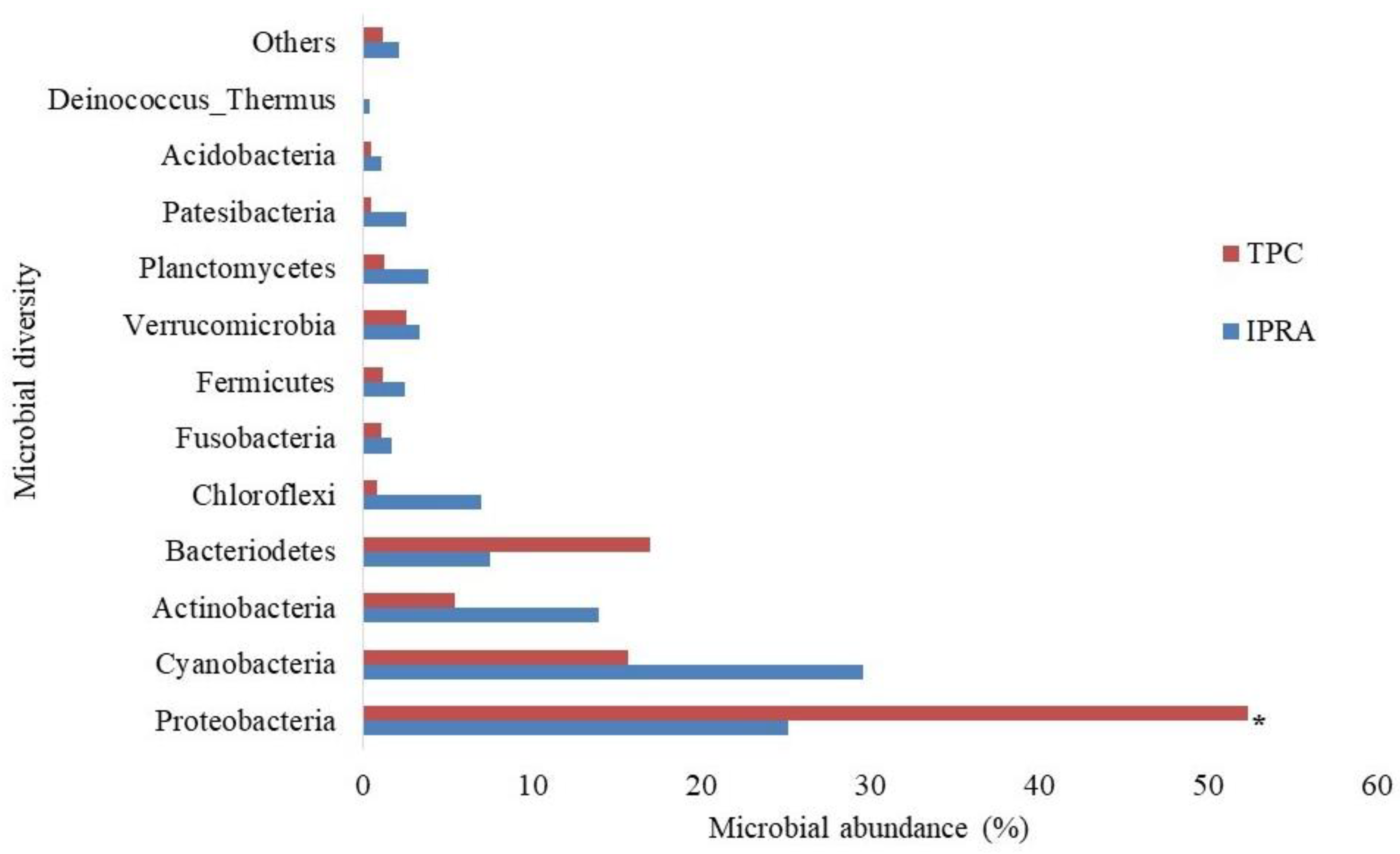
The study documented 12 different phyla of microbial mass in IPRA and TPC water environments (Figure 3). Cyanobacteria (29.59%) was the most abundant microbial mass, followed by Proteobacteria, Actinobacteria, Bacteriodetes, and Chloroflexi in IPRA, while Proteobacteria showed the dominating abundance in TPC, followed by Cyanobacteria, Bacteriodetes, and Actinobacteria. The abundance of Proteobacteria was observed to be significantly higher (p < 0.05) in TPC, whereas the richness of Planctomycetes was observed to be significantly higher (p < 0.05) in IPRA than in TPC.
4. Discussion
The fish in the IPRA culture unit might have received the appropriate culture environment, feeding management, and exercise for growth; therefore, weight gain rate (WGR), hepatosomatic index (HSI), and visceral mass ratio (VMR) were all observed to be higher in IPRA, despite being non-significant in this study. This fact indicated that the overall pond environments were optimal for growth, health, and energy reserves and that they had the capacity to withstand ecological challenges. Stocking density, pond environment, feed quality, swimming frequency, and physiological processes are the factors that influence the growth and muscle quality of fish [36,37,38,39,40,41].Wang et al. [42] reported higher total production with a relatively smaller harvest size in the IPRA at a higher stocking density. Similarly, Liu et al. [43] documented comparable findings between lower body weight and SGR at high density than in medium- and low-density treatments and reported that the maintenance of optimum water quality is essential for growth and for lowering the amount of metabolic waste. In the same vein, Liu et al. [38] observed higher LSI and CF with the addition of phytase to grass carp feed. The water quality parameters progressively worsened with age in both of the studied cultures, primarily because of the higher feed loads and the added fecal materials and feed residues. Intensive aquaculture with high stocking densities requires a large amount of feed [44,45] and is accountable for worsening water quality through the metabolic excretion of fish, causing excessive ammonia and organic load.
The crude fat concentration in the muscle was observed to be significantly higher in IPRA than that in TPC, but all other parameters were statistically similar. Higher muscle fat content in IPRA might be associated with the stocking density, diurnal activity, and feeding behavior of fish. Zhao et al. and Zhang et al. [7,19] found that water quality and diet affect the nutritional characteristics of grass carp muscle; fish turn sluggish, and a lesser amount of lipids are burned as energy, resulting in an accumulation in the muscles [46]. The differences in fish fat concentration were reported to be diverse according to different rearing systems [7]. Pyzlukasik et al. [47] reported that rainbow trout from intensive farming contained higher amounts of fat (5.39% compared to 3.13%) and less protein (19.23% compared to 20.34%) than trout from extensive culture. This finding is in agreement with Yang et al. [46] and Younis et al. [48], who reported a higher lipid content and low protein content in tilapia from ponds and in-pond cultured grass carp [49]. However, ref. [50] the documented increase in lipid metabolism and carbohydrate oxidation in grass carp reared at 40 kg/m3. In general, muscle fat content depends on the rearing conditions, reared fish species, and feed provided to the aquatic animals [18,51]. However, lower fat contents have been documented in wild fish species [52] and may be associated with higher alertness and activity. Interestingly, Valente et al. [53] reported an increase in lipid content in accordance with the degree of intensification in culture conditions in sea bream (Sparus aurata) and in other fish [54].
Muscle pH and WHC, a set of attributes defining muscle characteristics, are important aspects to evaluate muscle quality [55] and are associated with good mouthfeel [56,57]. Glycolytic potential, some biological reactions, and fatty acid concentrations and composition affect muscle pH [58] and strongly control the WHC. High muscle WHC may be due to compromised protein breakdown [59]. In this study, the pH and WHC of fish muscles were found to be statistically similar in both culture systems. This observation is similar to the findings of El Rammouz et al. [60], who reported anon-significant difference in pH and WHC after 2 to 6 h of postmortem in rainbow trout (O. mykiss) and stated that a higher WHC was observed at muscle pH 6.33.
Texture is an important characteristic for all members of the value chain, from producers to customers, in terms of satisfaction and acceptance of fish products [61]. The texture of fish muscle, especially hardness and juiciness, is affected mainly by the amount of moisture, fat, and protein, especially the collagen content [62]. Andersoen et al. [63] documented the correlation of fish muscle with a softer texture and fat content, while increased WHC might be linked with muscle hardening [62,64] and lower muscle pH with higher softness. Therefore, the higher hardness of muscle in TPC grass carp might be related to their lower fat content and higher pH. Fuentes et al. [61] demonstrated that muscle from wild stock had a higher hardness and springiness as their habitat was better than a cultured aquatic environment. The better adhesiveness of fish from the TPC was probably related to their higher muscle pH [59]. In this study, muscle texture profile analysis showed that all the textural attributes are statistically similar except for muscle adhesiveness, which was observed to be higher in IPRA fish muscle. These findings might be due to the better water quality and higher freedom of movement in TPC, which probably affected fish growth, muscle development, and the characteristic texture. This is because the low stocking density puts less stress on the fish in clean water. This result is in line with the findings of Zhao et al. [7], who reported a higher fillet lipid content with poor texture parameters such as hardness, resilience, and shear force in grass carp on artificial food. Furthermore, Refaey et al. [65] discussed the relationship between water quality and muscle texture and stated that low water quality spikes chronic stress in fish, disturbs homeostasis, and negatively affects the size and number of myofibers, influencing the muscle texture [66].
Concentration of geosmin and 2-MIB in fish muscle results in “earthy” and “musty” off-flavors [67], and Actinomycetes and a diverse array of blue-green algae are the major culprits [68] for this characteristic’s development. In this study, grass carp muscles were observed to contain an insignificantly higher amount of 2-MIB and geosmin content in TPC than those in IPRA at harvest, which might be due to a significantly higher Proteobacteria dominance. The 2-MIB content in the grass carp muscle showed an increasing trend for the whole culture period in both IPRA and TPC; however, muscle geosmin content was observed to be lower in September than in July and October in TPC, which correlated with a higher abundance of Cyanobacteria, Proteobacteria, Actinobacteria, Bacteriodetes, Chloroflexi, Planctomycetes, etc. Actinomycetes, Cyanobacteria, Proteobacteria, fungi, etc., were reported to produce geosmin and 2-MIB as secondary metabolites [25,68].In line with the results of this study, Qin et al. [24] and Tang et al. [13] reported comparable pictures of microbial diversity in pond water and described Proteobacteria, Bacteriodetes, Actinobacteria, and Planctomycetes as the dominant microorganisms in all types of culture systems and Proteobacteria as a leading geosmin producer [25].
The initial lower load of 2-MIB and geosmin with a consistent increasing trend for the complete culture period in this study (except at second sampling for 2-MIB and geosminin TPC) signals the obvious initial minimum number of Actinomycetes and blue-green algae in water, along with the subsequent multiplication in an aging culture period, and calls upon a profound sensory script at the harvest. The concentrations of geosmin and 2-MIB in grass carp flesh in both IPRA and TPC were above the concentration of sensory threshold, 0.9 µg/kg for geosmin [69] and 0.6 µg/kg for MIB [70], referenced with the rainbow trout standard, except for geosmin level at the initial two samplings in July and September. These findings are in line with Vallod et al. [71], who documented geosmin (0.48–3.05 µg/kg and 0.49–3.51 µg/kg) and 2-MIB (3.16–9.82 µg/kg and 2.11–7.25 µg/kg) in hybrid catfish and Nile tilapia, respectively, recording both odorous terpenoids above THC. Schrader et al. [28] also documented the highest geosmin and 2-MIB levels (1712 and 29,851 ng/kg) in hybrid catfish fillets. Similarly, Gutierrez et al. [72] also reported the high intensity of off-flavor in carps in ponds with a higher abundance of Cyanobacteria in the summer and beginning of autumn. The nutrient-enriched ponds at high temperatures may have supported the growth of Cyanobacteria, yielding off-flavors in the water and in fish muscle [71,73]. However, Varga et al. [74] described much lower levels of 2-MIB (0.19 ± 0.08 ng/g) and geosmin (0.13 ± 0.03 ng/g) in common carp.
5. Conclusions
The growth performances and muscle quality of grass carp cultured in TPC and IPRA were assessed and compared in this study. Along with aging culture duration, water quality parameters showed an increasing trend. With the exception of a few samples in TPC, the various forms of nitrogen, including nitrite nitrogen, ammonia, and total nitrogen, phosphate, and total phosphorus levels, were found to have a rising trend in IPRA and were discovered to have a very significant difference. Phosphate and total phosphorous were also significantly higher in IPRA. The crude lipid was observed to be significantly higher in IPRA fish muscle when compared to TPC during the experimental period. In terms of the physicochemical characteristics (pH and WHC), there were no appreciable variations in the muscle of grass carp between the two culture systems. The difference was evident upon inspecting flesh adhesiveness. TPC and IPRA clearly differed from one another, with TPC having superior flesh adhesiveness.
At the first sampling in July, it was found that TPC had higher levels of 2-MIB than IPRA did, but these levels were similar at the second sampling in September and at harvest in October. However, at the second sampling, it was discovered that the IPRA had higher levels of geosmin than the TPC did. While the 2-MIB content in fish muscle steadily increased throughout the IPRA culture period, the geosmin content in muscle in TPC declined more in September than it did in July and October. In IPRA and TPC water habitats, the study found 12 different phyla of microbial mass. In IPRA, Cyanobacteria (29.59%) had the highest dominance, followed by Proteobacteria, Actinobacteria, Bacteriodetes, and Chloroflexi, while in TPC, Proteobacteria showed the highest concentration, followed by Cyanobacteria, Bacteriodetes, and Actinobacteria. The abundance of Proteobacteria was observed to be higher in TPC, while the richness of Planctomycetes was higher in IPRA than TPC.
Despite a high stocking density in IPRA, the impact on nutrient composition was low, meaning that a low-fat content in grass carp is produced, and the textural quality of muscle is non-significant, aside from its adhesiveness. These things considered, the off-flavor in IPRA-produced fish had less of an impact on the aesthetic quality of flesh than TPC did with the aging of the culture duration. Taking into account all of these details, the study’s findings demonstrate that IPRA’s grass carp culture produces better-quality muscle than TPCs.
Author Contributions
Conceptualization, K.G. and L.L.; methodology, K.G., L.Y., K.L. and L.L.; software, K.G., L.Y., K.L. and N.B.; validation, K.G. and L.L.; formal analysis, K.G. and K.L.; investigation, K.G.; resources, L.L.; data curation, K.G., K.L., N.B. and L.L.; writing—original draft preparation, K.G.; writing—review and editing, K.G. and K.L., N.B. and L.L.; visualization, K.G., L.Y.; supervision, L.L.; project administration, L.L. and L.Y.; funding acquisition, L.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Shanghai Agriculture Applied Technology Development Programme grant number [X20210301] and The National Key Research and Development Programme of China grant number [2019YFD0900303].
Institutional Review Board Statement
All animal care and experimental procedures were approved by the Institutional Animal Care and Use Committee of Shanghai Ocean University [SHOU-DW-2019-058], Shanghai, China.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Acknowledgments
We thank Justice Frimpong Amankwah for editing and fine-tuning the manuscript language. The authors also thank Shanghai Agriculture Applied Technology Development Programme and The National Key Research and Development Programme of China for their financial assistance to this work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Subasinghe, R.; Soto, D.; Jia, J. Global aquaculture and its role in sustainable development. Rev. Aquac. 2009, 1, 2–9. [Google Scholar] [CrossRef]
- Crab, R.; Defoirdt, T.; Bossier, P.; Verstraete, W. Biofloc technology in aquaculture: Beneficial effects and future challenges. Aquaculture 2012, 356–357, 351–356. [Google Scholar] [CrossRef]
- Henriksson, P.J.G.; Troell, M.; Banks, L.K.; Belton, B.; Beveridge, M.C.M.; Klinger, D.H.; Pelletier, N.; Phillips, M.J.; Tran, N. Interventions for improving the productivity and environmental performance of global aquaculture for future food security. One Earth 2021, 4, 1220–1232. [Google Scholar] [CrossRef]
- Xie, C.; Li, J.; Li, D.; Shen, Y.; Gao, Y.; Zhang, Z. Grass carp: The fish that feeds half of china. In Aquaculture in China; Gui, J.-F., Tang, Q., Li, Z., Liu, J., De Silva, S.S., Eds.; John Wiley & Sons Ltd.: Chichester, UK, 2018; pp. 93–115. [Google Scholar]
- FAO. The State of World Fisheries and Aquaculture 2022. Towards Blue Transformation; FAO: Rome, Italy, 2022. [Google Scholar]
- Gisbert, E.; Mozanzadeh, M.T.; Kotzamanis, Y.; Estévez, A. Weaning wild flathead grey mullet (Mugil cephalus) fry with diets with different levels of fish meal substitution. Aquaculture 2016, 462, 92–100. [Google Scholar] [CrossRef]
- Zhao, H.; Xia, J.; Zhang, X.; He, X.; Li, L.; Tang, R.; Chi, W.; Li, D. Diet Affects Muscle Quality and Growth Traits of Grass Carp (Ctenopharyngodon idellus): A Comparison between Grass and Artificial Feed. Front. Physiol. 2018, 9, 283. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, P.; Nie, Z.; Li, Q.; Shao, N.; Xu, G. Growth, digestive enzymes activities, serum biochemical parameters and antioxidant status of juvenile genetically improved farmed tilapia (Oreochromis niloticus) reared at different stocking densities in in-pond raceway recirculating culture system. Aquacult. Res. 2019, 50, 1338–1347. [Google Scholar] [CrossRef]
- Fatima, S.; Komal, W.; Manzoor, F.; Latif, A.A.; Liaqat, R.; Ameen, S.; Janjua, R.S. Analysis of the growth performance, stress, profile of fatty acids and amino acids and cortisol in Tilapia (Oreochromis niloticus), cultured at high stocking density using in-pond raceway system. Saudi J. Biol. Sci. 2021, 28, 7422–7431. [Google Scholar] [CrossRef]
- Martin, T.E. Are microhabitat preferences of coexisting species under selection and adaptive? Ecology 1998, 79, 656–670. [Google Scholar] [CrossRef]
- Nielsen, J.; Hyldig, G.; Larsen, E. ‘Eating Quality’ of Fish—A Review. J. Aquat. Food Prod. Technol. 2002, 11, 125–141. [Google Scholar] [CrossRef]
- Listrat, A.; Lebret, B.; Louveau, I.; Astruc, T.; Bonnet, M.; Lefaucheur, L.; Picard, B.; Bugeon, J. How Muscle Structure and Composition Influence Meat and Flesh Quality. Sci. World J. 2016, 2016, 3182746. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Zhao, L.; Cheng, Y.; Yang, Y.; Sun, Y.; Liu, Q. Control of cyanobacterial blooms in different polyculture patterns of filter feeders and effects of these patterns on water quality and microbial community in aquacultural ponds. Aquaculture 2021, 542, 736913. [Google Scholar] [CrossRef]
- Alamri, S.A.; Mohamed, Z.A. Selective inhibition of toxic cyanobacteria by β-carboline-containing bacterium Bacillus flexus isolated from Saudi freshwaters. Saudi J. Biol. Sci. 2013, 20, 357–363. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Tong, F.; Tang, T.; Ao, Z.; Wei, Z.; Yang, F.; Shu, Y.; Liu, S.; Mai, K. Comparative evaluation of nutritional value and flavor quality of muscle in triploid and diploid common carp: Application of genetic improvement in fish quality. Aquaculture 2021, 541, 736780. [Google Scholar] [CrossRef]
- Wen, J.; Jiang, W.; Feng, L.; Kuang, S.; Jiang, J.; Tang, L.; Zhou, X.; Liu, Y. The influence of graded levels of available phosphorus on growth performance, muscle antioxidant and flesh quality of young grass carp (Ctenopharyngodon idella). Anim. Nutr. 2015, 1, 77–84. [Google Scholar] [CrossRef]
- Granada, L.; Lopes, S.; Novais, S.C.; Lemos, M.F.L. Modelling integrated multi-trophic aquaculture: Optimizing a three trophic level system. Aquaculture 2018, 495, 90–97. [Google Scholar] [CrossRef]
- Zhang, X.; Wang, J.; Tang, R.; He, X.; Li, L.; Takagi, Y.; Li, D. Improvement of Muscle Quality of Grass Carp (Ctenopharyngodon idellus) With a Bio-Floating Bed in Culture Ponds. Front. Physiol. 2019, 10, 683. [Google Scholar] [CrossRef]
- Zhang, X.; Shen, Z.; Qi, T.; Xi, R.; Liang, X.; Li, L.; Tang, R.; Li, D. Slight Increases in Salinity Improve Muscle Quality of Grass Carp (Ctenopharyngodon idellus). Fishes 2021, 6, 7. [Google Scholar] [CrossRef]
- Edwards, P. Aquaculture environment interactions: Past, present and likely future trends. Aquaculture 2015, 447, 2–14. [Google Scholar] [CrossRef]
- Dauda, A.B.; Ajadi, A.; Tola-Fabunmi, A.S.; Akinwole, A.O. Waste production in aquaculture: Sources, components and managements in different culture systems. Aquac. Fish. 2019, 4, 81–88. [Google Scholar] [CrossRef]
- Gutierrez, R.; Itayama, T.; Iwami, N.; Whangchai, N. Analysis of Geosmin and 2-Methylisoborneol Off-flavors In Tilapia Cage-Cultures in Thailand. In Proceedings of the 2nd MJU-Phrae National Research Conference, Phrae, Thailand, 1–2 September 2011. [Google Scholar]
- Hathurusingha, P.I.; Davey, K.R. A predictive model for taste taint accumulation in Recirculating Aquaculture Systems (RAS) farmed-fish–demonstrated with geosmin (GSM) and 2-methylisoborneol (MIB). Ecol. Model. 2014, 291, 242–249. [Google Scholar] [CrossRef]
- Qin, Y.; Hou, J.; Deng, M.; Liu, Q.; Wu, C.; Ji, Y.; He, X. Bacterial abundance and diversity in pond water supplied with different feeds. Sci. Rep. 2016, 6, 35232. [Google Scholar] [CrossRef] [PubMed]
- Lukassen, M.B.; de Jonge, N.; Bjerregaard, S.M.; Podduturi, R.; Jorgensen, N.O.G.; Petersen, M.A.; David, G.S.; da Silva, R.J.; Nielsen, J.L. Microbial Production of the Off-Flavor Geosmin in Tilapia Production in Brazilian Water Reservoirs: Importance of Bacteria in the Intestine and Other Fish-Associated Environments. Front. Microbiol. 2019, 10, 2447. [Google Scholar] [CrossRef] [PubMed]
- Brown, T.W.; Tucker, C.S. Pumping performance of a slow-rotating paddlewheel for split-pond aquaculture systems. N. Am. J. Aquacult. 2013, 75, 153–158. [Google Scholar] [CrossRef]
- Chopin, T.; Buschmann, A.H.; Halling, C.; Troell, M.; Kautsky, N.; Neori, A.; Kraemer, G.P.; Zertuche-González, J.A.; Yarish, C.; Neefus, C. Integrating seaweeds into marine aquaculture systems: A key toward sustainability. J. Phycol. 2001, 37, 975–986. [Google Scholar] [CrossRef]
- Schrader, K.K.; Tucker, C.S.; Brown, T.W.; Whitis, G.N. Earthy and Musty Off-Flavor Episodes in Catfish Split-Pond Aquaculture Systems. N. Am. J. Aquacult. 2018, 80, 26–41. [Google Scholar] [CrossRef]
- Wang, H.; Zhu, Y.; Zhang, J.; Wang, X.; Shi, W. Study on changes in the quality of grass carp in the process of postmortem. J. Food Biochem. 2018, 42, e12683. [Google Scholar] [CrossRef]
- Brown, T.W.; Chappell, J.A.; Boyd, C.E. A commercial-scale, in-pond raceway system for Ictalurid catfish production. Aquacult. Eng. 2011, 44, 72–79. [Google Scholar] [CrossRef]
- Zhang, X.; Zheng, W.; Zhang, H.; Chai, Y.; Ruan, G. Comparison of Muscle Quality of the Yellow Catfish Cultured in In-Pond Raceway Systems and Traditional Ponds. Water 2022, 14, 1223. [Google Scholar] [CrossRef]
- Akinwole, A.; Dauda, A.; Ololade, A. Haematological response of Clarias gariepinus juveniles reared in treated wastewater after waste solids removal using alum or Moringa oleifera seed powder. Int. J. Aquacult. 2016, 6, 1–8. [Google Scholar]
- Zhang, J.; Kaneko, G.; Sun, J.; Wang, G.; Xie, J.; Tian, J.; Li, Z.; Gong, W.; Zhang, K.; Xia, Y.; et al. Key Factors Affecting the Flesh Flavor Quality and the Nutritional Value of Grass Carp in Four Culture Modes. Foods 2021, 10, 2075. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, Y.; Shao, J.; Wang, J.; Li, R. Genes associated with 2-methylisoborneol biosynthesis in cyanobacteria: Isolation, characterization, and expression in response to light. PLoS ONE 2011, 6, e18665. [Google Scholar] [CrossRef] [PubMed]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An improved dual-indexing approach for multiplexed 16S rRNA gene sequencing on the Illumina MiSeq platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef] [PubMed]
- Johnston, I.A. Genetic and environmental determinants of muscle growth patterns. In Fish Physiology; Elsevier: Amsterdam, The Netherlands, 2001; Volume 18, pp. 141–186. [Google Scholar]
- Hua, K.; Koppe, W.; Fontanillas, R. Effects of dietary protein and lipid levels on growth, body composition and nutrient utilization of Channa striata. Aquaculture 2019, 501, 368–373. [Google Scholar] [CrossRef]
- Liu, L.; Zhou, Y.; Wu, J.; Zhang, W.; Abbas, K.; Xu-Fang, L.; Luo, Y. Supplemental graded levels of neutral phytase using pretreatment and spraying methods in the diet of grass carp, Ctenopharyngodon idellus. Aquacult. Res. 2014, 45, 1932–1941. [Google Scholar] [CrossRef]
- Masser, M.P. In-pond raceways. In Aquaculture Production Systems; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 387–394. [Google Scholar]
- Yuan, J.; Ni, M.; Liu, M.; Wang, H.; Zhang, C.; Mi, G.; Gu, Z. Analysis of the growth performances, muscle quality, blood biochemistry and antioxidant status of Micropterus salmoides farmed in in-pond raceway systems versus usual-pond systems. Aquaculture 2019, 511, 734241. [Google Scholar] [CrossRef]
- Brown, T.W.; Boyd, C.E.; Chappell, J.A. Approximate Water and Chemical Budgets for an Experimental, In-pond Raceway System. J. World Aquacult. Soc. 2012, 43, 526–537. [Google Scholar] [CrossRef]
- Wang, Y.; Xu, G.; Nie, Z.; Shao, N.; Li, Q.; Xu, P. Growth Performance of Bluntnose Black Bream, Channel Catfish, Yellow Catfish, and Largemouth Bass Reared in the In-Pond Raceway Recirculating Culture System. N. Am. J. Aquacult. 2019, 81, 153–159. [Google Scholar] [CrossRef]
- Liu, Q.; Hou, Z.; Wen, H.; Li, J.; He, F.; Wang, J.; Guan, B.; Wang, Q. Effect of stocking density on water quality and (Growth, Body Composition and Plasma Cortisol Content) performance of pen-reared rainbow trout (Oncorhynchus mykiss). J. Ocean. Univ. China 2016, 15, 667–675. [Google Scholar] [CrossRef]
- Li, W.; Cheng, X.; Xie, J.; Wang, Z.; Yu, D. Hydrodynamics of an in-pond raceway system with an aeration plug-flow device for application in aquaculture: An experimental study. R. Soc. Open Sci. 2019, 6, 182061. [Google Scholar] [CrossRef]
- Ni, M.; Liu, M.; Lou, J.; Mi, G.; Yuan, J.; Gu, Z. Stocking density alters growth performance, serum biochemistry, digestive enzymes, immune response, and muscle quality of largemouth bass (Micropterus salmoides) in in-pond raceway system. Fish Physiol. Biochem. 2021, 47, 1243–1255. [Google Scholar] [CrossRef]
- Yang, H.; He, J.; Lv, M.; Huang, G.; Wen, L.; Bi, X.; Hu, T.; Ma, H. Preliminary effects of dietary protein levels on muscle quality and digestive enzyme activities in GIFT-Oreochromis niloticus. Aquac. Stud. 2018, 18, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pyz-Łukasik, R.; Chałabis-Mazurek, A.; Gondek, M. Basic and functional nutrients in the muscles of fish: A review. Int. J. Food Prop. 2020, 23, 1941–1950. [Google Scholar] [CrossRef]
- Younis, E.M.; Al-Quffail, A.S.; Al-Asgah, N.A.; Abdel-Warith, A.A.; Al-Hafedh, Y.S. Effect of dietary fish meal replacement by red algae, Gracilaria arcuata, on growth performance and body composition of Nile tilapia Oreochromis niloticus. Saudi J. Biol. Sci. 2018, 25, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Pyz-Łukasik, R.; Paszkiewicz, W. Species Variations in the Proximate Composition, Amino Acid Profile, and Protein Quality of the Muscle Tissue of Grass Carp, Bighead Carp, Siberian Sturgeon, and Wels Catfish. J. Food Qual. 2018, 2018, 2625401. [Google Scholar] [CrossRef]
- He, Y.; Yu, H.; Zhao, H.; Zhu, H.; Zhang, Q.; Wang, A.; Shen, Y.; Xu, X.; Li, J. Transcriptomic analysis to elucidate the effects of high stocking density on grass carp (Ctenopharyngodon idella). BMC Genom. 2021, 22, 620. [Google Scholar] [CrossRef]
- Orban, E.; Lena, G.D.; Nevigato, T.; Casini, I.; Santaroni, G.; Marzetti, A.; Caproni, R. Quality characteristics of sea bass intensively reared and from lagoon as affected by growth conditions and the aquatic environment. J. Food Sci. 2002, 67, 542–546. [Google Scholar] [CrossRef]
- Grigorakis, K.; Taylor, K.; Alexis, M. Organoleptic and volatile aroma compounds comparison of wild and cultured gilthead sea bream (Sparus aurata): Sensory differences and possible chemical basis. Aquaculture 2003, 225, 109–119. [Google Scholar] [CrossRef]
- Valente, L.M.P.; Cornet, J.; Donnay-Moreno, C.; Gouygou, J.P.; Bergé, J.P.; Bacelar, M.; Escórcio, C.; Rocha, E.; Malhão, F.; Cardinal, M. Quality differences of gilthead sea bream from distinct production systems in Southern Europe: Intensive, integrated, semi-intensive or extensive systems. Food Control 2011, 22, 708–717. [Google Scholar] [CrossRef]
- Flos, R.; Reig, L.; Oca, J.; Ginovart, M. Influence of marketing and different land-based systems on gilthead sea bream (Sparus aurata) quality. Aquacult. Int. 2002, 10, 189–206. [Google Scholar] [CrossRef]
- Rasmussen, R.S. Quality of farmed salmonids with emphasis on proximate composition, yield and sensory characteristics. Aquacult. Res. 2001, 32, 767–786. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, Y.; Zhang, Y.; Ning, Z.; Li, Y.; Zhao, Q.; Lu, H.; Huang, R.; Xia, X.; Feng, Q.; et al. The draft genome of the grass carp (Ctenopharyngodon idellus) provides insights into its evolution and vegetarian adaptation. Nat. Genet. 2015, 47, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Feng, L.; Jiang, W.-D.; Wu, P.; Liu, Y.; Jiang, J.; Kuang, S.-Y.; Tang, L.; Zhou, X.-Q. Different dietary protein levels affect flesh quality, fatty acids and alter gene expression of Nrf2-mediated antioxidant enzymes in the muscle of grass carp (Ctenopharyngodon idella). Aquaculture 2018, 493, 272–282. [Google Scholar] [CrossRef]
- Fernandez, X.; MÅGÅRd, M.; Tornberg, E.V.A. Glycolytic Potential in Porcine Longissimus Muscle before and after Transport: An in Vivo Study. J. Muscle Foods 1992, 3, 83–89. [Google Scholar] [CrossRef]
- Bee, G.; Anderson, A.L.; Lonergan, S.M.; Huff-Lonergan, E. Rate and extent of pH decline affect proteolysis of cytoskeletal proteins and water-holding capacity in pork. Meat Sci. 2007, 76, 359–365. [Google Scholar] [CrossRef]
- El Rammouz, R.; Abboud, J.; Abboud, M.; El Mur, A.; Yammine, S.; Jammal, B. pH, rigor mortis and physical properties of fillet in fresh water fish: The case of rainbow trout (Oncorynchus mykiss). J. Appl.Sci. Res. 2013, 9, 5746–5755. [Google Scholar]
- Fuentes, A.; Fernández-Segovia, I.; Serra, J.A.; Barat, J.M. Comparison of wild and cultured sea bass (Dicentrarchus labrax) quality. Food Chem. 2010, 119, 1514–1518. [Google Scholar] [CrossRef]
- Cheng, J.H.; Sun, D.W.; Han, Z.; Zeng, X.A. Texture and Structure Measurements and Analyses for Evaluation of Fish and Fillet Freshness Quality: A Review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 52–61. [Google Scholar] [CrossRef]
- Andersen, U.B.; Thomassen, M.S.; Rørå, A.M.B. Texture properties of farmed rainbow trout (Oncorhynchus mykiss): Effects of diet, muscle fat content and time of storage on ice. J. Sci. Food Agric. 1997, 74, 347–353. [Google Scholar] [CrossRef]
- Pearce, K.L.; Rosenvold, K.; Andersen, H.J.; Hopkins, D.L. Water distribution and mobility in meat during the conversion of muscle to meat and ageing and the impacts on fresh meat quality attributes—A review. Meat Sci. 2011, 89, 111–124. [Google Scholar] [CrossRef]
- Refaey, M.M.; Li, D.; Tian, X.; Zhang, Z.; Zhang, X.; Li, L.; Tang, R. High stocking density alters growth performance, blood biochemistry, intestinal histology, and muscle quality of channel catfish Ictalurus punctatus. Aquaculture 2018, 492, 73–81. [Google Scholar] [CrossRef]
- Taylor, R.; Fjaera, S.; Skjervold, P. Salmon fillet texture is determined by myofiber-myofiber and myofiber-myocommata attachment. J. Food Sci. 2002, 67, 2067–2071. [Google Scholar] [CrossRef]
- Tucker, C.S. Off-flavor problems in aquaculture. Rev. Fish. Sci. 2000, 8, 45–88. [Google Scholar] [CrossRef]
- Guttman, L.; van Rijn, J. 2-Methylisoborneol and geosmin uptake by organic sludge derived from a recirculating aquaculture system. Water Res. 2009, 43, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Robertson, R.F.; Hammond, A.; Jauncey, K.; Beveridge, M.C.M.; Lawton, L.A. An investigation into the occurrence of geosmin responsible for earthy–musty taints in UK farmed rainbow trout, Onchorhynchus mykiss. Aquaculture 2006, 259, 153–163. [Google Scholar] [CrossRef]
- Persson, P.-E. Sensory properties and analysis of two muddy odour compounds, geosmin and 2-methylisoborneol, in water and fish. Water Res. 1980, 14, 1113–1118. [Google Scholar] [CrossRef]
- Gutierrez, R.; Whangchai, N.; Sompong, U.; Prarom, W.; Iwami, N.; Itayama, T.; Nomura, N.; Sugiura, N. Off-flavour in Nile tilapia (Oreochromis niloticus) cultured in an integrated pond-cage culture system. Maejo Int. J. Sci. Technol. 2013, 7, 1. [Google Scholar]
- Vallod, D.; Cravedi, J.; Hillenweck, A.; Robin, J. Analysis of the off-flavor risk in carp production in ponds in Dombes and Forez (France). Aquacult. Int. 2007, 15, 287–298. [Google Scholar] [CrossRef]
- Lukassen, M.B.; Saunders, A.M.; Sindilariu, P.-D.; Nielsen, J.L. Quantification of novel geosmin-producing bacteria in aquaculture systems. Aquaculture 2017, 479, 304–310. [Google Scholar] [CrossRef]
- Varga, D.; Sándor, Z.; Hancz, C.; Csengeri, I.; Jeney, Z.; Papp, Z. Off-flavour compounds in common carp (Cyprinus carpio L.) flesh in context of type of fish pond. Acta Aliment. 2015, 44, 311–315. [Google Scholar] [CrossRef]
Figure 1.
Water quality parameters of IPRA and TPC. Various forms of nitrogen ((a)—NO2−N, (b)—NO3−N, (c)—ammonia nitrogen) and (d)—PO43−P were tested over the experimental period. * Indicates significant (p < 0.05), ** indicates highly significant (p < 0.01).
Figure 1.
Water quality parameters of IPRA and TPC. Various forms of nitrogen ((a)—NO2−N, (b)—NO3−N, (c)—ammonia nitrogen) and (d)—PO43−P were tested over the experimental period. * Indicates significant (p < 0.05), ** indicates highly significant (p < 0.01).

Figure 2.
2-MIB and geosmin levels in flesh of grass carp from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC). Note: ** indicates highly significant (p < 0.01).
Figure 2.
2-MIB and geosmin levels in flesh of grass carp from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC). Note: ** indicates highly significant (p < 0.01).

Figure 3.
Pond water microbial diversity in in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC). Note: * indicates significant (p < 0.05).
Figure 3.
Pond water microbial diversity in in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC). Note: * indicates significant (p < 0.05).


{kind=link}
{kind=link}
{kind=link}
Table 1.
Water quality indicators of complete culture period in in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
Table 1.
Water quality indicators of complete culture period in in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
| Water Quality Indicators | IPRA | TPC | ||||
|---|---|---|---|---|---|---|
| Min | Max | Mean ± SD | Min | Max | Mean ± SD | |
| Temperature (°C) | 22.00 | 32.60 | 26.9 ± 4.29 | 22.80 | 34.00 | 27.4 ± 4.58 |
| Dissolve oxygen (mg/L) | 6.50 | 8.10 | 7.4 ± 0.63 | 6.90 | 8.00 | 7.7 ± 0.18 |
| pH | 7.20 | 7.60 | 7.4 ± 0.12 | 7.50 | 7.90 | 7.7 ± 0.18 |
| Total nitrogen (mg/L) | 3.05 | 9.51 | 5.69 ± 2.33 | 2.01 | 8.58 | 5.13 ± 2.65 |
| Total phosphorus (mg/L) | 2.11 | 7.71 | 4.32 ± 2.29 | 0.66 | 2.67 | 1.35 ± 1.00 |
| CODMn (mg/L) | 4.62 | 13.22 | 10.03 ± 4.79 | 6.39 | 10.96 | 7.74 ± 2.24 |
| Chlorophyll-a (mg/L) | 112.06 | 441.09 | 268 ± 155.41 | 80.96 | 351.49 | 173.91 ± 116.17 |
| Total soluble solid (mg/L) | 57.50 | 82.49 | 71.83 ± 50.0 | 64.67 | 108.90 | 85.85 ± 42.90 |
Table 2.
Geosmin and 2-MIB analysis in fish.
| Instruments | Used Conditions |
|---|---|
| Solid-phase microextraction (SPME) | Volume of vial 15 mL sample volume 5 mL |
| Gas Chromatography–Mass Spectrophotometry: 6890N Network (Agilent, Santa Clara, CA, USA) | Extraction time and temperature: 20 min and 50 °C |
| Detector: 5975 insert XL (Agilent, Santa Clara, CA, USA) | Desorption time: 5 min |
| Column: DB-WAX (30 m × 250 µm × 0.25 µm) | Injector temperature: 250 °C Oven temperature: 60 °C (2.5 min), 250 °C (8 °C/min) |
| Fiber SPME: Stable Flex 65µm DVB/ | Gas flow: He 1 mL/min Ionization voltage: 70 eV |
| PDMS/DVB (Supelco, Bellefonte, PA, USA) |
Notes: Detection limit of this method was 4.72 ng/L and 5.48 ng/L for GSM and 2-MIB, respectively [34].
Table 3.
Growth performance of grass carp from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
Table 3.
Growth performance of grass carp from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
| IPRA | TPC | |
|---|---|---|
| (Means ± SD) | (Means ± SD) | |
| Stocking weight (Kg.) | 0.42 ± 0.02 | 2.72 ± 0.14 |
| Weight (Kg.) | 0.86 ± 0.03 | 3.28 ± 0.02 * |
| Weight gain rate (%) | 117.93 ± 16.79 ** | 22.34 ± 4.85 |
| Specific growth rate (%/d) | 1.35 ± 0.20 | 1.67 ± 0.87 |
| Length (cm) | 34.25 ± 0.50 | 53.05 ± 3.80 |
| Visceral mass ratio (VMR) | 11.82 ± 1.03 | 11.5 ± 1.06 |
| Hepato somatic index (HSI) | 2.89 ± 0.18 | 2.50 ± 0.91 |
| Condition factor (CF g/cm3) | 2.14 ± 0.075 | 2.21 ± 0.52 |
Note: * Indicates significant (p < 0.05), ** indicates highly significant (p < 0.01).
Table 4.
Nutrient Composition of grass carp in IPRA and TPC.
| IPRA | TPC | |
|---|---|---|
| (n = 24) | (n = 27) | |
| Moisture% | 79.64 ± 0.81 | 78.49 ± 2.33 |
| Ash% | 1.469 ± 0.24 | 1.326 ± 0.20 |
| Crude lipid (CL%) | 1.707 ± 0.47 * | 1.383 ± 0.21 |
| Crude protein (CP%) | 18.66 ± 1.93 | 18.71 ± 0.94 |
Note: Values represent means ± SD, * indicates significant difference (p < 0.05).
Table 5.
pH, Water holding capacity, and texture profile analysis of grass carp muscle from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
Table 5.
pH, Water holding capacity, and texture profile analysis of grass carp muscle from in-pond raceway aquaculture (IPRA) and traditional pond culture (TPC).
| pH, WHC, and Texture Profile Attributes | IPRA | TPC |
|---|---|---|
| pH | 6.26 ± 0.07 | 6.34 ± 0.22 |
| Water Holding Capacity (%) | 0.25 ± 0.07 | 0.24 ± 0.068 |
| Hardness | 411.75 ± 183.27 | 582.52 ± 398.45 |
| Springiness | 0.51 ± 0.04 | 0.50 ± 0.07 |
| Stickiness | 4.02 ± 1.78 | 4.17 ± 2.14 |
| Chewiness | 122.67 ± 49.82 | 180.28 ± 142.29 |
| Adhesiveness | 237.25 ± 91.71 | 344.14 ± 243.82 * |
| Cohesiveness | 0.59 ± 0.05 | 0.59 ± 0.06 |
Note: Values represent means ± SD, * indicates significant difference (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gharti, K.; Yan, L.; Li, K.; Boonpeng, N.; Liu, L. Growth and Muscle Quality of Grass Carp (Ctenopharyngodon idella) in In-Pond Raceway Aquaculture and Traditional Pond Culture. Water 2023, 15, 1771. https://doi.org/10.3390/w15091771
AMA Style
Gharti K, Yan L, Li K, Boonpeng N, Liu L. Growth and Muscle Quality of Grass Carp (Ctenopharyngodon idella) in In-Pond Raceway Aquaculture and Traditional Pond Culture. Water. 2023; 15(9):1771. https://doi.org/10.3390/w15091771
Chicago/Turabian StyleGharti, Kamala, Li Yan, Kang Li, Nattida Boonpeng, and Liping Liu. 2023. "Growth and Muscle Quality of Grass Carp (Ctenopharyngodon idella) in In-Pond Raceway Aquaculture and Traditional Pond Culture" Water 15, no. 9: 1771. https://doi.org/10.3390/w15091771
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.